Evolution of Apparently Nonadaptive Behavior
Nathan W. Bailey
OUTLINE
1. What is apparently nonadaptive behavior?
2. Behavior as a transaction
3. Random mutation versus adaptation: Cannibalism
4. Manipulation: Imposter birds and zombie snails
5. Evolution does not equal perfection: Sexual cannibalism
6. Same-sex sexual behavior: A case study
7. Insights from apparently nonadaptive behavior
Behaviors appear to be nonadaptive when their costs, in terms of reproductive fitness, appear to outweigh their benefits. As long as there is genetic variation, selection should eliminate such behaviors under those conditions. However, apparently nonadaptive behaviors are much more common than might be expected. Examples are numerous and include counterintuitive responses to infection by pathogens or parasites, sexual cannibalism, and same-sex sexual behavior. This chapter explores how an evolutionary framework can be used to understand how and why such behaviors evolve, and what causes them to be maintained within populations despite what appear to be fitness disadvantages.
GLOSSARY
Altricial. The condition of offspring in which they are unable to feed or fend for themselves without considerable assistance from parents for a period of time.
Altruism. A behavior that benefits a recipient individual at the expense of the fitness of the actor (contributor).
Apparently Nonadaptive Behavior. Behavior that appears to impose a fitness cost on the actor with no compensating benefit.
Arms Race. Cycles of adaptations and counteradaptations leading to the evolution of novel morphological or behavioral traits, occurring between two or more interacting species, or in some cases, between the sexes.
Brood Parasite. In birds, a species that exploits the reproductive behavior of another species by laying an egg in its nest. The chick is then fed by the host species at the expense of the host’s offspring.
Filial Cannibalism. The eating of offspring by their parents.
Overdominance. Heterozygote advantage; when alleles in the heterozygous condition increase fitness but decrease it when in the homozygous condition.
Parasite Manipulation. Behavioral, physiological, or morphological changes induced in a host by a parasite that increase the likelihood of transmission of the parasite.
Phenotypic Plasticity. The production of different phenotypes by a single genotype, depending on the environment in which it is expressed.
Reproductive Fitness. An individual’s contribution of offspring in the next generation, relative to other individuals in the population.
Same-Sex Sexual Behavior. Sexual behavior, such as courtship, mounting, or copulation, occurring between individuals of the same sex that also occurs between individuals of the opposite sex.
Sexual Antagonism. The expression of genes (or a phenotype) in one sex that confer a fitness advantage, but when expressed in the other sex cause a fitness disadvantage.
Sexual Cannibalism. The killing and consumption of a mate during or after copulation.
1. WHAT IS APPARENTLY NONADAPTIVE BEHAVIOR?
Evolutionary theory provides a robust framework for explaining the diversity of the natural world. Behavior contributes considerably to that diversity, but understanding how and why animals behave the way they do poses challenges, because many behaviors appear to be costly and nonadaptive. The power of evolutionary theory lies in its ability to generate testable hypotheses to explain the reasons behind even the most bewildering traits, and there are a large number of bewildering behaviors from which to choose.
Wire-tailed manakins (Pipra filicauda) appear to go out of their way to attract predators and physically exhaust themselves during courtship. Males have a conspicuous yellow breast, a bright red cap, and long black filamentous tail feathers, and they engage in spectacular behavioral displays to elicit attention from females. They fluff up the feathers on their back, wiggle their tail, bob their head up and down and back and forth, quiver their outstretched wings, and leap and bounce rapidly from side to side along horizontal branches, all the while emitting a series of buzzing and whistling vocalizations. Any biologist observing these extraordinary behaviors could be forgiven for wondering how such a flamboyant repertoire could ever evolve, when it clearly requires an enormous energetic expenditure and appears to invite predation. How could this behavior possibly be adaptive for a manakin?
Closer examination of the natural history of many animal species will reveal similar examples of behaviors that appear to be nonadaptive—in other words, behaviors that appear to decrease rather than increase fitness. The purpose of this chapter is to highlight how the framework of evolutionary theory can be used to study such behaviors and to illustrate how historical studies of puzzling behaviors have provided powerful, sometimes surprising, evolutionary insights. It concludes with an analysis of same-sex sexual behavior as a topical case study in apparently nonadaptive behavior.
As with all good theories, the apparent exceptions provide the best test of a theory’s robustness and generality. Behaviors that appear to defy evolutionary logic have historically provided some of the most decisive verification of the basic principles of evolution, and research in the field continues to focus on identifying the adaptive value of counterintuitive behaviors, understanding their genetic basis, and working out how they evolve. This evolutionary approach has a long history. The Nobel Prize–winning ethologist Konrad Lorentz wrote in the 1960s that aggression was more likely to occur between individuals of the same species than between individuals of different species. Lorentz puzzled over why animals of the same species behave aggressively toward one another. Aggression is risky and costly. Predation is likely to arrive in the form of a different species, and disease and parasitism present yet other challenges, so if an animal is to expend energy being aggressive, why waste it on a conspecific? Lorentz’s logic was that the fiercest competition for resources should occur among members of the same species, whether the resources are materials such as food, water, or nesting substances, or reproductive such as access to mates. In 1973 John Maynard Smith and George Price formalized the evolutionary dynamics of animal conflict using game-theoretical models to clarify the individual benefits of aggressive behavior (see chapter VII.3). There are many other scientists whose theoretical advances have been motivated by puzzling animal behavior. Around the same time as Lorentz, William D. Hamilton developed the theory of kin selection to help explain why animals sometimes behave altruistically, conferring benefits on others at great cost to their own survival or reproduction. The scientific study of apparently nonadaptive behaviors was catalyzed by Charles Darwin, who proposed sexual selection as a mechanism underlying the evolution of conspicuous, costly traits, such as the displaying behavior of the manakins. But are these mechanisms sufficient to explain the persistence of all apparently nonadaptive behaviors?
2. BEHAVIOR AS A TRANSACTION
All animals behave. And all behaviors carry costs and benefits. The currency of this transaction can be thought of as reproductive fitness, or the share of the genes in the next generation contributed by a parent. More genes, through the production of more offspring relative to other parents in the population, equate to higher fitness. Some behaviors obviously increase the fitness of the actor. For example, a guppy initiating an escape response to a predator clearly increases its fitness. But how is this behavior evaluated in terms of costs and benefits? In the case of the fleeing guppy, the costs are not terribly high—they might include a bout of energetic expenditure and time taken away from eating or reproducing. But the benefit is obvious: not dying. Unfortunately, the costs and benefits of other behaviors are not always so obvious, and in many cases the costs appear to exceed the benefits.
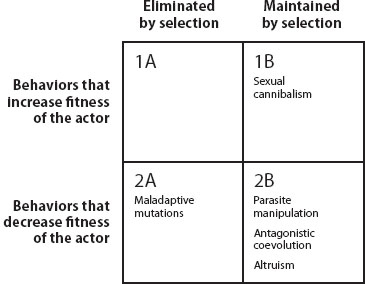
There are a number of evolutionary explanations for the existence of behaviors that give the appearance of imposing a fitness cost that is not compensated for by some other benefit, and they can be divided into two broad categories. The first is behavior that appears to decrease fitness but in reality is evolutionarily maintained within populations, whereas the second is behavior that appears to decrease fitness and is in fact selected against. It is important to note, however, that just because they appear to decrease the fitness of the individual engaging in them does not mean such behaviors always do decrease fitness. Sometimes the fitness benefit is “hidden” and can be revealed through experimentation. Nevertheless, in many cases behaviors that appear to be nonadaptive actually decrease the actor’s fitness but are still evolutionarily maintained (figure 1).
Figure 1. An evolutionary framework for studying apparently nonadaptive behavior. The horizontal rows indicate fitness effects of the behavior on the actor, and the columns indicate how selection acts on the behavior. Examples are given in each quadrant. The top row indicates apparently nonadaptive behaviors that have a “hidden” fitness benefit; in other words, the behavior yields a benefit that is not immediately apparent to scientific observers. Fitness-increasing traits can be counterselected (1A), but this is unlikely to happen as a result of direct selection on the trait; for example, the behavior might be lost because it is genetically correlated with another fitness-reducing trait. Apparently nonadaptive behaviors that increase fitness are likely to be evolutionarily maintained (1B), as in the case of sexual cannibalism in spiders. Some behaviors that appear nonadaptive truly do lower individual fitness (row 2) and are therefore counterselected (2A). However, most apparently nonadaptive behaviors of interest fall into the category of behaviors that incur a fitness cost to the individual, yet are nonetheless evolutionarily maintained in populations (2B). These include behaviors resulting from manipulation by other agents, behaviors arising because of selection at a level different from the individual, or correlated responses to selection such as sexually antagonistic coevolution.
3. RANDOM MUTATION VERSUS ADAPTATION: CANNIBALISM
Behaviors that are not evolutionarily maintained are not ordinarily observed or studied, because, by definition, selection has eliminated them. These include behaviors that arise as a result of random genetic mutation and confer a fitness disadvantage. However, lab studies frequently use mutant strains of Drosophila or mice containing genes that have been partially or wholly inactivated, and these are useful for illustrating how genetic mutations can affect the adaptive value of a behavior.
A mutant strain of lab mice called “Tokyo” provides a clear example of the genetic basis for a nonadaptive behavior. The Tokyo strain expresses a mutant vitamin D receptor, VDR. One of VDR’s ordinary functions is to act as a transcription factor than binds both DNA and vitamin D, but in the Tokyo strain this function is diminished because the mutant protein lacks a DNA-binding domain. Vitamin D is a hormone that regulates a variety of neurological processes underlying behaviors, and Allan Kelueff and his colleagues found that among other behavioral abnormalities, all female Tokyo mutants cannibalized all their pups, whereas females that did not express the mutant gene did not cannibalize any of their young. The mice in this study were kept in conditions that would not normally provoke offspring killing (filial infanticide), and it is reasonable to predict that a comparable mutation arising in a wild population of mice would not be evolutionarily maintained.
The killing of one’s own offspring seems so patently maladaptive that it would be easy to dismiss the phenomenon as unusual, pathological, and therefore of little interest. However, offspring killing is surprisingly common. Infanticide has been documented in many mammal species, from rodents to primates, in addition to birds, insects, fish, and rotifers, and it has different manifestations. There are many appealing adaptive explanations for killing the offspring of an unrelated conspecific, but it is less clear how it could be beneficial for a parent to kill its own young. Nevertheless, viewing filial infanticide through the lens of evolutionary theory and identifying the fitness costs and benefits can help reveal reasons for its persistence.
Sometimes it is advantageous for a female to kill her young if she gains resources from them, for example, by eating them. In the burying beetle Nicrophorus vespilloides, food for developing larvae is limited to a single liquefying animal carcass that the parents have prepared in advance. If the carcass is too small to support all the offspring that hatch and begin to develop, the parents have a strategy to cut their losses and ensure that at least some of them survive, rather than face the complete loss of the brood. That strategy is to kill and eat some—but not all—of their offspring. Wiping out a portion of the brood ensures that the remaining larvae have enough food to survive, and it has the added advantage of providing the parents with a meal. In other species, a female may sacrifice her young if the cost to her future reproductive success of rearing those young is greater than the cost of simply killing them and waiting to reproduce at a later date. The costs of rearing young are not insignificant in altricial species and particularly in mammals with long, energetically expensive periods of lactation, so the cost-to-benefit ratio of filial infanticide can shift dramatically during times of low resource availability or increased stress. For example, female mice (Mus musculus) can reduce their litter size when food is scarce. Sometimes this reduction is accompanied by filial cannibalism. In studies that examine this behavior in mice, it can be difficult to distinguish between active infanticide (i.e., physically killing pups) and simply allowing pups to die from starvation and then eating them. Intriguingly, however, female mice have the capacity to resorb developing fetuses, and they are more likely to do so when conditions are not favorable for rearing offspring.
4. MANIPULATION: IMPOSTER BIRDS AND ZOMBIE SNAILS
Animals do not always behave in their own best interests, and frequently that is because one individual is being manipulated to serve another’s needs. Some parasitic relationships resemble a marionette being controlled by an invisible puppeteer: parasites and pathogens can invade the body of their host and induce it to behave in a way that benefits the parasite at the expense of the host. In other cases, the manipulation is accomplished from afar. Either way, fascinating nonadaptive behaviors can result. A classic example is manipulation by cuckoos (Cuculus canorus). Female cuckoos lay mimetic eggs in host nests, and newly hatched cuckoo chicks, without any instruction or provocation, quickly set about pushing the host parent’s eggs up the wall of the nest on their tiny back, and then hurling them over the edge to their demise. Despite the obvious phenotypic differences between a cuckoo chick and what a host’s chick ought to look like, host parents appear to overlook the destructive behavior of the invader and instead expend a remarkable amount of time and energy feeding and protecting the developing cuckoo.
But how and where is this behavior nonadaptive? Cuckoo-feeding behavior is maladaptive for the host, because it reduces its own reproduction, but of great benefit to the brood parasitic bird by increasing the production of offspring, and this imbalance creates interesting evolutionary dynamics. What is adaptive for one species in the interaction is not adaptive for the other, but selection acts on both. Selection on hosts should result in the evolution of traits that decrease the risk of being deceived, while selection on the cuckoos should be expected to increase the ability to deceive hosts. These countervailing pressures can result in an evolutionary arms race wherein host species and parasite species evolve adaptations and counteradaptations, and many cuckoo studies support predictions made by the arms-race hypothesis. For example, because host species should evolve parasite egg discrimination, one would expect the brood parasites to evolve ever-better egg mimicry. Such evolution might in turn select for hosts that can distinguish foreign nestlings after they have hatched, and that is exactly what has been found in the superb fairywren (Malurus cyaneus), which is parasitized by two species of bronze-cuckoos (Chrysococcyx spp.). However, a theoretical study by Maria Servedio and Russell Lande showed that it is also possible for host egg discrimination and cuckoo egg mimicry to evolve to a stable point at which there is imperfect egg mimicry, weak host discrimination, and a slight tendency of hosts to reject cuckoo eggs.
Parasitic microorganisms that infect a host can also elicit nonadaptive behaviors by hijacking their host’s central nervous system to induce behaviors that increase their likelihood of transmission. Many neurophysiological mechanisms for manipulating host behavior have been identified. Pathogens can infect host neurons and disrupt their function, cause inflammation of the central nervous system by triggering host immune responses, induce neuronal apoptosis, and affect neurotransmitter and hormone levels in discrete structures of the host’s brain.
A fascinating example is that of snails infected with the trematode parasite Leucochloridium paradoxum. For this trematode flatworm to reproduce, it must migrate from its intermediate snail host to its definitive host, a bird. The trematode starts out as an egg within a snail. The egg hatches into a larva called a miracidium, and then the miracidium matures into a sporocyst. Multiple sporocysts migrate to the snail’s eye tentacles and form swollen, colorful brood sacs that throb rhythmically and resemble caterpillars. Infected pulsating snails further compound their doom by exhibiting positive phototaxis, which means they uncharacteristically move toward sunlight instead of away from it, thereby increasing their vulnerability to bird predators. Once inside a bird, L. paradoxum continues its life cycle. A similar locomotory manipulation occurs in insects infected with nematomorph worms. These extremely long, thin worms reproduce only in water, and during their infectious stage they impel the ordinarily land-bound insect host within which they are tightly packed to find water, jump in, and drown so the worms can emerge to swim away and mate.
Although host manipulation is clearly an adaptation for the parasite, these behaviors are not adaptive for the host—that is, the benefits to the host clearly do not outweigh the costs. A good deal of debate surrounds the question of whether selection has acted on parasites to be able to manipulate their hosts, or whether these abnormal behaviors are just by-products of physiological responses hosts mount when they are invaded. However, they differ from other behaviors that hosts mount during a pathological infection to kill pathogens or mitigate the negative effects of infection, such as the behavioral fever some insects induce by moving to a warmer area. As Richard Dawkins articulated in the early 1980s, nonadaptive behaviors often represent the influence of adaptations in other individuals, via the extended phenotypic effects of their genes. Viewing behavior as more than just a linear output starting from neurological processes operating within one individual’s body allows a much richer understanding of the selective pressures that maintain apparently nonadaptive behaviors. Behavior is by definition especially responsive to external influences, either from the physical environment or from social interactions with other individuals. This property makes it susceptible to the manipulative effects of genes carried in other individuals. However, the malleability of behavior can also be a major asset to an animal facing an uncertain environment.
5. EVOLUTION DOES NOT EQUAL PERFECTION: SEXUAL CANNIBALISM
No organism is perfectly adapted to its environment, because the environment consists of continually changing, interacting forces. These can be physical, such as ambient temperature or the pH of an aquatic habitat, or they can be biotic, for example, the extent of canopy cover in a rain forest or the abundance of a keystone species in an ecosystem. Social interactions among conspecifics can be particularly variable depending on the behavioral, morphological, or physiological traits expressed by interacting individuals. In such a constantly changing world, it often pays to be flexible.
Phenotypic plasticity (see chapter III.10 on reaction norms) describes the capacity for a single genotype to produce different trait values depending on the environmental conditions in which that genotype is expressed. Phenotypic plasticity is responsible for much of the behavioral variation that evolutionary biologists and behavioral ecologists observe and study. For example, in many amphibians and insects that use acoustic signals to attract mates, males can sing for females, or they can adopt a silent satellite strategy in which they position themselves close to singing males and then intercept females responding to the caller. Which strategy they adopt depends on a number of factors such as the population density, risk of predation, and their own body condition. Similarly, many female birds develop song preferences early in life. These preferences depend on nuances of the male songs to which they are exposed during a critical period of imprinting, which means that whom they choose to mate with as an adult depends greatly on the males they encountered as juveniles.
In an ideal world, selection would favor individuals with the ability to perfectly shape morphology, physiology, and behavior to whatever physical or social environment they happen to encounter, but this does not happen. Plasticity, like any other trait, has costs and limitations. Organisms must possess the necessary sensory organs and central nervous system circuitry to detect environmental variation, and then they must produce an appropriate response. The environment can also fluctuate a great deal, and in some cases environmental cues can be inaccurate.
Many other factors impede perfect adaptation. The physics of surface-to-area relationships limits how large or small animals can grow at a given temperature. The laws of chemistry and electricity dictate how fast nerve impulses can travel. Protein biochemistry determines the flexibility, elasticity, permeability, and durability of biological substances. The work of Geoff Parker and John Maynard Smith was instrumental in describing how selection may drive organismal evolution toward an optimal phenotype, but fluctuating environments compounded with other constraints such as these will constantly undermine the process.
Consider sexual cannibalism. Male redback spiders (Latrodectus hasselti) are tiny compared with females. They copulate by depositing sperm onto the end of one of their pedipalps, which are paired sensory appendages projecting from either side of the mouth, and inserting it into a female’s genital opening. Shortly after initiating this maneuver, the male flips his entire body directly into the female’s mandibles using his still-inserted palp as a hinge. Sometimes the female pierces him, injects digestive juices, and then sucks out his liquefied tissues (figure 2). The male thus sacrifices any opportunity for additional mating by becoming a snack for the female.
Figure 2. Female redback spider (Latrodectus hasselti) with cannibalized former mate hanging in the web above her fangs. (Photo by Ken Jones; copyright M.C.B. Andrade 2002.)
Maydianne Andrade and her colleagues have studied the behavioral and evolutionary dynamics of the stereotyped flipping behavior males engage in while mating, and they found that perhaps not surprisingly, it increases the likelihood that the female will eat the male. Males are especially prone to being eaten when the female is hungry. However, Andrade’s lab has also found that males that get eaten sire a greater proportion of the resulting offspring compared with males that escape, and this result is likely driven by the fact that females are less inclined to remate after eating a male.
At first glance it seems easy to explain why males commit nuptial suicide: males that do it sire more offspring. But surely there is a better way to sire more offspring that doesn’t involve killing yourself and thereby removing all possibility of reproducing in the future? A key question is, Why hasn’t evolution favored L. hasselti males that escape and go on to inseminate other females, perhaps donating a different, nonlethal, type of food item, as happens in many insect species? The answer lies in the fact that L. hasselti males almost never remate. In fact, they usually are not able to. The sperm-holding organs on their palps snap off after mating, and their chances of surviving long enough to even find another female are negligible because they are so tiny (figure 2). The evolution of self-sacrificing behavior makes much more sense in light of these vital constraints: if there is no chance of fathering more offspring in the future, and males do not provide direct care or resources to their young, then there is no fitness advantage to staying alive.
6. SAME-SEX SEXUAL BEHAVIOR: A CASE STUDY
The final portion of this chapter is devoted to a behavior that appears to be nonadaptive and has attracted increasing academic scrutiny over the last several decades: same-sex sexual behavior (SSB). The aim here is to demonstrate how evolutionary causes of a seemingly puzzling behavior, SSB, can be digested into different hypotheses to yield predictions that can then be tested. This process provides an example of the deductive power of evolutionary reasoning for exploring the ultimate reasons for the existence of apparently nonadaptive behaviors, by suggesting potential benefits to outweigh suspected costs.
Same-sex sexual behavior is present in many species, can occur in both sexes, and takes on a variety of forms. In general, it is defined as sexual behavior between individuals of the same sex that is (usually more commonly) expressed between individuals of opposite sex. It includes behaviors such as courtship, mounting, and copulation. Studies of same-sex interactions in nonhuman animals typically focus on same-sex behavior or same-sex preferences, rather than same-sex orientation, which is a more enduring preference and therefore more difficult to ascertain. SSB has been recorded at least since the time of Aristotle:
And some male birds have been seen to be so effeminate from their birth, that they neither crowed, nor desired sexual intercourse, and would submit themselves to any males that desired them. (Aristotle, Historia Animalium, in Cresswell [1891].)
In some species, the proportion of individuals in a population that exhibit SSB can be high; for example, more than 30 percent of pair-bonded birds in a Hawaiian population of Laysan albatross (Phoebastria immutabilis) were found to be female-female pairs, and in a study of the deep-sea squid Octopoteuthis deletron, researchers observed that about half of all sexual encounters were between males. The manner in which SSB manifests is as varied as the number of species in which it is found. To name just several, male toads (Bufo bufo) sometimes amplect other males, female Japanese macaques (Macaca fuscata) mount one another and engage in genital contact, and male flour beetles (Tribolium castaneum) will mount and attempt to deposit sperm on other males.
The pervasive assumption that SSB is never adaptive has been gradually but steadily replaced by a wide range of hypotheses for evolutionary mechanisms that maintain the behavior, and in many respects the human and primate literature has led this transition. There is no universal reason for the existence of SSB, as it appears to have different functions and origins in different animal species. However, SSB provides a vehicle for studying evolutionary mechanisms underlying many apparently nonadaptive behaviors, for example, sexually antagonistic selection, kin selection, intrasexual competition, cooperative breeding, and others. A recent theoretical analysis published by Sergey Gavrilets and William Rice is illustrative. Gavrilets and Rice examined the feasibility of two major genetic models for the evolutionary maintenance of SSB: overdominance and sexual antagonism. Overdominance describes a heterozygote advantage; genes that confer a fitness advantage when in the heterozygous condition might induce the expression of SSB when in the homozygous condition. Given a large enough heterozygote advantage, SSB could be maintained in a population even if it decreases the fitness of individuals that exhibit it. Under sexual antagonism, genes that provide a fitness advantage in one sex might antagonistically cause the expression of SSB in the other. Gavrilets and Rice were able to derive genetic and phenotypic conditions under which either scenario could maintain SSB within a population. Empirical evidence from humans supports predictions made by the sexual antagonism model: female relatives of male homosexuals produce more offspring than female relatives of male heterosexuals, and male homosexuality tends to run in maternal lineages.
Additional hypotheses have been advanced to explain the evolutionary maintenance of SSB in other species. SSB in primates, cetaceans, and other mammals has been examined to test whether it provides social cohesion, acting as a form of currency in interactions among group members. The evidence is mixed. In some cases, such as in bottlenose dolphins, male-male SSB represents approximately half of all sexual encounters and appears to facilitate the formation of social alliances within small groups of males. In other systems such as Japanese macaques, the social functions of SSB remain unclear.
In contrast with mammal studies, research in insect systems has focused on ways in which SSB might mediate intrasexual competition. Sara Lewis and her colleagues studied male-male mounting and copulation in the flour beetle T. castaneum. Mounting males sometimes transfer sperm onto the males they have mounted. Lewis’s group hypothesized that when the mounted males later mate with a female, some of the previously deposited sperm could fertilize females’ eggs “by proxy,” thus providing an adaptive benefit for male-male mounting. However, while fertilization by proxy can happen, it appears unlikely to occur frequently enough to provide an adequate fitness benefit to explain its maintenance.
The evolutionary reasons for SSB remain unresolved for most species. However, as exemplified by the studies described, research that tests hypotheses for the existence of an apparently nonadaptive behavior demonstrates how it can be studied within an evolutionary framework. These studies provide insights not only into the dynamics of a particular behavior but also into more general evolutionary questions, such as how cooperation or competition manifest in different species, what evolutionary forces shape behavior, and why the genetic architecture of behavior is important.
7. INSIGHTS FROM APPARENTLY NONADAPTIVE BEHAVIOR
Behaviors that appear to be nonadaptive are common. In some respects, labeling a behavior as nonadaptive is a matter of perspective. Behaviors that seem to be detrimental to the fitness of an individual are often actually adaptive on closer inspection, such as in the case of redback spiders. Other behaviors that appear detrimental really are fitness reducing, representing new mutations that have yet to be eliminated from the population, as in the case of genetic mutations seen in lab studies. However, there is a vast gray area between these two extremes describing behaviors that are evolutionarily maintained in populations despite what we perceive to be heavy costs. These include sexual cannibalism, brood parasitism, parasite manipulation, and same-sex sexual behavior, all of which have been empirically and theoretically studied to try to understand the evolutionary mechanisms causing their persistence. Same-sex sexual behavior has been utilized to explore the evolutionary mechanisms that maintain many nonadaptive behaviors, such as sexually antagonistic selection, kin selection, and intrasexual competition. By employing a toolkit of evolutionary hypotheses, future research on SSB has the potential to illuminate surprising and intriguing reasons for its existence that inform other areas of evolutionary biology.
Andrade, M.C.B. 1996. Sexual selection for male sacrifice in the Australian redback spider. Science 271: 70–72. A study examining the fitness benefits to males of self-sacrificing courtship behavior.
Bailey, N. W., and M. Zuk. 2009. Same-sex sexual behavior and evolution. Trends in Ecology & Evolution 24: 439–446. The most recent general review of same-sex sexual behavior that examines its causes and consequences across a wide range of species.
Cresswell, R. 1891. Aristotle’s History of Animals. London: George Bell and Sons. A comprehensive translation of Aristotle’s observational writings on animal natural history, behavior, and morphology.
Dawkins, R. 1982. The Extended Phenotype. Oxford: Oxford University Press. One of Dawkins’s most influential works, this book outlines his observation that genes in one individual can influence the development of traits in other individuals, through “extended” effects.
Gavrilets, S., and W. R. Rice. 2006. Genetic models of homosexuality: Generating testable predictions. Proceedings of the Royal Society B 273: 3031–3038. One of very few formal theoretical models of same-sex sexual behavior, this study examined genetic conditions under which the behavior might be expected to persist in populations.
Klein, S. L. 2003. Parasite manipulation of the proximate mechanisms that mediate social behavior in vertebrates. Physiology and Behavior 79: 441–449. A comprehensive review article on the evidence for parasite manipulation of behavior, presented from a mechanistic perspective.
Parker, G. A., and J. M. Smith. 1990. Optimality theory in evolutionary biology. Nature 348: 27–33. A seminal exposition of the rationale behind optimization models, and their implications for the power of selection to craft adaptations.
Servedio, M. R., and R. Lande. 2003. Coevolution of an avian host and its parasitic cuckoo. Evolution 57: 1164–1175. A useful discussion of the arms race hypothesis in systems of brood parasitism, plus a contrasting model for the evolutionary dynamics of such systems.
Tigreros, N., A. South, T. Fedina, and S. Lewis. 2009. Does fertilization by proxy occur in Tribolium beetles? A replicated study of a novel mechanism of sperm transfer. Animal Behaviour 77: 555–557. The most recent published study taking a hypothesis-testing approach to assess why male-male copulation occurs in flour beetles.
