SEVERAL times in this book, dinosaurs are identified as the ancestors of birds. This should have a major effect on our view of the dinosaurs. Instead of thinking of them as huge lizard- or elephant-like animals, we should see dinosaurs as bird-like behaviorally (see
chapter 13). Dinosaurs are no longer considered just a long-extinct group of animals because their descendants, the birds, live today as one of the world’s most successful animal groups. Indeed, some paleontologists do not make a distinction and simply regard birds as dinosaurs. In this chapter, I develop the evidence for a dinosaur ancestry of birds and discuss some aspects of the evolution of the “feathered dinosaurs.”
There are about 10,000 species of living
birds, making them the most diverse group of vertebrates except for the bony fishes. All birds belong to a single class of vertebrates, the
Aves (from the Latin for “bird”). It is difficult to imagine a person who has not seen a bird and cannot readily distinguish it from other animals. This distinction, of course, is based primarily on the powered flight and feathers that are the hallmarks of birds. The
feathers of birds are intricate structures that not only form flight surfaces (flight feathers) but also insulate the bird (downy feathers) (
figure 15.1).
Like their flight feathers, skeletal features characteristic of birds are associated with flight. The limb bones are hollow and are very lightly constructed (thin walls). Braces and struts within bird long bones provide structural support for their thin walls (
figure 15.2), and the long bones are
pneumatic. This means that air passes through small openings (ducts) in the bones when the bird respires. These
pneumatic ducts are connected to
air sacs and the bird’s lungs (
figure 15.3). Avian air sacs not only lighten a bird, but they also supplement its lungs by acting both as a supercharger that increases the efficiency of respiration and as a cooling system for the fast avian metabolism. Because birds lack sweat glands, they are strictly air cooled by their lungs and air sacs.
FIGURE 15.1
Bird feathers, the most distinctive avian feature, are intricate structures. (Drawing by Network Graphics)
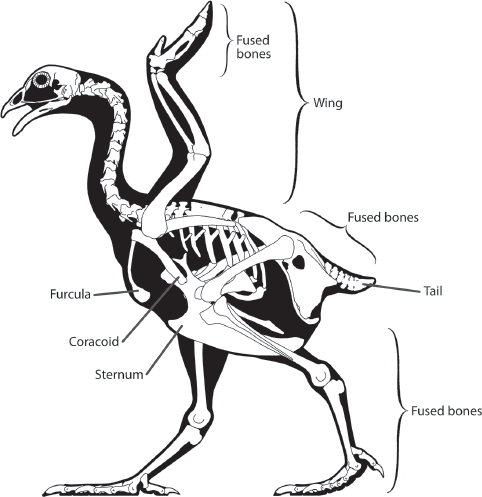
A second unique aspect of the avian skeleton is the fusion of many bones to form more rigid structures, which is particularly evident in the hind limb, pelvis, sacrum, and skull (
figure 15.4). For example, the collarbones (clavicles) are fused to form a single bone, the
furcula (the “wishbone”). Such fused structures contain far fewer separate bones than the same structures in most other vertebrates, including theropod dinosaurs.
Other distinctive features of all but the most primitive birds include modifications of the forelimb skeleton to form wings (
figure 15.5). The sternum (breastbone) is very large and keeled. The coracoid bone is very long and forms a brace with the sternum, and a large groove for the passage of a large tendon is present in the shoulder girdle. These structures are designed to anchor the huge and powerful chest muscles needed to flap the wings for flying. The limb itself is modified to form a wing. The carpals are fused so that the wrist is a simple hinge joint. The digits and metacarpals also are fused, and the joints at the wrist and elbow are modified to allow folding of the wing. Bumps on the ulna anchor flight feathers.
FIGURE 15.2
Avian long bones are hollow, have thin walls, and are braced by struts. (Drawing by Network Graphics)
FIGURE 15.3
The respiratory system of a bird is more than just its lungs. It includes air sacs connected to hollow bones. (Drawing by Network Graphics)
FIGURE 15.4
This skeleton of a chicken shows the fused bones in the limbs and hip that are characteristic of birds. (© Scott Hartman)
FIGURE 15.5
The shoulder girdle and forelimb skeleton of a bird is highly modified to form a wing. (© Scott Hartman)
Most paleontologists consider the Late Jurassic
Archaeopteryx to be the oldest known bird (
figure 15.6). It provides a unique glimpse of an animal in many ways part dinosaur, part bird.
Archaeopteryx thus represents what paleontologists would call a “transitional form” between two major groups of animals, the reptiles (dinosaurs) and birds. In popular terms,
Archaeopteryx is a “
missing link.”
Fossils of Archaeopteryx come from only one place, the Upper Jurassic Solnhofen Limestone in Bavaria, Germany. There, during the Late Jurassic, a lagoon was present behind coral reefs, at the bottom of which tiny fragments of calcium carbonate and the carbonate shells of microscopic marine organisms were deposited. The result was the preservation of delicate structures of the larger organisms that died there, making the Solnhofen Limestone an incredible graveyard of fossilized soft tissues. Indeed, the hallmark of the fossils of Archaeopteryx, their preserved feathers, is but one example of the high quality of fossilization in the Solnhofen Limestone.
The first known fossil of
Archaeopteryx is a single feather discovered in 1860. Since then, 11 skeletons have been discovered. The most exquisite, found in 1877, now resides in the Museum für Naturkunde in Berlin, Germany (
figure 15.7). The
Archaeopteryx specimens have been intensively studied for more than a century, and more has been written about them than about any other collection of a dozen fossils pertaining to one genus. Indeed, not all agree that the specimens belong to a single genus, so a second genus,
Wellnhoferia, has been named for some fossils previously called
Archaeopteryx.
The skeleton of Archaeopteryx shares many features with small theropod dinosaurs. Without feathers preserved, we might reasonably identify Archaeopteryx as a theropod. Indeed, some specimens of Archaeopteryx were originally identified as theropod (or pterosaur) before their feather impressions were recognized.
FIGURE 15.6
Late Jurassic Archaeopteryx was the first bird. (© Scott Hartman)
FIGURE 15.7
The Berlin specimen of Archaeopteryx is the most exquisitely preserved fossil of the first bird. (Courtesy John Ostrom, Yale Peabody Museum of Natural History)
Archaeopteryx was about the size of a crow or pigeon. It had a wingspan of about half a meter and an estimated weight of 300 to 500 grams. Its skull had teeth, and very few of its bones were fused to each other—dinosaurian, not avian features. The hollow limb bones of Archaeopteryx had thick walls without pneumatic openings, unlike the thin-walled, pneumatic limb bones of later birds.
No sternum (breastbone) is known for
Archaeopteryx, so it apparently lacked the main anchor for flight muscles seen in other birds. Although
Archaeopteryx had a furcula (distally fused clavicles) like other birds, this feature was also present in many theropods (see
chapter 5). The shape of the pelvis of
Archaeopteryx—the pubis apparently pointed posteriorly—was very avian and also resembled that of some theropods. The long, bony tail of
Archaeopteryx also sets it apart from other birds.
The forelimb of
Archaeopteryx was theropod-like in some features. It had three fully developed digits and a flexible and unmodified wrist, radius, and ulna. Yet, the forelimb of
Archaepteryx was also very avian. It had claws adapted to tree climbing and bore an elliptical wing made of asymmetrical flight feathers.
The hind limb of
Archaeopteryx was also very bird-like. The head of the femur was turned medially, and the knee and ankle joints acted as hinges. The fibula was reduced, the proximal tarsals were fused to the tibia, and the distal tarsals were fused to the metatarsals. The long metatarsals were partly fused to each other, and there were three slender, forward-facing digits, as well as a fourth digit that faced backward.
Archaeopteryx evidently used its hind feet for climbing and perching. Many of these avian features of the hind limb of
Archaeopteryx were also present in some small theropods. Compare them, for example, in
Archaeopteryx and
Compsognathus, a theropod of similar size contemporaneous with
Archaeopteryx (see
figure 5.9).
There was actually only one feature of Archaeopteryx that long identified it as a bird. This was its feathers, which in structure and arrangement clearly ally Archaeopteryx with birds. To call Archaeopteryx a “feathered dinosaur,” or, in other words, to recognize it as an animal with an essentially theropod skeleton and avian feathers, is a reasonable conclusion.
Although most paleontologists agree that dinosaurs were the ancestors of birds, a vocal group of dissenters exists. Some of these paleontologists argue that birds evolved directly from Triassic archosaurs more primitive than dinosaurs. This old idea is difficult to disprove because Triassic archosaurs certainly were sufficiently primitive to have been the ancestors of birds. But, to argue for a Triassic archosaur as the ancestor of birds is to ignore the many evolutionary novelties shared by dinosaurs and birds that indicate they are more closely related to each other than either is to a Triassic archosaur. Indeed, those who so argue also claim that the similarities of dinosaurs and birds are largely the result of convergent evolution (
box 15.1).
Although first suggested in the nineteenth century, the dinosaurian ancestry of birds did not gain much credence until the 1970s. At that time, paleontologist John Ostrom drew attention to the many similarities shared by the skeletons of
Archaeopyteryx and theropod dinosaurs. He thus demonstrated that
Archaeopteryx had a basically theropod skeleton, with a few specializations seen in birds, and the asymmetrical flight feathers distinctive of birds. Subsequent cladistic analyses and discoveries of “feathered dinosaurs” in the Lower Cretaceous of northeastern China, including
Caudipteryx and
Microraptor, decided the dinosaurian origin of birds in the mind of most paleontologists. Yet, there remains a group of scientific skeptics who argue that the dinosaur–bird similarities are the result of evolutionary convergence inherited from very separate ancestors. To them, bird ancestry must be sought in archosaurs other than the dinosaurs.

BOX FIGURE 15.1
Scansoriopteryx, from the Middle Jurassic of China, has been identified as a maniraptoran dinosaur or an early bird. (Photograph and illustration © Stephen Czerkas)
The latest addition to this discussion is
Scansoriopteryx, a Middle Jurassic animal first described as a maniraptoran dinosaur (
box figure 15.1). In a recent restudy of this animal, paleontologist Stephen Czerkas and ornithologist Alan Feduccia argue that
Scansoriopteryx is a bird older than
Archaeopteryx. They assign it to Aves based on its perching foot, gliding wing, and asymmetrical wing feathers. Furthermore, Czerkas and Feduccia draw attention to the nondinosaurian features of
Scansoriopteryx, including the relative lengths of its arm and hand, the digit proportions of its hand, and its hip socket structure, which they point out is very different from the hip socket of upright walking dinosaurs.
Scansoriopteryx not only has feathered wings and arms, but a feathered hind wing. Czerkas and Feduccia argue that
Scansoriopteryx was a small arboreal bird not derived from dinosaurs. They see it as a four-winged arboreal glider that represents an early stage of avian evolution prior to
Archaeopteryx. As Czerkas and Feduccia conclude, “The origin of Aves is all the more complex because convergence within Dinosauria has obscured the true avian ancestry that, instead of being from within Dinosauria, is actually from nondinosaurian archosaurs.” The debate goes on!
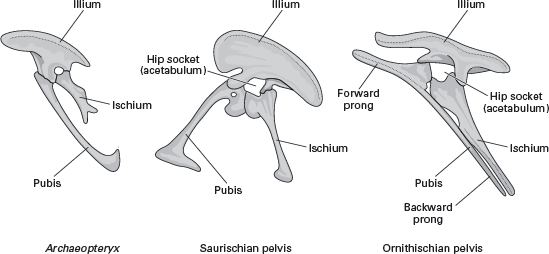
FIGURE 15.8
The avian pelvis is a modified saurischian pelvis that superficially resembles the ornithischian pelvis. (© Scott Hartman)
One challenge to the dinosaurian origin of birds comes from the claim that the three digits of the theropod hand are not the same digits as those in the hand of a bird. The dinosaur digits are supposedly I-II-III (the “thumb” is digit I), whereas the bird wing digits are supposedly II-III-IV. This means some evolutionary convergence must have occurred to produce strikingly different forelimbs in theropod dinosaurs and birds (see
figure 15.5). However, this claim has been countered by analysis that suggests that such a shift in digits could be achieved by a relatively minor gene change in the embryonic development of the earliest birds.
Two ideas about the origin of birds to avoid are that the ornithischian dinosaurs and the pterosaurs were avian ancestors. Certainly, the pelvis of birds does somewhat resemble that of the ornithischian (“bird-hipped”) dinosaurs. But a closer look reveals that the avian pelvis is actually a highly modified saurischian pelvis (
figure 15.8). Thus, the pubis of the saurischian pelvis was shifted backward, the ischium was shortened, and the posterior portion of the ilium was lost to produce an avian pelvis.
Pterosaurs were flying archosaurs that first appeared during the Late Triassic and became extinct at the end of the Cretaceous. Pterosaurs lacked feathers and the evolutionary novelties shared by dinosaurs and birds. As noted in
chapter 4, pterosaurs were archosaurs closely related to dinosaurs, but that’s as close as their relationship was to birds. Flight must have evolved twice, quite separately, among the archosaurs—once in pterosaurs and once in birds.
Because the hallmarks of birds, including Archaeopteryx, are their feathers and other skeletal modifications for flight, it is natural to ask how avian flight originated. Two major hypotheses have been advanced and remain the subject of discussion.
The oldest idea is that the immediate ancestors of
Archaeopteryx lived in the trees; they were arboreal. These arboreal bird ancestors evolved feathers to glide from branch to branch and to the ground. Flight powered by flapping the wings evolved later. Indeed, Archaeopteryx itself has claws that indicate it was capable of climbing trees. This hypothesis of the origin of avian flight can be termed the
arboreal hypothesis.
An alternative hypothesis is that
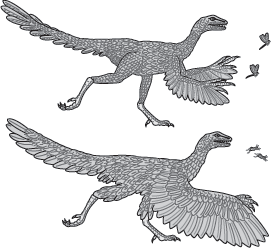
Archaeopteryx and its immediate ancestors were ground-living, fast runners (cursorial). According to this hypothesis, feathers evolved to insulate the dinosaur and, on the arms, may have provided additional surface area to aid in catching insects (
figure 15.9). Enlargement of these feathers followed to provide support and thrust when the newly evolved wings were flapped. This hypothesis can be termed the
cursorial hypothesis.
A criticism of the arboreal hypothesis points out that the wings of gliders, such as “flying” squirrels, are very different from those of birds. For example, in gliders, the wing membrane is attached to the body and extends between the fore and hind limbs, very different than the wing membrane of birds. Indeed, Archaeopteryx was not an arboreal glider, so no evidence is available for this gliding stage of evolution required by the arboreal hypothesis.
The arboreal hypothesis argues that birds took advantage of gravity, evolving at small size to take advantage of the “cheap energy” it provides when jumping out of a tree. Indeed, flight in arboreal gliders (lizards, snakes, mammals), pterosaurs, and bats has long been agreed to have started in high places. The relatively recent discovery of small, feathered theropods, such as Microraptor, that look like they could climb trees, also lends support to the arboreal origin of avian flight.
The principal criticism of the cursorial hypothesis is that it is challenged by gravity. Furthermore, the development of wings and feathers on the ground apparently indicates they first evolved for some function other than flying. This led to the idea that catching insects with feathered hands may have originally driven the evolution of the feathered wing. But, some paleontologists have difficult imagining what adaptive advantage the early stages of flapping the arms would have conferred on theropods. If anything, this might have slowed a theropod trying to catch insects or running away from a predator.
As is often the case with two competing ideas in science, the correct answer may blend portions of both. The currently known array of maniraptoran theropods and other possible bird ancestors includes animals capable not only of ground running but also of running up into trees, climbing in trees, and gliding from branches (
box 15.2). Feathers may have first evolved as insulation and camouflage and later developed further into aerofoils used in flying. In other words, the early steps of bird origins may have moved from the ground and then into the trees, thus combining elements of both hypotheses.
FIGURE 15.9
Two hypothetical stages in the cursorial origin of avian flight show a proto-bird catching insects with feathered hands (above) and a later, winged proto-bird like Archaeopteryx (below). (Drawing by Network Graphics)
The discovery of very bird-like theropods in the Lower Cretaceous lake beds of northeastern China was at first heralded as evidence establishing dinosaurs as the ancestors of birds. Indeed, dromaeosaurs, such as Microraptor, had asymmetrical flight feathers, the hallmark of birds. If such feathers diagnose birds, how could such dromaeosaurs be distinguished from birds? Isn’t Microraptor (and similar feathered dromaeosaurs) a bird?
Some paleontologist say yes! They identify many of the bird-like features of dromaeosaurs, such as Microraptor, as evidence that dromaeosaurs are descendants of birds, not dinosaurs. Indeed, some paleontologists have suggested that many of the maniraptorans are secondarily flightless descendants of birds. But, not all paleontologists agree with this.
BOX FIGURE 15.2
Mononykus, from the Upper Cretaceous of Asia, has been called a flightless bird or a theropod dinosaur. (© Scott Hartman)
One of the most bizarre of the maniraptorans is
Mononykus, from the Upper Cretaceous of Mongolia, with its very peculiar, spike-like forelimbs (
box figure 15.2). Although originally described as a flightless bird, some paleontologists argue that
Mononykus is a bizarre theropod dinosaur. When it comes to deciding whether some maniraptorans were birds or dinosaurs—the choice can truly be difficult! Stay tuned for more on this subject.
A book on dinosaurs would be remiss if it did not include a brief overview of the evolution of the descendants of dinosaurs, the birds.
Despite claims of older birds, most paleontologists regard Late Jurassic
Archaeopteryx as the oldest known bird. Cretaceous birds were much more advanced than
Archaeopteryx, although some still had teeth (
figure 15.10). The past few decades have seen the discovery of many Early Cretaceous birds from China (
box 15.3). These include long-tailed, clumsy fliers like
Confuciusornis, perhaps the best known Cretaceous bird represented by more than 1,000 fossil specimens (
figure 15.11).
Indeed, a diverse group of early birds appeared during the Cretaceous, the enantiornithines. These are the so-called “opposite birds” because the articulation of their shoulder bones, the coracoid and scapula, appears to be opposite to the anatomy seen in modern birds. Enantiornithine bird fossils are best known from the Cretaceous of China, Argentina, and Spain. Sinornis, from the Lower Cretaceous of China, is a representative enantiornithine.
FIGURE 15.10
Wingless diving birds of the Early Cretaceous, such as Baptornis, still had teeth. (Drawing by Network Graphics)
During the Early Cretaceous, northeastern China was a land of towering volcanoes and deep lake basins. Paper-thin layers of shale and limestone deposited in these lakes preserved a remarkable fossil record, including a surprising diversity of very modern-looking early birds. The first of these birds to be described, the sparrow-size
Sinornis, was a fully powered flier capable of perching in trees (
box figure 15.3). This primitive bird had a very archaic skull and skeleton that included teeth, a clawed hand, and limited fusion of the limb bones. Yet
Sinornis flew and perched very much like a modem bird. There is a huge evolutionary distance between
Sinornis and the oldest bird,
Archaeopteryx; evidently, during the 25 million years that separates them, much evolution took place in the early history of birds.
BOX FIGURE 15.3
Sinornis, from the Lower Cretaceous of China, was a surprisingly advanced bird. (© Scott Hartman)
Sinornis was only one of many new birds recently described from the Lower Cretaceous of northeastern China. One of the most remarkable of these is Confuciusornis, known from hundreds of specimens preserved with feather impressions. Confuciusornis is about the same size as Archaeopteryx. But, even though Confuciusornis is the oldest known toothless bird, it still has a clawed hand and some other primitive features. Confuciusornis is thus a classic example of what has been called mosaic evolution; it has a mixture of primitive (dinosaur-like) and advanced (modern bird–like) features. Confuciusornis, Sinornis, and other Early Cretaceous birds from China, as well as other Early Cretaceous birds from Spain and Mongolia, attest to an explosive early evolution of birds after their first appearance during the Late Jurassic. Birds were clearly very successful from the start.
During the Early Cretaceous, highly specialized, flightless diving birds also evolved. This early diversity among birds may indicate an ancestry older than
Archaeopteryx or that other Late Jurassic birds remain to be discovered. A major change took place by the end of the Cretaceous when the enantiornithines totally disappeared, and primitive, toothed birds also became extinct. Indeed, one paleontologist has referred to the Cretaceous as the “age of archaic birds.” Truly modern, toothless birds appeared by the beginning of the Cenozoic. During the past 66 million years, birds have been one of the most successful groups of vertebrates (
figure 15.12).
FIGURE 15.11
A superbly preserved Confuciusornis from the Lower Cretaceous of northeastern China (left) with detail of its tail feathers (right). (Courtesy Zhang Zihui)
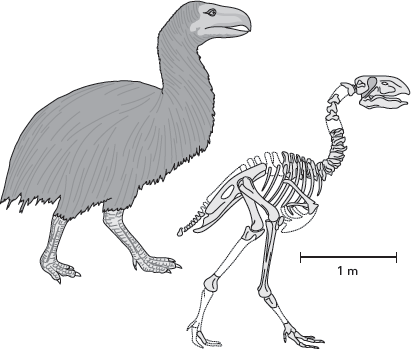
Birds have delicate, hollow bones; most are small animals; and many live in dry or forested environments. For these reasons, the fossil record of birds is less extensive than that of reptiles and mammals. Yet, despite this, we have fossils of water birds, land birds, and giant, flightless birds (
ratites) at least as far back as the Paleocene. The giant, flightless birds are particularly interesting because they evolved from flying ancestors and so represent a return to the flightless habits of the theropod ancestors of birds (
figure 15.13).
FIGURE 15.12
Birds have been very diverse and successful and underwent a Cretaceous diversification of archaic birds, followed by a second, Cenozoic, diversification of modern birds. (© Scott Hartman)
FIGURE 15.13
Giant, flightless birds, such as Diatryma, appeared as early as 58 million years ago. (Drawing by Network Graphics)
As stated at the beginning of this chapter, identifying theropod dinosaurs as the ancestors of birds should have a major impact on how we view the dinosaurs. We should no longer think of them as reptile-like in many aspects of their biology. Instead, much about dinosaur biology and behavior was very avian, as explained elsewhere in this book.
If dinosaurs were so bird-like, should we classify them with reptiles? Perhaps not, although various answers to this question have been proposed (
box 15.4). Regardless of the classification used, the close relationship of birds to dinosaurs is well established.
The cladogram of theropod dinosaurs presented earlier in this book (see
figure 5.1) can be modified slightly to show birds where most paleontologists believe they belong, as close relatives of the maniraptoran theropods (
box figure 15.4). If this cladogram is turned directly into a classification, that classification would simply identify birds as a particular type of tetanuran. In this classification, birds are just another type of dinosaur.
Some paleontologists have taken such a classification to heart. They cast doubt on the significance of “dinosaur extinction,” because, after all, the avian dinosaurs (= feathered dinosaurs, = birds) are still with us, some 10,000 species strong (see
chapter 16). These paleontologists speak flippantly of “carving the dinosaur” at a Thanksgiving dinner. In a more serious vein, they argue that putting the birds in a class distinct from that which contains the dinosaurs obscures their close relationship.
This may be the case, but putting birds into the same class as dinosaurs also obscures the key evolutionary novelties of birds: their feathers, skeletal features, and physiological mechanisms for sustained, powered flight. Despite the many similarities of theropod dinosaurs to birds, and their shared evolutionary novelties, birds are unique vertebrates. It is this uniqueness, and the great diversity of birds, that long ago led to their recognition as a separate class of vertebrates called Aves. For these reasons, some paleontologists prefer to distinguish this class from dinosaurs. But, others place dinosaurs (or at least theropod dinosaurs) in the Aves, and, as mentioned earlier, still others abandon Aves as a separate class and include birds in the Theropoda. Another solution was to introduce the term “Avialae” to encompass all the early birds, including Archaeopteryx, as distinct from birds as we know them, the Aves.
These differing classifications of birds reflect different philosophies of how a phylogeny should be turned into a classification; they do not reflect different ideas on the phylogenetic relationships of birds. All but a few paleontologists agree that the birds, or the nondinosaurian Aves, or the avian theropods, or whatever you choose to call them, were descended from theropod dinosaurs.
BOX FIGURE 15.4
This cladogram indicates that birds are closely related to dromaeosaurs. (Drawing by Network Graphics)
1. Birds are feathered vertebrates with skeletons highly modified to be lightweight and rigid to enable sustained powered flight.
2. Archaeopteryx, from the Upper Jurassic of Germany, is the oldest known bird.
3. Many skeletal features of Archaeopteryx are very similar to those of small theropod dinosaurs.
4. Evolutionary novelties of the skeleton shared by theropod dinosaurs and birds provide strong evidence that birds descended from dinosaurs.
5. Some paleontologists argue that the dinosaur–bird similarities are the result of evolutionary convergence and that the ancestry of birds lies among more primitive archosaurs.
6. There are two hypotheses of the origin of avian flight: the arboreal and the cursorial hypotheses.
7. The correct hypothesis of the origin of avian flight may blend portions of both the arboreal and cursorial hypotheses by recognizing possible bird ancestors capable not only of ground running but also of running up into trees, climbing in trees, and gliding from branches.
8. Modern birds evolved by the beginning of the Cenozoic.
9. Recognition of dinosaurs as the ancestors of birds forces us to rethink much of dinosaur anatomy and behavior and stress their bird-like aspects.
air sac
arboreal hypothesis
Archaeopteryx
Aves
bird
Confuciusornis
cursorial hypothesis
enantiornithine
feather
furcula
“missing link”
Mononykus
pneumatic
pneumatic duct
ratite
Scansoriopteryx
Sinornis
Solnhofen Limestone
1. How is a bird’s skeleton designed to make powered flight possible?
2. What evolutionary novelties were shared by theropod dinosaurs and birds?
3. Why do most paleontologists consider Archaeopteryx to be a bird? What similarities does it show to theropod dinosaurs?
4. What other possible ancestors of birds have been identified, and what might they suggest about the similarities of dinosaurs and birds?
5. Compare and contrast the arboreal and cursorial hypotheses of the origin of avian flight.
6. What do Cretaceous birds suggest about the origin and early diversification of the Aves?
7. What impact does their recognition as the ancestors of birds have on our ideas about dinosaurs?
Chiappe, L. M. 2007. Glorified Dinosaurs: The Origin and Early Evolution of Birds. Hoboken, N.J.: Wiley. (Book-length treatment of the origin of birds from dinosaurs)
Currie, P. J., et al., eds. 2004. Feathered Dragons: Studies on the Transition from Dinosaurs to Birds. Bloomington: Indiana University Press. (A collection of 14 articles on theropods and birds)
Czerkas, S. A., and A. Feduccia. 2014. Jurassic archosaur is a non-dinosaurian bird. Journal of Ornithology 155:841–851. (Argues that Scansoriopoteryx is a bird not descended from dinsoaurs)
Feduccia, A. 2014. Avian extinction at the end of the Cretaceous: Assessing the magnitude and subsequent explosive radiation.
Cretaceous Research 50:1–15. (Argues for a major extinction of birds at the end of the Cretaceous followed by the explosive diversification of modern birds)
Feduccia, J. A. 1996. The Origin and Evolution of Birds. Cambridge, Mass.: Harvard University Press. (A comprehensive review of the fossil record of birds, with a particularly detailed treatment of their Cenozoic evolution)
Feduccia, J. A. 2014. Riddle of the Feathered Dragons: Hidden Birds of China. New Haven, Conn.: Yale University Press. (Book-length treatment of the origin of birds, but from the perspective that they did not evolve from dinosaurs)
Naish, D. 2012. Birds, pp. 379–423, in M. K. Brett-Surman, T. R. Holtz, Jr., and J. O. Farlow, eds., The Complete Dinosaur. 2nd ed. Bloomington: Indiana University Press. (Concise review of bird origins and evolution)
Ostrom, J. H. 1976. Archaeopteryx and the origin of birds. Biological Journal of the Linnaean Society 8:91–182. (A comprehensive and now classic technical article on the origin of birds)
Paul, G. S. 2002. Dinosaurs of the Air: The Evolution and Loss of Flight in Dinosaurs and Birds. Baltimore: Johns Hopkins University Press. (Argues that many bird-like theropods are the flightless descendants of birds)
Pickrell, J. 2014. Flying Dinosaurs: How Fearsome Reptiles Became Birds. New York: Columbia University Press. (Very readable account of the discoveries, debates, and debacles of the science of the dinosaur–bird connection)
Xu, X., et al. 2003. Four-winged dinosaurs from China. Nature 421:335–340. (Provides a description of Microraptor)
To find the oldest fossil bird, Archaeopteryx, you must look hard, because Archaeopteryx fossils are rare. The pigeon-size Archaeopteryx remains one of the rarest of fossils, known only from about a dozen specimens collected from Upper Jurassic rocks in southern Germany. The famous Berlin specimen can be seen at the Museum für Naturkunde in Berlin, Germany. At the Natural History Museum in London, England, the London Specimen is on display. Other Archaeopteryx fossils on display in Europe are in smaller museums in the Netherlands and Germany. Closer to home, what has been called the Thermopolis specimen is on display at the Wyoming Dinosaur Center in Thermopolis. Excellent replicas (casts) of Archaeopteryx abound, particularly of the Berlin and London specimens. They are on display at many museums worldwide.