The “Order Insectivora” proposed by Simpson (1945) is considered a catchall term that included several morphologically disparate and unrelated groups of mammals and is not considered consistent or unified phylogenetically (Gunnell and Bloch, 2008). The term now commonly used is “insectivorous mammals,” which contains several different orders, each listed in this section (Gunnell and Bloch, 2008), and which more accurately reflects similarities in diet and ecology rather than being indicative of a phylogenetic relationship. Insectivorous mammals are characterized by a relatively small body size, elongate snouts, small eyes, relatively small brains, five-toed and plantigrade feet, and external ears that are small or absent. Their dentition was specialized for eating invertebrate prey, and they have rudimentary endothermy (Eisenberg, 1980; Symonds, 2005). As an ecological category, they also have a relatively long history in the geologic record (Symonds, 2005). The rocks at Badlands National Park preserve a diverse record of insectivorous mammals, although the fossils are fairly rare.
The order was set up to include extinct insectivorous mammals that had no clear relationship with living insectivorans. Little postcranial material has been found. More fossils need to be described to better document this group. Proteutherians tend to be characterized by a larger body size (Gunnell and Bloch, 2008).
The apatemyids are characterized by the lack of an ossified auditory bulla on the skull. The first lower incisor is enlarged and procumbent with a root extending posteriorly to the m3. The I1 lacks lateral enamel, and the canines are absent. The p3 is bladelike and single rooted. The upper molars are simple and lack mesostyles. Members of the family Apatemyidae range from the late Paleocene to the late Oligocene in North America (Gunnell et al., 2008).
Systematics and Evolution The type species for Sinclairella is S. dakotensis Jepsen, 1934. The specimen was collected from the upper part of Chadron Formation (about 11 feet below the limestone zone, which is usually considered as the boundary between the Chadron and Brule formations), near the head of the west fork of the east branch of Big Corral Draw, Shannon County, about 13 miles south-southwest of Scenic, South Dakota. The holotype and most complete specimen of Sinclairella have been lost for more than 30 years.
Distinctive Characters Sinclairella is characterized by its fairly large size compared to other insectivorous mammals. The upper molars are squared and have a large hypocone. The p4 is tiny, and the lower molars have reduced paraconids. The m3 is elongate with an expanded hypoconulid. The lower molar row is about four times as long as the lower premolar row (Gunnell et al., 2008).
Stratigraphic and Geographic Distribution Fossilized remains from Sinclairella can be found in the Chadronian through the Arikareean. Fossils are found in Wyoming, Colorado, South Dakota, Nebraska, Saskatchewan, and North Dakota.
Natural History and Paleoecology Sinclairella makes up a small part of the North American mammalian faunal sample until the end of the early Arikareean in the Great Plains. It is also the last surviving proteutherian in North America (Gunnell et al., 2008).
Two pantolestids, Chadronia margaretae Cook, 1954, and Cymaprimadon kenni Clark, 1968, were collected from north of Crawford, Nebraska, and Custer County, South Dakota, respectively. Both are from the White River Group but are beyond the area covered by this book. A new occurrence of Chadronia sp. has been documented in the Scenic Member of the Brule Formation in Badlands National Park, which extends the temporal range of pantolestids in North America (Boyd et al., 2014).
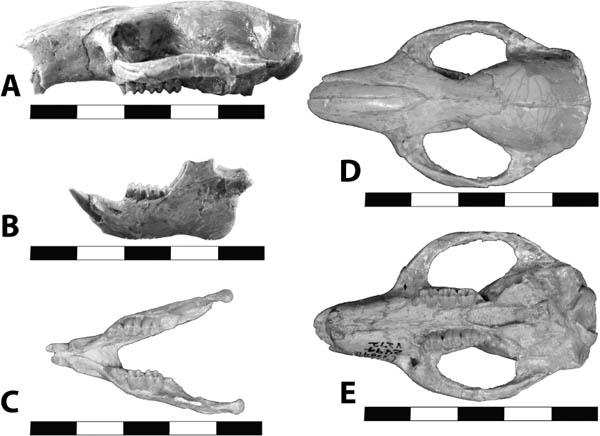
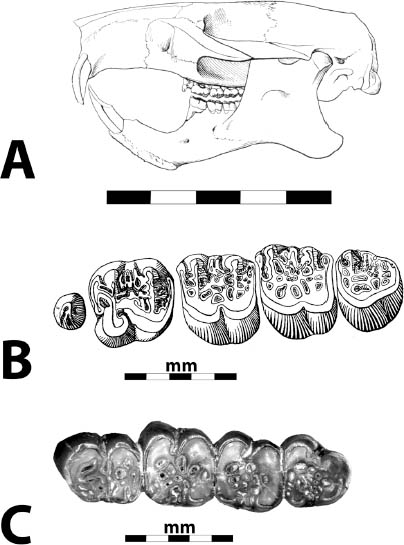
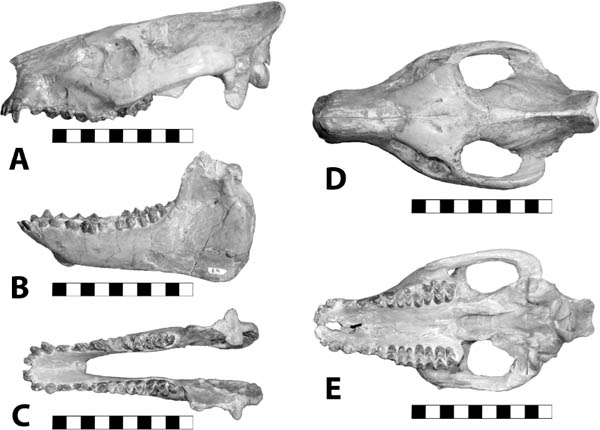
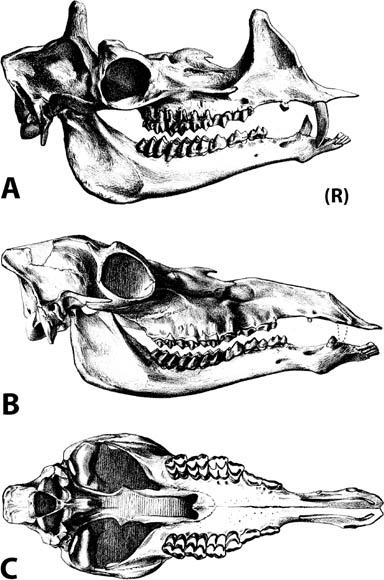
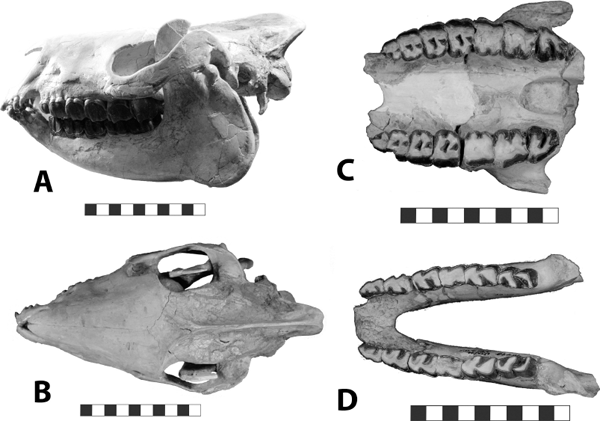
5.11. Insectivorous mammals from the White River Badlands (A) Leptictis sp., partial skull with right P2–M3 and left P1–M3, occlusal view, BADL 23756. (B) Proterix bicuspis, right P3–M3, BADL 11130/SDSM 7102 (C) Proscalops sp., skull, right lateral view, BADL 59754. (D) Proscalops sp., skull close-up showing antemolar region, right lateral view, BADL 59754. Scale bars for (A) and (B) are in centimeters; scale bars for (C) and (D) = 2 mm. Photo A by the authors; photo B by Edward Welsh; photos C and D by Mindy Householder. BADL 11130/SDSM 7102 from Badlands National Park, Interior, South Dakota, U.S.A., and the Museum of Geology, South Dakota School of Mines and Technology, Rapid City, South Dakota, U.S.A. BADL 59754 and BADL 23756 are from Badlands National Park, Interior, South Dakota, U.S.A. They are the property of the U.S. government.
The Leptictida are believed to be ancestral or related to several other groups. These include the Lipotyphla, the Apatemyidae, primates, and the Erinaceomorpha (Gunnell, Bown, and Bloch, 2008). They first appeared in the late Cretaceous of North America and range from the early Paleocene through the late Oligocene. Members of this order were either insectivorous or omnivorous. They had an elongate and mobile snout (Novacek, 1986). The forelimbs were much shorter than the hind limbs, and they had a distally fused tibia–fibula. The humerus was robust and was designed for digging for either food or shelter (Gunnell, Bown, and Bloch, 2008). The Leptictida were relatively small mammals, with an estimated body mass ranging from 400 to 700 g. They were also terrestrial mammals capable of rapid running and quadrupedal jumping (Rose, 1999).
The family is characterized by no more than five upper and lower premolars in total, often separated by a diastema. On the p5, the paraconid is large and the metaconid is reduced. The upper molars have district hypocones (Clemens, 1973; Novacek, 1986).
Systematics and Evolution The type species of Leptictis is L. haydeni Leidy, 1868. No type specimen was designated. Leptictis is the last of the order Leptictida in North America.
Distinctive Characters The skull is characterized by a twinned sagittal crest. The upper molars are transverse with narrow stylar shelves and large hypocones. The p4–p5 are similar in size. The P4 has a complex lingual moiety (Gunnell, Bown, and Bloch, 2008) (Fig. 5.11A).
Stratigraphic and Geographic Distribution Fossilized remains from Leptictis can be found in the Uintan through the early Arikareean. Fossils are found in South Dakota, Nebraska, Saskatchewan, Montana, Wyoming, Texas, Colorado, and North Dakota. In the White River Group of South Dakota, Leptictis is common in the Scenic Member but appears to have been rare in the Poleslide Member of the Brule Formation.
Natural History and Paleoecology Leptictis dakotensis likely was a quadrupedal cursor with possibly some bipedal hopping. It had relatively short forelimbs that were adapted for digging, a common behavior in terrestrial insectivores. Leptictis dakotensis has been recovered from burrowlike structures (Sundell, 1997; Rose, 1999, 2006). Certain structures in the nasals suggest that Leptictis had increased snout mobility similar to modern moles (Meehan and Martin, 2012).
The Lipotyphla include the moles, hedgehogs, and shrews. Modern forms are pentadactyl, plantigrade, and quadrupedal. The forelimbs are shorter than the hind limbs. They are characterized by terrestrial, fossorial, or semiaquatic locomotor patterns. The skulls were low with laterally facing small orbits. They had a small braincase and a long snout. The skulls have elongate nasals, indicating that the animals relied on olfaction for locating insect prey (Gunnell et al., 2008). Most of the fossils from this group are only known from jaws and teeth. They had dental specializations for eating soft-and hard-bodied insects. The postcrania are not well known.
The family Erinaceidae includes the modern hedgehogs, which today are found in Europe, Asia, and Africa but which in the past extended into North America. Modern hedgehogs belong to a separate subfamily called Erinaceinae, which originated in Asia and Europe approximately 35 Ma and continues to thrive today (Rich and Rasmussen, 1973). They were believed to be part of a faunal interchange between the Old World and North America (Rich and Rasmussen, 1973).
The dental formula for this family is I2–3/3, C1/1, P3–4/2–4, M3/3. The incisors are often enlarged, and the lower molars are progressively reduced from the m1 to m3. The lower molars are also rectangular with small or absent hypoconulids and weak exodaenodonty. The m1 has a strong, anteriorly angled paraconid. The upper molars are quadrate and often bunodont. The M1–2 have well-developed hypocones, and the M3 is reduced in size and oval in outline. The zygomatic arch is complete on skulls from members of this family (Gunnell et al., 2008).
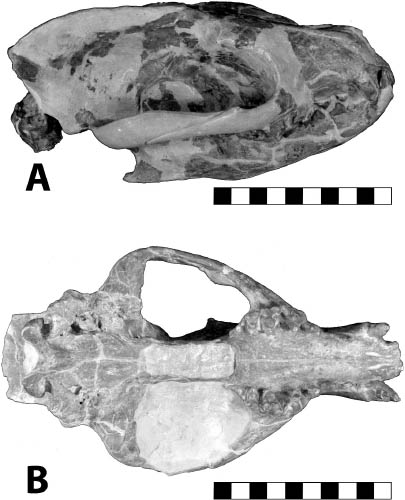
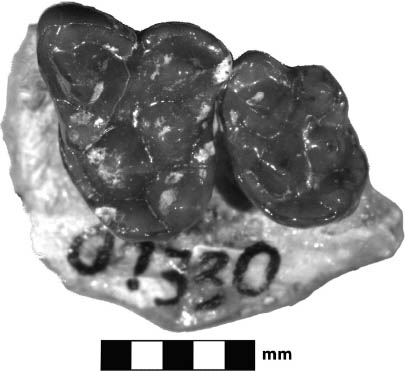
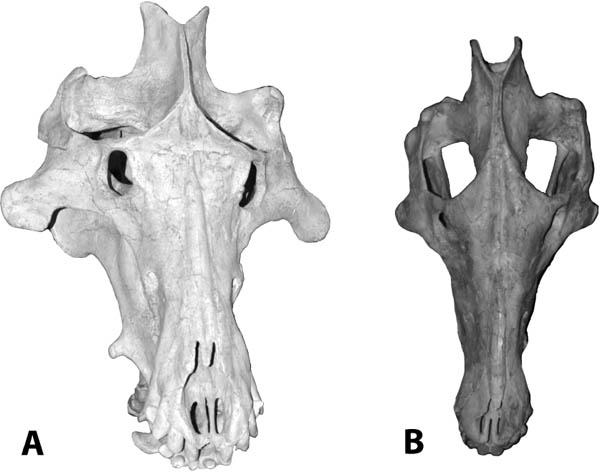
Systematics and Evolution The type species for Proterix is P. loomisi Matthew, 1903. The type specimen for Proterix is AMNH 9756, collected from Indian Draw, Cheyenne River, South Dakota, in the upper Oreodon beds (Lower Poleslide Member, Brule Formation) according to Gawne (1968). Proterix bicuspis (Macdonald, 1951), originally Apternodus bicuspis, SDSM 4048, was transferred to Proterix by Gawne (1968:11); it came from the Protoceras channel sandstone, 7 miles east of Rockyford, Shannon County, South Dakota.
Distinctive Characters The anterior portion of the skull is short with a completely ossified palate. On top of the skull, there are well-developed supraorbital crests and postorbital processes. The temporal crests form a strong sagittal crest. The nasals are elongate and extend posteriorly to the postorbital process. There is a deep depression anterior to the orbits for snout musculature. The zygomatic arches are well developed. The auditory bullae are completely ossified and extremely large. The P3–M3 are three rooted. The M3 is small and triangular. The M1 is quadrate and large with a subequal paracone and metacone. The m1 is five cusped, and the m2 and m3 are four cusped with a relatively broad cingulum (Rich, 1981) (Fig. 5.11B).
Stratigraphic and Geographic Distribution Fossilized remains from Proterix can be found in the Whitneyan through the early Arikareean. Fossils are found in the Poleslide Member of the Brule Formation in the White River Badlands of South Dakota. They are also found in Nebraska.
Natural History and Paleoecology Erinaceids are present throughout the Eocene and most of the Oligocene at a rather low diversity, but the group does continue into the Miocene. Beginning in the late early Arikareean, they begin to diversify and continue through most of the Miocene (Gunnell et al., 2008).
The family Proscalopidae, including ancient moles, are the earliest known Lipotyphla to exhibit highly specialized burrowing adaptations. Unlike modern moles, which use their forearms to break the soil during burrowing, in the Proscalopidae the head is used as the primary means of burrowing. This is indicated by the fusion of the cranial bones in adults, and the rostrum is distinctly longer than the cranium. The cranium is broad and deep with occipital condyles placed ventrally and well below the plane of the dentaries. The premaxillae are marked with prominent lateral shelves that extend anteriorly to the nares. The tympanic bulla is ossified but only slightly inflated (Gunnell et al., 2008).
The anteriormost incisors are enlarged and piercing in most genera. There are seven or fewer antemolars with at least some premolars that are single rooted. The lower molar cingulids are incomplete labially, and the molars are brachydont to hypsodont in latest forms.
The postcranial skeleton in fossil forms is also well adapted for burrowing. The neck is shortened, and some cervical vertebrae co-ossify to form a rigid base. The deltoid crest of the scapula is greatly elevated and arches broadly over the glenoid. The humerus and forelimb are highly modified for rotary burrowing (Reed and Turnbull, 1965; Barnosky, 1981, 1982). The humeral head is laterally compressed, and the distal end is greatly expanded, with distinct fossa for insertion of a major muscular ligament. The olecranon process of the ulna is transversely expanded. The tibia and fibula are unfused.
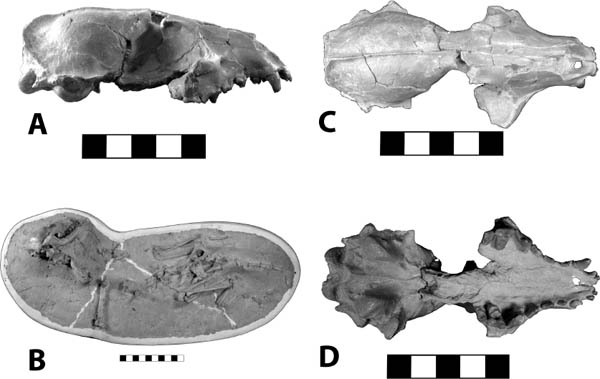
Systematics and Evolution The type species for Proscalops is P. miocaenus Matthew, 1901. The type specimen for Proscalops (AMNH 8949a) was collected in Whitneyan rocks of northeast Colorado. The locality of the holotype of P. tertius (AMNH 19420) is uncertain. It was collected by G. L. Jepsen from the “White River Formation, Badlands South Dakota.” Reed (1961) noted that it may have come from the Brule Formation, but no precise information is available (Gunnell et al., 2008).
Distinctive Characters The antemolar region is relatively short with six antemolar teeth. The molars are relatively high crowned. The coronoid process is high and spicular. The anterior lower incisor is gliriform, and the lower molars are brachyhypsodont to hypsodont. The upper molar metaconules and paracones are prominent (Fig. 5.11C, D) (Gunnell et al., 2008).
Stratigraphic and Geographic Distribution Proscalops is found in the Chadron and Brule formations of the White River Badlands in South Dakota. Fossils are also found in Wyoming, Colorado, Nebraska, Saskatchewan, and Montana.
Soricomorphs, or shrews, have a more derived and specialized dentition, which indicates a more strictly insectivorous diet. Many soricomorphs have molars with high, pointed cusps on tall trigonids, with either low and short talonids or well-developed broad talonids. Upper molars tend to be tritubercular with small or absent hypocones, and tall and sharply pointed cusps in more primitive forms. Skulls tend to be low with short infraorbital canals, enlarged lacrimal canals, relatively short rostrae, and relatively globular braincases. Zygomatic arches are often absent or weakly formed, and there is no ossified auditory bulla. There are few postcrania of fossil soricomorphs known (Gunnell et al., 2008).
The snout is long and tubular, abruptly narrowing above the P3 in later forms. The zygomatic arch is lost and the lacrimal foramen is large, opening into the orbit. The nasals are elongate, widest above the P3, and not fused to the other skull bones.
The dental formula for this family is I3/3, C1/1, P4/4, M3/3. The I1 is enlarged. The P1 is doubled rooted, and the P4 lacks a metacone. The M1–3 have a strong stylar shelf. The M3 has a strong parastyle and a weak metacone. The M1–2 have a separate hypoconal root. The lower premolars are double rooted, the p3 becomes progressively reduced, and the p4 becomes progressively simplified (McKenna, 1960).
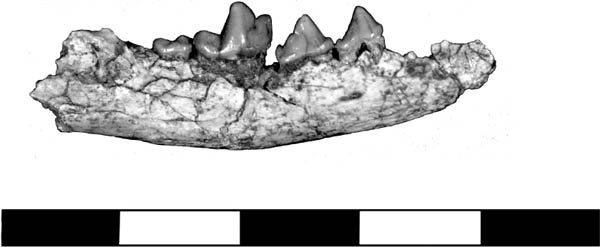
Systematics and Evolution The type species for Centetodon is C. pulcher Marsh, 1872. The type specimen for Centetodon is YPM 13507, which was found in Sweetwater County, Wyoming. Centetodon magnus Clark, 1936, originally described as Metacodon magnus, was found in the Chadron Formation in the White River Badlands.
Distinctive Characters Centetodon is a small soricomorph. The snout abruptly narrows at the level of the P3. The palatal margin is elevated at the anterior to the midpoint of the P4 (Lillegraven, McKenna, and Krishtalka, 1981).
Stratigraphic and Geographic Distribution Centetodon has a long temporal range. Lillegraven, McKenna, and Krishtalka (1981) thoroughly reviewed the Bridgerian and younger Centetodon, making it one of the most completely studied early Tertiary insectivores. Centetodon is the last of the Geolabidae and survives almost to the end of the Arikareean. Fossilized remains from Centetodon can be found in the Wasatchian through the late Arikareean. Fossils are found in Texas, Utah, Wyoming, Colorado, California, Saskatchewan, North Dakota, South Dakota, Nebraska, Montana, and Florida.
Members of this family are about the size of moles and were probably fossorial. The rostrum was long, moderately wide, and deep. The sutures tend to fuse easily. The upper cheek teeth are hypsibrachydont, and the ectolophs slant strongly lingually. The I1 is greatly enlarged, the P1 is absent, the P3 has an extremely reduced protocone, and the P4 is submolariform with a small hooklike cusp behind and above the mesostyle at the posterior end of the sectorial blade of the ectoloph. The M1–2 are much larger than the p2 and are somewhat molariform. The p4 and lower molars are high and have anteroposteriorly narrow trigonids with the upper half of the crown curving posteriorly (Stirton and Rensberger, 1964).
Systematics and Evolution The type and only species of Clinopternodus is C. gracilis Clark, 1937. The type specimen is YPM 14197 and was found in the Chadron Formation in Shannon County, South Dakota.
Distinctive Characters Clinopternodus is one of the larger genera in this family. The lower canine and the crowns of the p3–m1 are relatively procumbent and lingually curved. The tips of the crown are not recumbent. The p2 is absent, and the p3 has a relatively low crown. The m1 talonid is equal to two-thirds of the height of the trigonid (Stirton and Rensberger, 1964).
Stratigraphic and Geographic Distribution Fossilized remains from Clinopternodus are only found in the Chadronian. Its geographic range is also limited. Fossils are found in South Dakota and Wyoming. Clinopternodus is only known at one locality where it has been identified to species level.
The anatomy of the Apternodontidae is well known (Schlaikjer, 1933; McDowell, 1958; McKenna, 1960; Reed and Turnbull, 1965; Asher et al., 2002). The family consists of relatively small soricomorph insectivores that, on the basis of scant postcranial material, were almost certainly fossorial.
The lacrimal foramen is enlarged and laterally facing, and the anterior margin of the infraorbital canal is concave. A sagittal crest is present and continuous with a strong nuchal crest. The posterolateral braincase is expanded into lambdoid plates, and the entoglenoid process is enlarged. The postglenoid process is absent. The zygomatic arch is incomplete, and there is no ossified auditory bulla (Asher et al., 2002).
The dental formula for the Apternodontidae is as follows; I2/3, C1/1, P3/3, M3/3. The anterior incisors are enlarged and procumbent. The upper and lower P4 is molariform, and the upper molars are zalambdodont with metacones absent. The coronoid process extends anteriorly (Asher et al., 2002).
Systematics and Evolution The type species of Apternodus is A. mediaevus Matthew, 1903. The type specimen of Apternodus is AMNH 9601, which was found in Chadronian rocks near Pipestone Springs, Jefferson County, Montana.
Distinctive Characters Same as Family. Apternodus is the only genus in the family Apternodontidae.
Stratigraphic and Geographic Distribution Fossilized remains from Apternodus occur from the Duchesnean to the late Orellan. It has a long temporal range of about 8 million years. Fossils are found in Wyoming, South Dakota, Montana, Nebraska, Texas, Colorado, and Saskatchewan. In the White River Badlands, Apternodus is found in the Chadron Formation.
The posterior braincase is unspecialized, and a prominent entoglenoid process is present. The foramen ovale is enlarged, and the alisphenoid canal is absent. The ethmoid foramen and sinus canal exit anterior to the sphenorbital fissure. The lacrimal foramen is large and laterally oriented (Asher et al., 2002).
The molars are zalambdodont. The upper molars have distinct protocones, anterior cingula, and lack metacones. The lower second and third incisors are tricuspid. The lower molars have reduced talonid basins. The M3 talonid cusp is taller than the paraconid. The coronoid process is pocketed medially (Asher et al., 2002).
Systematics and Evolution The type species of Oligoryctes is O. cameronensis Hough, 1956. The type specimen of Oligoryctes is USNM 19909, which was collected from the White River Formation along Beaver Divide in Wyoming. Oligoryctes altitalonidus Clark, 1936, was found in the Chadron Formation in the White River Badlands.
Distinctive Characters Same as Family. Oligoryctes is the only genus in the family Oligoryctidae.
Stratigraphic and Geographic Distribution Fossilized remains from Oligoryctes can be found in the Uintan to the Whitneyan. The genus has a long temporal range of about 14 million years. Fossils are found in Wyoming, Nebraska, Montana, South Dakota, California, North Dakota, Colorado, and Saskatchewan.
This family is commonly referred to as shrews. The skull has a relatively short rostrum in which the bones fuse early. The cranium, although small, is inflated. The lacrimal duct curves upward and forward from a small, ventrally exiting foramen on an infraorbital bridge. The zygomatic arches are thin and the jugal is absent. The tympanic ring is large, lying nearly horizontal and close to the skull. The auditory bullae are not ossified (Gunnell et al., 2008:112).
The dentary tapers anteriorly with a long procumbent symphysis. The coronoid process is oriented vertically to a horizontal ramus. The upper first incisor is hook shaped, and the teeth between the I1 and P4 are unicuspid and single rooted. The P4 is semimolariform with a well-developed and basined lingual shelf and a large paracone extended posteriorly into a prominent blade. The M1–2 is dilophodont and roughly rectangular with a well-developed cingular hypocone and crested protocone. The upper dentition is reduced by one or more antemolars, and the lower dentition is reduced by two or more antemolars (Gunnell et al., 2008:113). The incisors of shrews are red from hematite in the enamel. Little is known of the postcranial skeleton of most fossil shrews. The skeleton of shrews in general is unspecialized (Reed, 1951).
Systematics and Evolution The type species for Domnina is D. gradata Cope, 1873. The type specimen for Domnina is AMNH 5353, collected from the White River Formation, Logan County, Colorado. Domnina gradata is also found in the White River Badlands of South Dakota.
Distinctive Characters Domnina is the least specialized of the shrews, with only a weakly differentiated condyle on the jaw and five or six lower antemolars. The m1–3 entoconid crest is high and joined to the metaconid but distinctly separated from the postcristid (Gunnell et al., 2008:113).
Stratigraphic and Geographic Distribution Fossilized remains from Domnina can be found in the Uintan to the Hemingfordian. It has a broad temporal range, lasting for 25 million years. Fossils are found in Wyoming, Colorado, South Dakota, Nebraska, Saskatchewan, Montana, California, North Dakota, and Idaho. At Badlands National Park, Domnina has been found in the Scenic Member of the Brule Formation in the North Unit of the park.
Natural History and Paleoecology Domnina is considered the first soricid, or true shrew. A large radiation of the soricids begins in the late Arikareean with a broad diversity through the Miocene and Pliocene. In North America today, the two most common genera are Sorex and Blarina.
The Order Lagomorpha includes pikas, rabbits, and hares. The earliest record of the order is from the Eocene of Asia, and the group first appears in the late Uintan of North America (Dawson, 2008). Early rabbits are placed in the subfamily Palaeolaginae, and this includes Palaeolagus and Megalagus, the two rabbit taxa found in the White River Badlands. The various subfamilies of rabbits are distinguished by differences in the folding of the enamel patterns of the lower third premolar and upper second premolar (Dawson, 2008).
Members of the order are characterized by two pairs of upper incisors in which a smaller peglike incisor is positioned behind the larger anterior grooved tooth of the pair. The cheek teeth, three upper and two lower premolars, and two to three upper and lower molars are hypsodont and ever-growing. The facial portion of the maxillae may have a single or multiple fenestrae. The bony palate is short and often a narrow bar. The auditory bullae is enlarged and formed from the ectotympanic. The distal end of the humerus is lacking an epicondylar foramen. The tibia and fibula are fused distally with the fibula contacting the calcaneum (Dawson, 2008).
The Leporidae includes both rabbits and hares and today is represented by four genera in North America: Lepus, Sylvilagus, Brachylagus, and Romerolagus. Characteristic locomotion in the family includes short bursts of speed, as in rabbits, or sustained bounding locomotion, as in hares (Dawson, 2008).
Systematics and Evolution The type species of Palaeolagus is P. haydeni Leidy, 1956. The type specimen of Palaeolagus, ANSP 11031, was collected in at the head of Bear Creek in the Badlands of South Dakota (Leidy, 1869). Palaeolagus is a common member of many Oligocene mammalian faunas, and many species have been described. The number of species also reflects the long chronology of the genus, which extended into the early Miocene.
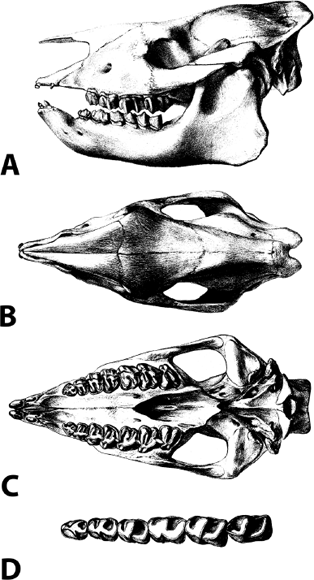
Distinctive Characters The teeth in Palaeolagus are more hypsodont and have better-developed cement tracts than Megalagus and earlier rabbits. The overall size is smaller than that of Megalagus (Fig. 5.12) (Dawson, 2008).
Stratigraphic and Geographic Distribution Palaeolagus is found in the Chadronian through the Arikareean. In the White River Badlands, Palaeolagus is found in the Brule Formation. Fossils are found in South Dakota, Nebraska, Wyoming, Colorado, Saskatchewan, Montana, North Dakota, and Oregon (Dawson, 2008).
5.12. Palaeolagus haydeni. (A) Skull and jaws. (B) Partial skull, occlusal view. (C) Right and left mandibles, occlusal view. Scales in centimeters. Photos by the authors of specimens from the Department of Earth Sciences teaching collection, University of Northern Colorado.
Natural History and Paleoecology In lagomorphs, one of the evolutionary changes seen is in the increase in hypsodonty, which coincides with the cooling and drying that occurred at the Eocene–Oligocene transition and the related changes in vegetation. Change in dentition was also accompanied by changes in posture that affected locomotion (Dawson, 2008).
Systematics and Evolution The type species of Megalagus is M. turgidus (Cope, 1873), which was originally described as Palaeolagus turgidus. The type specimen of Megalagus, AMNH 5635, was collected from the White River Formation, Logan County, Colorado. Evolutionary trends in Megalagus parallel similar trends in Palaeolagus, but at any given time, Megalagus tends to lag behind Palaeolagus in most dental characters (Dawson, 2008).
Distinctive Characters Megalagus is characterized by upper molariform teeth with buccal roots. The overall size is larger than Palaeolagus except for one species, P. intermedius (Dawson, 1958).
Stratigraphic and Geographic Distribution Megalagus is found in the Chadronian through the Hemingfordian. In the White River Badlands, Megalagus is found in the Chadron and Brule formations. Fossils are found in South Dakota, Nebraska, Saskatchewan, Montana, North Dakota, Colorado, and Florida.
Rodents are considered to be a successful group, a result of their taxonomic diversity and wide geographic distribution. Over 1800 living species are contained within this group, making up 50 percent of all living mammals. The rodent fossil record can be found on all continents except Antarctica. One of the defining features of rodents is their unique teeth and jaws, which played a significant role in their adaptations and evolutionary radiation. Rodents are characterized by a single pair of deep-rooted incisors that grow throughout their lives. The incisors are used to gnaw wood and nuts and to clip vegetation and husks of fruit. The incisors bear enamel only on the anterior surface so that the softer dentine behind wears faster and provides a sharp edge that is self-sharpening. Rodent jaws are also characterized by a diastema between the single incisor and the first cheek tooth, which is either the fourth premolar or first molar, a gap that in other mammals would be located at the second and third incisors, the canine, and the first three premolars (Fig. 5.13) (Benton, 2000).
5.13. Diagram showing individual features of a rodent skull and jaws (Ischyromys sp.). Images modified from Wood (1937:plate 27, figs. 1, 1a).
Rodents first appear in North America during the late Paleocene and early Eocene. By the late Eocene and Oligocene, they are quite diverse and well adapted to a variety of environments.
The family Ischyromyidae is one of the earliest rodent families to appear in the fossil record, first appearing in the late Paleocene to early Eocene in North America and also in Europe. They were the most diverse taxonomically during the Uintan (Korth, 1994b). Ischyromids are the most common rodents in the Orellan and have been used for biostratigraphic correlation (Prothero and Whittlesey, 1998). During the early Oligocene, their diversity decreases and they become extinct in the Whitneyan (Korth, 1984, 1994b; Heaton, 1993; Howe, 1996). The family Ischyromyidae may have been the ancestor to later Tertiary rodent families, such as Sciuridae and Aplodontidae (Emry and Korth, 1996).
Systematics and Evolution The type species of Ischyromys is I. typus from the Badlands of South Dakota. Currently seven species are recognized. The genus first appears in the middle Wasatchian and became extinct at the end of the Whitneyan. Ischyromys typus is known from the Scenic and Poleslide members of the Brule Formation. Ischyromys parvidens is from the Scenic Member of the Brule Formation. Ischyromys spp. from the Chadron Formation have not been identified to species (Anderson, 2008).
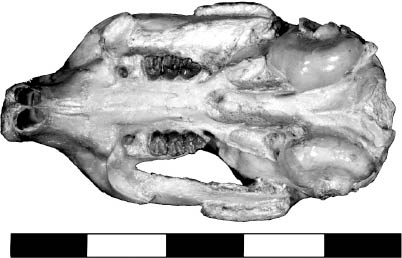
Distinctive Characters Members of the family Ischyromyidae are defined by a robust skull, mandible, and skeleton. The skull has a heavy rostrum, a small incisive foramina, and a sagittal crest on the cranium (Fig. 5.14A, D). These rodents have a protrogomorphous morphology where the masseter muscles attach to the zygomatic arch (primitive form). They also have the complete rodent dental formula of two upper premolars and one lower. The upper third premolar is a peg. In the subfamily Ischyrominae, the auditory bullae were co-ossified with the skull, and the cheek teeth were lophodont (Fig. 6.14B, C, E) (Black, 1968, 1971; Wahlert, 1974; Anderson, 2008). Ischyromys was a medium-size rodent with incisors that were oval in cross section; the lower molar crown pattern was crestlike; and the length of the molars was slightly greater than the width. Also the scaphoid and lunate bones (wrist) are fused in Ischyromys (Anderson, 2008).
Stratigraphic and Geographic Distribution Fossilized remains of Ischyromys occur in the Wasatchian through the early Whitneyan. Fossils are found in Wyoming, Colorado, South Dakota, California, Nebraska, Texas, Montana, Saskatchewan, North Dakota, and Utah.
Natural History and Paleoecology On the basis of its skeletal anatomy, Ischyromys was thought to have adaptations for terrestrial quadripedal locomotion ranging from scampering to subfossorial (Korth, 1994b; Anderson, 2008). It was also considered to be the most common rodent in the Orellan of the White River Group and the last genus of this family in the fossil record (Korth, 1994b).
The family consists of a holarctic radiation of small to medium and sometimes large rodents. There is a single surviving species, Aplodontia rufa, the mountain beaver from the Pacific Northwest of North America. Fossils from the family Aplodontidae are found in North America, Europe, and Asia. The center of diversity was during the mid-Cenozoic in North America, with repeated dispersals. This family was considered diverse in the western part of North America during the Oligocene and Miocene epochs. There were two distinctive groups that evolved during the mid-Tertiary. The Allomyinae–Aplodontinae subfamilies appeared in the later early Oligocene, diversified, and survives today as Aplodontia. The Aplodontids are small- to medium-size rodents that show various dental specializations including extra cusps on cheek teeth, development of an ectoloph on the upper cheek teeth, and the presence of molar styles. The skull and skeleton show a terrestrial and fossorial skull and postcranial adaptations. Aplodontids possibly evolved from the early Tertiary ischyromyoid rodents from North America. A recent molecular study shows a close link to squirrels (Adkins, Walton, and Honeycutt, 2002). Early morphologists noted that aplodontoid molars resemble squirrel molars in the arrangement of cusps and the lack of a hypocone (Flynn and Jacobs, 2008).
Systematics and Evolution The type species of Prosciurus is P. vetustus Matthew, 1903. The type specimen is AMNH 9626, collected from an unknown location. Prosciurus relictus Cope, 1873, has been collected from the Brule Formation, in the White River Badlands of South Dakota.
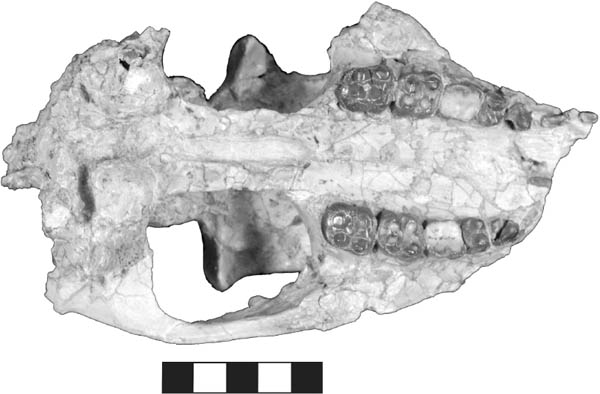
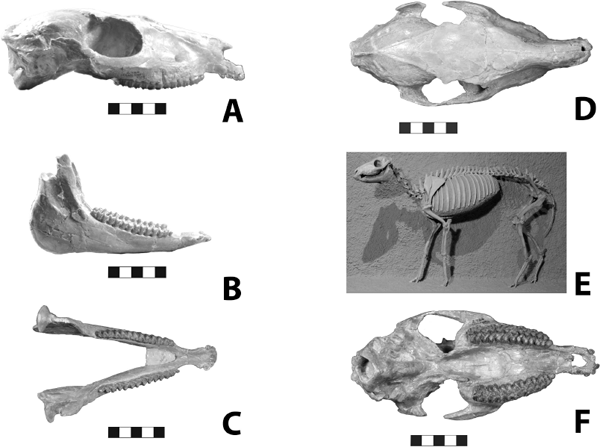
5.14. Ischyromys plaicus, BADL 53890/SDSM 2994. (A) Skull, left lateral view. (B) Mandibles, left lateral view. (C) Mandibles, occlusal view. (D) Skull, dorsal view. (E) Skull, occlusal view. Scales in centimeters. Photos by the authors of specimen from the Museum of Geology, South Dakota School of Mines and Technology, Rapid City, South Dakota, U.S.A., and is the property of the U.S. government.
Distinctive Characters Prosciurus had brachydont and cuspate upper molars with a continuous anteroloph–protocone–posteriorloph crest. The hypocone is absent or minute, and the metaloph is incomplete. The lower molars are rhombic, with broad, shallow basins (Flynn and Jacobs, 2008).
Stratigraphic and Geographic Distribution Fossilized remains of Prosciurus can be found in the Uintan to Whitneyan in North America. Fossils are found in Wyoming, Nebraska, Saskatchewan, North Dakota, Montana, Colorado, and South Dakota.
Natural History and Paleoecology Prosciurus was originally proposed as a subgenus for Sciurus (Matthew, 1903). It is one of the earliest aplodontines and possibly was the ancestor to the later Mylagualidae (Flynn and Jacobs, 2008).
Systematics and Evolution The type species of Pelycomys is P. rugosus Galbreath, 1953. The type specimen is KU 8343, which was collected from the White River Formation, Logan County, Colorado. Pelycomys is only identified to genus in the White River Badlands of South Dakota.
Distinctive Characters Pelycomys was a large rodent with brachydont cheek teeth that were subtriangular to sub-rhombic. The cusps were round and inflated (Flynn and Jacobs, 2008).
Stratigraphic and Geographic Distribution Pelycomys occurs in the Chadronian to Arikareean in North America. Fossils are found in Colorado, Wyoming, Nebraska, Montana, and South Dakota. In the White River Badlands, Pelycomys can be found in the Sharps Formation.
The family Sciuridae, also known as squirrels, chipmunks, prairie dogs, and marmots, comprise about 275 living and numerous extinct species. Squirrels are found worldwide except for Australia, southern South America, and Antarctica (Jones et al., 1983). The first sciurids appear in the Chadronian of North America and in similar age rocks in Europe (Goodwin, 2008). The oldest squirrel, Douglassciurus jeffersoni, from Flagstaff Rim, Wyoming, is one of the most complete and best described of the fossil squirrel specimens from North America (Emry and Thorington, 1982). Douglassciurus jeffersoni has a similar skeleton to a modern tree squirrel. All members of Family Sciuridae have sciuromorphic zygomasseteric structures except for D. jeffersoni. Within Badlands National Park, only two sciurid genera are found, Cedromus and Protosciurus (Goodwin, 2008).
5.15. Cedromus wilsoni, USNM 256584. Skull, occlusal view. Scale in centimeters. Photo by the authors. Courtesy of Smithsonian Institution, Washington, D.C., U.S.A.
Systematics and Evolution The type species for Cedromus is C. wardi Wilson, 1949. The type specimen is UCM 19808 from the Orellan rocks of Colorado.
Distinctive Characters Cedromus is characterized by a low, broad skull, short rostrum, and a primitive zygomasseteric structure. The zygoma is anteriorly broadened and tilted lateral to the infraorbital foramen. The upper molars had a small hypocone and large metaconules (Fig. 5.15). The lower molars are characterized by a posterior cingulum confluent with the entoconid and by the mesostylids having a greater buccal expansion (Korth and Emry, 1991).
Wilson (1949) interpreted Cedromus to be an ischyromyid but suggested the possibility that it could have been a primitive squirrel. Korth and Emry (1991) placed Cedromus in with the sciurids on the basis of well-preserved skull material.
Stratigraphic and Geographic Distribution Fossilized remains from Cedromus can be found in the Orellan through the Whitneyan. Fossils are found in South Dakota, Wyoming, Nebraska, Colorado, and Saskatchewan. In the White River Badlands, Cedromus is found in the Brule Formation.
Natural History and Paleoecology There are four major groups of squirrels: chipmunklike squirrels, tree squirrels, flying squirrels, and terrestrial squirrels, which include ground squirrels, prairie dogs, and marmots. The fossil squirrels found in the White River Badlands were believed to be arboreal in nature because terrestrial squirrels had not yet evolved. They are believed to have appeared and radiated in the late Arikareean to Barstovian (early to middle Miocene) (Goodwin, 2008).
Systematics and Evolution The type species of Protosciurus is P. condoni Black, 1963. The type specimen is UO F-5171, from the early Arikareean of Oregon. Protosciurus is only identified to genus in the White River Badlands of South Dakota. The skull of Protosciurus is not fully sciuromorphous but is more advanced than Douglassciurus or Cedromus. Overall, Protosciurus is still considered to be primitive compared to other squirrel groups (Goodwin, 2008).
Distinctive Characters The teeth are low crowned, subquadrate, with low lophs and a broadened protocone. The skeleton is gracile and is similar to those of nut- and seed-eating squirrels today (Black 1963, 1972).
Stratigraphic and Geographic Distribution Fossilized remains of Protosciurus occur in the Orellan to Hemingfordian in North America. Fossils are found in South Dakota, Florida, Oregon, Nebraska, Texas, California, and Saskatchewan. In the White River Badlands, Protosciurus is found in the Sharps Formation.
Natural History and Paleoecology On the basis of a single tibia assigned to this genus, it is thought that Protosciurus was more arboreal than terrestrial (Frailey, 1978).
The family Castoridae includes fossil and modern beavers. Living members of this family can weigh as much as 40 kg, with fossilized forms weighing as little as 1 kg. Beavers reached their maximum size in the Pleistocene with Castoroides, which may have weighed as much as 100 kg. Modern beavers are semiaquatic, but ancient forms were terrestrial, and some showed burrowing adaptations. Early beavers had relatively low crowned teeth but achieved full hypsodonty in the late Tertiary. Beavers originated in North America and migrated to Europe (Flynn and Jacobs, 2008). The greatest diversity of castorids occurred in the Arikareean with the radiation of palaeocastorine beavers. A third radiation involved the castorine beavers, which occurred in the Blancan.
5.16. (A) Agnotocastor praetereadens, AMNH 1428, skull, occlusal view. (B) Agnotocastor sp., BADL 22370/SDSM 56624, partial right mandible with lower p4–m3, occlusal view. Scales in centimeters unless otherwise noted. Photo A by the authors; and photo B by Edward Welsh. AMNH 1428 is from the American Museum of Natural History, New York, New York, U.S.A. BADL 22370/SDSM 56624 is from Badlands National Park, Interior, South Dakota, U.S.A., and the Museum of Geology, South Dakota School of Mines and Technology, Rapid City, South Dakota, U.S.A., and is the property of the U.S. government.
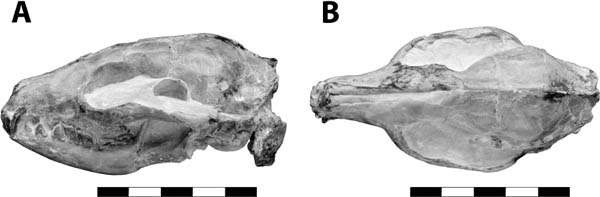
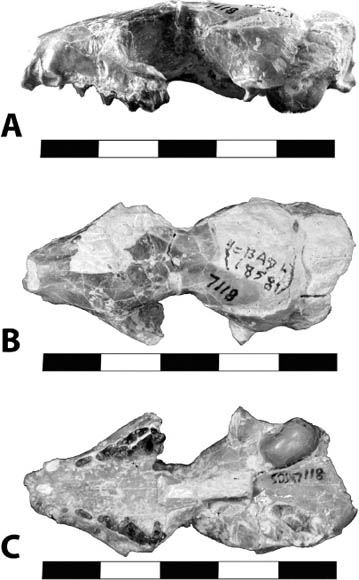
Systematics and Evolution The type species of Agnotocastor is A. praetereadens Stirton, 1935. The type specimen of Agnotocastor is AMNH 1428, collected near the Cheyenne River, South Dakota, in the Poleslide Member of the Brule Formation (Whitneyan). Agnotocastor is considered the earliest and most primitive castorid (Flynn and Jacobs, 2008). Because it appeared in both Asia and North America at the same time, it is unclear what direction the migration occurred (Korth, 1994b).
Distinctive Characters Agnotocastor is characterized by relatively low crowned teeth, and a narrow rostrum and skull (Fig. 5.16). The auditory bulla is kidney shaped. The lower incisors are rounded. The P3 is present and the P4 is square. All of the molars are rectangular (wider than long) (Fig. 5.16) (Flynn and Jacobs, 2008).
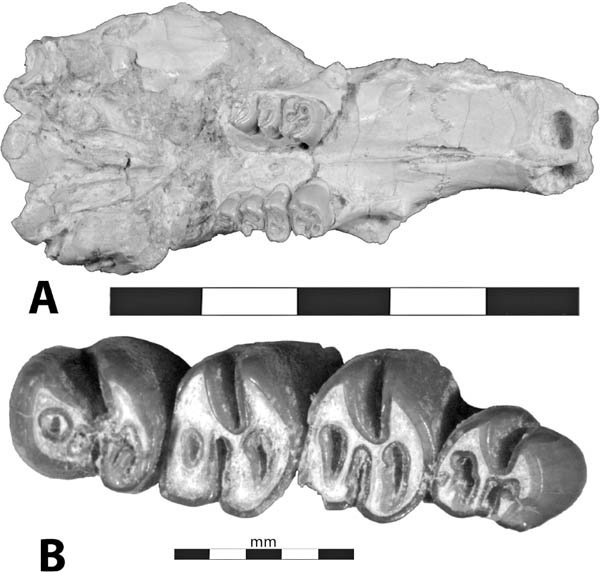
5.17. Eutypomys sp. (A) Skull and jaws, left lateral view (Wood, 1937:224). (B) Eutypomys thomsoni, AMNH 12254, left P3–M3 (Wood, 1937:225). (C) Eutypomys sp. left p3–m3, BADL 63385. Scales in centimeters unless otherwise noted. Photo C by Levi Moxness. BADL 63385 is from Badlands National Park, Interior, South Dakota, U.S.A., and is the property of the U.S. government.
Stratigraphic and Geographic Distribution Fossilized remains from Agnotocastor occur in the Chadronian to the early Arikareean. Fossils are found in South Dakota, Colorado, Wyoming, Nebraska, Florida, and Montana. In the White River Badlands, Agnotocastor is found in the Chadron and Brule formations.
Natural History and Paleoecology Agnotocastor was a terrestrial beaver, but there is no evidence that it burrowed. The radiation of Palaeocastor, with its famous Daemonelix burrows (devil’s corkscrews), is separate from the Agnotocastor lineage. Palaeocastor is only contemporaneous with Agnotocastor during the Whitneyan and continues into the Miocene (Korth, 1994b).
Systematics and Evolution The type species of Palaeocastor is P. nebrascensis (Leidy, 1856), which originally was described as Steneofiber nebrascensis. The exact location where the type was found is unknown.
Distinctive Characters Palaeocastor is characterized by high-crowned cheek teeth, which readily distinguishes it from Agnotocastor; large, rounded bullae; convergent suborbital ridges; moderate lambdoidal crest; incisors that are somewhat rounded; and premolars that are close to molars in size (Flynn and Jacobs, 2008). Reynolds (2002) estimated the body mass of Palaeocastor to be about 1 kg.
Stratigraphic and Geographic Distribution Palaeocastor occurs in the Whitneyan to the Hemingfordian. Fossils are found in Wyoming, South Dakota, Nebraska, North Dakota, Oregon, Texas, Saskatchewan, and Montana. The fossil record for Palaeocastor at Badlands National Park is limited to a few localities within the Sharps Formation (Parris and Green, 1969; Flynn and Jacobs, 2008).
Natural History and Paleoecology Martin (1987b) noted that skeletal remains of both P. fossor and Capacikala magnus were recovered from Daemonelix burrows, showing that Palaeocastor and Capacikala dug these amazing spiral features. No Daemonelix burrows have been found in Badlands National Park.
Systematics and Evolution The type species of Capacikala is Steneofiber gradatus Cope, 1878, and the species was originally placed in the genus Steneofiber. The type specimen of Capacikala, AMNH 7008, was collected from the John Day Formation, Grant County, Oregon. Martin (1987b) and Korth (1994b, 2001) proposed the subfamily Palaeocastorinae, which included both Palaeocastor and Capacikala.
Distinctive Characters Capacikala is considered a small beaver with a short, deep skull and orthodont incisors that have flattened anterior faces. There are double temporal muscle scars that do not meet in a single sagittal crest. The bullae are large and expand ventrally. The cheek teeth decrease in size posteriorly (Flynn and Jacobs, 2008).
Stratigraphic and Geographic Distribution Fossilized remains from Capacikala can be found in the early to middle Arikareean in North America. Fossils are found in Wyoming, South Dakota, Montana, and Oregon. Both Palaeocastor and Capacikala were found in channel deposits within the Sharps Formation in Cedar Pass at Badlands National Park (Parris and Green, 1969).
Natural History and Paleoecology The subfamily has adaptations for a subterranean habitat, including a flattening of the enamel surface of the incisors to form a chisellike digging tools and a shortened rostrum (Flynn and Jacobs, 2008).
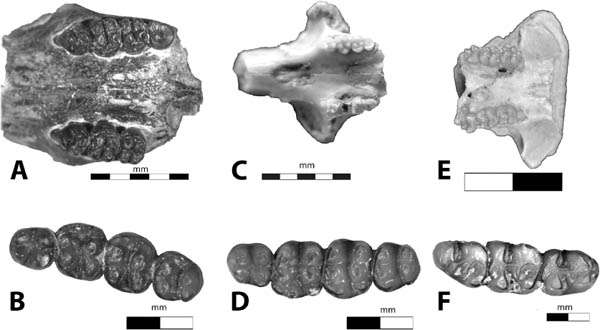
5.18. Rodents from the White River Badlands (A) Proheteromys nebraskensis, palate with right P4–M2 and left P4–M3, BADL 18589/SDSM 10001. (B) Proheteromys nebraskensis right p4–m3, BADL 18588/SDSM 10000. (C) Heliscomys sp., rostrum with no incisors, UCM 45468. (D) Heliscomys sp., left p4–m3, BADL 22286/SDSM 56540. (E) Eumys elegans, palate with right and left M1–M3, FM 38871. (F) Eumys sp., left m1–m3, BADL 22282/SDSM 56536. Scales in centimeters unless otherwise noted. Photos (A), (B), (D), and (F) by Edward Welsh; photos (C) and (E) by the authors. BADL 18589/SDSM 10001, BADL 18588/SDSM 1000, BADL 22286/SDSM 56540 are from Badlands National Park, Interior, South Dakota, U.S.A., and the Museum of Geology, South Dakota School of Mines and Technology, Rapid City, South Dakota, U.S.A. UCM 45468 is from the University of Colorado Museum of Natural History, Boulder, Colorado, U.S.A. FM 38771 is from the American Museum of Natural History, New York, New York, U.S.A.
Eutypomyidae is a sister group with Castoridae, on the basis of a mix of primitive and advanced tooth patterns. The teeth are high crowned and based on a four-cusp pattern, which is an advanced feature in the family Castoridae. The muscle and jaw morphology demonstrates a fully sciuromorphous condition (Flynn and Jacobs, 2008). The rostrum is elongate.
Systematics and Evolution The type species of Eutypomys is E. thomsoni Matthew, 1905. The type specimen of Eutypomys, AMNH 12254, is from the Scenic Member of the Brule Formation on Quinn Draw, Cheyenne River, South Dakota.
Distinctive Characters Eutypomys has a complicated crown pattern consisting of a complex of secondary loops that result in separate enamel islets. The complex eutypomyid tooth morphology is built upon a fundamentally primitive dentition including a 5/4 cheek tooth formula (Fig. 5.17) (Flynn, Lindsay, and Martin, 2008).
Stratigraphic and Geographic Distribution Fossilized remains of Eutypomys occur in the Duchesnean to Arikareean in North America. Fossils are found in South Dakota, Nebraska, Saskatchewan, Montana, North Dakota, Wyoming, and Texas. Eutypomys is found through a long section of the fossil record, and individual species are valuable stratigraphic indicators. Eutypomys thomsoni is an indicator species of the Early Late Orellan Miniochoerus gracilis Interval Zone (Prothero and Whittlesey, 1998), and the genus Eutypomys helps typify the Orellan land mammal age.
The cricetids are a successful group of rodents. Their record extends from the late Eocene to the Recent. They originated in Asia, migrated into North America, and radiated into the Neotropics during the Pliocene. They are small rodents and are adapted to a large variety of environments, including xeric, boreal, aquatic, and semifossorial. Most species are herbivorous. Both fossil and modern cricetids are successful because of their high fecundity and broad ecological tolerance (Lindsay, 2008). In terms of jaw musculature, the family Cricetidae represents a transition from subhystricomorphous to a myomorphous zygoma. The mandible is slender, and the cheek teeth are restricted to three molars, upper and lower and on each side. The incisors are narrow anteriorly and gently convex. The enamel surface is ornamented by numerous ridges. The braincase is inflated and the rostrum is narrow. The interorbital area has a smooth hourglass shape, and the auditory bullae are inflated and usually small. The hind limb is longer than the forelimb, and the fibula is fused to the tibia (Korth, 1994b).
Systematics and Evolution The type species of Eumys is E. elegans Leidy, 1856. The type specimen of Eumys is ANSP 11027, from the Scenic Member, Brule Formation, South Dakota. Besides the type species, eight other species are considered valid (Lindsay, 2008:465). Eumys represents the first known cricetid in North America. The eumyines represented the first radiation of cricetids, which achieved its greatest diversity in the Orellan with seven species (Lindsay, 2008:465).
Distinctive Characters Eumys was a small- to mediumsize rodent with brachydont dentition. The skull has a broad rostrum and a median sagittal crest. The dentition is slightly lophodont with robust cusps. Both upper and lower third molars are quite large (Fig. 5.18E, F) (Lindsay, 2008).
Stratigraphic and Geographic Distribution Fossilized remains of Eumys can be found in the Duchesnean to Arikareean in North America. Fossils are found in Wyoming, Colorado, South Dakota, Nebraska, Saskatchewan, Montana, and North Dakota. Eumys brachyodus is reported from the Poleslide member of the Brule Formation, and E. obliquidens and E. parvidens from the Scenic Member of the Brule Formation (Lindsay, 2008).
Natural History and Paleoecology Eumys is generally considered the most common and best-known Oligocene cricetid (Lindsay, 2008).
Systematics and Evolution The type species of Scottimus is S. lophatus Wood, 1937. The type specimen of Scottimus, MCZ 5064, is from the Poleslide Member of the Brule Formation from South Dakota. Besides the type species, currently five other species in the genus are considered valid.
Distinctive Characters Scottimus was a small brachydont rodent with a parallel-sided snout, parasagittal crests, and cheek teeth developing lophs directed anteroposteriorly between the labial cusps and on the lingual side. The upper molars decrease in size posteriorly (Lindsay, 2008:462).
Stratigraphic and Geographic Distribution Fossilized remains of Scottimus occur in the Orellan to Arikareean. Fossils are found in South Dakota, Wyoming, Nebraska, Montana, Colorado, and Saskatchewan.
Natural History and Paleoecology Scottimus was part of the first radiation of cricetids during the late Eocene to Oligocene pulse that began in the Chadronian with Eumys and climaxed in the early Arikareean. Cricetids declined during the latter part of the Arikareean. A second radiation began in the Barstovian (Lindsay, 2008).
The family Dipodidae includes the modern jumping mice and the birch mice of North America and Asia, which are often referred to as zapodids. They are small- to medium-size rodents and have saltatorial adaptations, especially in the hind limb. There are not many species within this family. Because of their preference for moist conditions, dipodoids can be useful climate indicators. The skull has a hystricomorphous zygomasseteric structure. The molars show a “cricetid” plan, and opposite cusps are joined by transverse crests. Evolutionary trends are toward increasing lophodonty and an emphasis on shearing crests. The upper incisors contain a longitudinal groove (Flynn, 2008a). The postcrania are rarely preserved in the fossil record. Living forms are gracile with limbs modified for saltatorial locomotion.
Systematics and Evolution The type species of Diplolophus is D. insolens Troxell, 1923. The type specimen of Diplolophus, YPM 10368, was collected from the Brule Formation near Scottsbluff, Nebraska. Diplolophus is an early rodent characterized by three cheek teeth. Close affinities with the geomyids (Korth, 1994b; Wood, 1980) suggest a relationship with Geomorpha (extant pocket gophers, pocket mice, and kangaroo rats).
Distinctive Characters Diplolophus is characterized by its large body size, mesodont, and strongly lophodont dentition. The cheek teeth have weak longitudinal connections in the upper molars and are absent in the lowers. The second molars are wider than long (2008a).
Stratigraphic and Geographic Distribution Diplolophus occurs in the Duchesnean through Orellan. Fossils are found in Colorado, South Dakota, Nebraska, and California. In the White River Badlands, Diplolophus is found in the Brule Formation.
Systematics and Evolution The type species of Plesiosminthus is P. schaubi from Saint-Gérand-le-Puy, France (early Miocene), and five species are recognized in North America. Plesiosminthus clivosus Galbreath, 1953, the type specimen of Plesiosminthus, is KU 9279, collected from the Pawnee Creek Formation in Logan County, Colorado. In Badlands National Park, Plesiosminthus is only identified to genus (Parris and Green, 1969).
Distinctive Characters Plesiosminthus is described as a small rodent with grooved upper incisors. The cheek teeth are brachydont, and the M1 has a strong anterocone. The upper molars have a long mesoloph and a reduced M3. The m1 has a prominent mesoconid, variable mesolophid, and a low and tiny anteroconid (2008a). Plesiosminthus is in the same subfamily (Sicistinae) as the modern birch mice.
Stratigraphic and Geographic Distribution Fossilized remains of Plesiosminthus can be found in Arikareean through Clarendonian rocks. Fossils are found in South Dakota, Nebraska, Saskatchewan, Wyoming, Montana, Colorado, and Delaware. In the White River Badlands, Plesiosminthus is found in the Sharps Formation (Parris and Green, 1969).
Natural History and Paleoecology The extant Sicistinae excavate shallow burrows and are found in both forest and steppe environments.
The Heteromyids are a diverse family, with six genera and 300 species. Extant genera include the pocket mice and kangaroo rats. Their locomotor adaptations range from bipedal hoppers to burrowers. These small to medium sciuromorphous rodents have a rich fossil record, especially in the Oligocene and Miocene of the Great Plains (Flynn, Lindsay, and Martin, 2008). The rodents within this family have thin skull bones, and the side of the snout is perforated. Their molars have six cusps with large styles and stylids. The p4 is small and narrow anteriorly. The incisors are laterally compressed, and the postcrania are gracile. The limbs are elongated, and the tibia and fibula are fused (Korth, Wahlert, and Emry, 1991).
Systematics and Evolution The type species of Proheteromys is P. floridanus Wood, 1932. The type specimen of Proheteromys, FSGS V5329, was collected from the Torreya Formation (Miocene) in Gadsden County, Florida. Proheteromys is only known at the genus level in Badlands National Park.
Distinctive Characters Proheteromys is characterized by smooth asulcate upper incisor enamel, mesodont dentition, and four-cusped premolars. The crown height varies with individual species. The anterior cusps on the p4 are close and joining early in wear to isolate a central basin. The lower molars have weak anterior and posterior cingula (Fig. 5.18A, B) (Flynn, Lindsay, and Martin, 2008).
Stratigraphic and Geographic Distribution Proheteromys has a broad range stratigraphically and geographically. However, its occurrence in Badlands National Park is rare. Fossilized remains of Proheteromys occur in the Whitneyan to Barstovian. Fossils are found in Texas, Florida, Colorado, South Dakota, Nebraska, Saskatchewan, California, Delaware, Wyoming, New Mexico, Oregon, and Montana. In the White River Badlands, Proheteromys is found in the Brule and Sharps formations.
The heliscomyids are some of the smallest rodents in North America, both fossil and recent (Korth, 1994b). The skull has a sciuromorphous zygomasseteric structure. The family Heliscomyidae was created by Korth, Wahlert, and Emry (1991). Taxa within this group used to be included within the primitive heteromyids. Green and Bjork (1980) noted indicators of a more complex evolution, and many species were grouped together that had divergent characteristics. This family is part of a larger group called Geomorpha (pocket gophers, pocket mice, and kangaroo rats) and is believed to be an early distinct radiation of the Geomyoidea (Korth, Wahlert, and Emry, 1991). The family was considered primitive because it lacked many of the features that characterize the modern pocket gophers, pocket mice, and kangaroo rats.
Systematics and Evolution The type species of Heliscomys is H. vetus Cope, 1873. The type specimen of Heliscomys is AMNH 5461, collected from the White River Formation in Logan County, Colorado. Besides the type species, three other species are currently recognized as valid.
Two distinct lineages are found within the genus Heliscomys (Korth, Wahlert, and Emry, 1991). One lineage, H. vetus–H. mcgrewi–H. woodi, has a reduction of the premolars and the simplification of molars (Orellan and Arikareean). A separate lineage, H. subtilis, has enlarged and an increased complexity of the premolars (Chadronian–Barstovian). The greatest diversity is in the Orellan, with three genera and five species.
Distinctive Characters Heliscomys is a small geomyine with brachydont teeth showing isolated cusps and strong cingulae-bearing accessory cusps. The upper premolar is triangular with one main anterior cusp and two posterior cusps. The lower premolar has four main cusps and narrows anteriorly. The mandible was dorsoventrally slender, and the diastema is shallow and short. The incisors are small, delicate, and laterally compressed with a narrow, gently convex anterior enamel surface (Fig. 5.18C, D) (Flynn, Lindsay, and Martin, 2008).
Stratigraphic and Geographic Distribution Heliscomys occurs in the Duchesnean to Hemingfordian. Fossils are found in Saskatchewan, California, Wyoming, South Dakota, Nebraska, Montana, Colorado, and Florida. Only H. vetus has been reported from the Scenic Member of the Brule Formation in the Badlands. There are no species known in the Whitneyan, but this absence of Heliscomys spp. could be due to a collecting bias (Korth, 1994b).
The family is characterized by a large optic foramen and reduced incisive foramina. The entostyle is elongated anteroposteriorly. The lower molars have lingual styles, and the cheek teeth are brachydont (Wahlert, 1983).
Systematics and Evolution The type species of Hitonkala is H. andersontau Macdonald, 1963. The type specimen of Hitonkala, SDSM 56120, was collected from Shannon County, South Dakota.
Distinctive Characters Hitonkala is a small florentiamyid with a single anterior cusp on the P4. The p4 is highly molariform. The M1 has a separated protostyle and hypostyle (Flynn, Lindsay, and Martin, 2008). Korth (1993) completed a study of the Hitonkala cranial anatomy that helps clarify the unique features of the family Florentiamyidae.
Stratigraphic and Geographic Distribution Hitonkala occurs in the early to late Arikareean. Fossils are found in Wyoming, South Dakota, Nebraska, and Saskatchewan. Parris and Green (1969) noted the presence of Hitonkala in the Sharps Formation in Cedar Pass at Badlands National Park.
Eomyidae is a complex and rich group that dominated the small-mammal faunas of North America during certain times. They are considered a sister taxa to Geomorpha (living gophers and mice). The family is now extinct. Because of increased screen washing, the group has become better documented in the fossil record. Superficially, eomyids are squirrellike in their body proportions and ground dwelling to arboreal in habits. They are small- to medium-size rodents and are found in North America, Europe, and Asia. Primitive genera occur in the late to middle Eocene of North America and appear during the Oligocene in Europe and Asia. Eomyids persisted to the end of the Miocene in North America (Flynn, 2008b).
Eomyids have a sciuromorphous jaw musculature, but their molars follow a cricetid morphology and are composed of four major cusps joined by thin strong lophs. The mandible is slender with a long diastema. There are no fossorial adaptations to the skull – for example, heavy bone, strong contacts, or a broad posterior skull. Skulls are generally gracile, and there is no sagittal crest. The premolars are submolariform and nearly rectangular. The molars are often wider than long. The cheek teeth are bunodont to pentalophodont. The eomyid postcrania are slender, which is similar to squirrels, and some taxa are inferred to have had gliding membranes, such as Eomys quercyi (Storch, Engresser, and Wuttke, 1996).
Most eomyids were associated with a mesic and forest habitat and with arboreality. Ecomorphological counterparts to Eomyidae might be living small to medium squirrels. However, their dentition follows a more conservative muroid pattern.
Systematics and Evolution The type species of Adjidaumo is A. minutus (Cope, 1873). The type specimen of Adjidaumo, AMNH 5362, was collected from the White River Formation, Logan County, Colorado.
Distinctive Characters Adjidaumo is characterized as a small eomyid and has brachydont cheek teeth, cuspate molars with low crests, and a strong anterior cingulum joined centrally. The incisor enamel is smooth and rounded (Flynn, 2008b). Adjidaumo is often confused with the European taxon Eomys, which migrated from North America.
Stratigraphic and Geographic Distribution Adjidaumo occurs in the Bridgerian to Whitneyan. Fossils are found in Texas, Wyoming, Colorado, South Dakota, Nebraska, Saskatchewan, British Columbia, and Montana. In the White River Badlands, Adjidaumo is found in the Chadron and Brule formations.
Natural History and Paleoecology Eomyids experienced a range expansion across several continents. The confusion between Adjidaumo and Eomys is a classic example of taxonomy driven by continent of occurrence. It is difficult to differentiate between the two taxa on the basis of morphology of individual teeth. McKenna and Bell (1997) placed Adjidaumo in Eomys. Flynn (2008b) concluded that there are enough features to distinguish the two genera.
Systematics and Evolution The type species of Paradjidaumo is P. trilophus (Cope, 1873), which was originally described as Gymnoptychus trilophus. The type specimen of Paradjidaumo, AMNH 5401, was collected from the White River Formation in Logan County, Colorado.
Distinctive Characters Paradjidaumo is a small eomyid. Its molars have developed lophs and moderately high crowns. The valleys, especially the central ones, are narrow and deep. The lower incisor is flattened anteriorly and bears a longitudinal ridge (Black, 1965).
Stratigraphic and Geographic Distribution Paradjidaumo occurs in the Bridgerian to Hemingfordian. Fossils are found in Utah, Wyoming, Colorado, South Dakota, Nebraska, Montana, North Dakota, California, British Columbia, and Saskatchewan. In the White River Badlands Paradjidaumo is found in the Chadron and Brule formations.
Natural History and Paleoecology Paradjidaumo has a broad range stratigraphically and geographically but is still fairly rare at Badlands National Park.
One of the earliest group of mammals to fill the ecological niche as carnivores are members of the Order Creodonta. A primitive group of mammals, they first appeared in the late Paleocene and survived until the end of the Oligocene in North America. Their extinction probably was the result of increased competition from the more advanced members of the Order Carnivora. There are two families within the Creodonta, Oxyaenidae and Hyaenodontidae, but only one genus of the latter is known from the White River Group. Members of the order range in body size from around a small cat to a hyena or lion. In addition to being active predators, like modern carnivores, they probably also scavenged and ate carrion.
Compared to modern carnivores with a similar body size, the size of the brain in creodonts was smaller and occupies a lower position on the skull. Another distinctive feature is a large, prominent sagittal crest and a large temporal fossa, suggesting they possessed powerful temporalis muscles for closing the jaw. Unlike modern carnivores, the auditory bulla is unossified in most creodonts. There is a well-defined postorbital constriction in the skull that distinctly separates the rostrum from the braincase. As in modern carnivores, the canines are large and pointed for holding and killing prey. Modern carnivores have one set of slicing teeth (carnassials), which are always formed by the same upper and lower tooth. In contrast, creodonts could have two or three pairs of carnassial teeth that could be formed by either the upper first molar and lower second molar or the upper second molar and lower third molar. These molars were located farther back on the skull than in modern carnivores. The body skeleton is generalized, but some forms were plantigrade and some digitigrade, like modern carnivores. Unlike modern carnivores with a fused scaphoid and lunar, all of the carpals in creodonts are separate. Their claws had a prominent groove down their midline (fissured), a feature not present in modern carnivores. The femur has a distinct third trochanter, a feature not present in modern carnivores.
The Hyaenodontidae first appear in the Wasatchian and became extinct at the end of the early early Arikareean. They are characterized by having sectorial molars (modified for slicing) and highly specialized carnassials. The bones of the limbs and feet of members of the family indicate they were often cursorial.
Systematics and Evolution The type species for Hyaenodon is Didelphis (H.) leptorhynchus from the Oligocene of France. Currently 10 species are recognized in North America and range in age from the Duchesnean to earliest Arikareean (Gunnell, 1998).
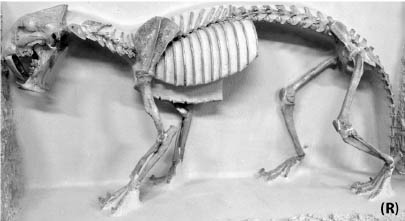
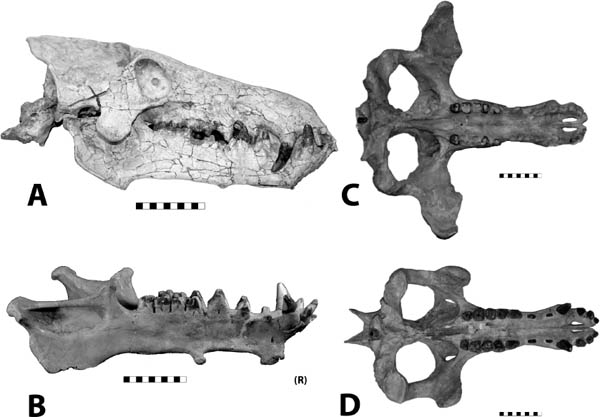
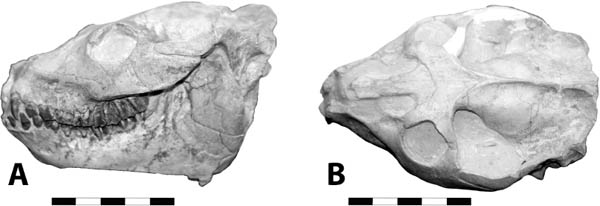
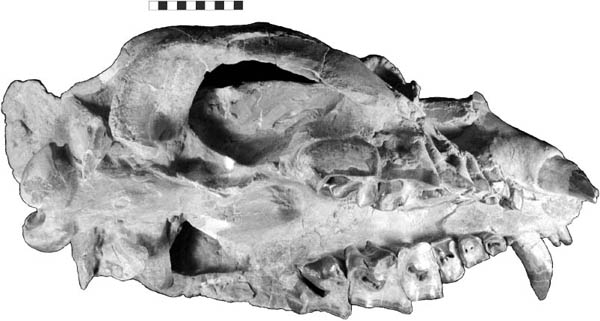
Distinctive Characters There is a great size range for species of Hyaenodon; an estimated body mass of an adult or subadult H. horridus, the largest North American species, is about 40 kg (Egi, 2001). In Hyaenodon both the upper first and second molars and lower second and third molars are modified as carnassials and were well developed for shearing meat. The feet are digitigrade, indicating it was cursorial and probably ran down its prey (Fig. 5.19A, B).
Stratigraphic and Geographic Distribution The earliest record of the genus is the early Duchesnean, but there are some indications that it may have appeared earlier in the late Uintan. It became extinct at the end of the early early Arikareean. Hyaenodon from the White River is represented by H. horridus, H. montanus, and H. crucians. Hyaenodon horridus is present in all members of the Chadron Formation, and a second species, H. crucians, is reported from the Peanut Peak Member. Hyaenodon brevirostris is reported from the Poleslide Member of the Brule Formation. Hyaenodon was the last surviving creodont in North America. The genus was widely distributed in North America and is known from South Dakota, California, Utah, Wyoming, Texas, Nebraska, Montana, New Mexico, Colorado, North Dakota, and Saskatchewan.
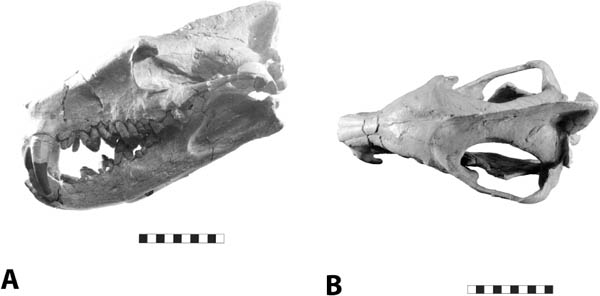
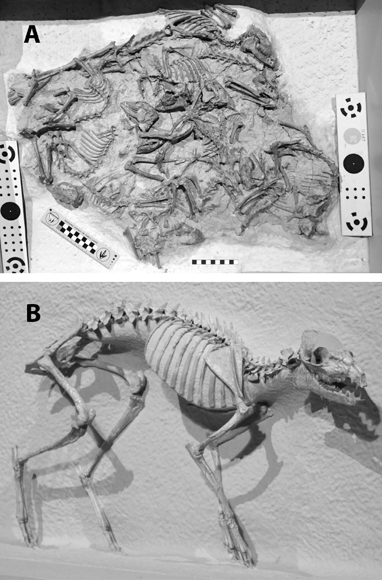
5.19. Hyaenodon horridus. (A) Skull and jaws, left lateral view, SDSM 242. (B) SDSM 242, skull, dorsal view. Scales in centimeters. Photos by the authors of specimens from the Museum of Geology, South Dakota School of Mines and Technology, Rapid City, South Dakota, U.S.A.
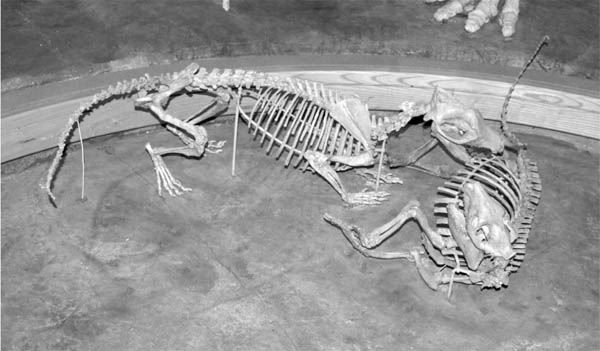
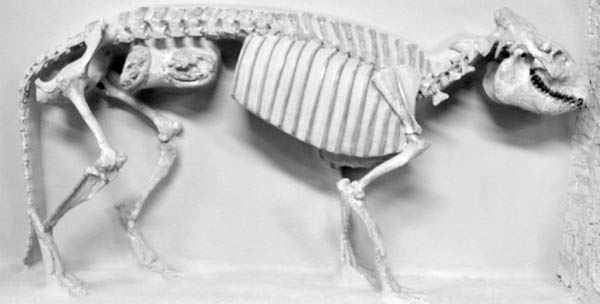
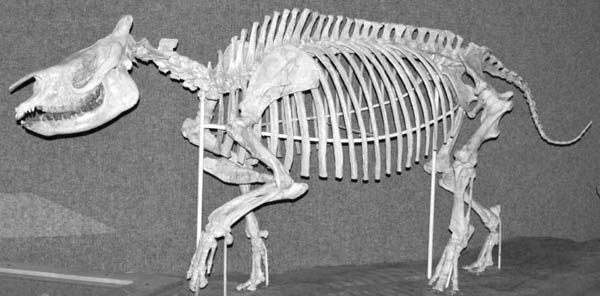
Natural History and Paleoecology The body size of Hyaenodon horridus is small compared to other species of the genus, and it may have preyed on any of the small artiodactyls present in the White River fauna, like Leptomeryx, Hypertragulus, or Hypisodus. A mount at the Denver Museum of Nature and Science has Hyaenodon killing a small individual of Merycoidodon (Fig. 5.20). The extinction of the creodonts is attributed to competition with members of the modern order Carnivora.
While the name of the order is indicative of the primary habits of its members as carnivores, this is not true of every member of the order, and various groups have adapted to quite different dietary preferences. Members of the Ursidae and Procyonidae tend to be omnivorous, and some bears, such as the panda and South American spectacled bear, are strictly herbivores. Likewise, members of the Canidae and Mustelidae will include varying amounts of both animal (including fish) and vegetable material in their diets. The Felidae are the most carnivorous members of the order and are often referred to as hypercarnivores. Carnivores may be both ambush or pursuit predators, and this is reflected in modifications of the skeleton that aided in running down prey, such as the cheetah or wolves. Some mustelids like martens and fishers are fully arboreal. Other carnivores may obtain food from aquatic environments and may be semiaquatic, like otters, or fully aquatic, like seals and sea lions.
All members of the order have large, well-developed canines that aid in catching and holding prey during hunting. Although reduced in the more omnivorous or herbivorous forms, members of the modern order Carnivora are characterized by having the shearing teeth, or carnassials, always being formed by the upper fourth premolar and lower first molar, unlike the creodonts, in which the position of the carnassials were further back in the skull and jaw, and could be formed by either first upper and second lower molars, or the second upper and third lower molars.
Another distinctive feature of the Order Carnivora is the fusion of two carpal bones, scaphoid and lunar, into a composite structure, the scapholunar. Claws are present in all forms and in some groups can be retracted, a common character of the Felidae, but also present in some other families as well. The manus and pes may be plantigrade or digitigrade, and while the first digit on either the manus or pes may be reduced or lost in some forms, the number of digits is never less than four. While the amount may vary, the radius remains mobile and able to rotate around the ulna to aid in the maneuverability of the forelimb while catching prey.
While members of the carnivore family Nimravidae superficially resemble modern cats (family Felidae), they represent a distinct family of carnivores. They first appear in Europe and North America in the late Eocene (early Chadronian in North America), but the family became extinct in the early Miocene (Clarendonian) and did not have any descendants. The general resemblance to true cats is based on the short face, but they retain a short braincase unlike the elongated braincase of members of the Felidae. Like true cats, the carnassials are highly modified for cutting meat, the upper first molar is reduced in size, and they have lost the upper second and third molars. Most of the major characters that distinguish members of the Nimravidae from the Felidae are in the anatomy of the braincase and the ear region, such as an incomplete auditory bulla. Some but not all nimravids developed a prominent elongation of the mental process of the mandible to form a “sheath” for the elongated upper canine.
5.20. Reconstruction of the creodont, Hyaenodon, attacking the oreodont, Merycoidodon. Hyaenodon crucians (DMNS 1604) and Merycoidodon culbertsoni composite skeleton (DMNS 84 and DMNS 54). Photo by the authors. Denver Museum of Nature and Science, Denver, Colorado, U.S.A. All rights reserved.
Primitive members of the family were plantigrade, although there is some evidence that advanced members of the family had become digitigrade, like modern felids. Another feature they shared with modern cats was retractable claws with a well-developed sheath housing the base of the nail. Their postcranial skeleton suggests they were capable of climbing trees, although Dinictis and Nimravus appear to have more cursorial habits than other nimravids. The scapula has a distinct process for the teres major muscle, suggesting the forearms could be retracted with force.
Except for a later genus, Dinaelurus, which is found only at John Day, Oregon, which has conical canines, all other nimravids have enlarged bladelike upper canines and were functional saber-tooths. The elongated (saber-tooth) upper canine morphology can be divided into the dirk-toothed forms with fine serrations (Hoplophoneus and Eusmilus) and a scimitar-toothed group (Dinictis and Nimravus) in which the serrations are coarse. The canines had serrations on both their mesial and distal edges. The coarseness of the serrations seems to be correlated with locomotor habits, with the dirk-toothed forms being short legged and plantigrade, so were more similar to bears in their body proportions, while the scimitar-toothed forms were more cursorial, like modern felids, in their postcranial anatomy. The dirk-toothed forms are thought to have been ambush predators, while the scimitar-toothed forms were probably capable of chasing their prey over short distances (Martin, 1998). It is interesting that the White River fauna contained five genera of nimravids with saber-toothed adaptations, although it is only during the Whitneyan that all five genera coexisted. It is possible that Eusmilus replaced Hoplophoneus as a dirk-toothed form and Nimravus, a scimitar-toothed form, replaced Dinictis. Both Eusmilus and Nimravus appear at the beginning of the Whitneyan, and Hoplophoneus and Dinictis had become extinct by the end of the Whitneyan. It is possible that these new genera may have competed with and ecologically replaced the older genera.
Systematics and Evolution The type species of Dinictis is D. felina and is based on a poorly preserved skull and jaw from the White River of the Dakota Territory. The genus is monotypic, and only one species is recognized as valid. Disappearing at the end of the Whitneyan, Dinictis may have been the ancestor to Pogonodon, which appeared in the Orellan.
Distinctive Characters Dinictis is a dirk-toothed nimravid, while Hoplophoneus is a scimitar-toothed form, suggesting they occupied different ecological niches as predators and probably focused on different prey species. The upper canine is shorter and broader than in Hoplophoneus. The occiput of Dinictis is inclined while in Hoplophoneus it is vertical (Martin, 1998).
Stratigraphic and Geographic Distribution Dinictis, along with Hoplophoneus, are the two earliest genera of nimravids in North America, and both first appear in the early Chadronian. In the Big Badlands of South Dakota, Dinictis is present in the Peanut Peak Member of the Chadron Formation and the Poleslide Member of the Brule Formation. It is also found in faunas in Wyoming, Colorado, Nebraska, and Saskatchewan.
Natural History and Paleoecology Nimravids appear in North America in the late Eocene, at about the same time or perhaps a little later than their appearance in Eurasia. While the first North American forms are already represented by cursorial forms, some of the early nimravids have features of the skeleton that suggest arboreal adaptations. Dinictis is more cursorially adapted than its contemporary, Hoplophoneus, a shorter-limbed form, which was probably, in contrast, a solitary ambush predator. Except for the brontotheres, most of the available prey species were small, and because the late Eocene–early Oligocene fauna was dominated by oreodonts, small horses, and small rhinocerotids, these were the most likely prey for Dinictis and other nimravids (Martin, 1998).
Systematics and Evolution The type species of Pogonodon is P. platycopis from the John Day Formation, Oregon, and the genus includes two other species, P. eileenae and P. paucidens. Species now referred to Pogonodon have at times been placed in Dinictis, but the two genera seem to have been distinct lineages since the early Oligocene (Orellan) (Martin, 1998).
Distinctive Characters Pogonodon is a large nimravid in the size range of a mountain lion to jaguar. Like Dinictis, it is a scimitar-toothed form, but the upper canine is short, broad, and coarsely serrated. The skull has a high sagittal crest, and there is a distinct flange on the mandible. Pogonodon retains a lower second premolar, which is absent in Hoplophoneus but has lost the lower second molar, which is retained in most specimens of Dinictis (Bryant, 1996). A distinctive feature of the pes is the presence of an articulation between the calcaneum and the navicular.
Stratigraphic and Geographic Distribution The biostratigraphic range of the genus is from the Orellan to the late early Arikareean. Pogonodon platycopis is the only species present in the White River Group in South Dakota and is found in the Brule Formation (Orellan to Whitneyan) (Martin, 1998). The species is also known from Wyoming, Montana, and Oregon.
Natural History and Paleoecology The extinction of Pogonodon coincides with that of Eusmilus and Nimravus at the end of the late early Arikareean. Although Pogonodon appears first in the Orellan and the other two genera first appear in the Whitneyan, their biostratigraphic ranges essentially overlap. This would suggest that they may have been ecological competitors for most of their existence because they overlapped not only in time but also in geography and have been found together in the same faunas. Martin (1998) noted that the late Eocene–Oligocene fauna was dominated by oreodonts and small rhinocerotoids, and this prey diversity may have permitted the coexistence of these three genera, perhaps ecologically equivalent to that of lions, leopards, and cheetahs in Africa today, although none of these genera have the advanced cursorial adaptations seen in cheetahs.
Systematics and Evolution The type species of Hoplophoneus is Machairodus primaevus from the Oreodon Zone of the White River Group in Colorado. Eleven species have been described for Hoplophoneus – more than for any other nimravid. Currently three other valid species besides the type are recognized.
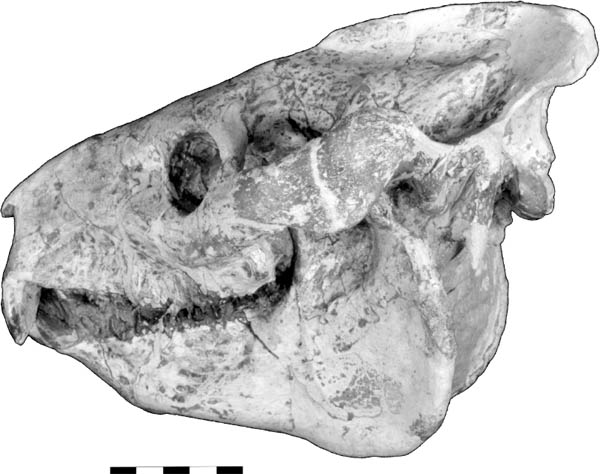
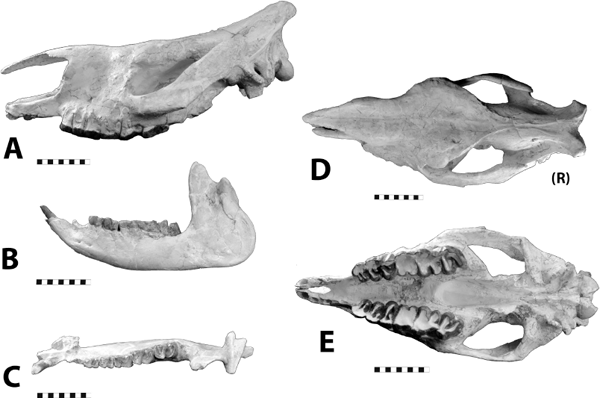
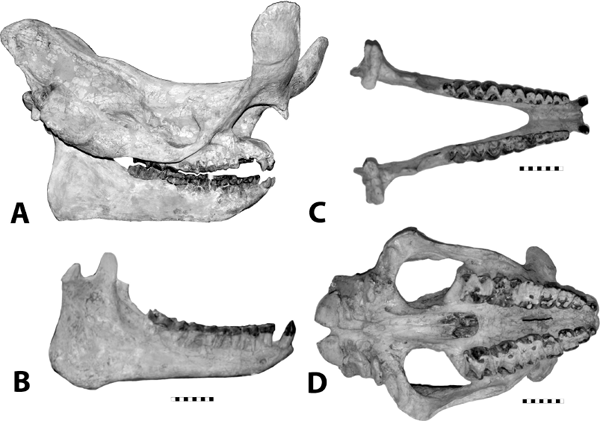
Distinctive Characters It is a dirk-toothed form with fine serrations on the upper canine, and the mandibular flange is larger and more prominent than in Dinictis. Hoplophoneus also has a prominent sagittal crest, and the occiput is vertical (Fig. 5.21A). The mastoid process is enlarged, suggesting powerful neck musculature. Like other saber-toothed cats, the deciduous canine was retained until the permanent canine was fully erupted. The permanent canine erupted along the inner edge of the deciduous canine (Fig. 5.21D, E). In the lower jaw it has only two premolars and one molar, while in Dinictis the lower dentition consists of three premolars and two molars (Fig. 5.21C). The mandible has a large flange (Fig. 5.21B) (Martin, 1998).
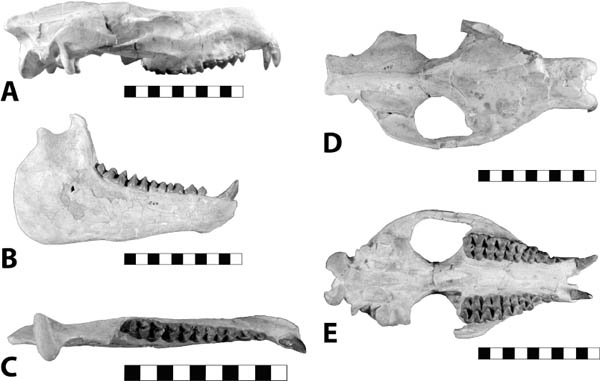
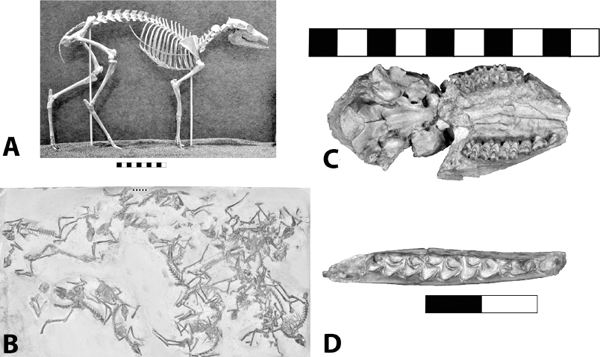
5.21. Hoplophoneus primaevus. (A) Skull, left lateral view, SDSM 2544. (B) Mandibles, right lateral view, reversed, SDSM 2544. (C) Mandibles, occlusal view, SDSM 2544. (D) Hoplophoneus sp. skull, anterior end, occlusal view with arrow marking permanent canines beginning to erupt, reversed, UCM 19160. (E) Hoplophoneus sp., skull, occlusal view, reversed, UCM 19160. Scales in centimeters. Photos by the authors. SDSM 2544 is from the Museum of Geology, South Dakota School of Mines and Technology, Rapid City, South Dakota, U.S.A. UCM 19160 is from the University of Colorado Museum of Natural History, Boulder, Colorado, U.S.A.
Stratigraphic and Geographic Distribution The stratigraphic range for the genus is from the Chadronian through Whitneyan. Hoplophoneus mentalis is present in the Crazy Johnson and Peanut Peak members of the Chadron Formation. Hoplophoneus primaevus and H. occidentalis are present in the Brule Formation. The genus is also found in Wyoming, Nebraska, and Montana.
Natural History and Paleoecology Hoplophoneus was a contemporary of Dinictis, and both genera first appear in the early Chadronian and became extinct at the end of the Whitneyan. Hoplophoneus was larger than Dinictis, with a size ranging from similar to a mountain lion (H. primaevus) to that of a jaguar (H. occidentalis) (Fig. 5.22).
On May 30, 2010, a Hoplophoneus skull was discovered in Badlands National Park by a 7-year-old girl while participating in a park-led Junior Ranger program. The skull was preserved in a carbonate-cemented mudstone bed in the middle to upper Scenic Member of the Brule Formation. The cranium is exceptionally well preserved and shows unique punctures, possibly bite marks from another nimravid. Badlands National Park, in cooperation with Rapid City Regional Hospital, scanned the skull by computed tomography (CT). The CT scan provided evidence that another Hoplophoneus attacked this individual (Fig. 8.1).
5.22. Hoplophoneus primaevus, SDSM 2528, skeleton (reversed), right lateral view. Photo by the authors of specimen from the Museum of Geology, South Dakota School of Mines and Technology, Rapid City, South Dakota, U.S.A.
Systematics and Evolution The type species of Eusmilus is E. bidentatus from the fissure deposits of Quercy, France, are considered to be Oligocene in age. Another genus from South Dakota, Egmoiteptecela, is considered to be a junior synonym of Eusmilus (Martin, 1998). Egmoiteptecela is from the Lakota Egmo, “cat”; ite, “face”; and ptecela, “short.” The name thus literally means “short-faced cat.”
Distinctive Characters Eusmilus is the smallest of the nimravids in North America, and unlike other nimravids, there does not appear to have been an increase in size through time. It is a dirk-toothed form with its cheek teeth greatly reduced in number, with two upper premolars and one to two lower premolars and a single upper and lower molar. Of all the nimravids found in the White River, its mandible has the greatest development of the flange in proportion to the animal’s size (Martin, 1998).
Stratigraphic and Geographic Distribution Eusmilus first appears at the beginning of the Whitneyan and became extinct at the end of the late early Arikareean at the Oligocene–Miocene transition. Eusmilus sp. has been identified from the Poleslide Member of the Brule Formation in the Big Badlands. The genus is also known from Nebraska and Oregon.
Natural History and Paleoecology All members of the tribe Eusmilini are small (Bryant, 1984), including Eusmilus, which is about the size of a modern bobcat, making it the smallest of all the saber-toothed predators present in the White River faunas (Bryant, 1984; Martin, 1992). Emerson and Radinsky (1980) estimated the body mass of Eusmilus at about 33.8 kg. Presumably this size difference allowed Eusmilus to avoid competition with other contemporary saber-tooths by preying on smaller species. Despite its smaller body size, the cranial morphology of Eusmilus was in many respects as derived as that of the later Barbourofelis, including the enlargement of the carnassials and reduction of the premolars (Martin, 1992). The presence of Eusmilus in contemporary faunas in both North America and Europe suggest a widespread Holarctic distribution for the genus during the Oligocene. Martin (1992) proposed that although the smaller species of oreodonts may have been the principal prey for Eusmilus, given its cranial specializations, it may have been able to have taken larger prey – up to the size of modern pigs or deer. In contrast, Emerson and Radinsky (1980) pointed out that the relationship between body size of predator and prey might be modified if a smaller predator such as Eusmilus preyed on different age classes, i.e., younger and hence smaller individuals of a given prey species, or if it was hunting socially and was thus able to take down larger individuals. We do not yet have any information regarding Eusmilus as to whether either or both of these alternatives are applicable. Van Valkenburgh (1985) concluded, on the basis of limb morphology, that although all members of the predatory guild in the early Oligocene were slower and more robust than their modern counterparts, the spacing of predator types was similar to that seen in modern faunas, and that the ecological processes that structured the guild were not appreciatively different than those that structure the guild of predators in modern ecosystems.
Systematics and Evolution The type species of Nimravus is N. brachyops from the John Day Formation, Arikareean of John Day, Oregon. Currently there are five species considered to be valid.
Distinctive Characters This was a scimitar-toothed form comparable in size to a leopard. The upper canine is coarsely serrated. The protocone is reduced on the upper fourth premolar, and the lower first molar has a distinct talonid. The second and third lower molars may be present in some individuals. Unlike Hoplophoneus, it did not develop an enlarged flange on the mandible. Its jaw is also distinguished from the other nimravids by having an enlarged masseteric fossa that extends anteriorly to under the lower carnassial (Martin, 1998).
Stratigraphic and Geographic Distribution The genus first appears in the Whitneyan and becomes extinct at the end of the late early Arikareean, at the Oligocene–Miocene boundary. Nimravus brachyops is present in the Poleslide Member of the Brule Formation in the Big Badlands. The genus is also known from California, Nebraska, North Dakota, and Oregon.
Like Eusmilus, Nimravus first appears in the Whitneyan and becomes extinct at the Oligocene–Miocene boundary, although there are some fragmentary specimens that suggest this genus may have persisted into the early late Arikareean, so Nimravus may have survived into the earliest Miocene.
Natural History and Paleoecology Intraspecific fights between the different saber-toothed taxa in the White River seem to have taken place. A skull of Nimravus in the Museum of Geology at the South Dakota School of Mines and Technology has an elongate puncture in the left frontal sinus that had partially healed. The size of the puncture is similar in size to the canine of Eusmilus (Scott and Jepsen, 1936).
The Amphicyonidae first appear in North America in the late Eocene (Duchesnean) and became extinct in the early late Miocene (Clarendonian), although they survived later in Europe and India. In North America, their greatest diversity was in the Miocene, and there are three genera in the White River: Daphoenus, Paradaphoenus, and Brachyrhynchocyon.
The general skull form is a short rostrum with an elongated postorbital region. As some members of the family increased in size, the skull became disproportionately enlarged, including a greatly enlarged area for the temporalis muscle and an extreme development of the sagittal crest. Like the nimravids, a major feature that distinguishes amphicyonids from other carnivores is the construction of the ear region, with the ectotympanic bone as the primary bone contributing to the formation to the bulla, which was not inflated and thus is somewhat similar to what is seen in bears. Most amphicyonids retain the basic eutherian tooth formula of I3/3-C1/1-P4/4-M3/3, and only in a few derived forms are premolars lost or the upper third molar, which happens independently in some genera. The incisors and canines tend to be large. In some lineages, the molars tend to decrease in size from the first to third, and in many the third molar is eventually lost; in other lineages, the posterior molars becomes the largest and with their enlargement become broad, flat, crushing surfaces, especially in the second molar, so they look like the molars of bears, giving rise to the popular name for the group, bear-dogs. Larger amphicyonids are plantigrade; smaller forms like those from the White River are digitigrade, but not as advanced in cursorial abilities as dogs. Digitigrade amphicyonids like Daphoenus retain slicing carnassials, while the larger plantigrade forms tend to lose the shearing capacity of the carnassials and have broader molars, thus more closely resembling bears in two ways. There is pronounced sexual dimorphism in amphicyonids. The males had a baculum, and a significant difference is size has been documented in Brachyrhynchocyon dodgei from the late Eocene (Chadronian).
5.23. Daphoenus vetus. (A) Skull and jaws, right lateral view, YPM PU 12651. (B) Skull, occlusal view, YPM PU 12648. Scales in centimeters. Photos by the authors. Courtesy of the Division of Vertebrate Paleontology, YPM PU 12651 and YPM PU 12648, Peabody Museum of Natural History, Yale University, New Haven, Connecticut, U.S.A.
Systematics and Evolution The type species for Daphoenus is D. vetus from South Dakota, but the exact locality is unknown. The type was collected in the White River beds of the Dakota Badlands and was described by Leidy in 1869 as Amphicyon vetus. It is one of the earliest finds of a White River mammal. Currently there are five described species.
Distinctive Characters Species of Daphoenus range in size from small- to midsize within the amphicyonids. The skull form and the canine teeth are dimorphic, with specimens interpreted as males larger with a more robust rostrum and canines. The skull is more elongate then the short, broad skull of Brachyrhynchocyon, but in both genera, the premolars increase in size from the front to the back. It retains the primitive eutherian dental formula with mesiodistally elongate and narrow premolars, especially the upper and lower second and third premolars (Fig. 5.23).
Stratigraphic and Geographic Distribution The genus first appears in the late middle Eocene (Duchesnean) and becomes extinct in the late Oligocene (late early Arikareean). It is the most common genus within the family, and for the Great Plains, essentially all known specimens are from the White River Group of South Dakota, Nebraska, Wyoming, and Colorado. Daphoenus vetus is the only species identified from the Big Badlands and is found in the Crazy Johnson and Peanut Peak members of the Chadron Formation and throughout the Brule Formation (Hunt, 1998b). The genus is also found in Texas, California, Wyoming, Oregon, Nebraska, Colorado, Montana, North Dakota, and Saskatchewan.
Systematics and Evolution The type species is P. cuspigerus from the John Day Formation of Oregon, and a second species, P. minimus, is recognized both on the Great Plains and at John Day.
Distinctive Characters This genus is a small amphicyonid, with an estimated body mass between 4 and 6 kg (Figueirido et al., 2011); the general form of the premolars is similar to those of Daphoenus, except for their smaller size. While the sample size is small, the lower premolars tend to be laterally compressed and bladelike. Paradaphoenus cuspigerus is the oldest and most primitive species and can be distinguished from contemporary amphicyonids such as Daphoenus by the closed trigonid with a slightly developed paraconid. The upper first molar has a V-shaped protocone. The talonid of the lower first molar is basined with the hypoconid and entoconid, creating a low ridge that forms the border of the basin. The genus is currently only known from two geographic areas: the Great Plains and the John Day Basin. The two populations are distinguished by the presence of a small upper third molar in the John Day sample that appears to be absent in the Great Plains sample. The genus lacks an enlarged or inflated auditory bulla, so the basioccipital is wide (Hunt, 2001).
Stratigraphic and Geographic Distribution The genus first appears in the Orellan and became extinct in the late early Arikareean. Paradaphoenus cuspigerus is present in the Scenic Member of the Brule Formation in South Dakota and it is also known from the Brule Formation in Nebraska (Hunt, 1998b).
Natural History and Paleoecology There is little evolutionary change in body size or skull proportions of the genus from the Orellan to the Arikareean, including the advanced form P. cuspigerus from John Day. The genus seems to be strictly North American. The rarity of specimens prevents any real determination of its probable diet.
Systematics and Evolution The type species of Brachyrhynchocyon is B. dodgei from the Seaman Hills of Wyoming, and a second species, B. montanus, is also recognized.
Distinctive Characters It is a small form but highly sexually dimorphic, with males having a broad, massive skull while in females the skull is narrower and more gracile (Gustafson, 1986). The skull is short and broad, thus giving rise to its name, Brachyrhynchocyon, “short-rostrum dog.” The genus retains the full primitive eutherian dental formula. It can be distinguished from its contemporary in the White River Group, Daphoenus, in having mesiodistally short, wide, and robust premolars. In both forms, the premolars increase in size posteriorly and will develop accessory cusps on the posterior margins of the second, third, and fourth premolars. The mandible is thick and massive (Hunt, 1996).
Stratigraphic and Geographic Distribution The genus is known only from the late Eocene (Chadronian). In the Big Badlands, it is found in all members of the Chadron Formation (Hunt, 1998b). Outside of South Dakota, it is found in Texas, Wyoming, Colorado, Nebraska, Montana, and Saskatchewan.
Natural History and Paleoecology A few postcranial skeletons are known for Brachyrhynchocyon. They indicate a robust, muscular, wolverinelike build to the body that, coupled with the shortened muzzle, which is also wolverinelike, suggests it may have filled a similar niche.
The Canidae includes the living dogs such as wolves, coyotes, jackals, and foxes. The distribution of the family today is widespread and includes North and South America, Eurasia, and Africa, with the human introduction of the dingo into Australia. The early fossil record and evolutionary history of the family is in North America, and they did not disperse into other continents until the late Miocene (Hemphillian). The family first appears in the late middle Eocene (Duchesnean). Three subfamilies are recognized: Hesperocyoninae, Borophaginae, and Caninae. The first two are extinct (Wang and Tedford, 2008).
Members of the Canidae are generally considered the most primitive or generalized of all the carnivores. As is the case with most carnivores, one of the primary characters that distinguishes the family is the ear region. Canids have a fully ossified and inflated auditory bulla formed primarily by the entotympanic and a smaller contribution from the ectotympanic. The overall shape of the skull is elongate, with a prominent tapering rostrum and strong zygomatic arches. The temporal fossae are well defined by either a prominent sagittal crest, generally better developed in males, or paired temporal cristae. The dental formula is similar to that of primitive eutherian mammals except that the upper third molar has been lost. Most canids are mesocarnivores with an omnivorous diet, although in the fossil record there are forms that were hypercarnivores, feeding almost exclusively on meat, as well as hypocarnivorous forms where meat is only a minor component of their diet.
5.24. Hesperocyon sp., SDSM 4385. (A) Skull, right lateral view. (B) Skeleton, right view, SDSM 2513 Hesperocyon gregarious. (C) Skull, dorsal view, SDSM 4385, Hesperocyon sp. (D) Skull, occlusal view, SDSM 4385, Hesperocyon sp. Scales in centimeters. Photos by the authors of specimens from the Museum of Geology, South Dakota School of Mines and Technology, Rapid City, South Dakota, U.S.A.
The extinct subfamily Hesperocyoninae is considered the most primitive of the canids. Although members of the subfamily have the canid type of auditory bulla, there are some genera that lack a bony tubular external auditory meatus. Recent analysis of this subfamily (Wang, 1994) did not support a monophyly for the subfamily, so even though it is still used, it is most likely that it is a polyphyletic group with multiple clades that only share primitive characters of the family.
Systematics and Evolution The type of Hesperocyon is H. gregarius. The locality given by Cope (1873a:3) in the type description is “abundant in the Oreodon beds of the Miocene formation of Colorado.” This has subsequently been interpreted as a reference to the Orellan rocks of the White River Formation of northeastern Colorado. Two other species are presently included in the genus, “H.” coloradensis and “H.” pavidus (Wang, 1994).
When Cope (1873) described the type of Hesperocyon, he originally placed it in the genus Canis as C. gregarius. The species was placed in the new genus Hesperocyon by Scott (1890). Cope did not designate a type specimen but merely stated that there were abundant specimens of Canis gregarius in his collections. Other species traditionally included in the genus were subsequently transferred to other genera by Wang (1994). Hesperocyon temnodon is now in Mesocyon and H. wilsoni in Prohesperocyon. The distinctions between the three remaining genera are often made on the basis of size and minor differences in morphology. Given how common sexual dimorphism is in the canids, it may be that this, as well as stage of ontogenetic development, may account for the features used to distinguish the different species.
Distinctive Characters Hesperocyon is a small animal with an elongate body and long tail. The manus is subdigitigrade, and it appears to have had retractable claws, a character not present in most canids. The protocone of the upper fourth premolar (carnassial) is reduced in size and medially positioned on the tooth. The lower carnassial (first molar) has an elongated shearing blade. Fossil bacula have been found with some articulated specimens. Fortunately a number of articulated skeletons of Hesperocyon, primarily H. gregarius, have been found, and mounted skeletons of the species are on display in museums (Fig. 5.24).
Stratigraphic and Geographic Distribution Hesperocyon gregarius ranges from the middle Duchesnean to the end of the Whitneyan, “H.” parvidus ranges from the beginning of the Orellan into the mid-Arikareean, and “H.” coloradensis is restricted to the Orellan. Hesperocyon gregarius is present in the Peanut Peak Member of the Chadron and both members of the Brule formations; a second species, “H.” parvidus, has been reported from the Poleslide Member of the Brule Formation. The genus is also known from California, Wyoming, Colorado, Nebraska, Montana, North Dakota, Saskatchewan, and Oregon, as well as in the southeastern United States in Georgia.
Natural History and Paleoecology Hesperocyon gregarius has the best fossil record of any member of the subfamily because of the large number of specimens, including articulated skeletons, from the White River Group. The cooccurrence of Hesperocyon and eomyid rodents has been interpreted by past workers as indicative of a predator–prey relationship (Cope, 1874; Clark, Beerbower, and Kietzke, 1967). Although coprolites are common in the White River, none can be attributed unequivocally to Hesperocyon to confirm what comprised its primary prey. In contrast, a premolar of Hesperocyon was identified in a coprolite along with snake remains that would have been produced by a larger predator in the ecosystem (Parris and Holman, 1978), and LaGarry (2004) reported remains of Hesperocyon in coprolites from the Orella Member of the Brule Formation in northwestern Nebraska, indicating it was prey as well as predator. Clark, Beerbower, and Kietzke (1967) proposed that the preferred habitat for Hesperocyon was the river-border forest environment.
Systematics and Evolution The type species is M. coryphaeus from John Day Formation in Oregon. Traditionally the genus has included 10 species, but Wang (1994) transferred three species to Parenhydrocyon and one to Cynodesmus, restricting Mesocyon to three species: the type, M. brachyops, and “M.” temnodon. The genus as defined by Wang may be paraphyletic because the three species are linked by shared primitive characteristics and not by any apomorphies.
Distinctive Characters Mesocyon is a primitive midsize canid. The genus is distinguished from Hesperocyon and Parenhydrocyon by having a long, robust, ventrally directed paroccipital process. Primitive characters shared by the three species in the genus include a wide upper second molar and the absence of robust premolars. In the derived species there is a round fossa on the supraoccipital (Munthe, 1998).
Stratigraphic and Geographic Distribution The genus is restricted to the Oligocene, appearing in the Orellan and becoming extinct in the late early Arikareean, with some tentative records reported from the early Miocene (late Arikareean). The genus has been found in faunas in South Dakota, California, Oregon, Wyoming, Colorado, Nebraska, and North Dakota.
Natural History and Paleoecology Given the taxonomic uncertainty of this genus, nothing has been published on its paleoecology.
Systematics and Evolution The type species of Ectopocynus is E. simplicidens from the Runningwater (formerly Marsland) Formation of western Nebraska, Middle Hemingfordian, early Miocene. The genus includes two other species, E. antiguus and E. intermedius. It should be noted that while three species are recognized, they are represented by a total of six specimens (Munthe, 1998), so a better understanding of the taxonomy of this genus and its relationships to other canids cannot be fully evaluated until more specimens become available. Wang (1994) did note some dental similarities between Ectopocynus simplicidens and Hesperocyon gregarius.
Distinctive Characters The lower premolars are short, blunt, and robust, and the accessory and cingular cusps are extremely reduced or absent. The lower first premolar is absent. The metaconid on the lower first and second molars is also reduced.
Stratigraphic and Geographic Distribution The genus appears in the Whitneyan and became extinct at the end of the early Hemingfordian. Ectopocynus antiquus is known from the Poleslide Member of the Brule Formation (Whitneyan) in the Big Badlands. It is also known from Wyoming.
Natural History and Paleoecology This is a poorly known taxon represented by only six specimens, thus limiting inferences about both phylogenetic relationships and paleoecology.
5.25. Sunkahetanka geringensis, YPM PU 13602. Right mandible, left lateral view. Scale in centimeters. Photo by the authors. Courtesy of the Division of Vertebrate Paleontology, YPM PU 13602, Peabody Museum of Natural History, Yale University, New Haven, Connecticut, U.S.A.
Systematics and Evolution The type species of Cynodesmus is C. thooides from the Deep Creek Beds of Montana, considered to be early Arikareean in age. In the past the genus has included species that are now placed in all three subfamilies of the Canidae. Over 14 species have been described for this genus, but currently only the type species and C. martini are considered valid (Wang, 1994).
Distinctive Characters As defined by Wang (1994), the genus can be distinguished from other members of the Hesperocyoninae by the strong paroccipital process on the skull with a posterior keel and a broadened upper second molar.
Stratigraphic and Geographic Distribution The stratigraphic range is from the Whitneyan to the end of the early Arikareean of western Nebraska and South Dakota, and the genus is present in the earliest Arikareean of western Montana. Specimens referred to the genotypic species are known from the Big Badlands, so all records of this taxon are Whitneyan in age. It is also known from Nebraska and Montana.
In Lakota, Sunka means “dog,” he means “tooth,” and tanka means “large.” Thus the name means “large-toothed dog.” Systematics and Evolution The type species of Sunkahetanka is S. geringensis (Barbour and Schultz, 1935) from the Gering Formation in Morrill County, Nebraska. The type specimen is USNM 1092, including the skull, mandible, and skeleton. There is only one species assigned to this genus. Sunkahetanka was proposed by Macdonald (1963:214) for Mesocyon geringensis as a “derivative of the Mesocyon line.” Wang (1994) considered Sunkahetanka as transitional between the more primitive Mesocyon and Cynodesmus and the more advanced Philotrox and Enhydrocyon.
Distinctive Characters Sunkahetanka is distinctive from Cynodesmus by its more massive imbricated premolars and the reduced metaconid on the lower molars. The skull is also heavier and the dentition more robust than Mesocyon or Cynodesmus (Fig. 5.25) (Wang, 1994).
Stratigraphic and Geographic Distribution Sunkahetanka is restricted to the topmost part of the Brule Formation in western Nebraska and to the Sharps Formation of western South Dakota (early Arikareean). At Badlands National Park, Sunkahetanka was found in Cedar Pass in the Sharps Formation (Wang, 1994).
5.26. Osbornodon sp., BADL 63382. (A) Skull, right lateral view. (B) Right mandible, right lateral view. (C) Skull, occlusal view. (D) Right mandible, occlusal view. Scales in centimeters. Photos by the authors of specimens from Badlands National Park. Specimens are the property of the U.S. government.
Systematics and Evolution The type species of Osbornodon is O. fricki from New Mexico and includes species that originally were placed in Cynodesmus, Mesocyon, Tomarctus, and Brachyrhynchocyon. Currently the genus includes five species.
Distinctive Characters Osbornodon sesnoni is a large canid. Many features of the skull of Osbornodon are similar to those seen in later members of the Caninae, with a long and slender rostrum, slender premolars, quadrate molars, and basined talonids on the lower molars (Fig. 5.26). It is distinguished from the other hesperocyonines by having more quadrate upper molars. The upper and lower second molars are enlarged, and the talonids of the lower first and second molars are basined (Wang, 1994).
Stratigraphic and Geographic Distribution As defined by Wang (1994), this genus has the longest stratigraphic range of the Hesperocyoninae, first appearing in the Orellan and becoming extinct at the end of the early Barstovian. Osbornodon sesnoni has been recovered from the Poleslide Member of the Brule Formation and is Whitneyan in age but is only known from a small number of specimens. Osbornodon renjiei occurs in the Poleslide Member of the Brule Formation. The genus is also known from faunas in Florida, Texas, Wyoming, Nebraska, and North Dakota.
The borophagine canids comprise one of the three subfamilies within the Canidae (Wang, Tedford, and Taylor, 1999). The subfamily is closely related to the subfamily Caninae, and the two are distinguished from the Hesperocyoninae by the presence of a basined, bicuspid talonid on the lower first molar. This feature seems to have allowed the borophagines to avoid competition with the hypercarnivorous hesperocyonines by allowing them to move into a hypocarnivorous niche soon after the two lineages diverged. Many of the early primitive borophagines had a dentition reminiscent of the dentition of members of the Procyonidae and were initially considered to be members of that family. Borophagines filled the ecological niches today filled by canids, procyonids, and hyaenids. Many genera of borophagines exploited the hypocarnivorous niche in the Tertiary, but later members became specialized as bone crushers.
The subfamily is known only from North America and first appears in the early Oligocene (Orellan) with the appearance of Otarocyon macdonaldi. The borophagines are the dominant canids in the late Tertiary until they became extinct in the Pliocene (Blancan). During this time interval they are among the best represented carnivorans present in a fauna. Among the different lineages of borophagines, the hypercarnivorous forms are better known than the hypocarnivorous, with many species of the latter known from only a handful of specimens. Although many of the genera have long, unbroken records during the Tertiary, there are taxa with significant gaps in their fossil record, such as the lack in the record of Otarocyon in the Whitneyan to connect the earlier O. macdonaldi (Orellan) and later O. cooki (early Arikareean). The eventual extinction of the borophagines is attributed to the appearance of felids, large mustelids, and giant ursids that came from Eurasia, first appearing in North America in the Hemphillian; they may have been ecological competitors. After the appearance of these immigrants, the only surviving borophagines are those adapted to a durophagous (bone-cracking) diet.
5.27. Archaeocyon leptodus, BADL 61910/SDSM 10501. (A) Skull and jaws, left lateral view. (B) Skull, dorsal view. Scales in centimeters. Photos by Xiaoming Wang of specimen housed at the Museum of Geology, South Dakota School of Mines and Technology, Rapid City, South Dakota, U.S.A. Specimen is the property of the U.S. government.
Systematics and Evolution The type species of Archaeocyon is A. pavidus, a species originally assigned to Pseudocynodictis from the Sespe Formation (late Whitneyan or early Arikareean), Ventura County, California. Two other species, A. leptodus and A. falkenbachi, are also recognized. Archaeocyon is a basal taxon near the separation of the Caninae–Borophaginae lineage from the hesperocyonines, as it has all of the characters that distinguish the two groups. Because of its basal position, Archaeocyon, like Oxetocyon and Otarocyon, occupies an ambiguous position in terms of its relationships to the other borophagines because it lacks derived characters of its own and in many ways more closely resembles Hesperocyon.
Distinctive Characters As a primitive member of the Caninae–Borophaginae lineage, Archaeocyon has the distinctive features of the group, including a weak parastyle and a lingual cingulum extending onto the anterior edge of the protocone and surrounding it on the on the upper first molar; the lower first and second molars have basined talonids. However, it lacks the derived characters seen in the Caninae, such as a slender, horizontal ramus of the mandible, narrow and elongate premolars separated by diastema, premolars with the posterior accessory cusps reduced or absent, and a reduced protocone on the upper fourth premolar. Compared to later borophagines, it is primitive in that the auditory bulla is not enlarged, instead having a posteriorly oriented paroccipital process that only rarely contacts the bulla; it has a posteriorly restricted hypocone on the upper first molar; and on the upper second molar, the lingual cingulum does not connect to the metaconule (Fig. 5.27) (Wang, Tedford, and Taylor, 1999).
Stratigraphic and Geographic Distribution Archaeocyon pavidus has been reported from the Poleslide Member of the Brule Formation (Whitneyan) in southwestern South Dakota (Jackson and Shannon counties), and A. leptodus is reported from the east side of Cedar Pass from the base of the Sharps Formation (early Arikareean), Jackson County, South Dakota (Wang, Tedford, and Taylor, 1999). The genus is known from the Whitneyan of South Dakota, Nebraska, and Wyoming; the late Whitneyan or early Arikareean of California; the early Arikareean (Sharps Formation) of South Dakota, North Dakota, Nebraska, Wyoming, Montana, and Oregon; and the medial Arikareean of Wyoming (Wang, Tedford, and Taylor, 1999).
5.28. Oxetocyon cuspidatus, BADL 11010/SDSM 7330. Left maxillary fragment with M1–M2. Photo by Edward Welsh with some modification of image of specimen from Badlands National Park. Specimen is housed at the Museum of Geology, South Dakota School of Mines and Technology, Rapid City, South Dakota, U.S.A. Specimen is the property of the U.S. government.
Natural History and Paleoecology The primitive dentition of Archaeocyon, which lacks specializations for slicing meat, suggests a tendency to hypocarnivory in contrast to the dentition seen in Hesperocyon, which is more hypercarnivorous. The diet of a hypocarnivore includes less than 30 percent meat. This suggests that Archaeocyon, like many early borophagines, was more omnivorous in their diets, perhaps consuming a larger percentage of vegetation in its diet, thus avoiding ecological competition with its contemporary, Hesperocyon.
Systematics and Evolution Oxetocyon is known from a single species, O. cuspidatus. The type was collected from the Poleslide Member of the Brule Formation in the Big Badlands. It represents the first of many genera within the subfamily that evolved hypocarnivory.
Distinctive Characters Oxetocyon was a small canid with low crowned (bunodont) teeth. The lingual cingulum of the upper first molar has three distinct cusps – thus the species name. The molar is enlarged and squared so that the tooth is symmetrical with a transverse cleavage in the middle of the crown (Fig. 5.28). The modification of the tooth into a more crushing mode for processing food suggests that vegetation may have been predominant in its diet.
Stratigraphic and Geographic Distribution It is only known from the Poleslide Member of the Brule Formation (Whitneyan) in the Big Badlands but has been reported from the Whitneyan and early Arikareean in Nebraska (Tanner, 1973; Wang, Tedford, and Taylor, 1999).
Natural History and Paleoecology Oxetocyon is one of the four earliest borophagine canids in North America found in the Big Badlands. Although the borophagines are often popularly characterized as the bone-crushing canids with hyaenalike dentition, this is only true for a few of the later genera like Borophagus. Remains of this genus are extremely rare, and little can be deduced regarding its phylogenetic relationships or paleoecology.
Systematics and Evolution The type species of Otarocyon is O. cooki, a species originally placed in Cynodesmus from the Sharps Formation (early Arikareean), Shannon County, South Dakota. A second species, O. macdonaldi, is also recognized. This genus appears abruptly in the Orellan without any apparent predecessor.
Distinctive Characters Compared to Archaeocyon, Otarocyon is a highly derived taxon that is defined by a number of anatomical features. The auditory bulla is hypertrophied and the rostrum is shortened. The braincase is broadened, with a short temporal fossa defined by a pair of temporal crests. The premolars are single cusped and short with tall crowns. The protocone on the upper fourth premolar is enlarged, and the upper first molar has an elevated cingulum (Wang, Tedford, and Taylor, 1999).
Stratigraphic and Geographic Distribution Otarocyon macdonaldi is known from the Scenic Member of the Brule Formation (Orellan), Pennington County, South Dakota, and from the Toston Formation (Orellan?) of Montana (Wang, Tedford, and Taylor, 1999). The genus is known from the Orellan of South Dakota and Montana, the early Arikareean of South Dakota, and the early to medial Arikareean of Wyoming.
Natural History and Paleoecology The closest living analog to Otarocyon is the living fennec fox, Vulpes zerda, which inhabits the deserts of North Africa and the Arabian Peninsula. Both are small canids and share a number of anatomical similarities, such as the expanded braincase, short nasal processes of the frontal, parasagittal temporal crests, and an enlarged auditory bulla. There is a hiatus of 25 Ma between the two taxa, and the fennec has many derived features that clearly place it with other living foxes, so the characters must have been independently derived. However, this remarkable convergence permits some inferences about the soft anatomy of Otarocyon. The enlarged bulla suggests that like fennecs, Otarocyon was sensitive to low-frequency sounds and probably had a large external ear. This implies that it may have lived in an open environment. The presence of at least a patchy network of open environments in the Orellan has been proposed by Retallack (1983b) on the basis of the paleosols.
Systematics and Evolution The type species of Cynarctoides is C. acridens, originally placed in the genus Cynarctus, from the Upper Harrison Beds (late Arikareean) near Agate, Sioux County, Nebraska. In addition to the type species, six other species are considered valid (Wang, Tedford, and Taylor, 1999). Cynarctoides, like many primitive hypocarnivorous borophagines, was originally thought to be a primitive procyonid. The multiple recognized species of the genus form a nearly continuous series of stages that clearly show an evolutionary trend in terms of the peculiar form of hypocarnivory in this genus. This includes retaining a primitive upper fourth premolar that lacks a hypocone and that never has a lingual cingulum. The lower molars tend to have distinctive oblique crests.
Distinctive Characters The many species of Cynarctoides are all distinguished by parasagittal crests and a slender, shallow horizontal ramus of the mandible. More derived species have a narrow rostrum, longer jaws with narrow and long premolars, and lower molars with conical and high-crowned cusps.
Stratigraphic and Geographic Distribution Cynarctoides lemur has been reported from the Poleslide Member of the Brule Formation in South Dakota and is the only possible Whitneyan record of the genus (Wang, Tedford, and Taylor, 1999). The genus is best known from the Arikareean and is clearly present in the early Arikareean (Sharps Formation) of South Dakota, Nebraska, and Oregon; the medial or late Arikareean of South Dakota and Florida; the late Arikareean of Colorado, Nebraska, Wyoming, and New Mexico; the early Hemingfordian of Nebraska, Idaho, Texas, and New Mexico; the late Hemingfordian of Nebraska, Wyoming, and New Mexico; and the early Barstovian of Nebraska, New Mexico, and California (Wang, Tedford, and Taylor, 1999).
Natural History and Paleoecology As a hypocarnivorous borophagine, the inference is that the genus probably had an omnivorous diet.
Bears have a long history in North America, first appearing in the late Eocene (Chadronian), and are represented in the modern fauna by grizzly (Ursus arctos) and black (Ursus americanus) bears. Three subfamilies are recognized, each representing a separate evolutionary radiation within the family. The fossil history of bears in North America represents repeated dispersal events from Eurasia, followed by extinction and a refilling of the bear niche by the next appearance of bears. Bears tend to have large canines and incisors, but they often reduce in size or lose the anterior premolars. The most distinctive feature of their dentition is the large quadrate molars that permit them to crush their food. Although the shearing ability of the carnassials in modern bears is greatly reduced, many of the fossil species have a more primitive dentition, including carnassials with well-developed shearing capability. Bears have a generalized postcranial skeleton including a greater ability to rotate the forearm than in other carnivores. They retain five digits on the manus and pes and have large, well-developed, nonretractable claws. The scapula has a prominent secondary spine along the posterior margin.
Systematics and Evolution The type of Parictis is P. primaevus, from the John Day Beds of Oregon, and seven species have been described. In the White River Group, these include P. dakotensis, P. major, P. parvus, and P. gilpini. This large number of species from a restricted period of time and geographic area suggests that the taxonomy of the genus needs to be reexamined; many of these species may become junior synonyms.
It is a member of the Amphicynodontinae, the most primitive subfamily of bears in North America, which includes two other genera, Allocyon and Kolponomos (only found in Alaska, Oregon, and Washington). Only Parictis is known from the White River Group and is the earliest representative of the Ursidae in North America
Distinctive Characters Parictis is a small animal, comparable in size to the contemporary dog, Hesperocyon. The dentitions of the two taxa are quite different, with the premolars of Parictis being more robust and transversely widened. Each of the premolars also has a distinctive cingulum (shelf) around the crown. The premolars lack anterior accessory cusps (Fig. 5.29). Post-Chadronian members of the family have more robust premolars that are transversely widened. Bones of the postcranial skeleton have not been described (Hunt, 1998b).
Stratigraphic and Geographic Distribution The genus first appears in the Chadronian and became extinct in the Orellan, with some specimens suggesting possible survival into the Whitneyan. In the White River Group, Parictis has been described from the Peanut Peak Member of the Chadron Formation and Scenic Member of the Brule Formation. It is also known from Wyoming, Nebraska, Montana, Colorado, and Oregon.
5.29. Parictis cf. parvus. YPM PU 16265. Right mandible with p3–m2. Scale in centimeters. Photo by the authors. Courtesy of the Division of Vertebrate Paleontology, YPM PU 16265, Peabody Museum of Natural History, Yale University, New Haven, Connecticut, U.S.A.
The Mustelidae includes the living weasels, minks, ferrets, badgers, wolverines, otters, and skunks, although some current taxonomists place skunks in their own family, Mephitidae. The family first appears in North America in the late Eocene (middle Chadronian), approximately 35.5 Ma. The majority of fossil mustelids in North America are the result of multiple dispersal events from the Old World with minimal in situ evolution. Of the 38 recognized fossil mustelid genera in North America, 26 are taxa that originated in the Old World (Baskin, 1998). To understand the fossil history of the North American mustelids, knowledge of the fossil record of the Eurasian members of the family is critical. The first appearance of many of the genera in the Tertiary of North America often provides an important biostratigraphic marker.
Members of the Mustelidae are characterized by having low braincases, wide occiputs, small orbits, anteriorly positioned carnassials, a moderately inflated auditory bulla, and short jaws that permit a powerful bite. The third molars are lost, and the second molars are reduced in size or may be absent as well. They are small- to medium-size carnivores with a tendency to elongated bodies and relatively short legs for their body size.
Systematics and Evolution The type of Mustelavus is M. priscus, and the genus is monotypic. This is a primitive mustelid, and the genus has also been placed in the Procyonidae (raccoon family), indicating the similarity of primitive members of the two families. Like other mustelids, the second molars of Mustelavus are reduced, whereas in procyonids these teeth tend to be enlarged, so currently it is considered to be a primitive mustelid. It is closely related to Mustelictis of the Old World but has a more primitive dentition (Baskin, 1998).
Distinctive Characters The skull of Mustelavus has a moderately inflated auditory bulla that contacts the base of the posteriorly projecting paroccipital process and broadly contacts the ventral part of the mastoid process. The dentition is secant, and all four premolars are present. The rounded and conical protocone on the upper carnassial is anteriorly placed, and the tooth has a deep carnassial notch. The upper first molar has a subtriangular outline of the occlusal surface. The upper second molar is small with three roots. The lower carnassial has a tall trigonid and a low-basined talonid (Baskin, 1998).
Stratigraphic and Geographic Distribution Mustelavus is known from Peanut Peak Member of the Chadron Formation (late Chadronian) and the Scenic Member of the Brule Formation (Orellan). It is only known from the Big Badlands (Baskin, 1998).
5.30. Palaeogale sp., SDSM 7118. (A) Skull, left lateral view. (B) Skull, dorsal view. (C) Skull, occlusal view. Scales in centimeters. Photos by the authors of specimen from the Museum of Geology, South Dakota School of Mines and Technology, Rapid City, South Dakota, U.S.A.
Systematics and Evolution The type species is P. minuta from Europe, and three species have been described in North America, P. sectoria, P. dorothiae, and P. sanguinarius, from the middle Chadronian to early Hemingfordian. Palaeogale is an enigmatic carnivore. Although it is currently placed with the mustelids, its relationships within the carnivora is still under debate. Relationships to an extinct felinelike family, Viverravidae, have been proposed (Hunt, 1974, 1989; Flynn and Galiano, 1982).
Distinctive Characters Palaeogale is a small animal. The auditory bulla is complete, inflated, and undivided. The upper fourth premolar has a parastyle and carnassial notch. The upper second molar is vestigial to absent, and it lacks an upper third molar (Fig. 5.30A, B) (Baskin, 1998).
5.31. Archaeotherium sp., BADL 34255/SDSM 67299. Articulated manus. Specimen was collected from the Big Pig Dig at Badlands National Park. Photo by the authors. Specimen is the property of the U.S. government. Specimen is housed at the Museum of Geology, South Dakota School of Mines and Technology, Rapid City, South Dakota, U.S.A.
Stratigraphic and Geographic Distribution The genus has a long stratigraphic history, first appearing in the middle Chadronian and becoming extinct at the end of the early Hemingfordian. There is a possible record of P. sectoria from the Scenic Member of the Brule Formation (Baskin, 1998). It is also known from Wyoming, Colorado, Montana, and Nebraska, and possibly Texas and Florida.
Members of the Order Artiodactyla, the even-toed or paraxonic ungulates in which the axis of the hand and foot passes between the third and fourth digits, comprise the most common hooved mammals in terms of taxonomic diversity and number of individuals in the modern fauna. A primary characteristic of the order is that the plane of symmetry of the front and hind foot passes between digits three and four, which are the largest digits (Fig. 5.31). Although being even toed is considered a primary characteristic of the order, some primitive forms may have a first digit, but it is usually vestigial, so it is the axis of symmetry that is the primary character. Digits 2 and 5 may be present and functional in some groups, such as peccaries, pigs, and hippopotamus, but in most groups, they become reduced in size, are vestigial, and are often lost early in the evolutionary history of these groups. In many forms, the third and fourth metapodials fuse in adults to become a single bone, often referred to as the cannon bone. Another primary skeletal character shared by all members of the order is the structure of the astragalus, which is a double pulley with both the proximal and distal ends having a similar shape. The calcaneum has a prominent convex articular surface for the distal fibula. Artiodactyls are primarily herbivores, although some, such as pigs, may be omnivorous, including scavenging; some extinct groups, such as entelodonts, are thought to have been scavengers as well. The order includes both browsers and grazers, and these differences in diet are often reflected in the dentition, with the former having brachyodont (low crowned) teeth while the latter are hypsodont (high crowned). In some advanced forms, the roots of the cheek teeth may be lost and the teeth are ever-growing, or hypselenodont. There are two primary types of dentition: a bunodont form with distinct cusps, such as is seen in peccaries and entelodonts (and humans), and a form referred to as selenodont in which the cusps form a semicircular or crescentic shape, and which is present in some White River taxa like the oreodonts and camelids, as well as modern deer, antelopes, and bison. Premolars are generally smaller than the molars and only rarely become molariform except occasionally the fourth premolar. The ulna and radius remain separate in some forms but may fuse in others, with the radius being the primary support of the body weight and the ulna becoming reduced to the proximal and distal ends. The femur never has a third trochanter. In primitive forms the fibula is complete, but in many groups it becomes reduced, losing the shaft so only the proximal and distal ends remain. The distal fibula articulates with the calcaneum and fits into a distinct facet in the distal tibia to serve as a lock to assist holding the astragalus in place. Although in most forms the ungual phalanx is modified into a hoof, there are some exceptions, as in the agriochoerids, in which the ungual is a claw.
Entelodonts are an extinct group of artiodactyls phylogenetically related to the living pigs and peccaries, as well as the hippopotamus. The family first appears in the late Eocene (Duchesnean) and became extinct in the early Miocene (late Hemingfordian). A possible earlier form, Brachyhyops, is known, but its phylogenetic relationships are unclear. The family was never diverse in North America, with only four known genera. One of these, Archaeotherium, is known from the White River Group and a second, Daeodon, is known from the Arikareean.
Entelodonts are distinguished by the proportionately large head for their body size. In Archaeotherium it is about 27 percent of the head–body length (Joeckel, 1990). The rostrum of the skull is greatly elongated and exceeds the postorbital length of the skull, and the braincase is small relative to the size of the animal. The zygomatic arches have expanded flanges that extend laterally and ventrally. The expansion of the squamosals gives the occiput a broad appearance. The brain cavity is small in comparison to the overall size of the skull, but the olfactory lobes were large, suggesting that smell was important to the animal. The frontal and parietal sinuses are enlarged and extend over the brain cavity. On the mandible there are two mandibular tubercles on the ventrolateral edge of the horizontal ramus (Fig. 5.32B) (Joeckel, 1990).
All three upper and lower incisors are pointed and round to subtriangular in cross section and increase in size laterally. The canines are large with serrations on the posterolateral and posteromedial margins in unworn teeth, but these are commonly worn off in older individuals. The cheek teeth are bunodont; all premolars and molars are present; and the premolars are large. The premolars are separated by diastema reflecting the elongation of the rostrum. The upper molars are brachyodont with six cusps, except for the third molar, which has a reduced posterior margin. The lower molars have four cusps, are square in outline, and increase in size posteriorly (Fig. 5.32B) (Effinger, 1998).
The neck is short and massive, with well-developed processes for the attachment of the neck musculature and ligaments, as would be expected given the disproportionately large head of the animal. The limb bones are relatively slender for their body size. The humerus is long and massive. The ulna and radius are co-ossified, and the ulna has a prominent olecranon process. The fibula is separate from the tibia in Archaeotherium, but in later forms the two bones become fused. The tibia is shorter than the femur but more massive. Like pigs and peccaries, the entelodonts are functionally didactyl, with large but short third and fourth metapodials and vestigial second and fifth metapodials (Effinger, 1998).
5.32. Archaeotherium sp. (A) Skull and jaws, right lateral view, UCM 19161. (B) Jaws, right lateral view, DMNS EPV 400. (C) Skull (male), occlusal view, DMNS 12720. (D) Skull (female), occlusal view, DMNS 1607. Scales in centimeters. Photo A by Katie McComas; photos (B–D) by the authors of specimens from DMNS and UCM. UCM 19161, University of Colorado Museum of Natural History, Boulder, Colorado, U.S.A. DMNS EPV 400, DMNS 12720, and DMNS 1607, Denver Museum of Nature and Science, Denver, Colorado, U.S.A. All rights reserved.
Systematics and Evolution The type species of Archaeotherium is A. mortoni from the Oligocene of the Badlands of South Dakota. Currently six species are considered valid. Megachoerus Troxell, 1920, was originally considered a distinct genus but is now considered a subgenus of Archaeotherium (Foss, 2007).
Distinctive Characters The body mass of Archaeotherium is estimated at about 150 kg (Joeckel, 1990). The skull morphology of Archaeotherium is extremely variable, reflecting pronounced sexual dimorphism and differences indicating changes with ontogenetic growth, making selection of characters to provide a consistent taxonomy difficult. The jugal flange in Archaeotherium is relatively straight and increases in thickness distally so that the lateral margin is swollen. The orbit is completely enclosed with a complete postorbital bar. The coronoid process of the mandible is small, suggesting they had a wide gape when opening the mouth. The mandibular symphysis is elongated, and the two halves are solidly fused together even in young individuals. The angular process of the mandible flares laterally. The upper first and second molars are subquadrate in outline, and the first has a distinct notch on the labial side that separates the tooth into anterior and posterior halves. The carpus retains the trapezium, which is lost in advanced entelodonts. The metatarsals are unfused but tightly articulated. Both the radius and ulna and the tibia and fibula are fused. The limb segment ratio suggests they had a cursorial terrestrial mode of locomotion (Figs. 5.31, 5.32, 5.33).
5.33. Archaeotherium mortoni. (A) Skull (male), dorsal view, DMNS 900. (B) Skull (female), dorsal view, DMNS 902. Photos by the authors of specimens from DMNS. DMNS 900, and DMNS 902, Denver Museum of Nature and Science, Denver, Colorado, U.S.A. All rights reserved.
Stratigraphic and Geographic Distribution This genus has the longest time range of all the North American entelodonts, first appearing in the early Chadronian and becoming extinct in the mid-Arikareean at the Oligocene–Miocene boundary.
Archaeotherium mortoni is known from the Ahearn Member of the Chadron Formation, the Crazy Johnson Member of the Chadron Formation, and the Scenic Member of the Brule Formation in the Big Badlands. Archaeotherium coarctatum is known only from the Ahearn Member of the Chadron Formation. Archaeotherium wanlessi is reported from the Scenic and Poleslide members of the Brule Formation, while A. lemleyi is known only from the Poleslide Member. The genus is also known from faunas in Texas, Wyoming, Nebraska, Montana, Colorado, and Saskatchewan.
Natural History and Paleoecology The presence of grooves worn along the alveolar margins of the lower canines and upper incisors has been suggested as indicative of rooting by entelodonts by Scott and Jepsen (1940), although Peterson (1909) had previously interpreted them as being scavenging omnivores, and perhaps in some cases active hunters. Evidence of scavenging by entelodonts is a skull of Merycoidodon with a fragment of a deciduous lower incisor of Archaeotherium embedded in the temporal bone of the skull (Mead, 1998). An analysis of the jaw mechanics and tooth wear by Joeckel (1990) indicates they were capable of ingesting large food items that included both mechanically resistant food items like roots and tubers and relatively hard materials like hard fruits and nuts, and possibly bones. Many specimens have broken teeth with subsequent polish on the remaining surface, possibly caused by biting on bone. It is possible that some coprolites found in the Big Badlands attributed to carnivores may have been produced by Archaeotherium. It appears that the preferred habitat of entelodonts was savanna–woodland or grassland.
Our knowledge of the paleobiology of Archaeotherium has recently been greatly enhanced by the discovery of a bone bed in Badlands National Park dubbed the Big Pig Dig, named for the numerous Archaeotherium fossils found at the site. A total of 19 genera were found at the site, with the minimum number of individuals for the four most common taxa as follows: eight Subhyracodon, 29 Archaeotherium, 11 Leptomeryx, and eight Mesohippus (Shelton et al., 2009). In all, a total of 19,290 specimens were collected from the site, and 187 m2 were excavated. Both A. mortoni and A. wanlessi were identified at the Big Pig Dig site, which is in the lower portion of the Scenic Member of the Brule Formation, about 5.5 m above the contact between the Chadron and Brule formations (Miller, 2010). Many of the bones of Subhyracodon preserve evidence of having been scavenged by Archaeotherium.
Hunt (1990) determined that the younger Agate bone bed represents the short-term, attritional accumulation of animals around a watering hole during drought conditions. The Agate bone bed also preserves evidence of bone processing and trampling (Hunt, 1990). The descendent of Archaeotherium, Daeodon, is well represented in the Agate deposits, and just as at the Big Pig Dig, many of the gouges and puncture marks found on the bones of the rhinoceros Menoceras closely match the teeth of Daeodon in size and shape, supporting the idea that entelodonts had a long history as scavengers.
Entelodonts appear to have displayed aggressive behavior. Many skulls display marks and pathologies that appear to have been the result of battles between males either fighting for territory or establishing dominance for mating. It is also possible some of the marks were made during mating (Sinclair, 1921a). Sexual dimorphism is inferred but not fully established. However, differences in the degree of development of the jugal flange, size of the canines, and size of the mandibular tubercles are highly suggestive that sexual dimorphism was present (Effinger, 1998).
Systematics and Evolution The type species of Daeodon is D. shoshonensis from the John Day Formation of Oregon (Lucas, Emry, and Foss, 1998). This animal is better known in the literature as Dinohyus based on D. hollandi recovered from bone beds in the Anderson Ranch Formation (late Arikareean) at Agate Fossil Beds National Monument in Nebraska. Daeodon is the terminal member of the family. Although five species have been described for this genus, currently only a single species is recognized (Lucas, Emry, and Foss, 1998). While Daeodon is presumed to have evolved from Archaeotherium, Lucas, Emry, and Foss (1998) suggested that the genus emigrated from Asia via Beringia in the early Arikareean. The two genera overlap during the early Arikareean.
Distinctive Characters With an estimated body mass of 750 kg, Daeodon is the largest member of the family, greatly exceeding in size all other genera (Joeckel, 1990). Despite its large size and a skull that was 25 percent to 35 percent of the head plus body length, the jugal flange is proportionately small for the size of the skull compared to other entelodonts. The mandibular tubercles are small, and the anterior pair is smaller than the posterior. The alveolar border of the premaxillae is relatively short, resulting in a crowding of the incisors. The tibia and fibula are co-ossified. The manus lacks the trapezium, and the fifth metatarsal may be absent in some individuals (Effinger, 1998).
Stratigraphic and Geographic Distribution This monotypic genus first appears in the early early Arikareean and became extinct at the end of the early Hemingfordian. It is only known from the Arikareean Cedar Pass fauna in the Sharps Formation of the Big Badlands (Parris and Green, 1969). By the late Arikareean the genus was widely distributed across North America and has been found in California, Oregon, Texas, South Dakota, Wyoming, Nebraska, New Jersey, Mississippi, Alabama, South Carolina, and Florida.
Natural History and Paleoecology Like Archaeotherium, Daeodon was predominately a scavenger, and many of the Menoceras bones in the Agate bone beds have gouges that closely match the premolars and canines of Daeodon in shape and size. These teeth on the skulls and jaws also often have extreme wear, suggesting they were used for crushing or biting bone. The preferred habitat appears to have been savanna–woodland or grasslands.
The relationship of this family to other artiodactyls has not been resolved. Traditionally it has been included as a subfamily within the Old World family Dichobunidae, but recent work indicates this creates an artificial paraphyletic family. It is a primitive group with bunodont (cusped) teeth in which the cusps tend to be inflated. The upper molars are triangular and tritubercular, while the lowers are quadritubercular. All of the cheek teeth from the third premolars to the third molars are robust. The first molar is the largest molar, and the molars decrease in size posteriorly. These are small taxa ranging in size from rabbit to about a modern peccary. Two genera are known from the White River Badlands, Leptochoerus and Stibarus.
The primitive tooth morphology suggests that the leptochoerids were omnivores and probably fed on a wide variety of foods, including fruits, seeds, and leafy vegetation. It is inferred, given their size, that they lived in dense undergrowth of woodlands. Their eventual extinction may have been the result of the loss of this habitat with the decline of forest and the spread of more open habitat in the Oligocene.
Systematics and Evolution The type species of Leptochoerus is L. spectabilis from the Brule Formation of the Badlands of South Dakota. Three other species are considered valid: L. elegans, L. supremus, and L. emilyae.
Distinctive Characters The skull has a short rostrum. The lower molars are more massive than in its contemporary, Stibarus. The upper and lower second premolar and lower third premolar tend to be tall, single-cusped, elongate teeth. The ulna and radius are separate, and the ulna is well developed. The manus had four functional digits. The fibula is reduced, and the distal end is fused with the tibia. The pes has four functional digits, and the cuboid and navicular are co-ossified, a feature characteristic of later cervids and bovids.
Stratigraphic and Geographic Distribution The genus first appears in the Chadronian and becomes extinct at the end of the late early Arikareean, approximately 23 Ma. Leptochoerus spectabilis is known from both the Ahearn and Crazy Johnson members of the Chadron Formation. Of the other three species described, L. elegans is described from the Scenic Member and L. supremus from the Poleslide Member of the Brule Formation. The genus is also known from Wyoming, Colorado, Nebraska, Montana, and North Dakota.
Systematics and Evolution The type species of Stibarus is S. obtusilobus, and two other species are known, S. quadricuspis and S. yoderensis.
Distinctive Characters The skull differs from Leptochoerus in having a pinched rostrum. There are two infraorbital foramina in Stibarus. As the bunodont lower molars become worn, the cusps develop a subcrescentic shape. The upper and lower second premolars and lower third premolar are low and have three cusps, with the medial cusp being the largest. The upper molars are triangular with a sharp lingual apex. Nothing is known of the postcranial skeleton of this animal.
Stratigraphic and Geographic Distribution Stibarus first appears in the early Chadronian and becomes extinct at the end of the Whitneyan. Only Stibarus obtusilobus, the type species, is known from the White River Group in the Scenic Member of the Brule Formation.
Members of the Tayassuidae, popularly known as peccaries or javelinas, originated in North America and underwent their primary evolution on this continent with later dispersals into South America in the Pliocene and to Eurasia. They first appear in the late Eocene (Chadronian). Today they are represented by the collared peccary in the southwestern United States, and their range extends southward into South America, which has three living species, the collared, white-lipped, and Chacoan. Modern peccaries are social animals and live in mixed-sex herds; this is believed to have been the case in the fossil forms as well.
Peccaries have prominent straight upper and lower canines with the uppers buttressed by massive lateral processes. These may be sexually dimorphic in some forms. Later Tertiary taxa may have large, laterally flaring zygomatic processes reminiscent of the entelodonts. The frontal is marked by deeply excavated supraorbital canals. The occiput is broad, and the external auditory meatus is enclosed in an elongate tympanic process of the squamosal. The cheek teeth are bunodont to zygodont, with some forms having teeth that approach being bilophodont. The ulna and radius may fuse in some forms. The third and fourth metapodials are large, and the second and fifth are reduced in size and may be lost. The metacarpals may remain separate, although the metatarsals fuse in some forms (Wright, 1998).
Systematics and Evolution The type and only species of Perchoerus is P. probus, collected by F. V. Hayden in the Badlands of South Dakota.
Distinctive Characters As a primitive member of the family, it is mostly distinguished by lacking characters seen in later forms. Features of the braincase that distinguish the genus include a dorsally directed external auditory meatus; the articular surface of the glenoid fossa extends slightly ventral to the basioccipital but is still above the occlusal plane; and there is a robust occipital crest. Like modern peccaries, the dentition is brachyodont, with quadrate molars having four cusps that are connected by small lophs. In the upper molars, the paracone and protocone are separated by a paraconule (Fig. 5.34) (Wright, 1998).
5.34. Perchoerus probus. YPM PU 12579. Skull, occlusal view. Scale in centimeters. Photo by the authors of specimen from YPM. Courtesy of the Division of Vertebrate Paleontology, YPM PU 12579, Peabody Museum of Natural History, Yale University, New Haven, Connecticut, U.S.A.
Stratigraphic and Geographic Distribution Perchoerus probus is known only from the Poleslide Member of the Brule Formation (Whitneyan).
Natural History and Paleoecology Fossils referable to Perchoerus are rare in the White River beds; only a few skulls and no skeletons have been found. Because of its rarity, little is known about the paleoecology of this genus. Presumably, like modern peccaries, Perchoerus was probably primarily herbivorous, but it may have occasionally consumed animals. Although modern peccaries are social animals and live in mixed-sex herds, it is difficult to determine if this was the case for Perchoerus, given its rarity and given that most of the known specimens consist of scattered remains.
Systematics and Evolution A second genus of peccary, Thinohyus, has been reported from the Big Badlands of South Dakota but has not been identified to species (Wright, 1998). The type and only described species is T. lentus is from the Arikareean sediments of John Day, Oregon.
Distinctive Characters Thinohyus is a rare taxon, and its diagnostic features are based essentially on the type specimen (Wright, 1998) and are consequently confined to features of the skull. It lacks features that distinguish the Perchoerus–Tayassu clade so can be considered the sister group to all other peccaries (Wright, 1998). In its dentition, the protocone and paracone are separated by a paraconule. Features of the skull include the postglenoid canal passing between the posttympanic process of the squamosal and the postglenoid process of the jugal; the tympanic process of the squamosal is dorsoventrally shallow; and the articular surface of the glenoid fossa is approximately in the plane of the basioccipital (Wright, 1998).
Stratigraphic and Geographic Distribution Thinohyus in the Big Badlands is known only from the Scenic Member of the Brule Formation. It is also known from the Wounded Knee fauna in South Dakota; Nebraska; and Oregon.
Natural History and Paleoecology Given its rarity, nothing has been written about the natural history of this taxon.
Members of the Anthracotheriidae are a group of primitive artiodactyls that seem to be somewhat convergent with modern hippos in their habits and appear to have been semiaquatic, although not all researchers agree on this interpretation (Kron and Manning, 1998). Also known from Europe and Africa, their diversity in North America is small, with six known genera, four of which are found in the White River Group of South Dakota. The earliest record is from the Duchesnean, and they became extinct at the end of the early Hemingfordian. The greatest diversity of anthracotheres occurs in the White River, with three genera, Heptacodon, Aepinacodon, and Bothriodon, present in the Chadronian. Heptacodon persisted into the Orellan and Whitneyan, where it overlapped with Elomeryx, which survived into the earliest Arikareean. The history of the anthracotheres in North America seems to be one of ecological replacement resulting from intercontinental dispersal rather than evolution in place, at least at the generic level (Kron and Manning, 1998). In the White River Group, Aepinacodon was replaced by Bothriodon, which was then replaced by Elomeryx.
The skull of anthracotheres tends to be long and slender, including an elongate rostrum and a narrow braincase with a well-developed sagittal crest. The internal nares are positioned posteriorly behind the upper third molar elongating the palate, supporting the idea that they may have been semiaquatic. As a general rule, they retain the primitive eutherian dental formula with the occasional loss of the first premolars. The cheek teeth are brachyodont. As the skull becomes more elongated in later forms, diastema develop between the premolars. The upper molars are large and square, with five cusps and a W-shaped ectoloph. The primary resemblances to hippos in the postcranial skeleton are the short, stout limbs. The body was not as massive as in hippos, and the skeleton lacks any graviportal modifications. The distal limb bones indicate they did not have a fully unguligrade stance. The manus retains five digits, but the pes has only four. Both the metacarpals and metatarsals are unfused (Kron and Manning, 1998). Sexual dimorphism has been documented in anthracotheres on the basis of the canines, with males having larger canines than the females, although the degree of difference is not as pronounced as in hippos or pigs. Differences in the anterior of the rostrum in Elomeryx armatus have also been interpreted as indicative of sexual dimorphism, with males having a premaxilla that is indented anterior to the canine, whereas in females there is no indentation (Macdonald, 1956).
Systematics and Evolution The type of Aepinacodon is A. deflectus from the Big Badlands of South Dakota, and a second species, A. americanus, is recognized. The name Aepinacodon was proposed by Troxell (1921:334) for the North American Oligocene anthracotheres not referable to Elomeryx, Octacodon, or Heptacodon. Among these he included Hyopotamus americanus Leidy, H. deflectus Marsh, and Ancodon rostratus Scott. Scott rejected this name in his White River monograph and referred the species A. americanus and A. rostratus to the European genus Bothriodon and ignored Marsh’s species H. deflectus. Macdonald (1956) revived Troxell’s genus in his review of the North American anthracotheres.
Distinctive Characters The genus is distinguished from Bothriodon by its shorter facial region of the skull, resulting from the shorter diastema between the canine and first upper premolar. The internal nares open just behind or at the level of the third molars. The canine is incisiform. The upper third premolar has a more prominent protocone.
Stratigraphic and Geographic Distribution Aepinacodon deflectus is known only from the Peanut Peak Member of the Chadron Formation, and A. americanus is known only from the Crazy Johnson Member of the Chadron Formation. Besides South Dakota, the genus is known from Nebraska and northern Colorado.
Systematics and Evolution The type of Bothriodon is B. velaunus from the early Oligocene of Western Europe. There are two species recognized in North America, B. rostratus and B. advena.
Distinctive Characters The diastema between the canine and first upper premolar is more pronounced than in Aepinacodon, so the rostrum of the skull is more elongate. The protocone of the upper third molar is reduced compared to Aepinacodon (Kron and Manning, 1998).
Stratigraphic and Geographic Distribution Bothriodon rostratus is found in the Scenic Member of the Brule Formation and the Orella Member of the Brule Formation in Nebraska, and B. advena is known from the Cypress Hills of Saskatchewan. The genus has also been reported from Wyoming (Kron and Manning, 1998).
Systematics and Evolution The type species of Elomeryx is E. armatus from the Badlands of South Dakota, and a second species, E. garbanii, has also been described.
5.35. Elomeryx armatus. YPM 10177. Skull, occlusal view. Scale in centimeters. Photos by the authors of specimens from YPM. Courtesy of the Division of Vertebrate Paleontology, YPM PU 10177, Peabody Museum of Natural History, Yale University, New Haven, Connecticut, U.S.A.
Distinctive Characters Elomeryx was about the size of a wild boar. The rostrum of the skull in Elomeryx is shorter than other anthracotheres such as Aepinacodon, and the diastema between teeth are less pronounced. The orbits are set high on the skull, another feature that has suggested the possibility of hippolike habits. The palate has a tubular extension that opens behind the last molar in Elomeryx, which also supports the idea of amphibious habits. The canine is large and extends ventrally in contrast to Bothriodon, which has small canines. The manus has five digits while the pes has four (Fig. 5.35) (Kron and Manning, 1998).
Stratigraphic and Geographic Distribution Elomeryx armatus is known from the Poleslide Member of the Brule Formation. The Orellan “Metamynodon channels” and the Whitneyan “Protoceras channels” of the Big Badlands contain the larger concentrations of its remains. The genus is also known from California, Nebraska, and Saskatchewan.
Natural History and Paleoecology The earliest records of Elomeryx are from the middle Eocene of Asia. It appears in Europe in the latest Eocene and is presumed to have dispersed into North America in the Oligocene. Although remains of Elomeryx are generally rare, the concentrations of its remains in the Orellan “Metamynodon channels” and Whitneyan “Protoceras channels” of the Big Badlands have been cited as evidence that anthracotheres were hippolike in their habits. On the basis of their association with nonaquatic taxa, Wilson (1975) proposed that the Protoceras Channel fauna actually represented a stream border habitat and that many of the taxa were in fact forest dwellers that just happened to be preserved in the channel deposits.
Systematics and Evolution The type of Heptacodon is H. curtus from the Whitneyan Protoceras channels of the Badlands of South Dakota, and four other species are recognized: H. gibbiceps, H. pellionis, H. occidentale, and H. quadratus.
Distinctive Characters The genus Heptacodon is quite distinctive. Its rostrum is short compared to other anthracotheres. The orbit is positioned more anteriorly on the skull, with its anterior margin almost to the first molar. The zygoma is deep. The mandible has an upturned symphysis. Heptacodon differs from the other contemporary anthracotheriids in having a fused mandibular symphysis without trace of the suture, while in most anthracotheriids the two halves of the mandible remain unfused. The canines are shifted laterally, and the upper and lower anterior premolars are not separated by diastema. The upper molars are heavily styled, and the anterior premolars are simple and generally rounded with nonangular cusps on both the upper and lower teeth (Macdonald, 1956). It differs from the other North American genera Bothriodon, Aepinacodon, and Arretotherium and Euro-American Elomeryx in the presence of a strong postprotocristid on its teeth, and the tooth rows lack a significant diastema between the canine and first lower premolar or between the first and second lower premolars (Kron and Manning, 1998).
5.36. Agriochoerus antiquus, SDSM 264. (A) Skull, right lateral view. (B) Right mandible, right lateral view. (C) Right mandible, occlusal view. (D) Skull, dorsal view. (E) Skull, occlusal view. Scales in centimeters. Photos by the authors of specimens from the Museum of Geology, South Dakota School of Mines and Technology, Rapid City, South Dakota, U.S.A.
Stratigraphic and Geographic Distribution Heptacodon first appears in the Duchesnean and became extinct at the end of the Whitneyan. Heptacodon curtus is known from the Poleslide Member of the Brule Formation in the Big Badlands and from the Orella and Whitney members of the Brule Formation in Nebraska. Heptacodon occidentale and H. quadratus are known from the Scenic Member of the Brule Formation and H. gibbiceps from the Poleslide Member of the Brule Formation. Stratigraphic distribution is the Duchesnean of Saskatchewan, Oregon, Texas and Utah; the Chadronian of Wyoming, South Dakota, and Colorado; and the Orellan and Whitneyan of South Dakota (Holroyd, 2002).
Natural History and Paleoecology According to Macdonald (1956), there is sexual dimorphism in the size of the canines, as in Elomeryx.
The families Agriochoeridae and Merycoidodontidae are closely related forms often referred to by the general term oreodonts. The Agriochoeridae is considered the more primitive of the two families.
The Agriochoeridae appear in the middle Eocene (late Bridgerian), and most genera became extinct in the late Eocene (Chadronian) with one genus, Agriochoerus, surviving until the end of the Oligocene. Eight genera are recognized, although most have not been formally named (Lander, 1998), leaving Agriochoerus as the only genus in the White River that has been formally described.
Members of the family retain many primitive features in the skull and the postcranial skeleton that have been lost in the Merycoidodontidae. Features that distinguish the agriochoerids from the merycoidodonts include the reduction and loss of the upper incisors; presence of a diastema between the canines and premolars; bothriodonlike molars; brachyodont molars and molariform third and fourth premolars; absence of a lachrymal pit; an incompletely closed orbit; a much heavier and more carnivorelike skeleton; and a far longer and thicker tail. The manus has five digits retaining the pollex, which is lost in the merycoidodonts, but the pes consists of four digits and unguals that are claws in some of the larger taxa.
5.37. Merycoidodon culbertsoni, skeleton, female found with two associated juveniles, right lateral view. SDSM 28129, photo by the authors of specimen from the Museum of Geology, South Dakota School of Mines and Technology, Rapid City, South Dakota, U.S.A.
Systematics and Evolution The type species of Agriochoerus is A. antiquus from the Brule Formation of the White River Badlands, South Dakota. Numerous species and subspecies have been described. Lander (1998) in his review of the genus reduced this to four subspecies of A. antiquus, and a second species, A. matthewi, with two subspecies.
Distinctive Characters Agriochoerus is larger than the other contemporary members of the family. The rostrum is constricted below the canine and second premolar. The upper incisors and first premolar are reduced or lost. The diastema between the upper first and second premolar is reduced or lost, while the one between the lower first and second premolars is expanded (Fig. 5.36). The ungual phalanges are clawed. Generally the skeleton of Agriochoerus resembles that of the carnivorous creodonts or fissipeds.
The cervical vertebrae, especially the axis, with its great hatchetlike neural spine, and the posterior thoracic and lumbar vertebrae are strongly built; the tail is extremely long and heavy; and the limb bones and clawed feet are all suggestive of a predator (Lander, 1998).
Stratigraphic and Geographic Distribution This genus has the longest biostratigraphic range of all members of the family, first appearing in the early Uintan, becoming extinct at the Oligocene–Miocene boundary. Given its long period of existence, numerous species and even subspecies have been described, many of which have been subsequently synonymized. The genotypic species, A. antiquus, is the only species recognized from the White River. It is known from the Peanut Peak Member of the Chadron Formation and the Scenic Member of the Brule Formation.
Natural History and Paleoecology There is no consensus on the habits of Agriochoerus. W. D. Matthew interpreted it as a tree climber, and the skeleton in the American Museum of Natural History is mounted so as to suggest this habit. Alternatively, it has been suggested that they were fossorial, although this many have been limited to the smaller forms. It has also been suggested they may have dug up roots and tubers as a principal part of their food.
The second oreodont family is the Merycoidodontidae, which first appears in the Duchesnean and became extinct at the end of the Miocene.
5.38. Merycoidodon culbertsoni, DMNS EPV 64 (A) Skull, left lateral view. (B) Mandibles, left lateral view. (C) Mandibles, occlusal view. (D) Skull, dorsal view. (E) Skull, occlusal view. Scales in centimeters. Photos by the authors of specimens from DMNS. DMNS EPV 64, Denver Museum of Nature and Science, Denver, Colorado, U.S.A. All rights reserved.
Members of the Merycoidodontidae are smaller than most of their ungulate contemporaries. This suggests a shorter life span and generation time, resulting in a high reproductive turnover. Oreodonts are the most common mammalian taxon found in the White River Badlands, suggesting they must have been a common animal on the landscape. On exhibit at the South Dakota School of Mines and Technology Museum of Geology is a skeleton of a female of Merycoidodon culbertsoni with two juveniles that were found in association with her, suggesting that twins may have been the normal litter size for the species (Fig. 5.37).
Systematics and Evolution The type species of Merycoidodon is M. culbertsonii from the Brule Formation of the Big Badlands of South Dakota, described by Leidy in 1848. It is one of the earliest fossil taxa described from the White River. The taxon is based on two cotypes in the Academy of Natural Sciences in Philadelphia, a partial jaw with three molars and a maxilla with the second and third molars. Leidy (1852) later replaced his own genus, Merycoidodon, with a new genus Oreodon for culbertsoni, which has contributed to taxonomic confusion regarding this animal. The validity of this taxon is not accepted by some recent workers such as Lander (1998), who preferred the later name Prodesmatochoeus periculorum, as he considered Leidy’s name a nomen vanum. As Leidy’s first generic name is well established in the literature, and as some subsequent workers (Stevens and Stevens, 1996) have chosen to retain it, we have done likewise, although we recognize that there are many nomenclatural issues with its retention. While Leidy’s second proposed genus, Oreodon, has no taxonomic validity, the term oreodont is still used as the popular or common name for this group of artiodactyls. In addition to M. culbertsoni, Stevens and Stevens (1996) recognized M. presidioensis, M. bullatus, and M. major.
Distinctive Characters Merycoidodon was about the size of a sheep. The skull has a complete postorbital bar. The dentition forms a continuous series without any diastemas. The first lower premolar is caniniform. The other premolars are simple and trenchant, but the upper fourth premolar is more molariform in its shape. The molars are selenodont, formed by four crescents in two transverse pairs but are low crowned. The facial portion of the skull is short, and the orbit is enclosed by a complete postorbital bar. There is a well-developed fossa in front of the orbit. The auditory bulla is small (Fig. 5.38). The neck and limbs are short in proportion to the body, and it retains a long tail, which in proportion to its body size is longer than in any other member of the family. Although an artiodactyl, one of its primitive features is that the manus has five digits, and while the pollex is reduced in size, it is complete with all phalanges (Figs. 5.38, 5.39) (Lander, 1998).
5.39. Bones of the oreodont Merycoidodon depicting individual elements including teeth and bone showing various features. Figures compiled from Scott and Jepsen (1940:fig 133, plate 69, fig. 2; plate 71, figs. 2, 9a, 10, 11, 12, 14, 15).
Stratigraphic and Geographic Distribution Merycoidodon culbertsoni is found in the Peanut Peak Member of the Chadron Formation and the Scenic Member of the Brule Formation. Merycoidodon major is found in the “lower Leptauchenia beds,” now known as the Poleslide Member of the Brule Formation in the Big Badlands and the Whitney Member of the Brule Formation in Nebraska. Merycoidodon bullatus is known from the Scenic and Poleslide members of the Brule Formation of the Big Badlands and from the Whitney Member of the Brule Formation of Nebraska and approximately equivalent beds in Montana, North Dakota, and Colorado. The genus is also known from faunas in Wyoming and Texas.
Natural History and Paleoecology Merycoidodon is the most common oreodont found in the White River, and its abundance far exceeds that of any other known taxa from the region or as Scott (1962:368) phrased it, “In the middle and lower White River (lower Brule and Chadron substages) the chief oreodont genus is Merycoidodon, the bones of which so far outnumber those of all other mammals, that the collector comes to regard them as a nuisance.” Given the abundance of this taxon, it must have been a herd animal.
5.40. Leptauchenia sp. UCM 17448 (A) Skull and jaws, left lateral view. (B) Skull, dorsal view. Scales in centimeters. Photos by the authors of specimen from UCM. UCM 17448, University of Colorado Museum of Natural History, Boulder, Colorado, U.S.A.
Systematics and Evolution The type of Leptauchenia is L. decora from the Brule Formation, White River Valley, South Dakota; two additional species, L. major and L. lullianus, are also recognized. Leptauchenia major is found in the White River Badlands, and L. lullianus is found in Montana (Prothero and Sanchez, 2008).
Distinctive Characters Compared to other contemporary oreodonts like Merycoidodon, Leptauchenia has a smaller body size. The skull has a reduced rostrum. The skull is dorsoventrally flattened. The orbits are small and positioned high up on the skull with their tops projecting above the level of the forehead. There are large antorbital vacuities that extend from above the orbits almost to the premaxillae and a small, shallow lacrimal fossa. The size of the antorbital vacuities has resulted in a reduction of the nasals to mere splints (Fig. 5.40). Prominences at the anterior margin of the nasal opening in some specimens along the nasal–maxillary suture, which may have formed attachment points for the soft tissue of the nose, although Janis (1982) has proposed they may have supported nasal horns. That these structures are not present in all individuals suggests they may have been sexually dimorphic. The tympanic bulla is highly inflated, and the external auditory meatus is an elongated tube located high and posteriorly on the skull. The occiput is expanded laterally. The first upper incisor is reduced or absent. The cheek teeth are hypsodont but with thin enamel. The fore- and hind limbs are of similar length. The ulna and radius are separate, and there is a complete fibula. The cuboid and navicular are separate bones (Lander, 1998).
Stratigraphic and Geographic Distribution Leptauchenia first appears in the late Orellan and becomes extinct in the late Arikareean. Leptauchenia decora appears in the early Orellan and continues to the early Arikareean. This species is found in the Poleslide Member of the Brule Formation. Besides South Dakota, the species also occurs in faunas in Nebraska, Wyoming, Colorado, and Montana. Leptauchenia major appears in the late Whitneyan and continues to the late Arikareean. The earlier L. decora is smaller than the later L. major (Prothero and Sanchez, 2008). It is also found in the High Plains and western Montana. Leptauchenia lullianus, which ranges within the early to middle Arikareean, is larger than L. major (Prothero and Sanchez, 2008). Prothero and Sanchez (2008) consider all three species to have been contemporaries in the early Arikareean. Leptauchenia lullianus is also found in the High Plains and eastern Montana.
Natural History and Paleoecology Because of its high-set eyes and ears, and large nasal-facial vacuities, Leptauchenia was considered to have been semiaquatic or amphibious (Cope, 1884; Matthew, 1899; Scott, 1937; Scott and Jepsen, 1940). Schultz and Falkenbach (1956) argued that Leptaucheniines were specialized for a desert mode of life. They based their interpretations on the eolian type sediments in which many specimens were found. They were compared to desert bovids, which have similar large bullae, large facial vacuities, and specialized nasal areas. In contrast, they are almost absent in the Protoceras channel deposits of the Poleslide Member of the Brule Formation. Leptaucheniines have hypsodont teeth with highly worn crowns, deep-set jaws, and a thick, heavy set zygomatic region – all features well adapted for an abrasive, grittier diet. Recent studies of the postcranial skeleton have suggested locomotor similarities to the living hyraxes (Wilhelm, 1993).
5.41. Eporeodon major. YPM PU 13594. Skull and jaws, left lateral view. Scale in centimeters. Photos by the authors of specimen from YPM. Courtesy of the Division of Vertebrate Paleontology, YPM PU 13594, Peabody Museum of Natural History, Yale University, New Haven, Connecticut, U.S.A.
Systematics and Evolution The type of Eporeodon is E. occidentalis from the Bridge Creek Locality of the John Day Valley, Oregon. Thorpe (1937) recognized 15 species within the genus, most of which have now been synonymized with E. occidentalis. Currently only a second species, E. major, known only from Arikareean of South Dakota, is also considered valid.
Distinctive Characters Eporeodon is a small genus of oreodont. The skull has a short facial region and a shallow zygomatic arch, and the sagittal crest is not prominent. The dentition is brachyodont with rectangular premolars. Eporeodon major is larger than E. occidentalis (Fig. 5.41) (Lander, 1998).
Stratigraphic and Geographic Distribution Eporeodon appears in the Whitneyan and becomes extinct at the end of the Arikareean; most records of the genus are from the Arikareean. Eporeodon occidentalis is present in the Poleslide Member (Whitneyan) of the Brule Formation in the Big Badlands. The genus is known from multiple localities in South Dakota, Wyoming, Nebraska, Oregon (John Day Formation), Florida, California, Montana, Washington, and Idaho.
Systematics and Evolution The type species of Miniochoerus is M. gracilis from Shannon County, South Dakota. Other species considered valid include M. forsythae, M. chadronensis, M. affinis, and M. starkensis.
Distinctive Characters Miniochoerus were relatively small and cranially stereotyped oreodonts and retained a minute auditory bulla. Their teeth became distinctive, with thin-enameled, quickly abraded fossettes and fossetids and foreshortened premolars (Stevens and Stevens, 2007).
Stratigraphic and Geographic Distribution The genus first appears in the early Chadronian and becomes extinct at the end of the Whitneyan. Miniochoerus forsythae is found in Wyoming and South Dakota from the middle Chadron Formation. Miniochoerus chadronensis is from the upper Chadron Formation in Wyoming and the lower Brule Formation in Nebraska. Miniochoerus affinis is from the Scenic Member of the Brule Formation in South Dakota and the lower Orella Member of the Brule Formation in Nebraska. Miniochoerus gracilis is from the Orella Member of the Brule Formation in Niobrara County, Wyoming, and the Scenic Member of the Brule Formation in South Dakota. Miniochoerus gracilis is also found in Nebraska and Colorado. Miniochoerus starkensis is found in the Brule Formation, Fitterer Badlands, North Dakota.
Systematics and Evolution The type species of Mesoreodon is M. chelonyx from the Fort Logan Formation of Montana. Besides the type species, a second species, M. minor, is tentatively recognized by Stevens and Stevens (1996, 2007). Lander (1998) considered Mesoreodon a junior synonym of Eporeodon. The taxonomy of the genus has been complex, and the name Mesoreodon has been restricted to forms with wide and low skulls. Some of the confusion appears to reflect differences in the postmortem deformation of the skull. The genus appears to be the ancestor to Hypsiops and consequently to the Desmatochoerus–Promerycochoerus lineage of oreodonts.
Distinctive Characters The general form of the cranium is similar to that of Merycoidodon but with inclined premaxillae. The antorbital fossa is shallow and pitlike. The infraorbital foramen is positioned above the third and fourth premolars. The premolars are only slightly reduced, and all teeth are slightly narrower and higher crowned than other contemporary oreodont genera. According to Stevens and Stevens (1996, 2007), Mesoreodon is distinguished from Eporeodon by its larger size, the pitlike antorbital fossa, a higher rostrum, the slightly reduced premolars, and anteroposteriorly shorter molars that are higher crowned.
Stratigraphic and Geographic Distribution As defined by Stevens and Stevens (1996, 2007), the genus appears in the latest Whitneyan and became extinct in the early Arikareean. In the White River Badlands, it has only been found at Quiver Hill, south of Kadoka, South Dakota (Arikareean). The genus is also known from Oregon, Montana, Wyoming, and Nebraska.
Natural History and Paleoecology Nothing specific has been proposed regarding the natural history or paleoecology of this genus.
The family Camelidae is today represented by the Old World dromedary and Bactrian camels and the South America llamas, vicuñas, and guanacos. The family originated in North America, with the earliest records in the Uintan, approximately 45.9 Ma. The primary evolutionary history of the family is in North America, and they did not disperse into Eurasia until the late Miocene and into South American until the early Pleistocene.
Camelids lack cranial structures like horns, antlers, and ossicones. They retain a canine, and the third incisors and anterior premolars may become caniniform in their morphology. The skull has a sagittal crest. In front of the orbit there is a lacrimal vacuity and a maxillary fossa that is well developed in early camelids but may be reduced in size in later forms. The first and second upper incisors are reduced, but the lower incisors are large and spatulate. The true lower canine becomes incisiform, and some premolars are caniniform. The molars are four cusped with straight ectolophs and with fossettes that are closed both anteriorly and posteriorly after moderate wear. Camelids have long necks compared to most artiodactyls and are only exceeded by the giraffids in length. A distinctive character of the cervical vertebrae is that the vertebral artery passes inside the pedicle of the neural arch and its posterior portion is confluent with the neural canal. The ulna and radius are co-ossified. The fibula is reduced and lacks the midshaft so there is only a small proximal end, and the distal malleolus locks into the distal tibia. The cuboid and navicular remain separate even in advanced forms, and this is reflected in the distal end of the astragalus, which has a keel that fits between them. Only the second and third metacarpals and metatarsals are present; they have a distinctive distal flare, with the keel on the distal articular surfaces confined to the posterior margin. The metapodials may be fused or unfused depending on the taxon. Primitive camelids have an unguligrade stance, but many later genera are digitigrade (Honey et al., 1998).
Systematics and Evolution The type species of Poebrotherium is P. wilsoni from the Badlands of South Dakota; the genus includes two other species, P. chadronense and P. eximium. Poebrotherium wilsoni was the first Tertiary camelid described from North America.
5.42. Poebrotherium wilsoni (A) YPM PU 12722, skull and jaws, left lateral view. (B) SDSM 2913, mandible, occlusal view. (C) SDSM 2913, skull, dorsal view. (D) USNM 15917, Poebrotherium sp., skeleton, left lateral view. (E) SDSM 2913, skull, occlusal view. Scales in centimeters. Photos by the authors of specimens from SDSM, USNM, and YPM. Courtesy of the Division of Vertebrate Paleontology, YPM PU 12722, Peabody Museum of Natural History, Yale University, New Haven, Connecticut, U.S.A. SDSM 2913 is from the Museum of Geology, South Dakota School of Mines and Technology, Rapid City, South Dakota, U.S.A. USNM 15917, courtesy of Smithsonian Institution, Washington, D.C., U.S.A.
Distinctive Characters Compared to later camelids, Poebrotherium is small. The largest species is about the size of a sheep but with a more slender build and a long neck and body (Fig. 5.42D). It maintains a complete dental formula and lacks a diastema between the anterior teeth. The teeth are less hypsodont than other camelids except Poebrodon. The skull lacks a postorbital bar, a feature seen in later camelids (Fig. 5.42A–C). A primitive feature seen in some Poebrotherium is a bifurcation of the protocone of the upper molars. The humerus has a single bicipital groove. The ulna and radius are co-ossified. The fibula has already lost the shaft and is reduced to a proximal splint that is fused to the tibia, and the distal malleolus is set in the distal tibia to lock the astragalus in place. Digit 1, is lost and digits 2 and 5 are reduced to vestigial nubbins in both the front and hind feet. The third and fourth metapodials remain unfused (Honey et al., 1998).
Stratigraphic and Geographic Distribution This is the most common Oligocene camelid and is the most primitive except for Poebrodon, a smaller genus from the Uintan. Poebrotherium first appears in the Chadronian and became extinct at the end of the early Arikareean. Poebrotherium eximium is found in the Peanut Peak Member of the Chadron Formation and the Scenic Member of the Brule Formation, and P. wilsoni is known from the Scenic and Poleslide members of the Brule Formation. The genus is also known from Wyoming, Colorado, Nebraska, North Dakota, and Montana.
Natural History and Paleoecology In the Chadronian and Orellan, camelids are common in the White River deposits of Colorado and southeastern Wyoming but rare in South Dakota and farther north, suggesting differences in habitat in these regions that limited their presence at higher latitudes. Clark, Beerbower, and Kietzke (1967) observed that in the Big Badlands, Poebrotherium is typically found in sediments far from channels, indicating it may have preferred drier habitat.
Systematics and Evolution The type of Paralabis is P. cedrensis and is the only species currently recognized. The type was collected from the Titanotherium beds (late Chadronian) of northeastern Colorado (Lull, 1921).
This taxon has had a confusing taxonomic history. An unfortunate lapse in Matthew (1901) illustrated a lower jaw, AMNH 8969, as the type of Protomeryx cedrensis, but the species name campester was used in the text, which cited the same catalog number for the type. The assignment of the species to Leidy’s genus Protomeryx, which had been used as a catchall name for many early camels – and McKenna (1966) considered the name a nomen dubium – was changed when Lull (1921) transferred the species to his new subgenus, Paralabis, within Pseudolabis. Both of these are now considered separate genera (Honey et al., 1998). Stock (1935) clarified the taxonomy with the recognition of cedrensis as the valid species name.
Distinctive Characters Paralabis apparently represents an advanced descendant of Poebrotherium in the Whitneyan. The muzzle has become elongated. The lower second to fourth premolars are shortened, and the diastema is better developed, especially between the lower first and second premolars where the depth of the jaw is at a minimum (Lull, 1921).
Stratigraphic and Geographic Distribution Paralabis cedrensis is known from the Poleslide Member of the Brule Formation (Whitneyan) in the Big Badlands but has also been reported from faunas in Colorado and Nebraska.
Systematics and Evolution The type and only species of Pseudolabis is P. dakotensis from the Whitneyan Protoceras beds of South Dakota. It was the first camel reported from the Protoceras beds or channel deposits in the Poleslide Member of the Brule Formation. It is the earliest representative of the Stenomyline or gazellelike camelids.
Distinctive Characters Pseudolabis is larger than Poebrotherium. The skull has a deeply depressed maxillary fossa, and the premaxilla extends posteriorly to the first premolar. It is the first camelid to have a complete postorbital bar that encloses the orbit. The teeth are relatively hypsodont, as in later members of the subfamily, and compared to Poebrotherium, the teeth are more elongate and transversely narrower. As the earliest stenomyline camelid, it is primitive in having a relatively short rostrum and unreduced premolars. The upper first premolar is caniniform in its morphology. The upper fourth premolar has a double internal crescent. The molars lack mesostyles (Matthew, 1904; Honey et al., 1998).
Stratigraphic and Geographic Distribution First appearing in the Whitneyan, the stratigraphic range of the genus extends into the early late Arikareean. In the Big Badlands, Pseudolabis dakotensis is present only in the Poleslide Member of the Brule Formation (Whitneyan). It is also found in faunas in Wyoming and Nebraska.
The family Protoceratidae first appears in the middle Eocene (early Uintan) and became extinct in the early Pliocene (late Hemphillian) (Patton and Taylor, 1973). The family takes its name from Protoceras, the first genus of the family described. The family currently includes 13 genera, and three genera, Protoceras, Leptotragulus, and Heteromeryx, are known from the White River Badlands.
The family is perhaps best known for the later genus, Synthetoceras, with its elongated slingshotlike horn on the tip of its nasals, and while many genera of the family do have well-developed horns, they are often absent in the earlier more primitive genera. There is a small facial vacuity anterior to the orbit, which is completely enclosed. Although upper incisors are present in primitive genera, they are lost in later genera – a trend that parallels the situation seen in many living artiodactyls such as camelids, cervids, and bovids. The jaw flexes ventrally at the mandibular symphysis, and the coronoid process of the jaw is one of the shortest among all ungulates. The dentition is of the primitive bunoselenodont type, with the molars widened laterally but shortened anteroposteriorly. Although the teeth are selenodont, there is no bifurcation of the lobes. There are prominent parastyles and mesostyles on the cheek teeth and a well-developed lingual cingulum (Prothero, 1998b).
Protoceratids share many features of the postcranial skeleton with camels. Among these is the vertebral canal of the cervicals passing through the neural arch and not the transverse process. One evolutionary trend in the family is a modification of the limbs for increased cursoriality, such as the fusion of the ulna and radius in later genera, but they never achieve the same level of modification seen in camels or the pecorans. The lateral metapodials are reduced in later genera, but they are not totally lost, so even the later genera have four toes and the metapodials never fuse. They also resemble camelids in the keel of the distal end of the metapodial being confined to the plantar surface of the articulation for the first phalanx. As in the camels, the distal astragalus has a distinct distal keel, reflecting the nonfusion of the cuboid and navicular (Prothero, 1998b).
5.43. Protoceras celer, adapted from Scott and Jepsen (1940: plate 60, fig. 1, and plate 61, fig. 1, 1b). (A) Skull (male) reversed, left lateral view, SDSM 2814. (B) Skull (female), right lateral view, YPM PU 10655. (C) Skull (female), occlusal view, YPU PU 10655.
Systematics and Evolution The type species of Protoceras is P. celer from the upper part of the Poleslide Member of the Brule Formation of South Dakota, and two other species are recognized.
Distinctive Characters The skull has a posteriorly placed orbit, and there are strong sagittal and parietal crests. The rostrum of the skull is elongated, with shortened or retracted nasals, possibly suggesting the presence of a flexible proboscis or muzzle like a tapir or saiga antelope. The upper incisors have been lost, but in males, the upper canine is a short, strong, recurved tusk. In females, the canine is reduced and vestigial. The lower canine is incisiform, while the first premolar in the male is caniniform. The other premolars are elongated anteroposteriorly and narrow with sharp cusps, while the molars are low crowned (Fig. 5.43). The ulna and radius are starting to co-ossify but only at their distal ends; each is still a fully functional bone. The fibula is totally lost. The manus has four functional digits, while the pes is functionally two toed, but the metatarsals remain separate from each other. As in camelids, the cuboid and navicular remain separate but differ in having a fused ecto- and mesocuneiform, which occurs in later genera. The second and fifth metatarsals are long and slender splints and lack phalanges (Prothero, 1998b).
Stratigraphic and Geographic Distribution Protoceras first appears in the Whitneyan and became extinct in the early late Arikareean. Only one species, P. celer, is present in the Big Badlands from the Poleslide Member of the Brule Formation (Whitneyan). The genus is also known from faunas in Nebraska.
Natural History and Paleoecology The type specimen of Protoceras is based on a female skull, but with the discovery of additional specimens, the pronounced sexual dimorphism in Protoceras was quickly recognized (Osborn and Wortman, 1892). The male has three pairs of bony hornlike structures. There is a pair that arises from the parietals just behind the orbits, the dorsal margin of the orbit is expanded into a triangular structure, and there is a dorsal expansion of the maxilla above the narial opening. The structure and size of these seems to vary between species, but a detailed study of this variation and how much it is determined by the stage of ontogenetic development has not yet been done. In P. celer, the anterior protuberances on the maxillae are simple, broad recurved plates that are thickened and rugose on the margins, and the posterior horns are short and club shaped. In females, the anterior protuberances on the maxillae are missing, while the posterior pair is small and hornlike (Prothero, 1998b).
Remains of Protoceras are so common in channel deposits in the late Whitneyan of the Big Badlands that the deposits are known as the Protoceras channels. The structure of their teeth indicates folivorous (leaf-eating) habits, and with the retracted nasals, a flexible nose that is either tapirlike or moose-like suggests that they may have fed on semiaquatic plants (Janis, 1982). Their short limbs and unfused metapodials suggest that they were not fast runners and probably inhabited bushy terrain, such as riparian vegetation along watercourses rather than open country. The pronounced differences seen in the development of the hornlike structures of the skulls of males and females suggests they were important for visual display during mating and perhaps in males in establishing dominance. The horns of Protoceras are best seen from the side, while those in other members of the family are more readily seen from the front. Battles during rut between males may have consisted of neck wrestling and pushing and pulling, with horns of males interlocking and preventing serious damage to the contestants (Prothero, 1998b).
Systematics and Evolution The type of Leptotragulus is L. proavus, and two, or possibly three, other species are considered valid. The type is from the Uintan of Uintah County, Utah.
Distinctive Characters This is a small, primitive protoceratid. The metaconid on the lower fourth premolar is not well developed, but the parastylid is prominent on the sharply flexed anterior crest (Prothero, 1998b).
Stratigraphic and Geographic Distribution The stratigraphic range of the genus is from early Uintan to middle Chadronian. This genus is not well known from the White River Group of South Dakota. “Leptotragulus” profectus from the Peanut Peak Member of the Chadron Formation may actually belong to another genus, Trigenicus (Prothero, 1998b). The genus is also known from faunas in Utah, Wyoming, Texas, Nebraska, Montana, and Saskatchewan.
Systematics and Evolution The type of Heteromeryx is H. dispar from the middle Chadron Formation on Indian Creek, Cheyenne River, South Dakota. The type specimen is represented by a skull with a considerable part of the skeleton, including most of the forefeet and parts of the hind feet. It is the only species in the genus (Prothero, 1998b).
Matthew (1905) originally considered Heteromeryx to be in the family Hypertragulidae on the basis of the presence of a functionally tetradactyl manus with a didactyl pes. He considered the genus to be intermediate between Leptomeryx and Protoceras and a possible ancestor to Protoceras. The genus is now placed in the Protoceratidae (Prothero, 1998b).
Distinctive Characters Heteromeryx is smaller than Protoceras. The skull of Heteromeryx is rather short, with an elongate muzzle, and the orbits are enclosed by a complete postorbital bar and are located above the posterior molars. The type skull lacks horns; it is not known whether this is characteristic of the genus, or alternatively whether the skull is from a female. The nasals are reduced and separated from the maxilla by a narial notch that extends to the third premolar. It is considered to be a primitive member of the family because the skull lacks horns or protuberances. The teeth are short crowned. There are four premolars, the first small, simple, and single rooted, with long diastema before and behind it, and the others much as in Leptomeryx. The molars have heavy internal cingula and a rudimentary mesostyle. The ulna has a well-developed shaft and is co-ossified with a radius along its entire length. The manus is functionally tetradactyl, with four separate digits, as in Leptomeryx. The magnum and cuneiform are fused. The distal end of the fibula is separate from the tibia. The pes is didactyl, with separate metatarsals. The cuboid and navicular are distinct, but the ecto- and mesocuneiform are fused. The ungual phalanges are short and compressed (Prothero, 1998b).
Stratigraphic and Geographic Distribution The genus first appears in the late Uintan and became extinct at the end of the Chadronian. In the Big Badlands, it is only known from the Ahearn Member of the Chadron Formation (early Chadronian). It is also known from similar age deposits in Nebraska and Uintan to middle Chadronian faunas in Texas.
Hypertragulids have a skull with a narrow rostrum, orbits centrally positioned on the skull, a reduced postorbital bar so the orbit is confluent with the temporalis fossa, and an auditory bulla with a long bony external auditory meatus. There are no antlers in males, unlike their modern relatives, the cervids. They do, however, retain a large upper and lower canine, and they resemble the modern musk deer of Southeast Asia. The upper molars have strong ribs on their labial (cheek) margin of the crown. The forelimbs are shorter than the hind limbs, giving them a posture reminiscent of the musk deer. The ulna and radius are co-ossified. The manus is primitive, with five digits (pentadactyl); the metacarpals are unfused, and the lateral digits are complete with phalanges but are shortened relative to the central digits and probably functioned like dewclaws. The fibula is complete. The pes has four digits with the metatarsals unfused, and the second and fifth metatarsals are reduced in size but still retain phalanges. A distinctive feature of the family is the “bent” astragalus, with the proximal and distal portions at an oblique angle to each other.
Systematics and Evolution The type species of Hypertragulus is H. calcaratus from the Brule Formation of Colorado, and three other species are recognized.
Distinctive Characters While the overall body size of Hypertragulus is small, it is the largest member of the family except for Nanotragulus. The teeth are not as hypsodont as other members of the family. Both the upper and lower molars have prominent cingula on their anterior margins, and accessory conules are also present. Both the upper and lower second premolars are caniniform and separated from the other cheek teeth by diastema (Fig. 5.44) (Webb, 1998).
Stratigraphic and Geographic Distribution Hypertragulus calcaratus is the only species known from the Big Badlands, where it is found in the Scenic and Poleslide members of the Brule Formation. It is also found in California, Colorado, Nebraska, Montana, Chihuahua (Mexico), Oregon, and Wyoming.
Natural History and Paleoecology Sites containing multiple Hypertragulus skeletons have been found (Fig. 5.44), suggesting that this was a social animal that probably lived in herds.
Systematics and Evolution The type species of Hypisodus is H. minimus from the White River of Colorado, and only a single species is known.
Distinctive Characters Hypisodus is smaller than Hypertragulus and is about the size of a rabbit. It is distinguished by a narrow and shortened rostrum and an anteroposteriorly compressed braincase. The orbit is large in comparison to the size of the skull, and the postorbital bar is incomplete. The auditory bullae are greatly enlarged so that they meet at the midline of the skull (Matthew, 1902). All of the upper incisors – the canine and the first and second premolar – have been lost. The molars are hypsodont and increase in size posteriorly. The upper molars have flat labial walls. The coronoid process of the mandible is shorter and more slender than in Leptomeryx (Fig. 5.45). The third and fourth metatarsals are nearly fused, and the second and fifth metatarsals are reduced.
Stratigraphic and Geographic Distribution The genus appears in the Duchesnean and became extinct in the early Arikareean. Hypisodus minimus is only found in the Scenic Member of the Brule Formation, but the genus is also known in the Poleslide Member in the Big Badlands. The genus is widespread and is found in a number of localities in Colorado, Nebraska, and Wyoming.
5.44. Hypertragulus calcaratus. (A) Multiple skeletons on block, UCM 19659. (B) Skeleton, right lateral view, USNM V 16744. Scale in centimeters. Photos by the authors of specimens from UCM and USNM. UCM 19659, University of Colorado Museum of Natural History, Boulder, Colorado, U.S.A. USNM V 16744, courtesy of Smithsonian Institution, Washington, D.C., U.S.A.
5.45. Hypisodus sp., DMNS 13083. (A) Skull, left lateral view, reversed. (B) Right mandible, right lateral view. (C) Right mandible, occlusal view, reversed. (D) Skull, dorsal view. (E) Skull, occlusal view. Scales in centimeters. Photos by the authors of specimens from DMNS. DMNS 13083, Denver Museum of Nature and Science, Denver, Colorado, U.S.A. All rights reserved.
Natural History and Paleoecology On the basis of the large orbit and the enlarged tympanic bulla, Hypisodus was probably nocturnal in its habits.
Leptomerycids are another group of North American hornless ruminants. Features of the skeleton that characterize the ruminants that can be seen in fossils include the loss of the upper incisors, the presence of procumbent lower incisors, and the fusion of the cuboid and navicular into a composite bone, the cubonavicular (Webb, 1998).
The family was originally considered to be a subfamily within the Hypertragulidae until a study by Gazin (1955) demonstrated a suite of distinctive features that distinguished the two main North American groups. Members of the Leptomerycidae can be distinguished from the Hypertragulidae by their broader rostrum and by the presence of a complete postorbital bar. As a primitive ruminant, both Leptomerycids and Hypertragulids have orbits located at the midpoint of the skull, reflecting their shorter rostrum than in later forms.
One of the most distinctive features of members of the family is an enlarged procumbent lower first incisor. Unlike the Hypertragulids, the upper canine is reduced in size, and the first upper premolar is lost. The upper first and second premolars have protocones, and the upper fourth premolar has a strong lingual crescent. The first lower premolar is caniniform or reduced in size, and the other premolars form a continuous series of three-cusped teeth.
Postcranially, Leptomerycids can be distinguished from Hypertragulids by a number of generally more derived features. The odontoid process of the axis is spoutlike. The magnum and trapezoid in the carpus are fused, and the manus is tetradactyl. The astragalus has parallel sides and is not bent, and metatarsals 3 and 4 are fused. The midshaft of the fibula is lost, with only a vestigial proximal end, and the distal malleolus serves as a lock to aid holding the astragalus in place (Webb, 1998).
With the inclusion of Archaeomeryx and other Old World genera, the family Leptomeryicidae at one time geographically extended into Asia, but they have been recently placed in their own family, so the Leptomerycids are not considered to be strictly North American but rather derived from an Asian ancestor. The family first appears in the Duchesnean and became extinct in the Arikareean (Heaton and Emry, 1996).
Systematics and Evolution The type species of Leptomeryx is L. evansi from the White River Badlands of South Dakota. In addition to the type species, five other species are recognized as valid. Because Leptomeryx is hornless, dentition, mostly the lower (because jaws are commonly preserved), has been used to define the different species. This has resulted in the description of multiple species and has produced a confusing taxonomy as a result of the large amount of variation in the morphology of the lower premolars, especially the third. Consequently, individual species often cannot be identified on the basis of a single specimen, and a statistical sample is often needed (Heaton and Emry, 1996). The evolutionary history of Leptomeryx includes several important morphological transformations, including the evolution of Leptomeryx from Hendryomeryx in the latest Duchesnean or earliest Chadronian; the splitting of L. yoderi into L. speciosus and L. mammifer in the early to middle Chadronian; and the evolution of L. speciosus into L. evansi in the late Chadronian and early Orellan (Heaton and Emry, 1996).
Distinctive Characters This is a small artiodactyl about the size of a house cat. The general body form is similar to that of the living mouse-deer. The skull is elongate with a slender muzzle. The orbits are positioned at the midpoint of the skull. All of the upper incisors have been lost, and both the upper and lower canines are reduced in size, but the lower canine functions as an incisor and the first premolar is caniniform in shape. The other premolars are narrow and sharp pointed, and the molars are low crowned. The hind legs and feet are longer than the front. The ulna is complete and separate from the radius, but the fibula is reduced. The manus has four digits, all separate, but in the pes there are only two digits, and the metatarsals are fused into a cannon bone (Fig. 5.46).
Stratigraphic and Geographic Distribution The genus first appears in the late Uintan and became extinct at the end of the early Hemingfordian. The type species is the only described species from the White River Group and is known from the Orellan to Whitneyan Scenic and Poleslide members of the Brule Formation of South Dakota. The genus is also found in Colorado, Nebraska, Montana, Saskatchewan, California, Wyoming, and Texas.
Natural History and Paleoecology This is one of the more common artiodactyls present in the Chadronian and Orellan faunas of the Great Plains ranging from Saskatchewan to Texas. Fossil deposits with this genus are sporadic in their distribution, making environmental and evolutionary reconstruction difficult. Important evolutionary transitions are not always recorded (Heaton and Emry, 1996).
Because Leptomeryx is so common, sufficiently large samples have been recovered, including accumulations of multiple skeletons that permit population studies. According to Clark and Guensburg (1970), Leptomeryx occurs in collections in greater numbers than any other genus except Palaeolagus, although this statement is difficult to believe, given the abundance of the oreodont Merycoidodon. Many specimens have been recovered as complete articulated skeletons, and multiple individuals have been recovered in a large block, suggesting the presence of a single herd. Scott and Jepsen (1940) cited one assemblage containing 20 individuals. These death assemblages were found near water sources associated with a swamp on a grassy or savanna plain. When wear stages of the teeth were examined, six age groups were identified in the samples. The presence of distinct age classes in the populations suggests seasonal breeding and birthing. Maximum life span was inferred to have been about 8 years (Clark and Guensburg, 1970).
The order Perissodactyla is represented today by horses, tapirs, and rhinoceros and by two extinct groups, brontotheres and chalicotheres. The order first appears in the late Paleocene in North America, and fossils of the group are found on all continents except Australia and Antarctica. Members of the order are characterized by having the axis of symmetry of the feet pass through the middle of the third digit (mesaxonic), and while they are popularly called the odd-toed ungulates, some forms may actually have four digits, usually on the manus. Another characteristic of the order is the presence of a prominent and well-developed third trochanter on the femur. The astragalus is distinctive, with a single pulley surface for articulation with the tibia rotated so it is at an angle to the axis of the bone and a flattened and compressed navicular process. The oblique axis of the trochlea of the astragalus is also reflected in the distal end of the tibia in which the articular surfaces are oriented obliquely to the long axis of the shaft.
5.46. Leptomeryx sp. (A) Skeleton, right lateral view, PM 61157. (B) Block containing several skeletons, P 12320. (C) Leptomeryx evansi partial skull, occlusal view, USNM 157 holotype. (D) Leptomeryx evansi partial right mandible, occlusal view, USNM 157 holotype. Scales in centimeters. Photos A and B by William Simpson, Field Museum of Natural History; photos C and D by the authors of specimens from USNM. USNM 157, courtesy of Smithsonian Institution, Washington, D.C., U.S.A.
The family Tapiridae originated in North America, where it underwent most of its evolutionary history. Today there is a single genus, with three species in South America and a single species in Malaysia. The family shares a common ancestry with rhinos, but the two lineages diverged in the early Eocene, and the earliest tapir is from the early Uintan.
Although tapirs have a typical perissodactyl manus and pes in which the axis passes through the third digit and the third metapodial is the largest in the series, the manus has four digits. The first has been lost and the fifth is reduced in size and nonfunctional. The pes has three digits, having lost the first and fifth, but digits 2, 3, and 4 are all functional. First appearing in the middle Eocene (Bridgerian) in the genus Helaletes (found in Colorado, Montana, Nevada, Utah, and Wyoming), a characteristic of tapirs is the shortening of the nasals and the development of a short, fleshy, prehensile proboscis, a feature present in living tapirs. They have a complete placental dental formula of C3/3-I1/1-P4/4-M3/3 totaling 44 teeth. The incisors are chisel shaped, and the third upper incisor is the largest in the series and is conical. It is separated by a short diastema from the smaller canine. There is a much longer diastema between the canines and premolars, the first of which may be absent. While the dentition is brachyodont, the cusps have coalesced to form lophs. In the upper dentition, the lophs form a characteristic pattern that resembles the Greek letter pi (π), with a straight ectoloph on the labial side of the tooth. The lower teeth have two unconnected transverse lophs. As in all members of the order, the femur has a well-developed third trochanter. The fibula is complete (Colbert and Schoch, 1998).
Systematics and Evolution The type species of Protapirus is P. priscus from the phosphorite deposits of Quercy, France (late Eocene). Five species are recognized in North America.
Distinctive Characters Protapirus is in the medium size range for members of the family. The third and fourth premolars are nonmolariform to submolariform. Differences in tooth morphology used to describe the different species are based on differences in the premolars, and it is possible that these differences simply reflect individual variation within a population. The genus is in need of revision. The shortening of the nasals in Protapirus is not as pronounced as in modern tapirs, and they extend to above the canine, so the flexible proboscis may not have been as well developed. They are separated from the maxillae by an enlarged narial incision. The manus has four digits; the first digit is lost, and the fifth digit is reduced to a similar degree as in the Malayan tapir and thus may not have borne weight (Colbert and Schoch, 1998).
Stratigraphic and Geographic Distribution The genus has the longest stratigraphic range of all the North American tapirs, first appearing in the late Uintan and becoming extinct at the end of the early Hemingfordian. Protapirus simplex is reported from the Scenic Member of the Brule Formation, and P. obliquidens and P. validus are reported from the Poleslide Member of the Brule Formation of South Dakota (Wortman and Earle, 1893). The genus is also known from Oregon.
Natural History and Paleoecology Modern tapirs are forest animals and closely associated with water. It appears that they filled this adaptive niche of living in humid mesothermal regions with a diversity of plants that provided large quantities of forage early in their evolutionary history. As herbivores, they feed on small branches and leaves as well as fresh sprouts along with fallen fruit and water plants. They use their flexible proboscis to feed, and it allows them to selectively pull leaves, shoots, and fruits into their mouths. The evolutionary history and major changes in the skeleton are related to the refinement of the proboscis, molarization of the premolars, and a general increase in size, but the postcranial skeleton tends to remain conservative and similar in all taxa.
Systematics and Evolution The type species of Colodon is C. occidentalis from the Nebraska Territory. It was originally described by Leidy as Lophiodon, but the species was transferred to a new genus, Colodon, by Marsh. Currently the genus includes four other recognized species.
Distinctive Characters Colodon is considered a tapiroid (but not a true tapir) in the family Tapiridae by Colbert and Schloch (1998). The canines are small or absent, and the cheek teeth are short and wide. The upper first premolar has a lingual loph or cusp, but the other premolars are molariform with separated protocone and hypocone, and with the metaloph as prominent as the protoloph and connecting to the hypocone. The lower first premolar is absent, and the other premolars have wide talonids and a relatively large entoconid. The lower premolars have distinct posterior cusps that have not joined to form a loph. The third lower molar has a third lobe. The skull has a greatly enlarged narial incision and reduced nasals. Colbert (2005) noted that although Colodon is only about half the size of modern tapirs, the skull shared more characters of the skull with the modern genus Tapirus than the contemporary genus, Protapirus. These included several skeletal indicators that Colodon, like the living genus, probably had a prehensile proboscis: a retracted nasoincisive incisure similar to Tapirus, the occurrence of apomorphic fossae for cartilaginous meatal diverticula on the dorsal frontals and nasal, and ascending maxillae. Other similarities between Colodon and Tapirus include the telescoping of the skull, resulting from the anteroposterior shortening of the frontals, and the development of frontal sinuses; the conformation of the rostrolateral processes of the frontals; the descending processes of the nasals and the ascending process of the maxillae; and the apparent contact with the cartilaginous nasal septum by the premaxillae, as indicated by a CT scan of two species of Colodon from the Big Badlands (Colbert, 2005). As in Protapirus, the first digit is absent in the manus, but the bones of the fifth digit are more reduced in size.
Stratigraphic and Geographic Distribution The genus first appears in the early Uintan and the last definite appearance is in the Whitneyan, although possible records have been reported from the early Arikareean. In South Dakota, C. occidentalis is known from the Ahearn Member of the Chadron Formation, with questionable records from the Crazy Johnson and Peanut Peak members, and it is known from the Scenic and Poleslide members of the Brule Formation. Colodon occidentalis has also been reported from faunas in Nebraska and Saskatchewan. The genus is also known from Montana, Wyoming, California, and Oregon.
The family Equidae today is represented by a single genus, Equus, and includes horses, zebras, and donkeys and a diverse number of fossil genera. The primary characteristics of the family are the lack of separation of the foramen ovale and the optic foramen from the ethmoid and anterior lacerate foramen in the base of the skull. Members of the family have a diverse range of dentition, from brachyodont and bunodont to lophodont and hypsodont, reflecting their evolutionary transition from browsers to grazers. Despite the variety of tooth morphology, all members of the family have a structure called the postprotocrista on the upper third premolar (Hooker, 1989). In early horses, the morphology of the premolars is different from the molars, but with the appearance of Mesohippus in the White River Group, it and all subsequent horse genera have molarized premolars. The facial region tends to lengthen to accommodate the increasing height of the cheek teeth, resulting in a shift of the position of the orbit more posteriorly on the skull. Early equids have prominent facial fossae on the maxillae, but this becomes reduced and eventually lost in later genera. Although there is a general increase in body size through time (MacFadden, 1986), this is not true for all horse lineages, and some Pliocene (Blancan) genera are small and similar in size to horses from the White River Group.
5.47. Mesohippus bairdi, SDSM 2920. (A) Skull, right lateral view. (B) Mandible, right lateral view. (C) Mandible, occlusal view. (D) Skull, dorsal view. (E) Mesohippus sp. USNM V15898, skeleton. (F) Mesohippus bairdi, SDSM 2920, skull, occlusal view. Scales in centimeters. Photos by the authors of specimens from SDSM and USNM. SDSM 2920 is from the Museum of Geology, South Dakota School of Mines and Technology, Rapid City, South Dakota, U.S.A. USNM V15898, courtesy of Smithsonian Institution, Washington, D.C., U.S.A.
Like all perissodactyls, horses have a well-developed third trochanter on the femur. Early horses have a separate ulna and radius, but in advanced forms, the shaft of the ulna is reduced, leaving only the proximal and distal ends, which fuse to the enlarged radius. The fibula becomes reduced over time and is eventually lost in the modern genus, Equus. Another trend in the family is the continuous loss of the medial and lateral digits and increasing emphasis of the third metapodial to support the animal. Horses from the White River Group tend to be more primitive in their anatomy than later horses and have played a major role in understanding the evolution of horses as representing transitional forms between early and later members of the family.
Systematics and Evolution The type species of Mesohippus is M. bairdi, probably collected from Bear Creek, South Dakota. Nine species are currently recognized. There have been preliminary attempts to refine the species-level taxonomy of the genus, but additional work is required.
Distinctive Characters Mesohippus was a small animal with an estimated body mass of 42.2 kg for M. bairdi and 47.7 kg for M. barbouri (MacFadden, 1986). There is an increase in the size of the genus from the Chadron through the Brule. The geologically older and smaller individuals have hind legs longer then the front so the rump is higher than the shoulders, while in later forms the front and hind limbs are similar in length. The orbits are farther forward on the skull than in later horses and positioned above the first molar. There is no postorbital bar, and the orbit is confluent with the temporal fossa. There is a maxillary fossa in front of the lachrymal. Mesohippus retains the low-crowned (brachyodont) cheek teeth present in earlier horses, but is the first horse in which the premolars have become molariform (Fig. 5.47). The humerus has an undivided bicipital groove, a feature that evolves in later horses to facilitate sleeping upright (Hermanson and MacFadden, 1992), and the greater tuberosity extends medially over the bicipital groove. The ulna, although reduced in size, remains distinct from the radius. The fibula is complete but the distal end and often portions of the shaft fuse to the tibia (Scott, 1891). Some species of Mesohippus have four metacarpals but have reduced the number of metatarsals to three, so it is considered to be the first fully tridactyl horse (MacFadden, 1998).
Stratigraphic and Geographic Distribution The genus first appears in the Duchesnean and became extinct at the end of the early early Arikareean, although there are some questionable records for its presence in the late early Arikareean. Most specimens of Mesohippus from the White River Group are referred to M. bairdi, which is found in all members of the Chadron Formation. Mesohippus from the Scenic Member of the Brule are referred to a second species, M. westoni. Another species, M. exoletus, is reported from the Peanut Peak Member of the Chadron and the Scenic Member of the Brule, and M. barbouri is reported from the Scenic Member of the Brule Formation (MacFadden, 1998). Outside of the Big Badlands of South Dakota, the genus is widespread and is found throughout the western United States, including Wyoming, Nebraska, Colorado, Montana, North Dakota, Oregon, and Texas, then north into Canada and Saskatchewan. It is the first equid known in the southeastern United States and Florida (Patton, 1969).
Natural History and Paleoecology The diet of Mesohippus has been considered to be that of a browser, and Clark, Beerbower, and Kietzke (1967) considered it to be a member of the aquatic wet forest habitat of the White River Group. They considered Mesohippus, along with Merycoidodon, to be the dominant taxa of the gallery forests along river courses during the Orellan. Isotopic analysis of tooth enamel of Mesohippus from the Chadron and Brule formations in South Dakota produced carbon-13 values between −10 and −14.1 , indicating a diet of C3 vegetation. C3 grasses today grow at either higher latitudes or elevations that provide cooler environments but also grow in the understory of forests (Wang, Cerling, and MacFadden, 1994). The pure C3 signal in their tooth enamel supports the idea they were predominately browsers living in an environment dominated by C3 vegetation, such as trees, shrubs, and herbaceous dicots.
, indicating a diet of C3 vegetation. C3 grasses today grow at either higher latitudes or elevations that provide cooler environments but also grow in the understory of forests (Wang, Cerling, and MacFadden, 1994). The pure C3 signal in their tooth enamel supports the idea they were predominately browsers living in an environment dominated by C3 vegetation, such as trees, shrubs, and herbaceous dicots.
Systematics and Evolution The type species of Miohippus is M. annectens from the late Oligocene/early Miocene of John Day, Oregon. Numerous species have been described for this genus. Eighteen were listed by Stirton (1940), but currently six species are recognized.
Distinctive Characters Miohippus and Mesohippus are both present throughout the White River Group. The distinctions between the two genera are subtle. Miohippus is about the size of a sheep and is larger than most species of Mesohippus, with an estimated body mass for M. quartus of 53.8 kg (MacFadden, 1986). The primary difference used to distinguish the two genera is Miohippus has an articulation between the cuboid and third metatarsal, larger hypostyles on the teeth, a more elongate face, and a deeper facial fossa. The articulation between the third metatarsal and cuboid reflects a widening of the proximal end of the former bone and reflects its increase in size to support the animal with the reduction of the other digits (Prothero and Shubin, 1989).
Stratigraphic and Geographic Distribution Miohippus first appears in the early Chadronian and became extinct at the late Arikareean. In South Dakota, the type species and M. assiniboiensis have been reported from the Poleslide Member of the Brule Formation; M. obliquidens is present in the Peanut Peak Member of the Chadron Formation and all members of the Brule Formation; and M. gidleyi is reported from the Poleslide Member of the Brule Formation. The genus is also found in California, Oregon, Wyoming, Colorado, Nebraska, North Dakota, and Saskatchewan.
Natural History and Paleoecology Given the close morphological similarities between Mesohippus and Miohippus, as well as their overlapping geography and chronology, it is difficult to determine how they were ecologically different or how they may have filled different niches. Both would have been browsers but must have utilized different habitats on the landscape.
5.48. Hyracodon nebraskensis. (A) SDSM 28100, skull and jaws, left lateral view. (B) SDSM 28100, Skull, dorsal view. (C) USNM 336207, holotype, partial skull, occlusal view. (D) USNM 336207, holotype, left and right mandibles occlusal view. Scales in centimeters. Photos by the authors of specimens from SDSM and USNM. SDSM 28100 is from the Museum of Geology, South Dakota School of Mines and Technology, Rapid City, South Dakota, U.S.A. USNM 336207, courtesy of Smithsonian Institution, Washington, D.C., U.S.A.
This family of rhinoceros is known from Eurasia and North America from the middle Eocene to Oligocene, with five genera in North America. They are distinguished from other rhinos by their elongated limbs, suggesting a higher degree of cursoriality than their contemporaries. The skull tends to be primitive with a full dentition including all the incisors and canines, although there is incisor loss in some forms. The sagittal crest is well developed. The nasals are elongate, but there are no bosses indicative of the presence of horns. The rostrum and anterior part of the skull are laterally constricted. The dentition was more hypsodont than many of their contemporary mammalian taxa such as the horses and oreodonts. The upper third molar is reduced in size, unlike other rhinos. The pes is tridactyl, and the metapodials are elongated as part of the cursorial adaptation.
Systematics and Evolution The type species of Hyracodon is H. nebraskensis. Five species are currently recognized.
Distinctive Characters Hyracodon is the largest member of the family. In the Whitneyan, there were two contemporary species, H. nebraskensis and the larger H. leidyanus. Although the genus is among one of the longest lived, from approximately 10 Ma, there seems to have been little evolutionary change during this time. There is no indication that it had horns and no evidence that it was sexually dimorphic. The incisors are pointed and the canines are small. The first lower premolar is lost. The paracones on the premolars and molars are not separated posteriorly from the ectoloph (Fig. 5.48) (Prothero, 1998a).
5.49. Metamynodon planifrons, SDSM 3634. Skull, occlusal view. Scale in centimeters. Photo by the authors. SDSM 3634 is from the Museum of Geology, South Dakota School of Mines and Technology, Rapid City, South Dakota, U.S.A.
Stratigraphic and Geographic Distribution The genus appears in the Duchesnean and became extinct in the early early Arikareean, making the last member of the family in North America. In South Dakota, Hyracodon nebraskensis is present in all members of the Brule Formation, and H. leidyanus is present in the Poleslide Member of the Brule Formation. Hyracodon priscidens is present in the Peanut Peak Member of the Chadron Formation. The genus is also found in Utah, Wyoming, Colorado, Nebraska, Montana, North Dakota, Saskatchewan, and Texas.
Natural History and Paleoecology Remains of Hyracodon are fairly common in the White River Group, making this genus the best known of the family. No large quarry samples similar to other White River Group taxa have been found, so they may not have formed herds and like modern rhinos been primarily solitary in their habits. Hyracodon is ubiquitous in its distribution and has been found in deposits indicative of near-stream, swampy plains and open plains habitats.
It has been generally considered to be one of the most cursorial rhinos in terms of its skeletal anatomy. This interpretation has been challenged by Wall and Hickerson (1995) because the limb indices of Hyracodon do not compare to modern cursorial animals and are more similar to the hippopotamus and pigs. Clark, Beerbower, and Kietzke (1967) linked the decline of Hyracodon in the White River Group to the decrease of wet forest habitat and the increase in savanna and prairie open habitat. One would expect that if Hyracodon was cursorial in its adaptations, it would have increased in numbers with the expansion of open habitat.
The family Amynodontidae originated in Asia and dispersed into North America. This family of rhinocerotoids first appears in North America in the middle Eocene (late Bridgerian) and survived into the early Oligocene (Whitneyan). The family underwent a minor radiation in North America during the middle Eocene that seemed to coincide with the presence of lush tropical forest that existed in response to a homogenous equitable humid climate. They were medium to large in size and have been interpreted as semiaquatic in their habits. River channel deposits in the Orellan rocks of the White River Group have been named the Metamynodon channel sandstones. Their decline is attributed to the increasing aridity from the Eocene to Oligocene.
All members of the family have a large facial fossa on the skull. The size and location on the skull varies with different genera. There is a prominent sagittal crest, a primitive feature to all rhinos. All members of the family have greatly enlarged canines, and the size of the canines appears to be sexually dimorphic. There is a trend to reduce the number of premolars, and the upper and lower first premolar is absent in all forms. For most described amynodonts, the postcranial skeleton is unknown. The primitive build of the body seems to be subcursorial to mediportal, but metamynodontines are heavy limbed and barrel chested, and thus closely parallel features seen in the skeletons of hippos. Whether they had hippolike feeding habits, leaving the water to graze on land or feed on aquatic plants in the river, is unknown.
Systematics and Evolution The type species of Metamynodon is M. planifrons. In addition to the type species, two other species are considered valid.
Distinctive Characters The antorbital or facial fossa characteristic of members of the family is greatly reduced in Metamynodon. The orbits are positioned high on the skull, similar to hippos, supporting the notion of its semiaquatic habits. The second lower premolar is lost, and the third lower premolar is small or may be absent (Fig. 5.49).
Stratigraphic and Geographic Distribution The genus appears in the late Uintan and became extinct at the end of the Whitneyan. It has the longest stratigraphic range for the family and was the last genus of the family to become extinct. Metamynodon planifrons is found in all members of the Brule Formation. A second species, M. chadronensis, is known from the Peanut Peak Member of the Chadron Formation. The genus is also known from Mississippi, North Dakota, and Texas.
The name rhinoceros translates directly from Greek, “nose horn.” This feature is present in all five extant species of rhinos, although many extinct members of the family did not have a horn. Modern rhinos may have one horn or two aligned anteroposteriorly, growing from the top side of the nose. Unlike the horns of bovid artiodactyls, the horns of the rhinoceroses lack a bony core and are composed of compressed hair or keratin. They are attached to the skull via a bony boss, and the presence of this ossification on the nasals can be used to infer the presence of a horn in fossil forms. The family first appears in North America as immigrants from Asia, and it subsequently underwent a radiation, with 13 genera currently recognized. The family became extinct in North America at the end of the late Miocene (Hemphillian).
The skull of members of the family tends to be low and saddle shaped. The parietals support broad parasagittal crests. The nasals are long and slender and separated from the maxillae by a deep narial notch. While modern members of the family are characterized by the presence of a prominent horn or horns on the skull, many fossil members of the family lacked horns. A distinctive dental feature of the family is a chisel-shaped upper first incisor, which occludes with a tusklike lower second incisor. Except in the most primitive members of the family, all the other incisors, along with the canines, are lost. The upper molars have the typical pi pattern of lophs with a straight ectoloph. In primitive members of the family, the premolars are not fully molarized but become more so in advanced forms. In advanced forms, the teeth are more hypsodont, and they may develop additional internal ridges, often referred to as crochets, antecrochets, and cristae, that increase the surface area of the teeth and are an adaptation to ingesting more abrasive food.
5.50. Subhyracodon occidentalis, adapted from Scott and Jepsen (1941:plate 85, figs. 1, 2, 2a, 3). (A) Skull and jaws, right lateral view, AMNH 534, juvenile. (B) Skull, top view, AMNH 534, juvenile. (C) Skull, occlusal view, AMNH 534, juvenile. (D) Lower cheek teeth, p2–m3, YPM PU 13138.
5.51. Trigonias sp. (A) Skull, left lateral view, DMNS EPV.953. (B) Mandible (L), left lateral view, DMNS EPV.2674. (C) Mandible (L), occlusal view, DMNS EPV.2674. (D) Skull, dorsal view, DMNS EPV.953. (E) Skull, occlusal view, DMNS EPV.953. Scales in centimeters. Photos by the authors. Denver Museum of Nature and Science, Denver, Colorado, U.S.A. All rights reserved.
Bones of the postcranial skeleton tend to be massive and readily preserved as fossils. The ulna and radius remain separate, and a complete and separate fibula is retained even in advanced forms. Most rhinos are tridactyl, with digit 3 being the largest, although a functional fifth metacarpal is present in some genera.
Systematics and Evolution The type species of Subhyracodon is S. occidentalis was reported as coming from the “Missouri Territory,” now thought to be the Badlands of South Dakota. Two other species, S. mitis and S. kewi, are considered valid.
Distinctive Characters Subhyracodon is the largest of the true rhinos found in the Big Badlands and is the most common. Like many other fossil rhinos, it was hornless. It has lost the upper third incisor and canine, and the premaxillae are narrower and more delicate than in other contemporary rhinos. The premolar row is relatively shorter, and the upper second premolar has become molarized in comparison to earlier rhinos (Fig. 5.50). The limb bones are relatively heavy and more massive. The feet are tridactyl. One species of Subhyracodon evolved into Diceratherium in the Whitneyan, but the two genera coexist from the Whitneyan to early Arikareean, at which time Subhyracodon became extinct. One characteristic of the skull that distinguishes Diceratherium from Subhyracodon is the development of paired nasal ridges that suggest the presence of keratinous horns (Prothero, 1998c).
Stratigraphic and Geographic Distribution The genus first appears in the early Chadronian and survived until the early Arikareean. Subhyracodon occidentalis is present in the Scenic Member of the Brule Formation. Subhyracodon mitis is present in all members of the Chadron Formation. The two species are primarily distinguished by a difference in size, with S. mitis being the smaller of the two species. The genus is not only known from the Big Badlands of South Dakota but is also present in faunas in Wyoming, Colorado, Montana, North Dakota, Saskatchewan, and California, and it has also been found in Mississippi (Prothero, 2005).
5.52. Trigonias sp., DMNS 872. Skeleton, left lateral view. Composite specimen. Photo by the authors. Denver Museum of Nature and Science, Denver, Colorado, U.S.A. All rights reserved.
Natural History and Paleoecology Subhyracodon is particularly common in the Lower Nodular zone of the Scenic Member of the Brule Formation, which is Orellan in age. Numerous individuals of Subhyracodon have been recovered from an ancient water hole in Badlands National Park, called the Big Pig Dig. The large number of individuals probably accumulated over a short period of time, possibly during a drought. Many of the bones of Subhyracodon recovered from the Big Pig Dig show evidence of bite marks that suggest scavenging by the entelodont Archaeotherium. Besides the Big Pig Dig, a large number of individuals were recovered from the Nuttal Rhino Quarry and the Harvard Fossil Reserve in Wyoming and the Rockerville Site in South Dakota, as well as other localities in the Black Hills, including Wind Cave National Park. Rhino tracks referred to Subhyracodon preserved at Toadstool Geologic Park in Nebraska include numerous parallel trackways, suggesting that the genus may have formed small herds, unlike modern rhinos, which tend to be solitary.
As a larger-bodied rhino with more massive and stocky limb bones, Subhyracodon probably was not as cursorial as other forms such as the smaller Hyracodon. Its preferred habitat seems to have been riparian habitat along rivers, where it appears to have been a mixed feeder.
Systematics and Evolution The type species of Diceratherium is D. armatum from the John Day Formation of Oregon. Other recognized species of the genus include D. tridactylum, D. annectens, D. matutinum, and D. niobrarense. The genus evolved from Subhyracodon.
Distinctive Characters Unlike its ancestor, Subhyracodon, as the name suggests, Diceratherium had a pair of nose horns positioned side by side just before the tip of the nasals in males, as indicated by the rugosity of the bone. These are absent in the females. The skull is elongate, with a flat dorsal profile. It is also larger than Subhyracodon. The second to fourth premolars are more molariform than in Subhyracodon. The molars have strong cingula with simple crown patterns that lack either crochets or cristae. The limbs are long but the metapodials are short (Prothero, 2005).
Stratigraphic and Geographic Distribution Diceratherium is one of the longer-lived North American rhinos appearing in the Whitneyan; it did not become extinct until the Hemingfordian. Diceratherium armatum and D. annectens have been reported from the Peanut Peak Member of the Chadron Formation (Prothero, 1998c). The holotype of D. tridactylum is a complete skeleton of a female collected from the Protoceras beds, Poleslide Member of the Brule Formation (late Whitneyan) of South Dakota, and the species is known only from this unit in the Big Badlands. It is also known from California, Wyoming, Montana, Nebraska, Saskatchewan, and Oregon, as well as south to Arizona and Panama.
Systematics and Evolution The type of Trigonias is T. osborni from the Chadron Formation of the Big Badlands. A second species, T. wellsi, is also known from the Big Badlands. The primary difference between the two species of Trigonias is size, with T. osborni being the smaller of the two species (Prothero, 2005).
Distinctive Characters Trigonias is a medium-size rhino. It retains all of its anterior teeth except the lower third incisor and canine. The upper first incisor and lower second incisor show the typical rhino chisel/tusk modifications. There is no evidence for the presence of horns. The skull is elongate, with a low, saddle-shaped profile. The sagittal and occipital crests are moderately broad (Figs. 5.51, 5.52). The manus has a functional fifth metacarpal that is absent in most other genera (Prothero, 1998c).
Stratigraphic and Geographic Distribution Trigonias osborni is known from the Ahearn and Crazy Johnson members of the Chadron Formation. A second species, T. wellsi, is from the Ahearn Member of the Chadron Formation of the Badlands of South Dakota. The genus is known also from Nebraska, Colorado, Montana, Wyoming, California, and Saskatchewan.
Natural History and Paleoecology That the two species of Trigonias are known only from the Chadronian and have overlapping distributions is suggestive that they may represent males and females of the same species, although T. wellsi is known from a fewer number of localities. This may have been a more social species than other rhinos. A quarry site in Weld County, Colorado, produced multiple individuals.
Systematics and Evolution The type species of Penetrigonias is P. hudsoni from the upper part of the Chadron Formation at Chadron Flats, north of Toadstool Geologic Park, Sioux County, Nebraska. It is also known from Wyoming and the Ahearn Member of the Chadron Formation in the Big Badlands. Penetrigonias dakotensis is found in Wyoming, Colorado, and the Poleslide Member of the Brule Formation in the Big Badlands, while the third species, P. sagittatus, is known only from the Chadronian of the Cypress Hills of Saskatchewan.
Distinctive Characters This rhino is smaller than its contemporaries, Subhyracodon, Trigonias, or Amphicaenopus. The premaxilla is short but still retains all of its incisors, and it has the typical rhinocerotid incisor complex. There is some molarization of the premolars, but the degree to which it occurs is variable. The range of morphological variation present in the premolars resulted in the description of multiple species, taxonomically oversplitting the genus (Prothero, 1998c).
Stratigraphic and Geographic Distribution The genus has a long biostratigraphic range, first appearing in the Duchesnean and becoming extinct in the Whitneyan. Despite its long stratigraphic range, it has not yet been found in any Orellan faunas (Prothero, 2005). Penetrigonias dakotensis is known from the Poleslide Member of the Brule Formation in the Big Badlands of South Dakota and has been found in Wyoming and Colorado. Penetrigonias sagittatus is known only from the Chadronian of the Cypress Hills of Saskatchewan. The Duchesnean records are from Texas and Utah. Besides South Dakota, the Chadronian records are from Nebraska, Wyoming, Montana, and Colorado, and the single record from the Whitneyan is from the Protoceras beds of South Dakota.
Systematics and Evolution Amphicaenopus is only known from the type species, A. platycephalus, from the Whitneyan Protoceras channels of South Dakota.
Distinctive Characters This is a large rhino, larger than Subhyracodon and Diceratherium. The skull is broad and short with greatly expanded occipital crests. The lower second incisor is procumbent. The upper second and third premolars are not molariform, but the upper fourth premolar is nearly molariform. The nasals are short, and the notch between them and the maxilla that is commonly present in other rhinos is absent. The orbit is positioned farther forward on the skull, above the second molar, than in other rhinos (Prothero, 1998c).
Stratigraphic and Geographic Distribution The genus first appears in the early Chadronian and became extinct at the end of the Whitneyan. Like Penetrigonias, it has not been found in any Orellan faunas. In the Big Badlands of South Dakota, it is known from the Chadron Formation and the upper Poleslide Member of the Brule Formation (late Whitneyan).
Natural History and Paleoecology The body proportions of Amphicaenopus are hippolike and reminiscent of Metamynodon (Prothero, 2005). The lower incisors show striations due to wear, and Bjork (1978) suggested the presence of a flexible upper lip used to pull vegetation across the tusk.
5.53. Megacerops sp. (A) P12161, skull and jaws, right lateral view. (B) DMNS 3748, mandible, right lateral view. (C) DMNS 3747, mandible, occlusal view. (D) DMNS 3746, skull, occlusal view. Scales in centimeters. Photo (A) by William Simpson, Field Museum of Natural History; photos (B), (C), and (D) by the authors of specimens from DMNS. DMNS 3748, 3747, and 3746, Denver Museum of Nature and Science, Denver, Colorado, U.S.A. All rights reserved.
This extinct family, also sometimes referred to as titanotheres, first appears in the late Wasatchian and became extinct at the end of the Chadronian (the Eocene–Oligocene boundary). The phylogenetic relationship of this family to other perissodactyls has not been resolved. The family evolved in North America, but some forms dispersed into Asia, where some forms may have survived after the group became extinct in North America.
Early members of the family are small, but there is a trend of increasing size, and by the late Eocene, they are larger than modern rhinos and were the largest species in the fauna. Given the size of full-grown adults in the late Eocene, it is unlikely that any of the available predators, whether any of the five saber-toothed cats, creodonts, or even canids, preyed on these animals. Even juvenile brontotheres were disproportionately large compared to the contemporary predators and would have been difficult to kill.
While early forms had relatively long faces in proportion to overall skull size, in the later forms, the face is shortened while the skull posterior to the orbit has become greatly elongated. Later members of the family are characterized by their large body size and the development of prominent horns composed of bony extensions from the nasals and frontals. It is possible that the horns were covered with a keratinized skin, but there is no direct evidence for this. The size and robustness of the horns is sexually dimorphic, as is the body size, with males being significantly larger than females (Fig. 5.53A). Evidence that males used their horns on each other in in displays of dominance or during rut is provided by broken and healed ribs. In forms with large horns and skulls, the neck is short. Variation in the horns has been used as the basis for taxonomy in the group, resulting in a plethora of names being created since their initial discovery.
Brontotheres are interpreted as having been obligatory browsers on the basis of the morphology of their low-crowned dentition, which was used to either crush or shear vegetation. The incisors are small, are reduced in number, have a buttonlike crown, and do not show any signs of wear. While canines are present, they are short and do not project beyond the level of the cheek teeth. The reduction of the incisors and lack of wear suggests a prehensile lip, perhaps similar to that seen in living rhinos. The upper dentition has a characteristic W-shaped ectoloph separated from the two prominent lingual cusps. The lower molars have a distinctive M-shaped pattern formed by the fusion of the cusps into a single loph (Fig. 5.53C). The premolars never become molariform. They probably lived in warm temperate to subtropical environments that included forest to relative open woodland.
5.54. The first fossil discovered in the White River Badlands. Described by H. A. Prout of St. Louis, 1846–1847. Partial mandible of a brontothere. USNM 21820. Scale in centimeters. Photo by the authors. USNM 21820, courtesy of Smithsonian Institution, Washington, D.C., U.S.A.
As expected from the large size and massive body of the late Eocene forms, the bones of the postcranial skeleton are massive and robust. Modifications of the skeleton include graviportal features such as columnar limbs and reduced area of articulation between limb bones, indicating a reduced range of flexion and extension. The shaft of the femur has become flattened, and the third trochanter is reduced in size compared to the overall size of the animal. The ulna and radius remain separate, and the ulna is heavy in its build. A separate stout fibula is present. The manus has four digits; the pes has three. The arrangement of the bones in the manus and pes suggests that the feet had a pad of elastic tissue, similar to that seen in modern elephants and rhinos. As in modern elephants and rhinos, the unguals are reduced and would not have carried the weight of the animal.
Systematics and Evolution Five genera of brontotheres, all of large body size, have been described from the Chadronian: Megacerops, Brontops, Brontotherium, and Menodus/Menops. Each of these genera in turn had multiple species, 47 described by 1929 according to Mihlbachler, Lucas, and Emry (2004), primarily based on differences in horn structure, the differences of which may be due to sexual dimorphism, stage of ontogenetic development, or simple regional differences. The variation that has been described is apparently continuous among the variables commonly used to diagnose the Chadronian brontothere species. While Mader (1998) recognized three of the four genera as valid, Mihlbachler, Lucas, and Emry (2004) considered all of them, including the well-known genus Brontotherium, which Mader (1998) considered to be a junior synonym of Megacerops, to be one genus and the same as Megacerops, the oldest available name created by Leidy in 1870, with M. coloradensis as the genotypic species. The lower jaw (Figs. 1.2A, 5.54) described by Prout (1847) as the European taxon Palaeotherium is in fact a jaw of a brontothere and subsequently became the holotype specimen of Menodus giganteus (Pomel, 1849), which some authors consider to be the valid name for the Chadronian brontotheres but is considered a nomen dubium by Mihlbachler, Lucas, and Emry (2004). Not only did these different described taxa live during the same narrow time period, but many of the specimens placed in these different genera were collected from the same localities, further supporting the idea that the names created are based on variations within a single biological species. Currently only the type species of Megacerops, M. coloradensis, and one other species included in this genus are considered valid (Mihlbachler, Lucas, and Emry, 2004).
Distinctive Characters This is the largest member of the family. The number of incisors, both the uppers and lowers, is reduced to four or fewer and are globular in shape. On the basis of the reduction of the incisors, it has been inferred that Megacerops may have had a prehensile lip. There is no diastema, and the premolars are partially molarized. The posterior part of the zygomatic arch is thickened. The horns are short, anteriorly directed, and round to elliptical in cross section. The placement of the horns relative to the rest of the skull gives the skull a saddlelike appearance. The horns lack any evidence of grooves for blood vessels, which suggests they were covered with tough hide and lacked a keratinous covering. As previously noted, there is a wide variety of in the shape of the horns. Part of the variation is undoubtedly due to sexual dimorphism, and skeletons considered to be females are smaller than the males. This suggests that the horns may have also served as intraspecific displays, and once the species-level taxonomy is resolved, perhaps they will serve as species identification as well (Fig. 5.53).
Stratigraphic and Geographic Distribution Megacerops is known only from the Chadronian and is found in all members of the Chadron Formation. Regionally, it is known from South Dakota, Nebraska, Colorado, Montana, Wyoming, and Saskatchewan.
Natural History and Paleoecology Restricted to the Chadronian, Megacerops was the last brontothere in North America. On the basis of the dentition, the preferred habitat was probably forest or open woodland, which existed at that time. Their extinction at the end of the Eocene may reflect the changing climate, which included a general cooling and development of more open woodland habitat. Their disappearance at the end of the Chadronian makes them one of the primary taxa used to define the Eocene–Oligocene boundary.
The most distinctive features of Megacerops as the last brontothere are its extreme development of the horns, and that it appears to be sexually dimorphic. Osborn (1929) was the first to study the allometric relationship of the evolutionary development of the horns in brontotheres; his data were examined by Hersh (1934), who concluded that the evolutionary trend toward larger horns in the brontotheres leading to those seen in Megacerops was simply the result of selection for larger body size. The positive phylogenetic allometry between horn length and skull length is among the strongest known of such relationships in any group of vertebrates, and the seemingly disproportionate lengthening of bony frontonasal horns compared to an overall increase in the size of the skull has often been used as an illustration of a vertebrate macroevolutionary trend (McKinney and Schoch, 1985). The limited data available from juvenile specimens are inconclusive but indicate a strong role for accelerated horn growth late in ontogeny. On the basis of data derived from scaling patterns in modern ungulates, the increase in brontothere size was probably accompanied by an increase in individual life spans and longer developmental stages (Bales, 1996). The horns were apparently used for fighting and further strengthening of the skull; this activity is suggested by the expanded zygomatic arches. The horns seem to have functioned for frontal attacks, and two battling individuals could have locked horns in a test of strength and endurance, similar to that seen in bison and elk today (Mader, 1998). It appears that the primary target for attack was the side of the body, as broken and healed ribs are often present in skeletons.
In the White River Badlands, body fossils consist of mineralized bone and teeth. In contrast, trace fossils are burrows, tunnels, footprints, tooth marks, endocasts, nests, or egg chambers, which provide indirect evidence of animal activity (Fig. 5.55). Footprints can tell us how many animals were in an area, the direction they moved in, their size, and the speed they were going. Eggs and nests can provide information about ancient environments and climate (Lockley and Gillette, 1991). Trace fossils in terrestrial environments are uncommon because the agents that preserve them (lakes, ponds, streams, rivers) are not widespread.
In contrast to other types of trace fossils, evidence for fossil dung beetle activity is fairly common. Retallack (1984) described near spherical internal molds of silty micrite within the upper Scenic Member of the Brule Formation. Some of the spheres are irregular in shape and some are ellipsoidal. Many are slightly flattened. Under more powerful magnification, calcite crystal tubes were noted and were interpreted as fossil root traces. The sphere diameters ranged from 12.2 to 21.3 mm. These trace fossils are similar to the broad burrows made by scarabaeid beetles, which provision an underground nest with food for their developing larvae. The evidence for the use of dung is based on the following features: the irregular fibrous nature of the wall material, the high phosphorous content, and the abundance of large mammals that would provide the source of dung (Retallack, 1984). However, there is some debate regarding whether the dung beetle balls are the remains of a brood ball or are the mold of a pupal cell. Retallack (1984) was able to document deep vertical shafts 100 to 125 cm in length with short subhorizontal side passages 1 to 3 cm in length. The spherical internal molds were compared with the brood balls of modern scarabaeid beetles (Halffter and Matthews, 1966; Halffter, 1977). None of the fossils have thick, clayey shells; nor do they show any evidence of a former airspace around them. Nor is more than one near-spherical mold found at the end of each branch of the burrow (Retallack, 1984). The existence of dung beetles during the Oligocene time is indicated by the presence of the subfamily Scarabaeinae in the fossil record at other locations (Halffter, 1959; Balthasar, 1963; Matthews and Halffter, 1968; Wilson, 1977; Crowson, 1981; Grande, 1984).
5.55. Trace Fossils from the White River Badlands. (A) Brain endocast of Merycoidodon sp., BADL 930. (B) Rodent gnaw marks on bone, specimen found in Badlands National Park. (C) Photos of coprolites from Badlands National Park. Scales in centimeters. Photo (A) by the authors; photos (B) and (C) by the National Park Service of specimens from Badlands National Park. All specimens are the property of the U.S. government.
Although not as common as the beetle trace fossils, sweat bee cells provide important evidence of environments that were conducive to flowering plants. Retallack (1984) described tear-shaped internal molds of silty micrite from the same stratigraphic horizon as the beetle trace fossils. Upon closer examination, the tear-shaped molds had short entrance burrows of about 2 to 5 mm filled in with brown, silty claystone. The cells appear clustered around vertical shafts and were also filled with brown, silty claystone. Both the cells and the burrows had narrow crystal tubes interpreted as penetrating root traces (Retallack, 1983b). These tear-shaped molds were thought to have been created by sweat bees for the following reasons: the depth of the fossils within the paleosol, the clustering and horizontal orientation, the short entrance passages, the degree of organic coating, and the absence of spiral caps. These were all features found in other documented sweat bee nests (Apoidea, Halictidae, Halictinae) (Sakagami and Michener, 1962; Michener, 1974). Both wasps and bees were present during the Oligocene. A classic example are the bee fossils from the deposits from the Florissant Lake Beds, which are late Eocene in age.
Fossil trackways are a rare feature in the Big Badlands of South Dakota, with only three published occurrences (Chaffee, 1943; Lemley, 1971; Bjork, 1976). Chaffee (1943) reported brontothere tracks from the Chadron Formation and camelid tracks from the upper Brule Formation. There was some debate on whether the camelid tracks belonged to Poebrotherium or Pseudolabis (Bjork, 1976). Lemley (1971) published on a trackway from the Peanut Peak Member of the Chadron Formation, consisting of two parallel trackways about 17 feet in length, containing 24 tracks side by side. The prints are rounded and deep but were not assigned to a particular taxon beyond Mammalia (Lemley, 1971; Bjork, 1976).
Bjork (1976) describes a sandstone slab with six tracks representing two different mammals, proceeding in the same direction. The tracks are preserved as a negative, showing the sediment that filled the original tracks. Stratigraphically, the block was not found in place but is believed to be from the Poleslide Member of the Brule Formation, just above the Leptauchenia clays (Bjork, 1976). Four of the six tracks are from an artiodactyl approximately the size of a modern pronghorn. Bjork (1976) proposes that the mammal who made the tracks was a member of the Camelidae, possibly Pseudolabis dakotensis, Paratylopus primaevus, or Paratylopus matthewi. In addition to the camelid tracks, there are two poorly preserved carnivore tracks comprising the left front and hind foot. On the basis of the size and the extended claw marks, the track maker was believed to be Hesperocyon. It is not clear which animal came first. Bjork (1976) proposes the following scenario for the creation of the trackway. A shallow pool develops in the Leptauchenia clays, and the animals walk across the pool, leaving tracks in the mud. The tracks are buried by a sheet of sand.
In contrast to South Dakota, the Toadstool Geologic Park Trackway Site of western Nebraska is much more extensive (Nixon, 1996; LaGarry, Wells, and Terry, 1998). The trackway was discovered in the 1950s by Cecil Harvey (Harvey, 1960; Nixon, 1996; LaGarry, Wells, and Terry, 1998). The trackway consists of 75 sandstone slabs and 11 track morphologies. The site is believed to be a paleoshoreline along an ancient river system (LaGarry, Wells, and Terry, 1998). The site contains the tracks of a broad range of animals, including antelope, camels, entelodonts, birds, carnivores, rhinoceros, turtles, and invertebrates (Nixon, 1996; LaGarry, Wells, and Terry, 1998). As in South Dakota, the trackways are from the White River Group (Orella Member of the Brule Formation) and were created by the similar process of wet clays buried by a layer of sand (LaGarry, Wells, and Terry, 1998).
Coprolites, or fossilized excrement (Fig. 5.55C), have been well documented in the White River Badlands since the early twentieth century (Sinclair, 1921b; Wanless, 1923; Abel, 1926, 1935; Stovall and Strain, 1936; Lemley, 1971; Edwards and Yatkola, 1974; Retallack, 1983b). Coprolites from the White River Badlands are mostly from mammals, and more exist from carnivorous mammals than from herbivores (Edwards and Yatkola, 1974), although there have been two accounts of herbivore coprolites from the Chadron Formation (Stovall and Strain, 1936; Lemley, 1971). Coprolites from carnivorous mammals are characterized by elongate, cylindrical, and usually unsegmented masses. They can be tapered on one end and rough and ragged on the other (Edwards and Yatkola, 1974). This type of coprolite often contains bone and tooth fragments from undigested prey. Herbivore coprolites can be similar to modern bovids, with a disc shape and curling on the ends (Stovall and Strain, 1936; Lemley, 1971). The abundance of coprolites is not consistent throughout the late Eocene and Oligocene. Greater concentrations can be found within the Chadron Formation and the Scenic Member of the Brule Formation. According to Edwards and Yatkola (1974), this is due to the depositional environment. It appears that coprolite preservation requires rapid burial with fine-grained sediments in a low-energy fluvial environment. The sediments within the Poleslide member of the Brule Formation and the Sharps Formation are predominately eolian. Because of a cooler and dryer climate, there were fewer low-energy streams at that time. The channel deposits that make up portions of the Sharps Formation represent a high-energy environment that would quickly destroy any type of feces.