1. Three concepts of the ecological niche
2. The recess/role niche and seeking ecological equivalents
3. The population-persistence niche and mechanistically representing competition
4. The resource-utilization niche and understanding the evolution of species differences
5. Environmental niche modeling and analyzing niches on a macroscale
It may come as something of a surprise that ecological niche, a term so common in the popular media, has three distinct meanings among scientists, each with an associated conceptual basis: these are the recess/role niche, the population-persistence niche, and the resource-utilization niche.
character displacement. The situation in which two species are more different in geographic locations where they overlap than between locations where they occur alone
community. Those species populations occurring at some location
competition. Ecological interaction in which two or more species negatively affect one another by consuming common resources or by other harmful means
convergence. Development of increasing similarity over time, usually applied to species somewhat unrelated evolutionarily
niche dimension. Environmental variable along which a species’ niche is characterized, e.g., food size, and typically represented as the axis of a graph
polymorphism. The existence of two or more forms, differing in morphology or some other way, in the same population
population. Those individuals of a species occurring at some location
population growth rate r. The per capita rate at which a population changes size, typically computed as the birthrate minus the death rate
The first use of “ecological niche” appeared in a report on ladybugs written by R. H. Johnson nearly a century ago, although the term was used shortly thereafter by the zoologist Joseph Grinnell, who is generally given credit for its original development. The meaning was very close to figurative usage: the ecological niche of a species is its “role,” “place,” or more literally “recess” (in the sense of a “nook” or “cubbyhole”) in an ecological community. Thus, the California thrasher, one of Grinnell’s major examples, is a bird of the chaparral community that feeds mostly on the ground by working over the surface litter and eating both animal and plant items of a suitable size. Escape from predators is similarly terrestrial, with the well-camouflaged bird shuffling off through the underbrush on the rare occasions when it is threatened.
The idea that there exists a set of characteristic habitat and food types with accompanying behavioral, morphological, and physiological adaptations leads to the notion of ecological equivalents. These are defined as two or more species with very similar niche characteristics that occur in completely different localities. An example from Grinnell’s writings is the kangaroo rat of North America, which “corresponds exactly” to the jerboa (another desert rodent species) of the Sahara. The existence of ecological equivalents would imply that rather invariant rules determine the niches available for occupancy in a particular kind of environment, e.g., a desert. Moreover, niches can be empty in the sense that a suitable species does not occur within a locality, perhaps because it never got there or was unable to evolve in situ.
But to what extent do ecological equivalents really exist? Decades after Grinnell’s work, we now know (section 2, below) that although some examples can certainly be found, perhaps more commonly, species of similar environments (e.g., deserts) among distant localities are neither identical nor often even similar. Perhaps such considerations helped to engender the two other meanings of ecological niche, each with its accompanying set of ideas about how the ecological world works.
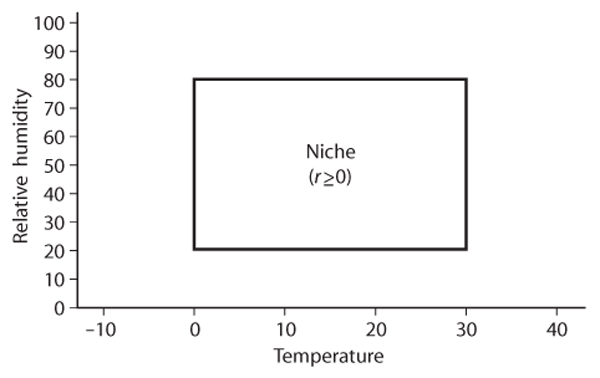
The population-persistence niche has its roots in papers written in the mid-twentieth century by the ecologist and limnologist G. E. Hutchinson. This concept focuses on the species, in this case its population, rather than on the environment. Hutchinson formulates the ecological niche as a quantitative description of the range of environmental conditions that allow a population to persist in some location; the term persist means having a positive or at least zero (break-even) population growth rate, r (if r is negative, the population dwindles away to extinction). An example of an environmental condition is temperature; a second example would be humidity (for organisms on land) or salinity (for organisms in water). If we represent an environmental condition by the axis of a graph, a range is an interval along that axis, e.g., temperature from 0°C to 30°C (figure 1). A second interval, say for relative humidity, might range from 20% to 80% along the humidity axis. We can have as many different environmental axes as necessary to characterize the population growth rate. If r for a given axis is uncorrelated with the values of variables of the other axis (e.g., if the range of temperatures allowing r ≥ 0 is the same for any value of humidity), then the niche is rectangular (as in figure 1); otherwise it will have other shapes. Hutchinson labeled his concept the cumbersome “n-dimensional hypervolume” (imagine three or more environmental axes). The more succinctly labeled fundamental niche is that portion of niche space where the species population can persist. The fundamental niche is visualized as being in the absence of other species that compete with the given species for resources and thereby affect its persistence. To account for this latter circumstance, Hutchinson defined the “realized niche” as that portion of the fundamental niche not overlapping the fundamental niches of competing species, plus that portion overlapping the competing species’ niches where the given species can still persist (have r ≥ 0).
Figure 1. Example of Hutchinson’s population-persistence niche. Rectangle encloses the ranges of temperature and humidity in which the species’ population can persist (where r ≥ 0).
Hutchinson’s concept is important for several reasons. First, it provides a precise, quantitative way to characterize the ecological niche. Second, it focuses on what the species itself does rather than on the opportunity for a species to exist or not in a community (the latter being the “recess” concept of Grinnell). Thus, ecological equivalents are not necessarily expected and, if they do not occur, are not troubling to the concept: for Hutchinson, there are no “empty niches.”
Such a precise formulation of the niche is not without its drawbacks, however. Chief among them perhaps is the difficulty of finding out what the population-persistence niche of a species actually is in nature. Presumably, for each point of the n-dimensional hypervolume—say for each value of temperature and humidity—one needs to culture populations or otherwise determine their population growth rate r; and one repeats this for different points until one has all combinations of temperature and humidity for which the population can persist. The difficulty of so doing for all but microorganisms (at best) is easy to imagine. A second problem is that certain niche characteristics as conceptualized by Grinnell are not easily ordered along an environmental axis. An example is food size: at any given real location, food comes in a variety of sizes (rather than there being one food size for each location). Of course, one can use average food size, but such a concept is not as plausible as using average temperature because animals come across a variety of food sizes on a daily basis. Animals of a particular body size (and therefore a particular size of feeding apparatus, e.g., mouth) have limitations on the extreme values of food size that can be consumed: items too large cannot be swallowed, and those too small cannot be handled deftly (or eaten in an energetically profitable way). Hence, a more detailed description than the average food size available at a location is desirable. Third, Hutchinson’s niche is onesided in the sense that it assumes a rather passive species that does not affect other species in the community in a way that eventually feeds back onto the given species. Fourth, Hutchinson focuses almost exclusively on one type of ecological interaction, competition between species; for example, his distinction between the fundamental and the realized niche. In this way, his concept was not as inclusive as that of Grinnell.
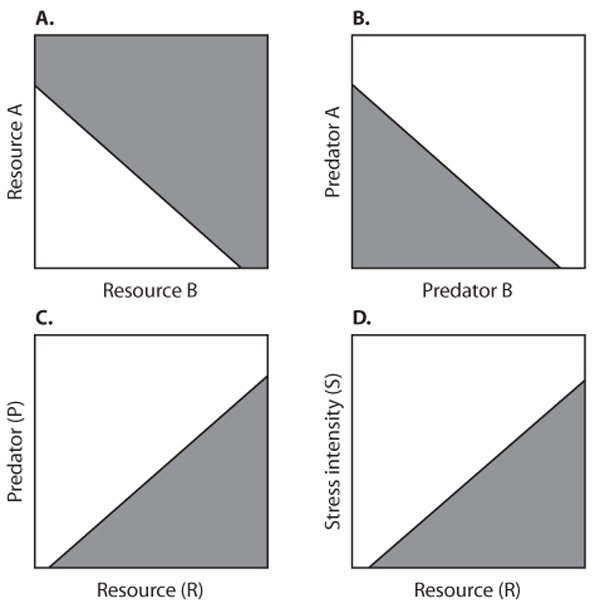
Figure 2. Chase-Leibold concept of the population-persistence niche. Each panel has two regions, a shaded region where r ≥ 0 and an unshaded region where r < 0. The niche is the shaded region. (A) A species with two substitutable resources (the axes measure resource density); (B) a species with two predators; (C) a species with a predator and a resource; (D) a species with a stress and a resource. (Figure courtesy of J. M. Chase.)
In part as a reaction to the latter two drawbacks, Jonathan Chase and Mathew Leibold have substantially extended the population-persistence niche. In a recent but already very influential book, they define the niche as a joint specification of environmental conditions or variables that allow a species to have r ≥ 0 along with the effects of that species on those environmental variables. Niche axes are quite broadly construed and can include a variety of factors that impact populations (and vice versa); examples include amount of a given resource, abundance of a given predator, and degree of a physical stress such as wind speed (figure 2). Thus, one can incorporate effects of species on environmental conditions, and one can specify a given region of niche space where a species has r ≥ 0 (figure 2). Although this model represents a vast improvement in the concept of population-persistence niche, the operational difficulty of measurement still exists: determining the niche for figure 2 (Chase and Leibold) is not much easier than for figure 1 (Hutchinson).
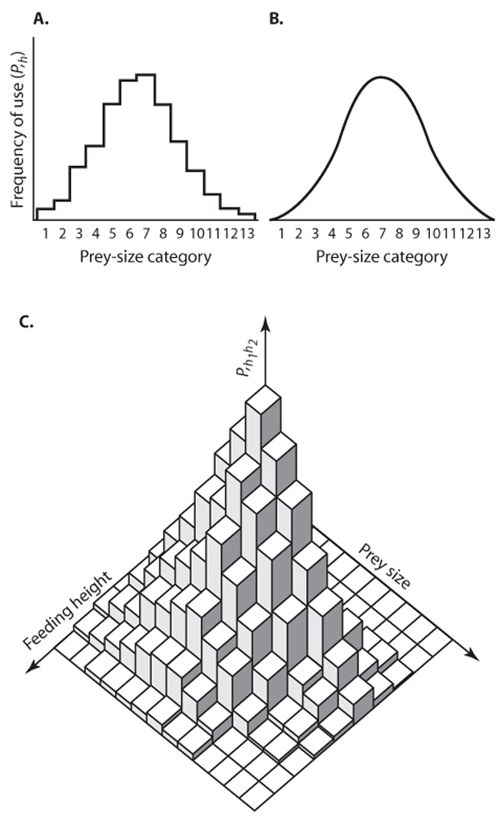
An eminently operational concept of the ecological niche, formulated by two evolutionary ecologists, Robert MacArthur and Richard Levins, is the resource-utilization niche, our third meaning. Like the population-persistence niche, the resource-utilization niche is quantitative and multidimensional, but it focuses entirely on what members of a species population in some locality actually do—in particular, how they use resources. The relative use (= utilization) of resources along a given niche axis can be described as a frequency distribution or histogram. Take, for example, the axis food size. We can (figure 3, top) draw a histogram showing the fraction of food of different sizes consumed by all members combined of a given population; e.g., the fraction of the total population’s foods between 5 and 6 mm. If we have a second dimension, say feeding height, we can graph the fraction of food items eaten at different heights in the vegetation. The two can be combined as a joint distribution or three-dimensional histogram (figure 3, bottom), and this can be further generalized (although not easily graphed) for as many dimensions as ecologists find important to describe the population’s resource use. A broad classification of the kinds of niche axes used for utilizations consists of habitat, food type, and time. Within habitat, microhabitat and macrohabitat are distinguished, whereby microhabitat has a smaller spatial scale (e.g., height in vegetation) than does macrohabitat (e.g., vegetation zone such as tropical rainforest or desert). Within food type, food size and hardness can be distinguished. Within time, daily and seasonal activity can be distinguished.
The resource-utilization niche immediately frees us from the problem with Hutchinson’s formulation that certain environmental variables cannot be meaningfully described using only the average. Indeed, the resource-utilization niche is nothing more than a precisely formulated description of the natural history of a species: its habitat, food types, and activity times, among other things. Such natural history can include nonfeeding habitats and activity times for behaviors such as predator escape and mating, all characterizable on its niche axes. Thus, we have a niche concept that precisely encapsulates what ecologists measure anyway. Indeed, Grinnell, the originator of the recess/role niche concept, measured such things in his study organisms but with the assumption that, in so doing, he was discovering something about the availability of niches in the community—an availability or opportunity to which the species more or less had to conform. The resource-utilization niche, in contrast, assumes nothing about rigidly determined niche recesses in a community, nor about the necessity of ecological equivalents, nor about the existence of empty niches. The resource-utilization niche was formulated a decade or so after the population-persistence niche but, unlike the latter, has remained rather unchanged up to the present. This is despite the fact that, by emphasizing resources, it is seldom extended beyond resource use, unlike both the recess/role niche of Grinnell and the population-persistence niche of Chase and Leibold.
Figure 3. An example of the resource-utilization niche. (A) A one-dimensional niche, where the dimension is prey size. Numbers give prey-size categories, indexed by h; (B) the same utilization smoothed; (C) utilization of two resource dimensions, preysize and feeding height. (Redrawn from Schoener, 1986.)
We now review seriatim the kinds of research engendered by the three concepts of the niche as well as a very recent research trend called ecological niche modeling that includes elements of all three.
In an early study of grassland birds inhabiting far-flung locations—Kansas, Chile, and California—Cody found that each community contained about the same number of species and the same ecological types: three or four passerines (small “perching” birds), a larger vegetarian “grouse-like” species, both a long- and a short-billed wader, and two or three raptors. Twenty pairs of ecological equivalents were identified between the two Mediterranean systems: Chile and California. However, later studies by Cody in other Mediterranean systems including Sardinia and South Africa showed a weaker pattern, especially for the latter, whose floras were very different.
In contrast to birds, plants in Chilean and Californian systems showed little convergence at the community level; for example, woody vegetation in Chile comprises less of the total cover but more total species and has a greater diversity of height layers than in California. Nonetheless, the major growth forms (e.g., broad-leaved evergreen, broad-leaved deciduous) are similar, even with regard to number of species, although several forms present in Chile (e.g., spinose-stemmed shrubs) are absent from California—an apparent empty niche. Major resemblances between plant growth forms among plants with very different evolutionary lineages occur rather commonly among plants; a striking example is given by American cacti and African euphorbs.
Perhaps the least evidence for ecological equivalents after systematic search is among colubrid snakes of North, Central, and South America. Cadle and Greene find few ecological equivalents (and little evidence for community similarity); instead, a number of types (fossorial earthworm eaters, nocturnal arboreal lizard/frog eaters) in some communities are conspicuously absent in others.
Probably the most extensive work on convergence and ecological equivalents has been done on lizards. An initial study by Fuentes, again comparing Chile and California, found convergences in community characteristics as well as in individual niche traits—microhabitat, daily activity time, and food type. In a second major study, Pianka found less evidence for similarity in community characteristics than difference among lizards of the three warm-desert systems of North America, Australia, and Africa. Nonetheless, striking ecological equivalents sometimes exist, such as the amazing resemblance between the horned toad of North America and the thorny devil of Australia (figure 4).
Figure 4. An example of ecological equivalents: the horned toad (Phrynosoma platyrhinos) of North American deserts and the thorny devil (Moloch horridus) of Australian deserts. (From Pianka, E. R. 2000. Evolutionary Ecology. San Francisco: Harper & Row. Used by permission of Pearson Education, Inc.)
Examples of ecological equivalents are most impressive when the species from widely different localities are relatively unrelated in terms of evolutionary descent: convergent evolution toward the same morphology and behavior would seem to support the idea of the niche as a functional optimum characteristic of particular types of communities (e.g., those in deserts) into which species repeatedly evolve. Nonetheless, a plausible hypothesis for lack of convergence is that major evolutionary stocks are so different that evolution is too constrained to produce much convergence. Melville, Harmon, and Losos recently examined two lizard families, the Iguanidae and Agamidae, of North America and Australia, respectively, which are closely enough related to belong to the same clade (Iguania) even though they have been geographically separate for as long as 150 million years. Using an approach that takes into account evolutionary relatedness, they found convergence in habitat use and locomotor morphology, including pairs of ecological equivalents, between the two deserts.
Another example of convergence among relatively closely related species is provided by the Anolis lizards of large West Indian islands: Cuba, Hispaniola, Jamaica, and Puerto Rico. Here, various ecomorphs—species occupying the same microhabitat—have independently evolved on the separate islands. Harmon, Kolbe, Cheverud, and Losos found that five functionally distinct morphological characters—body size, body shape, head shape, lamella (ridges on toes) number, and sexual size dimorphism—converge among the different islands as a function of habitat similarity. For example, lizards living on the ground and low trunks are more similar between Cuba and Hispaniola than either is to other ecomorphs (e.g., those living in tree crowns) co-occurring on the same island and to which they are more closely related.
A final recently discovered example of convergence occurs in a completely different group: orb-weaving spiders of the genus Tetragnatha of the Hawaiian islands. Blackledge and Gillespie found that spiders inhabiting different islands constructed remarkably similar webs. These convergences toward ecological equivalency, which they called “ethotypes” (ethology is the study of behavior, and this emphasizes the behavioral similarity), occurred independently in evolution. Like the Australian Iguania discussed above, the group as a whole consists of relatively closely related species.
In conclusion, although the evidence for ecological equivalents is certainly mixed, more and more examples are coming to light that make Grinnell’s rather old concept seem alive if not completely well. As Schluter has suggested, to the extent that ecological equivalents exist and are independently evolved, morphology, physiology, and behavior must constrain the efficiencies with which resources and other factors characteristic of particular kinds of ecosystems (e.g., deserts) can be dealt with—ecological equivalents mark peaks in the adaptive landscape.
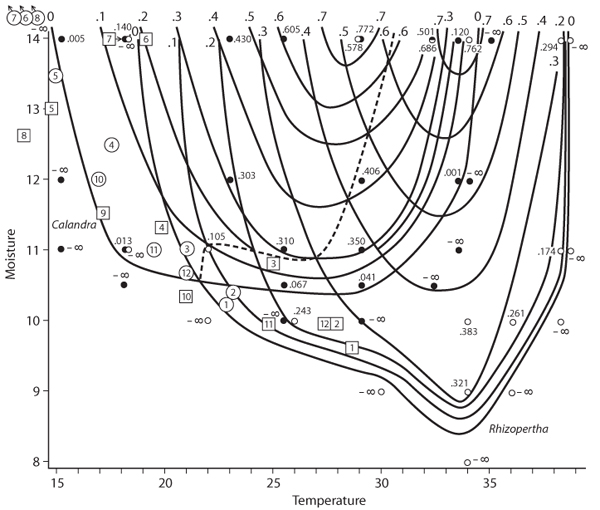
Maguire in 1973 may have been first to plot population growth rate r for real species as a function of niche dimensions and to make predictions about the competitive outcomes among them. In the l950s, Birch had studied several species of beetle infesting stored grain in Australia; figure 5 shows Maguire’s plot of Birch’s data with respect to temperature and moisture. Isoclines of positive values of r down to zero (no population growth) show different patterns for the two species, such that Calandra oryzae has a higher r for lower temperatures and somewhat greater moistures than Rhizopertha dominica. The dashed line in figure 5 separates regions of niche space where one versus the other species has the higher r. Assuming no complications, an environment on one or the other side of the line will favor one or the other species of beetle in competition.
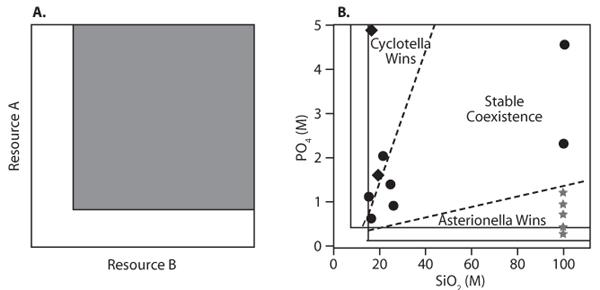
To illustrate their ideas about the population-persistence niche, Chase and Leibold replot data of Tilman for two species of diatoms, Asterionella and Cyclotella (figure 6). The situation is somewhat more complex than that shown in figure 2 because resources are not substitutable (which would mean that the populations can survive on either resource alone or on some combination) but rather are essential: figure 6A shows the general case, where a species must have a minimal amount of each resource in order that r ≥ 0. For two such species, coexistence is possible if each species can just survive (r = 0) for a different one of the two resources. In Tilman’s experiment, the resources are the nutrients silicate (SiO2) and phosphate (PO4), and the levels of each can be controlled in the laboratory. Asterionella is a specialist on SiO2, and Cyclotella on PO4. From the individual species growth curves on the separate resources, one can predict regions of niche space (plots of SiO2 versus PO4 concentration) where each species has a lower r = 0 and so is limited by a different resource. In that region (figure 6B), the species can coexist. Outside that region, one or the other species wins, depending on which resource is more abundant.
Figure 5. Maguire’s illustration of the population-persistence niche for the dimensions temperature (C) and percentage moisture of wheat, using the beetles Calandra oryzae (small strain) and Rhizopertha dominica. Numbers in squares indicate average monthly conditions in Bourke, New SouthWales, Australia; numbers in circles give same for Adelaide, South Australia. (Redrawn from Maguire, B., Jr. 1973. Niche response structure and the analytical potentials of its relationship to the habitat. American Naturalist 107: 213–246)
Such empirical studies are impressively successful in the highly controlled setting of the laboratory, but they are very difficult indeed to perform in the field. Chase and Liebold could find only one such field study, again by Tilman (and Wedin), in which several plant species vary in their ability to utilize nitrogen from the soil. These relative abilities were used rather successfully to predict competitive outcomes along a natural nitrogen gradient. Probably, practical difficulties largely explain why the population-persistence niche is a concept with mostly theoretical development. It seems most likely that it will be easiest to apply to organisms with the size and behavior that enable their populations to persist in small spatial units (sometimes called microcosms).
How similar can species be and still coexist? An answer was obtained in the last section for species having a small number of ecological requirements or resource types. What if species fed on a wide variety of resources, such as foods of different sizes found at different vegetation heights and preferring different temperatures? This situation applies to predators, such as Grinnell’s California thrasher, that eat a great variety of insects and other arthropods that in turn have their own populations with their own niche characteristics.
Figure 6. (A) Chase-Leibold population-persistence niche (shaded area) for essential resources. (B) Ranges of coexistence and exclusion for two species of diatoms competing for two essential resources in chemostats. Circles give nutrient-ratio treatments where Asterionella excludes Cyclotella; diamonds give treatments where Cyclotella excludes Asterionella. (Redrawn from Chase and Leibold, 2003; their figure 4.1 in turn redrawn from Tilman, D. 1977. Resource competition between planktonic algae: An experimental and theoretical approach. Ecology 58: 338–348)
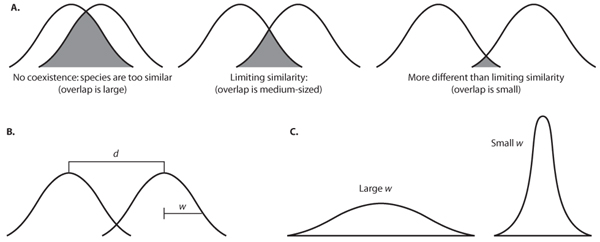
The 1967 paper in which MacArthur and Levins promoted the resource-utilization niche has as its main objective the understanding of how similar competing species can be and yet still coexist. It is sometimes said that species cannot coexist if they occupy the same niche, but the theory of MacArthur and Levins also posits that if the niches of the species are too similar (too much niche overlap), they still cannot coexist. To illustrate, imagine two species with the one-dimensional niche in figure 7; this dimension might be food size, and one species tends to eat larger food on average than the other. If the niches are too close (figure 7A, left), they are too similar (the niche overlap [shaded area] is too great), and the better competitor will eliminate the other from the community. That degree of closeness at which the species can just coexist (any closer and one is eliminated) is called the limiting similarity (figure 7A, middle); the niches can, of course, be farther apart and still allow coexistence (figure 7A, right).
Limiting similarity is measured in units of d/w, where d is the distance between peaks and w is the width of the niche (usually computed as the standard deviation of the utilization distribution; figure 7B). The larger the w, the more generalized the species; a specialist has a thin niche (small w; figure 7C).
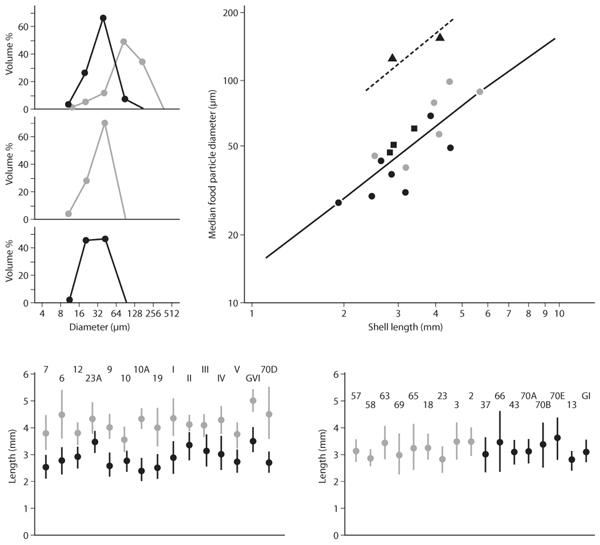
In MacArthur and Levins’s theory, a d/w slightly larger than 1.0 is the limiting similarity; much subsequent work has shown limiting similarity to vary greatly yet be about 1 (certainly to an order of magnitude). Indeed, sometimes real species differ by almost exactly this theoretical value. A sensational example is provided by two mud snails (Hydrobia) studied by Fenchel in Denmark. The snails ingest particles: diatoms and inorganic pebbles covered with minute sessile organisms. About 150 years before the study, a fjord collapsed, and one species invaded the other’s range. The resource-utilization niches of the species displaced away from one another, apparently independently, numerous times, to d/w ≈ 1 (figure 8, top left). Corresponding to this niche difference is a difference in body (shell) size such that larger species ingest larger particles (figure 8, top right), and the body sizes of the species had diverged (in a process called character displacement) to a ratio of 1.3–1.5 (figure 8, bottom). Consistent with the theory of Taper and Case (see below), this ratio is higher than the ratio of d’s for the two resource utilizations of 1.2.
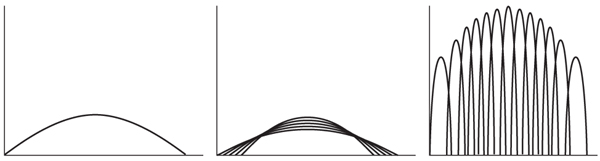
So far we have represented the resource-utilization niche as a distribution summing together the food-size or other niche characteristics of all individuals in a population. However, individuals may differ in their niche characteristics, sometimes just by chance opportunity (e.g., what they happen to come across to eat), but sometimes because they have different morphologies and behaviors that make them specialized for a certain portion of a niche axis (just as species can be specialized). Figure 9 shows the two extreme possibilities for such component individuals; note that each individual can be a generalist (figure 9, left) or a specialist (figure 9, middle), in either case producing the same utilization for all individuals combined (figure 9, right). What difference does it make which of the two situations one has? A series of specialist individuals may eventually allow the population as a whole to be more generalized in the absence of competing species, and this “polymorphism” might even lead to speciation (see chapter I.18). Such polymorphism, when measured in terms of those morphological characters corresponding to position on the niche axis (e.g., shell size corresponding to mean food-particle size), was uncommon in the literature at the time of Taper and Case’s paper, and this was consistent with their theoretical model in which the proportion of different kinds of individual niches evolves once the competing species meet geographically. Recently, however, Bolnick, Svänback, Arágo, and Persson looked at the resource-utilization niches themselves rather than the morphological characters that reflect them. They found that the bigger the w for the total population, the bigger the between-phenotype niche width, measured as the standard deviation of the d’s of the niches of the component individuals. It remains to be seen exactly how these apparently somewhat contradictory trends will be reconciled.
Figure 7. (A) One-dimensional resource-utilization niches of two species showing no coexistence because niches are too similar (left), limiting similarity—species can just coexist (middle), and greater than limiting similarityalso allowing coexistence (right). (B) Niche distance d and niche width w for two species. (C) Niches of generalist (large w) and specialist (small w) species.
A recent set of techniques, called environmental niche modeling (ENM), combines elements of all three niche concepts. The method characterizes the macrohabitat niche of a species by quantitatively summarizing geographic-information-system (GIS) information on climatic and similar variables at stations throughout the species’ geographic range. Such macrohabitat niche information is then used to predict the potential geographic range of the subject species. Because of its focus on macrohabitat, the scale is similar to Hutchinson’s version of the population-persistence niche. However, the method specifies the “empty niches” of Grinnell’s recess/role niche as those localities having the niche characteristics of the subject species but where that species does not, in fact, occur. Finally, it allows quantification of niche similarity between species via measures of niche overlap used for the typically finer scale of resource-utilization niches of MacArthur and Levins.
One of the most successful applications of ENM so far examines the question of whether the more closely related species are, the more similar are their niches. The question is important because if the answer is yes, evolutionary history must have a major influence in determining niche characteristics relative to the influence of the community in which the species now occurs. A study by Knouft, Losos, Glor, and Kolbe on the 11 species of the Anolis sagrei group in Cuba found no evidence that niches were more similar, the more closely related the species (evolutionary relatedness is assessed using molecular genetics). A second study, by Warren, Glor, and Turelli showed along with the previous study that the most recently diverged species had the greatest climatic-niche differences. The second study, however, gave somewhat more support for the hypothesis in general, in that niche similarity between closely related species of birds, butterflies, and mammals separated by the Isthmus of Panama was greater than expected by chance. However, somewhat contrary to the founding ENM study by Peterson, Soberon, and Sachez, niches were rarely identical, so the overall answer is in fact mixed, as is so often the case in ecology.
Figure 8. (top left) Resource-utilization niches for prey size among two species of gastropods (Hydrobia ulvae, gray circles; H. ventrosa, black circles) where the species overlap (top), where H. ulvae is alone (middle), and where H. ventrosa is alone (bottom). (top right) Median diameters of ingested food particles of four species of Hydrobia plotted against shell length. (bottom) Average lengths of Hydrobia ulvae (gray circles) and H. ventrosa (black circles) from 15 localities where the species co-occur (left) and 17 localities where one of the two species occurs alone. All samples from the Limfjord during summer 1974. (Redrawn from Fenchel, T. 1975. Character displacement and coexistence in mud snails (Hydrobiidae). Oecologia 20: 19–32)
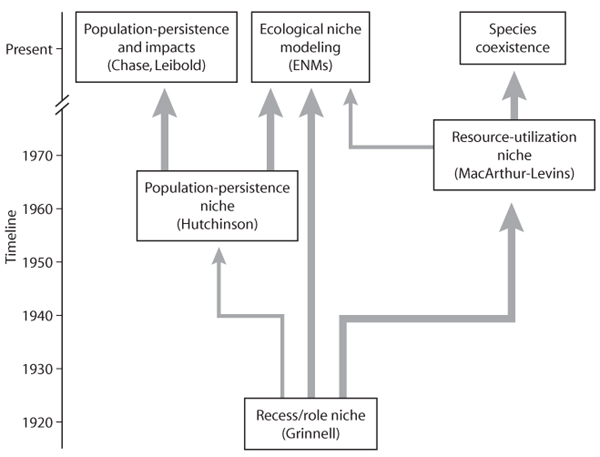
The research trends discussed in relation to the three niche concepts are summarized as an evolutionary tree in figure 10. In this diagram, the thicker arrows indicate a greater influence of one concept or research program on the next. Note that all three niche concepts, despite sometimes rather early beginnings, have stimulated research that is being actively pursued at the present time.
Figure 9. (left) Resource-utilization niche of a species population. (middle) Population of generalist individuals whose niches sum to the curve at top. (right) Population of specialist individuals whose niches sum to the curve at top. (After Klopfer, Peter H. 1962. Behavioral Aspects of Ecology. Princeton, NJ: Princeton University Press)
Figure 10. Timeline showing the development of niche concepts and the research programs stemming from them.
Chase, Jonathan M., and Mathew A. Leibold. 2003. Ecological Niches. Chicago and London: University of Chicago Press. A recent major revision of the population-persistence niche concept.
Grinnell, Joseph. 1917. The niche-relationships of the California thrasher. Auk 34: 427–433. One of the founding papers of the recess/role niche concept.
Harmon, Luke J., Jason J. Kolbe, James M. Cheverud, and Jonathan B. Losos. 2005. Convergence and the multidimensional niche. Evolution 59: 409–421. A very recent study of convergence and ecological equivalents that emphasizes the niche.
Hutchinson, G. Evelyn. 1957. Concluding remarks. Cold Spring Harbor Symposia on Quantitative Biology 22: 415–427. The founding paper of the population-persistence niche concept.
Knouft, Jason H., Jonathan B. Losos, Richard E. Glor, and Jason J. Kolbe. 2006. Phylogenetic analysis of the evolution of the niche in lizards of the Anolis sagrei group. Ecology 87: S29–S38. A recent exemplar of the environmental-niche-modeling technique.
MacArthur, Robert H., and Richard Levins. 1967. The limiting similarity, convergence and divergence of coexisting species. American Naturalist 101: 377–385. The founding paper of the resource-utilization niche concept.
Mooney, Harold A., ed. 1977. Convergent Evolution and Chile and California. Stroudsburg, PA: Dowden, Hutchinson and Ross. Contains major, detailed papers comparing two climatically similar regions.
Schoener, Thomas W. 1986. Resource partitioning. In J. Kikkawa and D. J. Anderson, eds. Community Ecology—Pattern and Process. Oxford: Blackwell Scientific Publications, 91–126. A review of how the resource-utilization niche is used in ecological research.
Schoener, Thomas W. 1989. The ecological niche. In J. M. Cherrett, ed. Ecological Concepts: The Contribution of Ecology to an Understanding of the Natural World. London: Blackwell Scientific Publications, 79–113. A detailed history of the development of niche concepts.