As far as we know, no other species on Earth has language; only humans talk. Sure, many animals communicate with each other in subtle and intricate ways. But we’re the only ones who gossip, take seminars, interview celebrities, convene board meetings, recite poems, negotiate treaties, conduct marriage ceremonies, hold criminal trials—all activities where just about the only thing going on is talking.

Fine, we also do many other things that our fellow Earth-creatures don’t. We play chess and soccer, sing the blues, go paragliding, design bridges, paint portraits, drive cars, and plant gardens, to name just a few. What makes language so special? Here’s the thing: language is deeply distinct from these other activities for the simple reason that all humans do it. There is no known society of Homo sapiens, past or present, in which people don’t talk to each other, though there are many societies where no one plays chess or designs bridges. And all individuals within any given human society talk, though again, many people don’t play chess or design bridges, for reasons of choice or aptitude.
So, language is one of the few things about us that appears to be a true defining trait of what it means to be human—so much so that it seems it must be part of our very DNA. But what, exactly, do our genes contribute? One view is that language is an innate instinct, something that we are inherently programmed to do, much as birds grow wings, elephants grow trunks, and female humans grow breasts. In its strongest version (for example, as argued by Steven Pinker in his 1994 book The Language Instinct), this nativist view says that not only do our genes endow us with a general capacity for language, they also lay out some of the general structures of language, the building blocks that go into it, the mental process of acquiring it, and so on. This view of language as a genetically driven instinct captures why it is that language is not only common to all humans but also is unique to humans—no “language genes,” no talking.
But many language researchers see it differently. The anti-nativist view is that language is not an innate instinct but a magnificent by-product of our impressive cognitive abilities. Humans alone learn language—not because we inherit a preprogrammed language template, but because we are the superlearners of the animal kingdom. What separates us from other animals is that our brains have evolved to become the equivalent of swift, powerful supercomputers compared with our fellow creatures, who are stuck with more rudimentary technology. Current computers can do qualitatively different things that older models could never aspire to accomplish. This supercomputer theory is one explanation for why we have language while squirrels and chimpanzees don’t.
But what about the fact that language is universal among humans, unlike chess or trombone-playing (accomplishments that, though uniquely human, are hardly universal)? Daniel Everett (2012), a linguist who takes a firm anti-nativist position, puts it this way in his book Language: The Cultural Tool: Maybe language is more like a tool invented by human beings than an innate behavior such as the dance of honeybees or the songs of nightingales. What makes language universal is that it’s an incredibly useful tool for solving certain problems that all humans have—foremost among them being how to efficiently transmit information to each other. Everett compares language to arrows. Arrows are nearly universal among hunter-gatherer societies, but few people would say that humans have genes that compel them to make arrows specifically, or to make them in a particular way. More likely, making arrows is just part of our general tool-making, problem-solving competence. Bows and arrows can be found in so many different societies because, at some point, people who didn’t grow their own protein had to figure out a way to catch protein that ran faster than they did. Because it was well within the bounds of human intelligence to solve this problem, humans inevitably did—just as, Everett argues, humans inevitably came to speak with each other as a way of achieving certain pressing goals.
The question of how we came to have language is a huge and fascinating one. If you’re hoping that the mystery will be solved by the end of this chapter, you’ll be sorely disappointed. It’s a question that has no agreed-upon answer among language scientists, and, as you’ll see, there’s a range of subtle and complex views among scientists beyond the two extreme positions I’ve just presented.
In truth, the various fields that make up the language sciences are not yet even in a position to be able to resolve the debate. To get there, we first need to answer questions like: What is language? What do all human languages have in common? What’s involved in learning it? What physical and mental machinery is needed to successfully speak, be understood, and understand someone else who’s speaking? What’s the role of genes in shaping any of these behaviors? Without doing a lot of detailed legwork to get a handle on all of these smaller pieces of the puzzle, any attempts to answer the larger question about the origins of language can only amount to something like a happy hour discussion—heated and entertaining, but ultimately not that convincing one way or the other. In fact, in 1866, the Linguistic Society of Paris decreed that no papers about the origins of language were allowed to be presented at its conferences. It might seem ludicrous that an academic society would banish an entire topic from discussion. But the decision was essentially a way of saying, “We’ll get nowhere talking about language origins until we learn more about language itself, so go learn something about language.”
A hundred and fifty years later, we now know quite a bit more about language, and by the end of this book, you’ll have a sense of the broad outlines of this body of knowledge. For now, we’re in a position to lay out at least a bit of what might be involved in answering the question of why people speak.
2.1 Why Us?
Let’s start by asking what it is about our language use that’s different from what animals do when they communicate. Is it different down to its fundamental core, or is it just a more sophisticated version of what animals are capable of? An interesting starting point might be the “dance language” of honeybees.
The language of bees
The dance language of honeybees was identified and described by Karl von Frisch (see von Frisch, 1967). When a worker bee finds a good source of flower nectar at some distance from her hive, she returns home to communicate its whereabouts to her fellow workers by performing a patterned waggle dance (see Figure 2.1). During this dance, she repetitively traces a specific path while shaking her body. The elements of this dance communicate at least three things:
1. The direction in which the nectar source is located. If the bee moves up toward the top of the hive, this indicates that the nectar source can be found by heading straight toward the sun. The angle of deviation away from a straight vertical path shows the direction relative to the sun.
2. The distance to the source. The longer the bee dances along the path from an initial starting point before returning to retrace the path again, the farther away the source is.
3. The quality of the source. If the bee has hit the nectar jackpot, she shakes with great vigor, whereas a lesser source of nectar elicits a more lethargic body wiggle.
Different bee species have different variations on this dance (for example, they might vary in how long they dance along a directional path to indicate a distance of 200 meters). It seems that bees have innate knowledge of their own particular dance “dialect,” and bees introduced into a hive populated by another species will dance in the manner of their genetic ancestors, not in the style of the adopted hive (though there’s some intriguing evidence that bees can learn to interpret foreign dialects of other bees; see Fu et al., 2008).
In some striking ways, the honeybee dance is similar to what we do in human language, which is presumably why von Frisch used the term language to describe it. The dance uses body movements to represent something in the real world, just as a map or a set of directions does. Human language also critically relies on symbolic representation to get off the ground—for us, it’s usually sequences of sounds made in the mouth (for example, “eat fruit”), rather than waggling body movements, that serve as the symbolic units that map onto things, actions, and events in the world. And, in both human languages and bee dances, a smaller number of communicative elements can be independently varied and combined to create a large number of messages. Just as bees can combine different intensities of wiggling with different angles and durations of the dance path, we can piece together different phrases to similar effect: “Go 3 miles northwest and you’ll find a pretty good Chinese restaurant”; or “There are some amazing raspberry bushes about 30 feet to your left.”
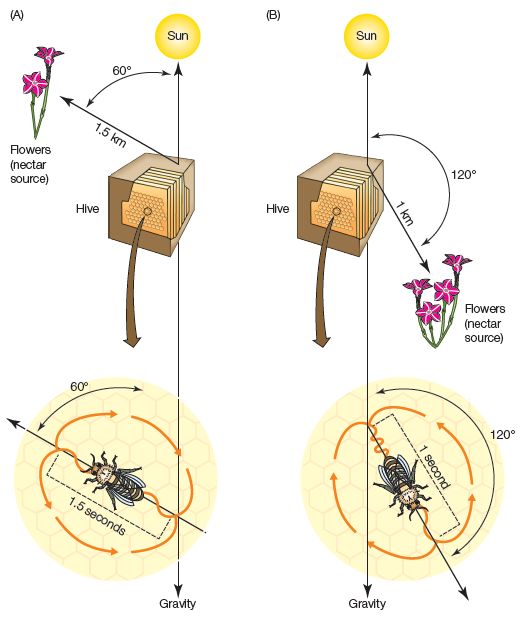
Figure 2.1 The waggle dance of honeybees is used by a returning worker bee to communicate the location and quality of a food source. The worker dances on the surface of the comb to convey information about the direction and distance of the food source, as shown in the examples here. (A) The nectar source is approximately 1.5 km from the hive flying at the indicated angle to the sun. (B) The nectar source is closer and the dance is shorter; in this case, the flowers will be found by flying away from the sun. The energy in the bee’s waggles (orange curves along the line of the dance) is in proportion to the perceived quality of the find.
Honeybee communicative behavior shows that a complex behavior capable of transmitting information about the real world can be encoded in the genes and innately specified, presumably through an evolutionary process. Like us, honeybees are highly cooperative and benefit from being able to communicate with each other. But bees are hardly among our closest genetic relatives, so it’s worth asking just how similar their communicative behavior is to ours. Along with the parallels I’ve just mentioned, there are also major differences.
Most importantly, bee communication operates within much more rigid parameters than human language. The elements in the dance, while symbolic in some sense, are still closely bound to the information that’s being communicated. The angle of the dance path describes the angle of the food source to the sun; the duration of the dance describes the distance to the food source. But in human language, there’s usually a purely arbitrary or accidental relationship between the communicative elements (that is, words and phrases) and the things they describe; the word fruit, for example, is not any more inherently fruit-like than the word leg. In this sense, what bees do is less like using words and more like drawing maps with their bodies. A map does involve symbolic representation, but the forms it uses are constrained by the information it conveys. In a map, there’s always some transparent, non-arbitrary way in which the spatial relations in the symbolic image relate to the real world. No one makes maps in which, for example, all objects colored red—regardless of where they’re placed in the image—are actually found in the northeast quadrant of the real-world space being described, while the color yellow is used to signal objects in the southwest quadrant, regardless of where they appear in the image.
Another severe limitation of bee dances is that bees only “talk” about one thing: where to find food (or water) sources. Human language, on the other hand, can be recruited to talk about an almost infinite variety of topics for a wide range of purposes, from giving directions, to making requests, to expressing sympathy, to issuing a promise, and so on. Finally, human language involves a complexity of structure that’s just not there in the bees’ dance language.
To help frame the discussion about how much overlap there is between animal communication systems and human language, the well-known linguist Charles Hockett listed a set of “design features” that he argued are common to all human languages. The full list of Hockett’s design features is given in Box 2.1; you may find it useful to refer back to this list as the course progresses. Even though some of the features are open to challenge, they provide a useful starting point for fleshing out what human language looks like.
Primate vocalizations
If we look at primates—much closer to us genetically than bees—a survey of their vocal communication shows a pretty limited repertoire. Monkeys and apes do make meaningful vocal sounds, but they don’t make very many, and the ones they use seem to be limited to very specific purposes. Strikingly absent are many of the features described by Hockett that allow for inventiveness, or the capacity to reuse elements in an open-ended way to communicate a varied assortment of messages.
For example, vervet monkeys produce a set of alarm calls to warn each other of nearby predators, with three distinct calls used to signal whether the predator is a leopard, an eagle, or a snake (Seyfarth et al., 1980). Vervets within earshot of these calls behave differently depending on the specific call: they run into trees if they hear the leopard call, look up if they hear the eagle call, and peer around in the grass when they hear the snake alarm. These calls do exhibit Hockett’s feature of semanticity, as well as an arbitrariness in the relationship between the signals and the meaning they transmit. But they clearly lack Hockett’s feature of displacement, since the calls are only used to warn about a clear and present danger and not, for example, to suggest to a fellow vervet that an eagle might be hidden in that tree branch up there, or to remind a fellow vervet that this was the place where we saw a snake the other day. There’s also no evidence of duality of patterning, in which each call would be made by combining similar units together in different ways. And vervets certainly don’t show any signs of productivity in their language, in which the calls are adapted to communicate new messages that have never been heard before but that can be easily understood by the hearer vervets. In fact, vervets don’t even seem to have the capacity to learn to make the various alarm calls; the sounds of the alarm calls are fixed from birth and are instinctively linked to certain categories of predators, though baby vervets do have to learn, for example, that the eagle alarm shouldn’t be made in response to a pigeon overhead. So, they come by these calls not through the process of cultural transmission, which is how humans learn words (no French child is born knowing that chien is the sound you make when you see a dog), but by being genetically wired to make specific sounds that are associated with specific meanings.
This last point has some very interesting implications. Throughout the animal world, it seems that the exact shape of a communicative message often has a strong genetic component. If we want to say that humans are genetically wired for language, then that genetic programming is going to have to be much more fluid and adaptable than that of other animals, allowing humans to learn a variety of languages through exposure. Instead of being programmed for a specific language, we’re born with the capacity to learn any language. This very fact might look like overwhelming support for the anti-nativist view, which says that language is simply an outgrowth of our general ability to learn complex things. But not necessarily. The position of nativists is more subtle than simply arguing that we’re born with knowledge of a specific language. Rather, the claim is that there are common structural ingredients to all human languages, and that it’s these basic building blocks of language that we’re all born with, whether we use them to learn French or Sanskrit. More on this later.
One striking aspect of primate vocalizations is the fact that monkeys and apes show much greater flexibility and capacity for learning when it comes to interpreting signals than in producing them. (A thorough discussion of this asymmetry can be found in a 2010 paper by primatologists Robert Seyfarth and Dorothy Cheney.) Oddly enough, even though vervets are born knowing which sounds to make in the presence of various predators, they don’t seem to be born with a solid understanding of the meanings of these alarms, at least as far as we can tell from their responses to the calls. It takes young vervets several months before they start showing the adult-like responses of looking up, searching in the grass, and so on. Early on, they respond to the alarm calls simply by running to their mothers, or reacting in some other way that doesn’t show that they know that an eagle call, for example, is used to warn specifically about bird-like predators. Over time, though, their ability to extend their understanding of new calls to new situations exceeds their adaptability in producing calls. For instance, vervets can learn to understand the meanings of alarm calls of other species, as well as the calls of their predators—again, even though they never learn to produce the calls of other species.
Seyfarth and Cheney suggest that the information that primates can pull out from the communicative signals they hear can be very subtle. An especially intriguing example comes from an experiment involving the call behavior of baboons. Baboons, as it happens, have a very strict status hierarchy within their groups, and it’s not unusual for a higher-status baboon to try to intimidate a lower-status baboon by issuing a threat-grunt, to which the lower-ranking animal usually responds with a scream. The vocalizations of individual baboons are distinctive enough that they’re easily recognized by all members of the group. For the purpose of the study, the researchers created a set of auditory stimuli in which they cut and spliced together prerecorded threat-grunts and screams from various baboons within the group. The sounds were reassembled so that sometimes the threat-call of a baboon was followed by a scream from a baboon higher up in the status hierarchy. The eavesdropping baboons reacted to this pairing of sounds with surprise, which seems to show that the baboons had inferred from the unusual sequence of sounds that a lower-status animal was trying to intimidate a higher-status animal—and understood that this was a bizarre state of affairs.
It may seem strange that animals’ ability to understand something about the world based on a communicative sound is so much more impressive than their ability to convey something about the world by creating a sound. But this asymmetry seems rampant within the animal kingdom. Many dog owners are intimately familiar with this fact. It’s not hard to get your dog to recognize and respond to dozens of verbal commands; it’s getting your dog to talk back to you that’s difficult. Any account of the evolution of language will have to grapple with the fact that speaking and understanding are not necessarily just the mirror image of each other.
Can language be taught to apes?
As you’ve seen, when left to themselves in the wild, non-human primates don’t indulge in much language-like vocalization. This would suggest that the linguistic capabilities of humans and other primates are markedly different. Still, a non-nativist might object and argue that looking at what monkeys and apes do among themselves, without the benefit of any exposure to real language, doesn’t really provide a realistic picture of what they can learn about language. After all, when we evaluate human infants’ capacity for language, we don’t normally separate them from competent language users—in other words, adults—and see what they come up with on their own. Suppose language really is more like a tool than an instinct, with each generation of humans benefiting from the knowledge of the previous generation. In that case, to see whether primates are truly capable of attaining language, we need to see what they can learn when they’re allowed to have many rich interactions with individuals who have already solved the problem of language.
This line of thinking has led to a number of studies that have looked at how apes communicate, not with other non-linguistic apes, but with their more verbose human relatives. In these studies, research scientists and their assistants have raised young apes (i.e., chimpanzees, bonobos, orangutans, and gorillas) among humans in a language-rich environment. Some of the studies have included intensive formal teaching sessions, with a heavy emphasis on rewarding and shaping communicative behavior, while other researchers have raised the apes much as one would a human child, letting them learn language through observation and interaction. Such studies often raise tricky methodological challenges, as discussed in Method 2.1. For example, what kind of evidence is needed to conclude that apes know the meaning of a word in the sense that humans understand that word? Nevertheless, a number of interesting findings have come from this body of work (a brief summary can be found in a review article by Kathleen Gibson, 2012).
First, environment matters: there’s no doubt that the communicative behavior of apes raised in human environments starts to look a lot more human-like than that of apes in the wild. For example, a number of apes of several different species have mastered hundreds of words or arbitrary symbols. They spontaneously use these symbols to communicate a variety of functions—not just to request objects or food that they want, but also to comment on the world around them. They also refer to objects that are not physically present at the time, showing evidence of Hockett’s feature of displacement, which was conspicuously absent from the wild vervets’ alarm calls. They can even use their symbolic skills to lie—for instance, one chimp was found to regularly blame the messes she made on others. Perhaps even more impressively, all of the species studied have shown at least some suggestion of another of Hockett’s features, productivity—that is, of using the symbols they know in new combinations to communicate ideas for which they don’t already have symbols. For example, Koko, a gorilla, created the combination “finger bracelet” to refer to a ring; Washoe, a chimpanzee, called a Brazil nut a “rock berry.” Sequences of verbs and nouns often come to be used by apes in somewhat systematic sequences, suggesting that the order of combination isn’t random.
As in the wild, trained apes show that they can master comprehension skills much more readily than they achieve the production of language-like units. In particular, it quickly became obvious that trying to teach apes to use vocal sounds to represent meanings wasn’t getting anywhere. Apes, it turns out, have extremely limited control over their vocalizations and simply can’t articulate different-sounding words. But the trained apes were able to build up a sizable vocabulary when signed language was substituted for spoken language, or when researchers adopted custom-made artificial “languages” using visual symbols arranged in systematic structures. This raises the very interesting question of how closely the evolution of language is specifically tied to the evolution of speech, an issue we will probe in more detail in Section 2.5.
But even with non-vocal languages, the apes were able to handle much more complexity in their understanding of language than in their production of it. They rarely produced more than two or three symbols strung together, but several apes were able to understand commands like “make the doggie bite the snake,” and they could distinguish that from “make the snake bite the doggie.” They could also follow commands that involved moving objects to or from specific locations. Sarah, a chimpanzee, could reportedly even understand “if/then” statements.
Looking at this collection of results, it becomes apparent that with the benefit of human teachers, ape communication takes a great leap toward human language—human-reared apes don’t just acquire more words or symbols than they do in the wild, they also show that they can master a number of Hockett’s design features that are completely absent from their naturalistic behavior. This is very revealing, because it helps to answer the question of when some of these features of human language—or rather, the capability for these features—might have evolved.
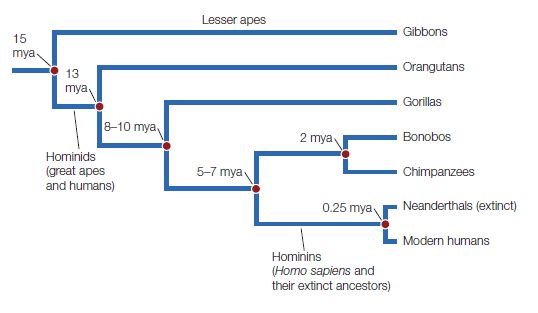
Figure 2.2 The evolutionary history of hominids. The term hominids refers to the group consisting of all modern and extinct great apes (including humans and their more immediate ancestors). This evolutionary tree illustrates the common ancestral history and approximate times of divergence of hominins (including modern humans and the now-extinct Neanderthals) from the other great apes. Note that a number of extinct hominin species are not represented here.
Biologists estimate that humans, chimpanzees, and bonobos shared a common ancestor between 5 and 7 million years ago. The last common ancestor with gorillas probably occurred between 8 and 10 million years ago, and the shared ancestor with orangutans even earlier than that (see Figure 2.2). Evidence about when the features of human language evolved helps to answer questions about whether they evolved specifically because these features support language.
Among nativists, the most common view is that humans have some innate capabilities for language that evolved as adaptations. Evolutionary adaptations are genetically transmitted traits that give their bearers an advantage—specifically, an adaptive trait helps individuals with that trait to stay alive long enough to reproduce and/or to have many offspring. The gene for the advantageous trait spreads throughout a population, as over time members of the species with that trait will out-survive and out-reproduce the members without that trait. But not all adaptations that help us to use language necessarily came about because they gave our ancestors a communicative edge over their peers. Think about it like this: humans have hands that are capable of playing the piano, given instruction and practice. But that doesn’t mean that our hands evolved as they did because playing the piano gave our ancestors an advantage over non-piano-playing humans. Presumably, our nimble fingers came about as a result of various adaptations, but the advantages these adaptations provided had nothing to do with playing the piano. Rather, they were the result of the general benefits of having dexterous hands that could easily manipulate a variety of objects. Once in possession of enhanced manual agility, however, humans discovered that hands can be put to many wonderful uses that don’t necessarily help us survive into adulthood or breed successfully.
The piano-playing analogy may help to make sense of the question, “If language-related capabilities evolved long before humans diverged from other apes, then why do only humans make use of them in their natural environments?” That is, if apes are capable of amassing bulky vocabularies and using them creatively, why are they such linguistic underachievers in the wild? The contrast between their communicative potential and their lack of spontaneous language in the wild suggests that certain cognitive skills that are required to master language—at least, those skills that are within the mental grasp of apes—didn’t necessarily evolve for language. Left to their own devices, apes don’t appear to use these skills for the purpose of communicating with each other. But when the cultural environment calls for it, these skills can be recruited in the service of language—much as in the right cultural context, humans can use their hands to play the piano. This state of affairs poses a challenge to the “language-as-instinct” view.
Nevertheless, it’s entirely possible that the skills that support language fall into two categories: (1) those that are necessary to get language off the ground but aren’t really specific to language; and (2) traits that evolved particularly because they make language more powerful and efficient. It may be that we share the skills in the first category with our primate relatives, but that only humans began to use those skills for the purpose of communication. Once this happened, there may have been selective pressure on other traits that provided an additional boost to the expressive capacity of language—and it’s possible that these later skills are both language-specific and uniquely human.
It seems, then, that when we talk about language evolution, it doesn’t make sense to treat language as an all-or-nothing phenomenon. Language may well involve a number of very different cognitive skills, with different evolutionary trajectories and different relationships to other, non-linguistic abilities.
Throughout this book, you’ll get a much more intimate sense of the different cognitive skills that go into human language knowledge and use. As a first step, this chapter starts by breaking things down into three very general categories of language-related abilities: the ability to understand communicative intent, a grasp of linguistic structure, and the ability to control voice and/or gesture.
2.2 The Social Underpinnings of Language
Imagine this scene from long ago: an early hominid is sitting at the mouth of his cave with his female companion when a loud roar tears through the night air. He nods soberly and says, “Leopard.” This is a word that he’s just invented to refer to that animal. In fact, it’s the first word that’s passed between them, as our male character is one of language’s very earliest adopters. It’s a breakthrough: from here on, the couple can use the word to report leopard sightings to each other, or to warn their children about the dangerous predator. But none of this can happen unless the female can clue in to the fact that the sounds in leopard were intentionally formed to communicate an idea—they were not due to a sneeze or a cough, or some random set of sounds. What’s more, she has to be able to connect these intentional and communicative sounds with what’s going on around her, and make a reasonable guess about what her companion is most likely to be trying to convey.
From your perspective as a modern human, all of this may seem pretty obvious, requiring no special abilities. But it’s far from straightforward, as revealed by some very surprising tests that chimpanzees fail at miserably, despite their substantial intellectual gifts. For example, try this next time you meet a chimp: Show the animal a piece of food, and then put it in one of two opaque containers. Shuffle the two containers around so as to make it hard to tell where it’s hidden. Now stop, and point to the container with the food. The chimpanzee will likely choose randomly between the two containers, totally oblivious to the very helpful clue you’ve been kind enough to provide. This is exactly what Michael Tomasello (2006) and his colleagues found when they used a similar test with chimpanzees. Their primate subjects ignored the conspicuous hint, even though the experimenters went out of their way to establish that the “helper” who pointed had indeed proven herself to be helpful on earlier occasions by tilting the containers so that the chimp could see which container had the food (information that the chimps had no trouble seizing upon).
Understanding the communicative urge
Chimpanzees’ failure to follow a pointing cue is startling because chimps are very smart, perfectly capable of making subtle inferences in similar situations. For example, if an experimenter puts food in one of two containers and then shakes one of them but the shaken container produces no rattling sound, the chimpanzee knows to choose the other one (Call, 2004). Or, consider this variation: Brian Hare and Michael Tomasello (2004) set up a competitive situation between chimpanzees and a human experimenter, with both human and chimp trying to retrieve food from buckets. If the human extended her arm toward a bucket but couldn’t touch it because she had to stick her hand through a hole that didn’t allow her to reach far enough, the chimpanzees were able to infer that this was the bucket that must contain the food, and reached for it. Why can chimpanzees make this inference, which involves figuring out the human’s intended—but thwarted—goal, but not be able to understand pointing? Tomasello and his colleagues argued that, although chimpanzees can often understand the intentions and goals—and even the knowledge states—of other primates, what they can’t do is understand that pointing involves an intention to communicate. In other words, they don’t get that the pointing behavior is something that’s done not just for the purpose of satisfying the pointer’s goal, but to help the chimpanzee satisfy its goal).
To some researchers, it’s exactly this ability to understand communicative intentions that represents the “magic moment” in the evolution of language, when our ancestors’ evolutionary paths veered off from those of other great apes, and their cognitive skills and motivational drives came to be refined, either specifically for the purpose of communication, or more generally to support complex social coordination.
Some language scientists have argued that a rich communication system is built on a foundation of advanced skills in social cognition, and that among humans these skills evolved in a super-accelerated way, far outpacing other gains we made in overall intelligence and working memory capacity. To test this claim, Esther Hermann and her colleagues (2007) compared the cognitive abilities of adult chimpanzees, adult orangutans, and human toddlers aged two and a half. All of these primates were given a battery of tests evaluating two kinds of cognitive skills: those needed for understanding the physical world, and those for understanding the social world. For example, a test item in the physical world category might involve discriminating between a smaller and a larger quantity of some desirable reward, or locating the reward after it had been moved, or using a stick to retrieve an out-of-reach reward. The socially oriented test items looked for accomplishments like solving a problem by imitating someone else’s solution, following a person’s eye gaze to find a reward, or using or interpreting communicative gestures to locate a reward. The researchers found that in demonstrating their mastery over the physical world, the human toddlers and adult chimpanzees were about even with each other, and slightly ahead of the adult orangutans. But when it came to the social test items, the young humans left their fellow primates in the dust (with chimps and orangutans showing similar performance).
There’s quite a bit of additional evidence showing that even very young humans behave in ways that are different from how other primates act in similar situations. For example, when there’s nothing obvious in it for themselves, apes don’t seem to be inclined to communicate with other apes for the purpose of helping the others achieve a goal. But little humans almost feel compelled to. In one study by Ulf Lizskowski and colleagues (2008), 12-month-olds who hadn’t yet begun to talk watched while an adult sat at a table stapling papers without involving the child in any way. At one point, the adult left the room, then another person came in, moved the stapler from the table to a nearby shelf, and left. A little later, the first adult came back, looked around, and made quizzical gestures to the child. In response, most of the children pointed to the stapler in its new location. According to Michael Tomasello (2006), apes never point with each other, and when they do “point” to communicate with humans (usually without extending the index finger), it’s because they want the human to fetch or hand them something that’s out of their reach.
Skills for a complex social world
So, humans are inclined to share information with one another, whereas other primates seem not to have discovered the vast benefits of doing so. What’s preventing our evolutionary cousins from cooperating in this way? One possibility is that they’re simply less motivated to engage in complex social behavior than we humans are. Among mammals, we as a species are very unusual in the amount of importance we place on social behavior. For example, chimpanzees are considerably less altruistic than humans when it comes to sharing food, and they don’t seem to care as much about norms of reciprocity or fairness. When children are in a situation where one child is dividing up treats to share and extends an offer that is much smaller than the share he’s claimed for himself, the other child is apt to reject the offer, preferring to give it up in order to make the point that the meager amount is an affront to fairness. A chimp will take what it can get (Tomasello, 2009).
In fact, when you think about the daily life of most humans in comparison to a day in the life of a chimpanzee, it becomes apparent that our human experiences are shaped very profoundly by a layer of social reality, while a chimpanzee may be more grounded in the physical realities of its environment. In his book Why We Cooperate (2009), Michael Tomasello points out how different the human experience of shopping is from the chimpanzee’s experience of foraging for food:
Let us suppose a scenario as follows. We enter a store, pick up a few items, stand in line at the checkout, hand the clerk a credit card to pay, take our items, and leave. This could be described in chimpanzee terms fairly simply as going somewhere, fetching objects, and returning from whence one came. But humans understand shopping, more or less explicitly, on a whole other level, on the level of institutional reality. First, entering the store subjects me to a whole set of rights and obligations: I have the right to purchase items for the posted price, and the obligation not to steal or destroy items, because they are the property of the store owner. Second, I can expect the items to be safe to eat because the government has a department that ensures this; if a good proves unsafe, I can sue someone. Third, money has a whole institutional structure behind it that everyone trusts so much that they hand goods over for this special paper, or even for electronic marks somewhere from my credit card. Fourth, I stand in line in deference to widely held norms, and if I try to jump the line people will rebuke me, I will feel guilty, and my reputation as a nice person will suffer. I could go on listing, practically indefinitely, all of the institutional realities inhibiting the public sphere, realities that foraging chimpanzees presumably do not experience at all.
Put in these terms, it becomes obvious that in order to successfully navigate through the human world, we need to have a level of social aptitude that chimpanzees manage without.
At some level, the same socially oriented leanings that drive humans to “invent” things like laws and money also make it possible for them to communicate through language. Language, law, and currency all require people to buy into an artificial system that exists only because everyone agrees to abide by it. Think about it: unlike vervets with their alarm calls, we’re not genetically driven to produce specific sounds triggered by specific aspects of our environment. Nor do our words have any natural connection to the world, in the way that honeybee dance language does. Our words are quite literally figments of human imagination, and they have meaning only because we all agree to use the same word for the same thing.
But it may not just be an issue of general social motivation that’s keeping our primate relatives from creating languages or laws of their own. It’s possible that they also lack a specific cognitive ingredient that would allow them to engage in complex social coordination. In order to do something as basic as make a smart guess about what another person’s voluntary mouth noises might be intended to mean, humans needed to have the capacity for joint attention: the awareness between two (or more) individuals that they are both paying attention to the same thing. Again, this doesn’t seem especially difficult, but Tomasello and his colleagues have argued that, to any reliable extent, this capacity is found only in humans. Chimps can easily track the gaze of a human or another ape to check out what’s holding the interest of the other; they can also keep track of what the others know or have seen. In other words, chimps can know what others know. But there’s no clear evidence that they participate in situations where Chimp A knows that Chimp B knows that Chimp A is staring at the same thing. Presumably, this is exactly the kind of attunement that our ancestors sitting by their caves would have needed to have in order to agree on a word for the concept of leopard.
It turns out that joint attention skills are very much in evidence in extremely young humans. Toward the end of their first year (on average), babies unambiguously direct other people’s attention to objects by pointing, often with elaborate vocalization. Months prior to this, they often respond appropriately when others try to direct their attention by pointing or getting them to look at something specific. It’s especially relevant that success with language seems to be closely linked to the degree to which children get a handle on joint attention. For example, Michael Morales and his colleagues (2000) tracked a group of children from 6 to 30 months of age. The researchers tested how often individual babies responded to their parents’ attempts to engage them in joint attention, beginning at 6 months of age; they then evaluated the size of the children’s vocabularies at 30 months and found that the more responsive the babies were even at 6 months, the larger their vocabularies were later on (see Researchers at Work 2.1). Another study by Cristina Colonnesi and her colleagues (2010) documented evidence of a connection between children’s pointing behaviors and the emergence of language skills. Interestingly, the connection was apparent for declarative pointing—that is, pointing for the purpose of “commenting” on an object—but not for imperative pointing to direct someone to do something. This is intriguing because when apes do communicate with humans, they seem to do much less commenting and much more directing of actions than human children do, even early in their communication. Both are very clearly communicative acts, and yet they may have different implications for linguistic sophistication.
It’s increasingly apparent, then, that being able to take part in complex social activities that rely on mutual coordination is closely tied to the emergence of language. Researchers like Michael Tomasello have argued that there is a sharp distinction between humans and other apes when it comes to these abilities. But there’s controversy over just how sharp this distinction is. It’s also not obvious whether these abilities are all genetically determined or whether skills such as joint attention also result from the deep and ongoing socialization that almost every human is subject to. What should we make, for example, of the fact that apes raised by humans are able to engage in much more sophisticated communication—including pointing—than apes raised by non-humans? Furthermore, we don’t have a tremendous amount of detailed data about apes’ capabilities for social cognition. Much research has focused on the limitations of non-human primates, but newer studies often show surprisingly good social abilities. For instance, many scientists used to think that chimpanzees weren’t able to represent the mental states of others, but it now appears that they’re better at it than had been thought (see Method 2.2; Call & Tomasello, 2010). So, while it’s clear that very young humans display impressive social intelligence compared to the non-human primates that are typically studied in experiments, we don’t really know how much of this skill comes from our biological evolutionary history and how much of it comes from our cultural heritage.
But whether or not the rich social skills of humans reveal a uniquely human adaptation, and whether or not these adaptations occurred largely to support communication (or more generally to support complex social activity), there are other skills that we need in order to be able to command the full expressive power of language. These other skills, in turn, may or may not be rooted in uniquely human adaptations for language, a theme we’ll take up in the coming section.
2.3 The Structure of Language
Being able to settle on arbitrary symbols as stand-ins for meaning is just one part of the language puzzle. There’s another very important aspect to language, and it’s all about the art of combination.
Combining units
Combining smaller elements to make larger linguistic units takes place at two levels. The first level deals with making words from sounds. In principle, we could choose to communicate with each other by creating completely different sounds as symbols for our intended meanings—a high-pitched yowl might mean “arm,” a guttural purr might mean “broccoli,” a yodel might mean “smile,” and so on. In fact, this is very much how vervets use sound for their alarm calls. But at some point, we might find we’d exhausted our ability to invent new sounds but still had meanings we wanted to express. To get around this limitation, we can take a different approach to making words: simply use a relatively small number of sounds, and repurpose them by combining them in new and interesting ways. For example, if we take just ten different sounds to create words made up of five sounds each without repeating any of the sounds within a word, we can end up with a collection of more than 30,000 words. This nifty trick illustrates Hockett’s notion of duality of patterning, in which a small number of units that don’t convey meanings on their own can be used to create a very large number of meaningful symbols. In spoken language, we can take a few meaningless units like sounds (notice, for example, that there are no specific meanings associated with the sounds of the letters p, a, and t) and combine them into a number of meaningful words. Needless to say, it’s a sensible approach if you’re trying to build a beefy vocabulary.
But the combinatorial tricks don’t end there. There may be times when we’d like to communicate something more complex than just the concepts of leopard or broccoli. We may want to convey, for instance, that Johnny kicked Freddy in the shin really hard, or that Simon promised to bake Jennifer a cake for her birthday. What are the options open to us? Well, we could invent a different word for every complex idea, so a sequence of sounds like beflo would communicate the first of these complex ideas, and another—say, gromi—would communicate the second. But that means that we’d need a truly enormous vocabulary, essentially containing a separate word for each idea we might ever want to communicate. Learning such a vocabulary would be difficult or impossible—at some point in the learning process, everyone using the language would need to have the opportunity to figure out what beflo and gromi meant. This means that as a language learner, you’d have to find yourself in situations (probably more than once) in which it was clear that the speaker wanted to communicate the specific complex idea that gromi was supposed to encode. If such a situation happened to never arise, you’d be out of luck as far as learning that particular word.
A more efficient solution would be to combine meaningful elements (such as separate words) to make other, larger meaningful elements. Even better would be to combine them in such a way that the meaning of the complex idea could be easily understood from the way in which the words are combined—so that rather than simply tossing together words like Jennifer, birthday, promised, bake, Simon, and cake, and leaving the hearer to figure out how they relate to each other, it would be good to have some structured way of assembling sentences out of their component parts that would make their meanings clear from their structure. This added element of predictability of meaning requires a syntax—a set of “rules” about how to combine meaningful units together in systematic ways so that their meanings can be transparent. (For example, once we have a syntax in place, we can easily differentiate between the meanings of Simon promised Jennifer to bake a birthday cake and Jennifer promised Simon to bake a birthday cake.) Once we’ve added this second level into our communication system, not only have we removed the need to learn and memorize separate words for complex ideas, but we’ve also introduced the possibility of combining the existing elements of our language to talk about ideas that have never before been expressed by anyone.
Structured patterns
It should be obvious that the possibility of combining elements in these two ways gives language an enormous amount of expressive power. But it also has some interesting consequences. Now, anyone learning a language has to be able to learn its underlying structural patterns. And since, for the most part, human children don’t seem to learn their native language by having their parents or teachers explicitly teach them the rules of language (the way, for example, they learn the rules of arithmetic), they have to somehow intuit the structures on their own, simply by hearing many examples of different sentences. You might think of language learning as being a lot like the process of reverse engineering a computer program: Suppose you wanted to replicate some software, but you didn’t have access to the code. You could try to deduce what the underlying code looked like by analyzing how the program behaved under different conditions. Needless to say, the more complicated the program, the more time you’d need to spend testing what it did.
In case you’re tempted to think that language is a fairly simple program, I invite you to spend a few hours trying to characterize the structure of your own native tongue (and see Box 2.2). The syntactic structures of human languages involve a lot more than just basic word orders. Once you start looking up close, the rules of language require some extremely subtle and detailed knowledge. For example, how come you can say:
Who did the hired assassin kill the mayor for?
meaning “Who wanted the mayor dead?” But you can’t say:
Who did the hired assassin kill the mayor and?
intending to mean “Aside from the mayor, who else did the hired assassin kill?” Or, consider the two sentences:
Naheed is eager to please.
Naheed is easy to please.
These two sentences look almost identical, so why does the first involve Naheed pleasing someone else, while the second involves someone else pleasing Naheed? Or, how is it that sometimes the same sentence can have two very different meanings? As in:
Smoking is more dangerous for women than men.
meaning either that smoking is more dangerous for women than it is for men, or that smoking is even more hazardous than men are for women.
It’s not only in the area of syntax that kids have to acquire specific knowledge about how units can be put together. This is the case for sound combinations as well. Languages don’t allow sounds to be combined in just any sequence whatsoever. There are constraints. For instance, take the sounds that we normally associate with the letters r, p, m, s, t, and o. If there were no restrictions on sequencing, these could be combined in ways such as mprots, stromp, spormt, tromps, rpmsto, tormps, torpsm, ospmtr, and many others. But not all of these “sound” equally good as words, and if you and your classmates were to rank them from best- to worst-sounding, the list would be far from random.
Here’s another bit of knowledge that English speakers have somehow picked up: even though they’re represented by the same letter, the last sound in fats is different from the last sound in fads (the latter actually sounds like the way we usually say the letter z). This is part of a general pattern in English, a pattern that’s clearly been internalized by its learners: if I were to ask any adult native speaker of English exactly how to pronounce the newly invented word gebs, it’s almost certain that I’d get the “z” sound rather than the “s” sound.
So there’s structure inherent at the level of sound as well as syntax, and all of this has to somehow be learned by new speakers of a language. In many cases, it’s hard to imagine exactly how a child might learn it without being taught—and with such efficiency to boot. For many scholars of the nativist persuasion, a child’s almost miraculous acquisition of language is one of the reasons to suspect that the whole learning process must be guided by some innate knowledge.
One of the leading nativists, Noam Chomsky (1986), has suggested that the problem of learning language structure is similar to a demonstration found in one of Plato’s classic dialogues. The ancient Greek philosopher Plato used “dialogues,” written as scenarios in which his teacher Socrates verbally dueled with others, as a way of expounding on various philosophical ideas. In the relevant dialogue, Socrates is arguing that knowledge can’t be taught, but only recollected, reflecting his belief that a person’s soul has existed prior to the current lifetime and arrives in the current life with all of its preexisting knowledge. To demonstrate this, Socrates asks an uneducated slave boy a series of questions that reveals the boy’s knowledge of the Pythagorean theorem, despite the fact that the boy could not possibly have been taught it.
Chomsky applied the term “Plato’s problem” to any situation in which there’s an apparent gap between experience and knowledge, and suggested that language was such a case. Children seem to know many things about language that they’ve never been taught—for instance, while a parent might utter a sentence like “What did you hit Billy for?” she’s unlikely to continue by pointing out, “Oh, by the way, notice that you can’t say Who did you hit Billy and?” Yet have you ever heard a child make this kind of a mistake?
Moreover, Chomsky argued that children have an uncanny ability to home in on exactly the right generalizations and patterns about their language, correctly chosen from among the vast array of logical possibilities. In fact, he’s argued that children arrive at the right structures even in situations where it’s extremely unlikely that they’ve even heard enough linguistic input to be able to choose from among the various possible ways to structure that input. In reverse-engineering terms, they seem to know something about the underlying language program that they couldn’t have had an opportunity to test. It’s as if they are ruling out some types of structures as impossible right from the beginning of the learning process. Therefore, they must have some prior innate knowledge of linguistic structure.
Are we wired for language structure?
If we do have some innate knowledge of linguistic structure, what does this knowledge look like? It’s obvious that, unlike vervets with their alarm calls or bees with their dances, humans aren’t born wired for specific languages, since all human infants can learn the language that’s spoken around them, regardless of their genetic heritage. Instead of being born with a preconception of a specific human language, Chomsky has argued, humans are prepackaged with knowledge of the kinds of structures that make up human languages. As it turns out, when you look at all the ways in which languages could possibly combine elements, there are some kinds of combinations that don’t ever seem to occur. Some patterns are more inherently “natural” than others. The argument is that, though different languages vary quite a bit, the shape of any given human language is constrained by certain universal principles or tendencies. So, what the child is born with is not a specific grammar that corresponds to any one particular language, but rather a universal grammar that specifies the bounds of human language in general. This universal grammar manifests itself as a predisposition to learn certain kinds of structure and not others.
If the idea that we could be genetically predisposed to learn certain kinds of language patterns more easily than others strikes you as weird, it might help to consider some analogous examples from the animal kingdom. James Gould and Peter Marler (1987) have pointed out that there’s plenty of evidence from a broad variety of species where animals show interesting learning biases. For example, rats apparently have trouble associating visual and auditory cues with foods that make them sick, even though they easily link smell-related cues with bad food. They also have no trouble learning to press a bar to get food but don’t easily learn to press a bar to avoid an electric shock; they can learn to jump to avoid a shock but can’t seem to get it through their heads to jump to get food. Pigeons also show evidence of learning biases: they easily learn to associate sounds but not color with danger, whereas the reverse is true for food, in which case they’ll ignore sound but pay attention to color. So, among other animals, there seems to be evidence that not all information is equal for all purposes and that creatures sometimes harbor useful prejudices, favoring certain kinds of information over others (for instance, for rats, which are nocturnal animals, smell is more useful as a cue about food than color, so it would be adaptive to favor scent cues over color).
Proposing a universal grammar as a general, overarching idea is one thing, but making systematic progress in listing what’s in it is quite another. There’s no general agreement among language scientists about what an innate set of biases for structure might look like. Perhaps this is not really surprising, because to make a convincing case that a specific piece of knowledge is innate, that it’s unique to humans, and furthermore, that it evolved as an adaptation for language, quite a few empirical hurdles would need to be jumped. Over the past few decades, claims about an innate universal grammar have met with resistance on several fronts.
First of all, many researchers have argued that nativists have underestimated the amount and quality of the linguistic input that kids are exposed to and, especially, that they’ve lowballed children’s ability to learn about structure on the basis of that input. As you’ll see in upcoming chapters, extremely young children are able to grasp quite a lot of information about structure by relying on very robust learning machinery. This reduces the need to propose some preexisting knowledge or learning biases.
Second, some of the knowledge that at first seemed to be very language-specific has been found to have a basis in more general perception or cognition, applying to non-linguistic information as well.
Third, some of the knowledge that was thought to be language-specific has been found to be available to other animals, not just humans. This and the more general applicability of the knowledge make it less likely that the knowledge has become hardwired specifically because it is needed for language.
Fourth, earlier claims about universal patterns have been tested against more data from a wider set of human languages, and some researchers now argue that human languages are not as similar to each other as may have been believed. In many cases, apparent universals still show up as very strong tendencies, but the existence of even one or a couple of outliers—languages that seem to be learned just fine by children who are confronted with them—raises questions about how hardwired such “universals” can be. In light of these exceptions, it becomes harder to make the case that language similarities arise from a genetically constrained universal grammar. Maybe they come instead from very strong constraints on how human cognition works—constraints that tend to mold language in particular ways, but that can be overridden.
Finally, researchers have become more and more sophisticated at explaining how certain common patterns across languages might arise from the fact that all languages are trying to solve certain communicative problems. We can come back to our much simpler analogy of the seeming universality of arrows. Arrows, presumably invented independently by a great many human groups, have certain similarities—they have a sharp point at the front end and something to stabilize the back end, they tend to be similar lengths, and so on. But these properties simply reflect the optimal solutions for the problem at hand. Language is far more complex than arrows, and it’s hard to see intuitively how the specific shape of languages might have arisen as a result of the nature of the communicative problems that they solve—namely, how to express a great many ideas in ways that don’t overly tax the human cognitive system. But an increasing amount of thinking and hypothesis testing is being done to develop ideas on this front.
2.4 The Evolution of Speech
In the previous sections, we explored two separate skills that contribute to human language: (1) the ability to use and understand intentional symbols to communicate meanings, perhaps made possible by complex social coordination skills; and (2) the ability to combine linguistic units to express a great variety of complex meanings. In this section, we consider a third attribute: a finely tuned delivery system through which the linguistic signal is transmitted.
The ability to speak: Humans versus the other primates
To many, it seems intuitively obvious that speech is central to human language. Hockett believed human language to be inherently dependent on the vocal-auditory tract, and listed this as the very first of his universal design features. And, just as humans seem to differ markedly from the great apes when it comes to symbols and structure, we also seem to be unique among primates in controlling the capacity for speech—or, more generally, for making and controlling a large variety of subtly distinct vocal noises. In an early and revealing experiment, Keith and Cathy Hayes (1951) raised a young female chimpanzee named Viki in their home, socializing her as they would a young child. Despite heroic efforts to get her to speak, Viki was eventually able to utter only four indistinct words: mama, papa, up, and cup. To understand why humans can easily make a range of speech-like sounds while great apes can’t, it makes sense to start with an overview of how these sounds are made.
Most human speech sounds are produced by pushing air out of our lungs and through the vocal folds in our larynx. The vocal folds are commonly called the “vocal cords,” but this is a misnomer. Vocal sounds are definitely not made by “plucking” cord-like tissue to make it vibrate, but by passing air through the vocal folds, which act like flaps and vibrate as the air is pushed up. (The concept is a bit like that of making vibrating noises through the mouth of a balloon when air is let out of it.) The vibrations of the vocal folds create vocal sound—you can do this even without opening your mouth, when you making a humming sound. But to make different speech sounds, you need to control the shape of your mouth, lips, and tongue as the air passes through the vocal tract. To see this, try resting a lollipop on your tongue while uttering the vowels in the words bad, bed, and bead—the lollipop stick moves progressively higher with each vowel, reflecting how high in your mouth the tongue is. In addition to tongue height, you can also change the shape of a vowel by varying how much you round your lips (for instance, try saying bead, but round your lips like you do when you make the sound “w”), or by varying whether the tongue is extended forward in the mouth or pulled back. To make the full range of consonants and vowels, you have to coordinate the shape and movement of your tongue, lips, and vocal folds with millisecond-level timing.
A comparative glance at the vocal apparatus of humans versus the great apes reveals some striking differences. Looking at Figure 2.4, you can see that the human larynx rests much lower in the vocal tract than that of chimpanzees. This creates a roomier mouth in which the tongue can move around and make acoustically distinct sounds. We also have a very broad tongue that curves downward toward the throat. Chimpanzees, whose tongues rest flat in their long and narrow oral cavity, have more trouble producing sounds like the vowels in bead or boo.
The distinct shape of our vocal tract comes at a tremendous cost: for the great apes, the height of the larynx means that they can breathe and swallow at the same time. We can’t, and so quite a few human beings die as a result of choking on their food or drink. It’s implausible that this potentially lethal trait would have evolved if it didn’t confer a benefit great enough to outweigh the risk. Some researchers have argued that speech is precisely such a benefit and that (genetically speaking) our species accepted some risk of choking as a fair trade for talking (Lieberman et al., 1969). Still, the link between speech and a lowered larynx is not clear. Many animals can and do actively lower their larynx during vocalization, possibly as a way to exaggerate how large they sound to other animals (see, e.g., Fitch, 2010).
In any case, having the right anatomy for speech is only part of the story. Somewhere in the evolutionary line between chimpanzees and us, our ancestors also had to learn to gain control over whatever articulatory equipment they had. As an analogy, if someone gives you a guitar, that doesn’t make you a guitar player (even if it’s a really terrific guitar). You still have to develop the ability to play it. And there’s reason to believe that, aside from any physical constraints they might have, non-human primates are surprisingly lacking in talent when it comes to manipulating sound. More specifically, they appear to have almost no ability to learn to make new vocal sounds—clearly a key component of being able to acquire a spoken language.
As we saw in Section 2.1, most primates come into the world with a relatively fixed and largely innate set of vocalizations. The sounds they produce are only very slightly affected by their environment. Michael Owren and his colleagues (1993) looked at what happened when two infant rhesus macaques were “switched at birth” with two Japanese macaques and each pair was raised by the other species. One revelation of this cross-fostering experiment was that the adopted animals sounded much more like their biological parents than their adoptive ones—obviously a very different situation than what happens with adoptive human infants (see Box 2.3).
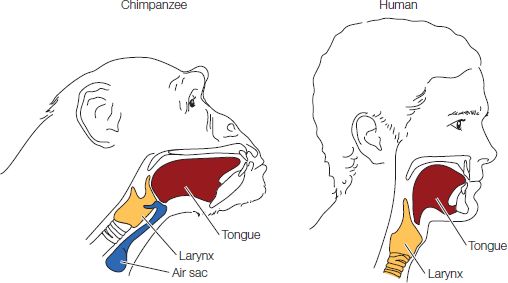
Figure 2.4 Comparison of the vocal anatomy of chimpanzees (which is similar to that of the other non-human great apes) and humans. Their lowered larynx and down-curving tongue allow humans to make a much wider variety of sounds than other primates. Humans also differ from other primates in the lack of air sacs (blue) in the throat; the precise consequences of this anatomical difference are not known. (After Fitch, 2000, Trends Cogn. Sci. 4, 258.)
The failure of primates to learn to produce a variety of vocal sounds is all the more mysterious when you consider that there are many species of birds—genetically very distant from us—who have superb vocal imitation skills. Songbirds learn to reproduce extremely complex sequences of sounds, and if not exposed to the songs of adults of their species, they never get it right as adults, showing that much of their vocal prowess is learned and not genetically determined (Catchpole & Slater, 1995). Many birds, such as ravens or mockingbirds, easily mimic sounds not naturally found among their species—for instance, the sounds of crickets or car alarms. And parrots are even able to faithfully reproduce human speech sounds—a feat that is far beyond the capabilities of the great apes—despite the fact that the vocal apparatus of parrots is quite unlike our own. This suggests that the particular vocal instrument an animal is born with is less important than the animal’s skills at willfully coaxing a large variety of sounds from it.
Sophisticated vocal learning is increasingly being found in other non-primate species. For example, seals, dolphins, and whales are all excellent vocal learners, able to imitate a variety of novel sounds, and there are even reports that they can mimic human speech (e.g., Ralls et al., 1985; Ridgway et al., 2012). Recently, researchers have found that an Asian elephant is able to imitate aspects of human speech (Stooger et al., 2012) and that bats are excellent vocal learners (Knörnschild, 2014). As evolution researcher W. Tecumseh Fitch (2000) puts it, “when it comes to accomplished vocal imitation, humans are members of a strangely disjoint group that includes birds and aquatic animals, but excludes our nearest relatives, the apes and other primates.”
Why are other primates so unequipped to produce speech sounds? Several researchers (e.g., Jürgens et al., 1982; Owren et al., 2011) have argued that not all vocalizations made by humans or other animals are routed through the same neural pathways. They’ve pointed out that both humans and other primates make vocalizations that come from an affective pathway—that is, these sounds have to do with states of arousal, emotion, and motivation. The sounds that are made via this pathway are largely inborn, don’t require learning, and aren’t especially flexible. Among humans, the noises that crying babies make would fall into this category, as would the exclamations of surprise, fear, or amusement that we all emit. Notice that, while languages have different words for the concept of a dog, laughter means the same thing the world over, and no one ever needs to learn how to cry out in pain when they accidentally pound their thumb with a hammer. Non-human primates seem to be, for the most part, limited to vocalizations that are made by the affective pathway, and the alarm calls of animals such as the vervets are most likely of this innate and inflexible affective kind.
But humans (and some other animals, including songbirds and aquatic mammals) can also make vocal sounds via a cognitive pathway. These sounds usually involve a great deal of auditory learning before they can be reliably produced, take practice, and are highly malleable. Language sounds (unlike giggles or cries of terror) are the result of the cognitive pathway and are, under this view, different from ape vocalizations not just in variety and complexity, but in their fundamental nature.
Language without speech
It would appear, then, that perhaps the sharpest discontinuity between human and non-human primates lies in the ability to produce the speech signal by which linguistic meaning is usually conveyed. But here, the evolutionary story has a very interesting twist: as it turns out, Hockett was wrong, and human languages aren’t universally spoken. It’s true that speech is the default mode along which to transmit language—in every known society whose members have normal hearing, people communicate by shaping the noises that come out of their mouths. But when humans are deprived of their usual auditory powers, they can readily adapt language and produce it by means of gesture instead of speech. Language, as it turns out, is not at all the same thing as speech and can exist independently of it. Far from being a form of pantomime, signed languages produced by deaf and hearing people alike have all the characteristics of fully fledged languages. This includes a level of “sound-like” structure where basic gestural elements are combined in various ways to form new words (displaying the property of duality of patterning), and a level of complex syntactic structure that includes recursion (see Box 2.2). In fact, as far as language researchers can tell, there are no fundamental differences between spoken and signed languages and the ways in which they’re learned, used, and understood by human beings. For the purpose of this book, just about all of the ideas that we explore about the psychology of language will apply equally well to signed and spoken languages, and I’ll normally use the term language to refer to language in either modality. And since English doesn’t have a separate word that means “to produce language by means of a signed linguistic system,” I’ll often use words such as speak or speaker without intending to exclude signed languages in any way.
So, we’ve arrived at an evolutionary paradox. It seems apparent that humans have evolved an anatomy and nervous system that outfit them perfectly for speech—and it seems quite plausible that these reflect adaptations for linguistic communication. But at the same time, humans aren’t dependent on speech to communicate linguistically. This state of affairs has led some researchers (e.g., Corballis, 1999) to hypothesize that the earliest forms of human language were gestural, and that at some later point in time, humans developed the capacity to speak. This idea fits well with several facts.
First of all, the great apes are spectacularly bad at learning to communicate with humans through any semblance of speech. But they do far better in learning to communicate through signed languages. In fact, if researchers had thought to look only at whether apes can learn spoken modes of language, we’d still be completely ignorant of the fact that they can easily learn to use hundreds of different meaningful symbols and even combine them in novel, if rudimentary, ways. So, while apes have close to zero capacity for speaking through their mouths, they can obviously control their hands and limbs well enough to make a great many distinct gestures and movements.
Second, primatologists have found that when apes communicate with each other in the wild, they do spontaneously use gestures. What’s especially intriguing is that their gestures tend to be more communicatively flexible than their vocalizations, as noted by several researchers, including Frans de Waal and Amy Pollick (2012). Vocal sounds are often limited to very specific contexts, but apes seem to be able to repurpose the same gesture in a variety of settings, much as we can flexibly use words. For example, a bonobo involved in a fight with another might stretch out its hand toward a third bonobo as a way of requesting help. But the same gesture might then be used in a very different situation, such as requesting food from a bonobo in possession of a treat. Unlike vocalizations (and again, like words in human language), gestures also seem to be more culturally specific, since chimpanzees have been found to use some gestures that are specific to their particular group.
Finally, when you trace the various communicative behaviors through the primate line, gesture makes a plausible precursor to human language compared to vocal communication. We’ve already talked about how the vocal alarm calls of vervets are quite unlike human speech vocalizations, suggesting that there’s a qualitative difference between the two. But it’s also interesting that apes, who are genetically most closely related to us, don’t make alarm calls and don’t seem to have vocalizations that have a clearly referential function. This makes apes unlike many species of monkeys and other mammals such as prairie dogs, and makes it less likely that alarm calls served as the bedrock on which language was built. At the same time, monkeys don’t appear to use manual gestures to communicate in the way that apes and humans do, hinting that gesturing is a trait that arose after apes diverged from their common ancestor with monkeys, but before humans diverged from our common ancestor with apes (see Figure 2.2).
But whether gestures served as the vehicle for the first complex, truly language-like systems is hard to know. Clearly, gesture can be used by modern humans when spoken language is not an option, which shows that language can be overlaid on more than one modality. But we simply don’t know when complex vocal skills might have emerged relative to some of the other abilities needed to support a fully fledged language. Maybe even more importantly, we don’t really have a sense of how these independent skills might have interacted and influenced each other once present in a species.
It may be that to answer these questions, we’ll have to broaden our scope and take a close look at animals that are more distant from us genetically than our favorite non-human research subjects, the primates. There’s not much evidence that the ancestors we share with other primates had speech capabilities, but there is evidence that vocal skills evolved independently in a number of quite different animals. This means we can look for clues as to why such skills might have developed, and how closely tied they might be to some of the other abilities that are needed for human language (see Box 2.4).
2.5 How Humans Invent Languages
In the previous sections, we spent a fair bit of time exploring the linguistic capabilities of animals and contrasting them with those of humans. One of the interesting findings from this body of research is that when animals interact with humans, they generally prove themselves capable of more sophisticated language skills than we’ve been able to observe in the wild (presuming, of course, that we’re looking for the right kind of “language” when we observe them in the wild). This would seem to be at least modest evidence in favor of the “language-as-tool” camp, which claims that rather than being innately wired for language, humans invented it to fill a need and, being the supremely social beings that we are, then transmitted this knowledge to subsequent generations. When animals are on the receiving end of this kind of cultural transmission, it seems that they edge somewhat closer to human-like language capability. But there’s another side to the cultural transmission story.
Communicating from scratch
It turns out that when developing a linguistic system, humans are surprisingly less dependent on cultural transmission than one might think. True enough, no child deprived of hearing German has ever grown up to speak German, fluently or otherwise. A central theme throughout this chapter is how our particular linguistic knowledge relies heavily on learning. But, as it happens, a child deprived of any real language at all does have the resources to invent from scratch at least the basic framework of a language that has just about all of the properties researchers agree are common to all human languages.
How do we know this? You might be relieved to know that language researchers don’t deliberately assign children to experimental conditions in which no one speaks to them for 10 years to see what happens. But researchers can see what happens in situations where children are born without the ability to hear the language spoken around them. More than 90% of deaf children are born to hearing parents, and since only a very small minority of hearing adults know any signed language, these children are born into a situation where they can’t make use of the particular linguistic input that the adults around them are able to provide.
Nevertheless, these youngsters are highly innovative when it comes to creating a gestural system for communication. This phenomenon is referred to as homesign. The fascinating thing about homesign is that it comes about at the initiative of the child, who has not yet had the opportunity to learn what a human language might look like, rather than being invented by the adult, who has already mastered at least one language. According to Susan Goldin-Meadow, one of the researchers who has studied gesture and homesign most extensively, the parents of these children do gesture, but usually as an accompaniment to speaking, and they generally don’t use their gestures in a way that systematically carries meaning. In contrast, children who homesign typically do make systematically meaningful gestures, as described in the following section (a more detailed description can be found in Goldin-Meadow, 2005).
When gestures replace language
In homesign, children produce separate signs as separate symbols. This is different from how people normally gesture while speaking. When people speak, they often gesture to show emphasis or to bring out some perceptual aspect of what they’re talking about. For example, while talking about a particular person, a gesture might be used to indicate that this person is large or curvaceous. But these gestures don’t have a referential value the way that the spoken names do—that is, they don’t stand in as symbols for that particular person. Children’s homesign gestures, however, do have such a referential function, and children tend to reuse the same gestures to convey the same meaning across a wide variety of situations—for example, to make requests, to ask questions, and to comment on a person or situation, whether present or not at the time.
Children also use combinations of signs to convey complex ideas—for example, rather than having a separate sign for holding an umbrella and holding a balloon, a child would invent a sign for the general notion of holding and combine that with signs for the concepts of balloon or umbrella. Combinations of this sort are used systematically in such a way that the relations between the units can be surmised from their order.
All fully fledged languages have bits of sound (or signs) that reflect inherently relational concepts—that is, words or parts of words like and, if, since; -s at the end of a word to mark plural or -ed to mark past, and so on. Homesigning children have been seen to invent signs to mark past and future, or the concept that’s captured by the word but. Goldin-Meadow has even argued that children show evidence of recursion, being able to create more complex sentential units out of simpler sentences.
In terms of the structural complexity of the homesign system, children deprived of any reliable linguistic input already exceed apes who have the benefit of a rich linguistic environment. They also exceed apes in the variety of uses to which they put their communication system. In addition to making requests or commenting on people and situations, children can use homesign to spin a narrative; to talk about hypothetical events; to make statements not just about particular things, but also about things in general (for example, to convey that dogs bark); to talk to themselves; and to comment on their homesign system. And there’s evidence that if they have to rely on their homesign system into adulthood (for example, if they never properly learn a commonly used signed language or a spoken language), they continue to innovate and add complexity over time.
The existence of homesign ought to make advocates of the language-as-tool scenario sit up and take notice. It’s clear that in addition to being born with the capacity to learn language, children are born with (at least to some degree) the capacity to invent language, and that these invented languages share some significant similarities with each other as well as with more developed languages. And, more tellingly, this inventive capacity seems to be exercised by all children who are not provided with a culturally transmitted language but who are otherwise raised in a highly social environment. Maybe it’s useful to return to the analogy of arrow-making, which Daniel Everett (2012) argues is a good way to think about language: given the usefulness of arrows, it’s not surprising that just about every human society developed some form of them, and it’s also likely that every human being of normal intelligence has the capacity to invent an arrow. But this doesn’t mean that every single person in any given society would invent an arrow if presented anew with the problem of how to catch protein that runs very fast. More likely, some would, some wouldn’t (maybe they’d explore other possible solutions), and those who didn’t would look over at the arrows made by their ingenious peers, say “What a great idea!” and promptly copy it. Yet language of some kind does seem to be truly universal even at the level of individuals, and not just at the level of groups.
Language: It takes a village
Homesign systems don’t develop into full languages if the engine behind them is a single person, suggesting that something more is needed than just one person’s inventiveness. But that something more doesn’t need to involve a more sophisticated form of language providing a model that can be copied. It turns out that when a number of people engage in the same system of homesign, it quickly ratchets up in its complexity and systematicity.
In most of North America, deaf people are either surrounded by hearing people who try to use their own spoken languages to communicate with them, or they have access to a community of speakers of American Sign Language (ASL), a fully formed language in its own right. Typically a deaf person’s experience involves both situations. But recent events in Nicaragua have provided researchers with a fascinating natural experiment in which a group of deaf people who did not know either signed or spoken language were brought together and began to communicate in a shared homesign system.
Before the 1970s, deaf people in Nicaragua had very little contact with each other, usually growing up in hearing families with little access to a broader educational system or services for the deaf. But in 1977, a school for the deaf was founded in the capital city of Managua and quickly expanded to serve hundreds of students in the region. The aim of the school was to teach the children Spanish via lip-reading, and it was not overly successful in this respect. But the really interesting linguistic action took place when children were allowed to use gesture to communicate with each other in the schoolyard or on the bus. The staff at the school initially took no part in this gestural system, but when it became apparent that this parallel system of communication was becoming the dominant means of communication for their students, they called in some experts in signed languages to provide some insight. Since then, the students’ emerging sign language has been documented by researchers, among them Judy Kegl, Annie Senghas, Marie Coppola, and Laura Polich.
What seems to have happened is this: The children arrived, each having developed, to a greater or lesser degree of complexity, some system of homesign that they’d used to communicate with their hearing families and friends. Once together, they quickly negotiated a shared, standard homesign system to use with each other. By the time the researchers showed up (Kegl et al., 1999), they noticed that, as with many homesign systems, the Nicaraguan kids showed some basic language-like patterns in their gestures. But they also noticed that as younger students arrived and were exposed to the shared homesign system, they began to alter it in ways that gave it greater communicative power. For example, in comparison to the older, original group of students, the younger signers used gestures more efficiently—they made more compact gestures that required less movement, and they combined them more fluently with each other. Whereas the older signers were found to be able to express 24 different events per minute, the younger signers almost doubled this expressive capacity, to 46 events per minute. Individual signs also came to carry more information, and the younger signers were more consistent in the ways they used signs.
Streamlining signing
Along with the efficiency gain, the researchers also documented some significant ways in which the signed system was being restructured and reorganized. As noted by Susan Goldin-Meadow and her colleagues (1996), when gestures accompany speech, they often have a pantomimed quality to them—for example, if you were pantomiming an event in which someone ate a cookie, you might imitate the action of taking a cookie out of the cookie jar and eating it. But when gestures are used instead of speech, they’re more likely to be produced as separate signs corresponding to separate concepts that are then combined together in sequence. So the same cookie-eating event might be communicated by pointing to a cookie, or by representing it as a round shape and then using a separate gestural symbol to communicate the notion of eating. In a deep way, this reflects a shift from a purely gestural system (playing a supporting role to language) toward a more linguistic one in its own right, where symbols are used for general concepts in such a way that they can be recombined with other elements. (You may have noticed this in the examples from Web Activity 2.4.)
Annie Senghas and her colleagues (2004) noticed that the younger learners of Nicaraguan Sign Language (NSL) were pushing their new language further along this trend, abandoning holistic signs in favor of more sequential ones (see Figure 2.5). To test this trend in a systematic way, the researchers showed older and younger signers some animated cartoons that depicted objects in motion (such as rolling down a hill) and had them use sign to communicate what had happened in the videos. (For comparison, they also recorded the gestures that were made by hearing Spanish speakers while orally describing the events.) In these events, it might be especially tempting to use a single holistic sign to simulate the motion. And in fact, when the videos were described by the Spanish speakers with accompanying gestures, the gestures were always holistic rather than sequential. The oldest signers were more likely than the Spanish speakers to break down the event into two component parts, produced one after the other, but they still produced holistic gestures more often than not. However, things were dramatically different with the younger signers; the two groups of students who arrived later than the original group were both found to use sequential signs most of the time.
Another interesting change was in the way in which the signers came to use spatial location for grammatical purposes. Signs within NSL are usually made in a neutral position, right in front of the body, so shifting away from that neutral position can be used to imbue the sign with some additional information. Mature sign languages make use of shifts in the spatial locations of signs for a variety of reasons—for marking past and future, for example, or even to distinguish between the grammatical subject and object of a verb. When Annie Senghas and Marie Coppola (2001) looked at how NSL signers used space, they found that older signers produced signs in non-neutral locations for several reasons—for example, to introduce new characters or topics, or to indicate different points in time. But the younger signers had settled on a very specific function of spatial signing: they used it when making the signs for verbs in order to show that the individual involved in two different actions was the same. For example, if they made the signs for see and pay in the same non-neutral location, this meant that the same person who was seen was also paid. But to the older signers, who hadn’t yet attached a specific function to spatial locations for signs, the same two verbs signed in this way could mean either that the same person was seen and paid, or that one person was seen, while another, different person was paid.
Figure 2.5 Holistic gestures (A) and sequential signs (B) exemplified by two individuals expressing the motion of rolling downhill. (A) A Spanish speaker gestures while talking. Notice how he has incorporated both the manner of motion (rolling) and the path (down) into a single gesture. (B) The person in these two photos is a third-cohort signer of Nicaraguan Sign Language; she has separated the manner and path of the motion into two separate signs, performed in sequence. (From Senghas et al., 2004, Science 305, 1779.)
Many other changes have been observed for NSL over time. Overall, these changes have had the general effect of putting more information into the conventionalized system of signs—the linguistic code—so that there is less of a burden on the viewer to rely on context or shared knowledge to fill in the gaps. Older languages that have developed over many generations rely on a great many abstract grammatical markers to mark subtle nuances of meaning. To get a feel for why all this grammatical marking might have emerged, let’s imagine a version of English that has a much sparser linguistic code than our current version, in terms of its structure and markers—think of a simplified form of English spoken by a small child, or by someone just beginning to learn English. For each of the simplified versions that follow (the a sentences), think about the additional contextual knowledge you’d need to have in order to understand it as equivalent to the b version of the sentence, but distinct from the c version:
(a) Christopher Columbus sail America.
(b) Christopher Columbus sailed to America.
(c) Christopher Columbus will sail from America.
(a) Francis build house last year.
(b) Francis was building a house all of last year.
(c) Francis built a house last year.
(a) Timmy marry girl. Girl have baby.
(b) Timmy is going to marry a girl who’s had a baby.
(c) The girl that Timmy married is going to have a baby.
It’s not hard to see how elaborating the linguistic code with the grammatical embellishments found in the more complex sentences might help improve the efficiency of communication and reduce the possibility of misunderstandings.
The sensitive period and innate language ability
Senghas and other NSL researchers have suggested that two ingredients were needed in order for this shared homesign to progress beyond its humble beginnings: (1) a community of speakers using the same linguistic system; and (2) a generational progression in which very young speakers were exposed to the structured input of their linguistic predecessors. The greatest competency in the language, and the most sophisticated innovations, were observed in new learners who had the benefit of the structured system that had been put in place by a previous cohort of students. In particular, students who were very young when exposed to this structured input benefited the most. This fits with the notion that there’s a sensitive period, a window of time during which children seem to have a special aptitude for learning language. In general, young children who are exposed to a foreign language learn it quickly and in a native-like manner, whereas most adults who start learning a foreign language never achieve native fluency, even after decades of exposure. It’s easy to see this in immigrant families in which the kids rapidly shoot past their parents’ or grandparents’ abilities to speak in their newly adopted tongue—much to the annoyance and envy of their elders.
Many nativists have argued that evidence of a sensitive period for language learning supports the view that language is innately specified. If, as the non-nativists claim, language is simply a by-product of our vigorous intellectual capacity, then it’s hard to explain why it should be that small children—who overall are not as smart as adolescents or adults—seem to have a leg up on their intellectual superiors when it comes to language learning. On the other hand, it’s not hard to explain under a nativist view, as there are other cases in nature where a genetically specified ability never develops properly if it hasn’t been activated within a certain window of time. For instance, if songbirds are kept from hearing the songs made by adults of their species during a certain period of their development, they never manage to sing normally afterward (e.g., see Brainard & Doupe, 2002).
The parallel to songbirds is especially intriguing, given that there’s a surprising amount of overlap between human language and complex birdsong, as discussed in Box 2.4. Birdsong, like human language, seems to involve a mixture of genetic programming and learning from experience, though it looks like the precise balance between genes and experience varies somewhat among bird species. But the songbird story gets especially interesting when we consider that researchers have found among our avian friends some parallels to the emergence of NSL. In a study by Olga Feher and her colleagues (2009), young male zebra finches were kept from contact with other birds (researchers use the term isolate to refer to a young bird in this situation). Isolates that aren’t exposed to adult song never grow to sing normally, but parallel to human homesigners, they do produce simpler, distorted songs with some of the elements present in the normal adult songs. It turns out that new baby birds will readily imitate the songs of isolates even though they sound abnormal. The researchers wondered: If baby birds only hear songs of isolates and are kept from hearing normal adult song, what do they grow up to sound like? Do they sound just like the first-generation isolates, or do they, like later cohorts of NSL signers, add on to the song system and change it? Their experiments found that the second-generation isolates, exposed only to the songs of the first generation, produced sounds that came closer to the normal songs of zebra finches than the first generation did. The researchers then had progressive generations of zebra finches learn from their predecessors and found that over the course of three or four generations, the songs of the zebra finches raised in this way came to sound very much like what the birds produce in a normal environment. It seems that birdsong, like human language, can reconstitute itself from nothingness into something approaching its full glory in a small number of generations.
So where does language come from?
It’s tempting to see the emergence of homesign and NSL as evidence that children are innately wired for language. Certainly, something is driving the creation of a linguistic system that has many of the features of fully developed languages. But we still don’t know exactly what that something is. We also don’t know exactly why it is that, in order to create a “full” language with all the complexities of more developed languages, kids need exposure to language from other people, even if it’s not exposure to a fully formed variety of language. Nor do we know which of the features commonly found across languages are universal because they spring from our genetic code, and which of these features arise simply because they represent the optimal solution for how to package and transmit the kinds of information that human beings are inclined to share with each other.
What is clear is that language is the result of an intricate collaboration between biology and culture. It’s extremely unlikely that all of the features of language are genetically determined, or conversely, that all of them are cultural inventions made possible by our big brains. Full answers to these questions will lean heavily on research within the growing field of biolinguistics, which is preoccupied with tracing the connections between genes and human linguistic behavior. As you will see from the next section, this is no easy feat, but recent advances in genetics research hint at the prospect of brisk progress and exciting revelations waiting in the wings.
2.6 Language and Genes
Ever since Down syndrome was definitively linked to a chromosomal anomaly in 1950, we’ve known that the ability to learn and carry out intellectual tasks can be disrupted by genetic “glitches.” Since then, scientists have identified a variety of learning disabilities that have a strong genetic basis and that show distinctive patterns of impairment.
These disorders are an important piece to the puzzle of language evolution. For example, they may be able to hint at whether there’s a separate, dedicated language system that has evolved in humans over time, or whether language has emerged as a by-product of humans’ generally muscular intelligence. These two perspectives fuel very different expectations about the link between language functioning and other aspects of intelligence. If language is an outgrowth of overall intellectual ability, then genetic anomalies that curtail other intellectual functions should have a dramatic effect on language. On the other hand, if language is a specially evolved “module” (much like a specialized organ), it might not be that tightly connected to other cognitive skills.
Williams syndrome: An island of preserved function?
Williams syndrome (WMS) has attracted the attention of language researchers because it appears to be a case where language function is fairly well preserved despite some striking impairments in other domains. WMS is caused by a specific genetic anomaly on chromosome 7. Together with certain facial features and cardiovascular problems, it usually results in learning disability, with the overall IQs of affected individuals typically falling in the 50–70 range. People with WMS tend to be socially gregarious and, as it turns out, are often very verbal. Several language researchers have been struck by this last trait, especially given the tremendous difficulties that people with WMS often show on a variety of cognitive tasks, including those that rely on numerical or visual–spatial skills.
Ursula Bellugi and her colleagues (2000) have documented the linguistic and non-linguistic skills of people with WMS compared with the cognitive profiles of people with Down syndrome, another genetic anomaly that leads to intellectual impairments. When Bellugi compared a group of adolescents with WMS and a group with Down syndrome, she found that the overall scores on tests for IQ and cognitive functioning were similar for the two groups. In particular, the WMS group showed quite dramatic difficulties with numerical concepts—for example, many of them said they would rather have “50 pennies” than “5 dollars,” and when asked to estimate the length of a bus, they gave responses such as “3 inches or 100 inches maybe” and “2 inches, 10 feet.” Needless to say, they had a great deal of trouble carrying out tasks like making change, balancing a checkbook, or cooking from a recipe. Some of their most dramatic difficulties were in spatially organizing parts of objects into coherent wholes.
Typically, individuals with WMS operate at about the level of an average 6-year-old when it comes to their conceptual understanding, but their conceptual weaknesses are often accompanied by very adult-sounding language. For instance, one young woman, who was literate and enjoyed reading about vampires, seemed to have trouble understanding the concept of vampires, a confusion that stood in stark contrast to her use of very advanced vocabulary; when asked to define a vampire, she offered that a it is “a man who climbs into ladies’ bedrooms at night and sinks his teeth into their necks.” When asked why vampires behave in this way, she said “vampires must have an inordinate fondness for necks” (Johnson & Carey, 1998).
In Bellugi’s comparison of Williams and Down syndromes, language was clearly more sophisticated among the Williams group. Their sentences were more fluent and complex, and they showed a stronger understanding of how syntactic structure contributes to meaning. For example, in a sentence like The man is chased by the horse, you need a good grasp of syntax to know who is doing the chasing and who is being chased—you can’t simply deduce this from the words man, chased, and horse. Individuals with Down syndrome performed almost randomly with such sentences when matching them up with pictures of the events they depicted, while the Williams group showed much better performance. Examples of the divergent strengths and weaknesses of the two groups are shown in Box 2.5.
Some language scientists have taken results such as Bellugi’s to be strong evidence for a genetically specified language module that is independent of overall intelligence. But a good amount of subsequent research has challenged this conclusion.
First of all, comparing individuals with WMS with a group of people with Down syndrome doesn’t necessarily provide the best assessment of their linguistic strengths, even if both groups are matched for overall IQ. A slightly different picture emerges when people with WMS are compared with typically developing kids of the same mental age—that is, a group of children who are at the same overall level of cognitive functioning. The logic is that if a person with WMS is at the cognitive level of a 6-year-old, then it makes sense to compare his language abilities with those of a typical 6-year-old. If these abilities are at about the same level, this suggests that the linguistic abilities of the person with WMS are closely linked to his other cognitive abilities. In order to conclude that the language module is preserved in WMS, we’d need to see evidence that language abilities actually exceed what we’d expect to find based on mental age alone (and perhaps are closer to that person’s chronological age).
In fact, on most detailed measures of language, subjects with WMS perform about as well as you’d expect based on their mental age (for a review, see Brock, 2007). The truth is, a typical 6-year-old has pretty good language skills too. The striking—but somewhat misleading—impression that a number of researchers had of the unexpectedly strong linguistic performance of the WMS individuals exemplified in Box 2.5 probably came about for several reasons: (1) when they are compared with subjects with Down syndrome, their language is good, but this is largely because Down subjects underperform on language relative to their mental age (a fact which, in and of itself, demands an explanation and suggests that language and other cognitive abilities aren’t always in sync); (2) the language abilities of WMS individuals are surprisingly good when compared with their difficulties with visual–spatial and numerical tasks—but this is because performance on these latter tasks is much worse than you’d expect based on their mental age; and (3) certain superficial features of WMS language (such as the use of rare words or unusual turns of phrases) give the impression of greater linguistic sophistication, but these words and phrases may be used without full control or understanding.
So, a closer look at the cognitive and linguistic profiles of people with WMS doesn’t really show a dramatic dissociation between language and overall cognitive ability. At the same time, results from a wide variety of language measures do suggest a couple of areas of strength for WMS subjects. Relative to their mental age, they score especially well on tests of receptive vocabulary (words that are recognized and understood) and the ability to hold auditory material in short-term memory. Relative weaknesses within the language domain also exist, however; for instance, WMS subjects have been found to score poorly on comprehension of complex syntactic structures (e.g., Mervis, 1999).
The research is far from complete, but the emerging message is this: WMS doesn’t appear to supply definitive evidence for an independent language module, but it does point to an intriguing separation of some specific cognitive skills, both linguistic and non-linguistic in nature. The hope is that by systematically studying many of the skills that are involved in learning and using language, researchers will ultimately come to a better understanding of the particular skills that have a genetic basis, and the consequences for language when these skills are disrupted.
Specific language disorder: An island of impaired function?
Having asked whether Williams syndrome is truly a disorder in which language is preserved while other cognitive functions are deeply affected, we will now look at the flip side of the same question: Is there a disorder in which language is selectively disrupted while other cognitive functions are intact?
Research around this question has focused on specific language impairment (SLI), which has been defined as a disorder in which children fail to develop language normally even though there’s no obvious reason for this—that is, no apparent neurological damage or disorders, no general cognitive impairment or delay, no hearing loss, and no abnormal home environment. Children with SLI usually start speaking later than their peers, and once they do talk, their language shows evidence of odd glitches, some of which linger into adulthood. They produce strangely ungrammatical sentences (for example, Who did Marge see someone? and Yesterday I fall over), and they persist in such errors well past the age when children normally make elementary syntax errors. Abnormalities at all levels of language structure have been found—that is, at the levels of sound structure, sentence structure, and the structure of complex words.
There’s quite strong evidence that the disorder has a hereditary component, as gleaned from family histories, from studies of identical and non-identical twins, and from the fact that a number of genetic anomalies have been found in people with SLI. To some researchers, SLI appears to offer evidence that certain aspects of language structure behave as modules that are independent of other cognitive functions and that have a direct basis in genetics. This is the point of view taken by researchers such as Heather van der Lely and Ken Wexler (e.g., van der Lely & Marshall, 2011; Rice & Wexler, 1996). These researchers disagree with each other on the details, but their general approach is to say that SLI is due to a genetically based disruption in the process of learning language structure. The end result is a “broken” grammar, or being stuck at a stage of arrested development in the learning of complex language structure. In short, they take a domain-specific perspective on SLI, in which the linguistic deficit strikes at mechanisms that are particular to language, rather than ones that are shared with other cognitive abilities.
But, as you might have guessed from our discussion of Williams syndrome, the picture is less clear close up than it appears from a distance. For starters, the dissociation between language and other cognitive functions in SLI is far from sharp. It’s certainly true that the problems that usually bring children with SLI into clinicians’ offices are their difficulties with language, rather than any other obvious signs of cognitive delay or impairment; in other aspects of their lives, these kids seem to be functioning fine. But more detailed testing shows that many children with SLI also show unusual performance on other tasks that are at best indirectly related to language structure (see Table 2.2). Basic speech perception is often impaired, with SLI kids having more trouble distinguishing between similar sounds like “ba” and “pa.” They might also have shorter memory spans, as measured by their ability to retain words or other units in memory over a short period of time. Some children also have trouble with control over their articulatory systems, or even with more general aspects of motor coordination. (See Joanisse & Seidenberg, 1998, for a review of non-linguistic deficits that can accompany the linguistic problems in SLI.)
What to make of these more general symptoms? Several different explanations are possible. Some researchers have argued that the non-linguistic impairments are a clue that the underlying problem isn’t specifically linguistic after all. Instead, they argue for a domain-general perspective that views SLI as a cognitive problem that’s not particular to language in and of itself but that ends up having especially weighty consequences for language just because language happens to draw heavily on precisely the skills that are impaired. In response to such arguments, defenders of the “broken grammar” view have countered that the mere presence of non-linguistic deficits can’t be construed as evidence that they cause the apparent problems with language. For instance, Heather van der Lely and Chloe Marshall (2011) take the position that these other cognitive deficits may coexist with the linguistic impairments, but that they don’t really have anything to do with the language deficit itself. After all, even in genetic disorders like Williams or Down syndrome, which can be traced back to a single chromosome, the cognitive impairments are also accompanied by other symptoms or features—for instance, certain typical facial characteristics, or congenital heart problems. Does this mean that the heart is part of the same “system” as the impaired cognitive functions? Hardly. Rather, the co-occurrence reveals that genes can sometimes have very diffuse effects, with ramifications for multiple systems at the same time. To show that the linguistic problems stem from the more general processing deficit, it’s important to show that the severity of the linguistic impairment is clearly related to the severity of the more general processing deficit.
There’s still a lot we don’t know about the link between the language problems and more general cognitive anomalies of people with SLI. A number of issues complicate the picture and will need to be carefully sorted out. For example, it’s unlikely that SLI makes up a single disorder with a single underlying cause. There’s quite a bit of variability in the linguistic and non-linguistic profiles of people who have been diagnosed as having SLI. Moreover, unlike Williams syndrome, the problem can’t be diagnosed through genetic testing, because no single genetic anomaly has been identified as being at the root of SLI. Many genetic variants have been shown to increase the risk of language impairment, with a number of these typically showing up in individuals described as having SLI, and people who seem to have completely typical language development may have some of these variants as well. All this has led researchers to suggest that SLI is a catchphrase for a cluster of cognitive vulnerabilities that end up disproportionately affecting language function, with each individual genetic variant having only a very small effect. This lack of specificity of otherwise unexplained language disorders is reflected in the fact that the current version of the Diagnostic and Statistical Manual (DSM-5; American Psychiatric Association, 2013), which is the primary handbook used by clinicians to diagnose psychiatric and cognitive disorders, has changed the term Specific Language Disorder to the more generic Language Disorder.
Adapted from Joanisse and Seidenberg, 1998, Trends Cogn. Sci. 2, 240.
It’s still possible that one or more of the genetic variants leading to problems with language learning may turn out to be specific to language, with little or no impact on any non-linguistic function. But overall, the research on SLI dispels the notion that there is an easy genetic link to be made between language functioning and other cognitive processes.
The new frontier: Tracing our ancestors’ genes
One of the great challenges facing the study of language evolution is that language itself leaves no fossils (given that language surely arose long before writing systems did, the earliest record of which is a mere 3,000 years old). As scientists, we’re stuck trying to make inferences about whether our ancestors did or did not speak based on whatever physical evidence they’ve left behind. Until very recently, the only physical evidence we had to work with was the shape of the bodily remains of our ancestors or now-extinct relatives, along with artifacts or drawings that hinted at complex culture or tool-making. Recent advances in genetic sequencing have changed all that. Incredibly, scientists have been able to sequence the genomes of two extinct hominin species, Neanderthals and Denisovans, with whom we share common ancestors after our lineage split off from the chimpanzee lineage. This new evidence, along with the genetic maps of other living primates and animals, throw open new doors for research. As scientists start to identify specific genes that are linked to language and other cognitive functions, they will be able to compare the human genome to those of extinct and living relatives, or even to very distant animals that have certain abilities that resemble ours.
An intriguing case study can be found in the flurry of research around the FOXP2 gene, located on chromosome 7. This gene captured the attention of language researchers when it was discovered by a team of geneticists (Fisher et al., 1998; Lai et al., 2001) to be the site of a specific mutation observed in a certain family (identified as the KE family) living in the United Kingdom. As previously documented by Myrna Gopnik and Martha Crago (1991), half of this family (16 members across 3 generations) had severe language learning problems as children, with virtually unintelligible speech and grammatical difficulties. There was a clear genetic cause to this language impairment: any individual who inherited the mutation from at least one parent displayed the language impairment, whereas family members without the mutation showed typical language development. It quickly became apparent that this mutation also led to other non-linguistic problems, such as trouble forming non-linguistic sounds (humming a tune, clicking the tongue) and producing facial movements on request (closing the left eye, biting the lower lip) and slightly lower overall IQs (Vargha-Kadem, 1994). Nevertheless, the gene’s clear association with language functioning made it a target for researchers interested in the evolution of language (see Fisher et al., 2015, for a review of relevant work).
FOXP2 is a gene that is found not only in our close primate relatives, but in many other species of vertebrate animals as well. This is a gene that regulates the activity of many other genes, and as a result, it plays an important role in brain development (in addition to the development of other organs). The gene has been very stable throughout much of its evolutionary history. However, the human version of FOXP2 reveals two genetically important differences compared with the chimpanzee version, and researchers speculate that these changes may play an important role in language. (The songbird version, for its part, has been implicated in the normal development of birdsong, as evident from studies that disrupt the gene’s functioning, resulting in dire consequences for the birdsong of the affected animals.)
An intriguing recent development is that our species is not the only one to bear the “human” version of FOXP2. It has also been identified in our extinct Neanderthal relatives (Krause et al., 2007). Does this mean that we are not the only articulate hominin species to have walked the planet? To answer this question, we will need to learn more about the exact role of FOXP2—and other genes—in how the brain develops, and what this means for learning and using the kind of information that is characteristic of language.
FOXP2 was the first gene to have been decisively identified as important for language. But language ability hinges on other genes as well, as evident from the fact that the vast majority of cases of language disorders can’t be linked to FOXP2. Still, the gene’s discovery has opened up new lines of research and spurred collaborative relationships and cross-fertilization of ideas across disciplines that previously didn’t have much to say to each other. These new developments and collaborations will enhance progress in understanding the roles of other language-relevant genes as they are discovered.
2.7 Survival of the Fittest Language?
So far, we’ve talked about language evolution and adaptation entirely from a biological perspective. That is, a trait is adaptive to the extent that it helps an organism survive and procreate. Organisms that survive and procreate pass on their genes, while those that die young or can’t attract a mate fail to leave a genetic legacy. As a result of this cold, hard reality, the genes for an adaptive trait spread throughout a population or species. In this scenario, what changes over time is the genetic makeup of the individuals that make up the species. In language nativist terms, the capacity for language is adaptive, and those humans who readily learned and used language had a reproductive advantage over those who didn’t, so over many generations, the general human population came to possess a highly efficient set of cognitive skills tuned for the learning and use of language. By this account, the human brain as shaped by our genes has changed over time and has become specifically good at language. But it’s also possible to talk about the evolution of language from a completely different angle: languages themselves are in a constant state of change, and not just when they’re being newly formed, as with Nicaraguan Sign Language.
Language changes
The English in which Chaucer wrote the Canterbury Tales is incomprehensible to most English-speaking people today. Americans have come to speak a variety of English that is different from the one people in the United Kingdom speak—and neither of those varieties is the same as the English that came out of Shakespeare’s mouth 500 years ago (see Box 2.6). In fact, your version of English isn’t even exactly the same as your grandparents’ version (even assuming your grandparents were native English speakers). It’s common for older generations to complain about the “poor grammar” of younger generations, and perhaps you’ve had someone older than you chastise you for saying “I’m good” instead of “I’m well,” or inform you that it’s wrong to say “less than three cookies”—it should be “fewer than three cookies”—or that “Who did you call?” should be “Whom did you call?” If so, maybe you should inform them that these aren’t grammatical errors—they’re grammatical mutations.
Languages, like genes within populations, are in constant flux. English has seen some significant changes over the years, including how words are pronounced, what words mean, and which grammatical markers or structures have been adopted. As one minor example, back in the 1600s, over the loud protests of stodgier generations, speakers of the English language stopped differentiating between singular and plural pronouns for the second person: you (plural) and thou (singular) became collapsed into the pronoun you, used for either the plural or singular. The phenomenon of language change has long been studied with great interest by linguists. In fact, at the time Charles Darwin was still mulling over his ideas about biological evolution, linguists were already well aware of language’s tendency to mutate and gradually shape-shift—often diverging over time into separate languages, just as genetic changes within groups of individuals can eventually result in distinct species (see Figure 2.6).
The conceptual tools from biology that are used to explain why the genetic makeup of a species changes over time turn out to be quite useful when applied to thinking about why languages change over time. (And I do encourage you to pull out these concepts when you find yourself needing to defend yourself against charges that you are contributing to the corruption of English.) In biological species, mutations occur randomly within individuals, but the vast majority of these don’t spread widely or lead to significant changes within the species. The same is true for language: you might have a distinctive way of pronouncing the sound “s” or you and your friends may have coined a new way to express that you like something (following in the long tradition of expressions like “groovy,” “nifty,” “cool,” “bomber,” “phat,” or “sick”), but most of these little innovations won’t catch on to the point of becoming embedded in what people around the world know as part of the English language.
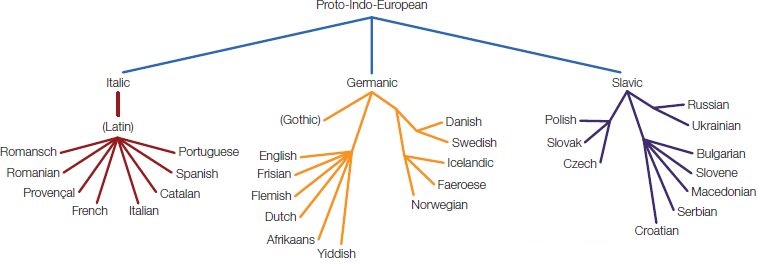
Figure 2.6 A dendrogram (tree) presenting the relationships among some of the modern European languages, showing how they diverged from a common ancestral language through a series of linguistic changes over time. Languages in parentheses are no longer spoken.
When mutations do take hold, whether in genes or language, they do so for one of two reasons, the first boring and the second very interesting. The “boring” reason is known, biologically speaking, as genetic drift: mutations spread randomly when some individuals in the population happen to reproduce more than others not because of any genetic advantages they possess, but simply because they got lucky (who knows, maybe their mate was sick at home during the freak flood that happened to wash away a number of other males who were out hunting that day). The far more interesting reason—and the one researchers suspect of nudging our species toward language—is directional selection: adaptive mutations that enhance an individual’s prospects of passing their genes on to lots of offspring spread throughout a population because they outcompete genotypes that don’t offer this advantage. Both of these forces help explain genetic diversity and how it is that a single species can split off into distinct species over time.
Similar explanations can be given for linguistic diversity, helping us understand why, worldwide, humans currently speak approximately 6,000 languages rather than just one. Just as with genetic drift within a species, some linguistic innovations just happen to be repeated more often than others, for no particular reason, and thus gain a long-term foothold in the language. But historical linguists, who study the detailed ways in which languages change over time, have noticed that language change is not fully random, and that across languages, certain patterns of change are far more likely than others to spread from one generation to the next. This raises the possibility that some innovations are more adaptive than others, when viewed from the perspective of the language’s ability to propagate itself. What characteristics give language a better chance of surviving and being passed on to the next generation? It seems logical that at least two criteria would need to be met:
1. The language needs to be communicatively useful. This means that the language should be able to convey the information that people want to get across in an efficient way. Efficiency probably needs to be seen from the perspectives of both the speaker and the hearer. Ideally, the delivery of information should take as little time as possible to utter (and to make sense of), and it also shouldn’t put too much strain on the processing resources of either the speaker or the hearer. Forms that take too long to utter, or that mentally tax the speaker, or that are subject to misunderstandings by the hearer will simply be used less often than optimal forms, and over time people will converge on the best communicative solutions.
2. New learners need to be able to learn the language. The features of a language that are difficult to learn will simply not be learned reliably, and as a result won’t be passed down to a new generation of learners. This means that new learners may play an important filtering role in shaping language. In fact, several researchers (e.g., Mufwene, 2008) have proposed that this filter is the driving force behind linguistic changes that take place when a language is passed from one generation to another—maybe it’s not so much that children innovate changes more than previous users of the language but that they play a key role in selecting which of a number of inconsistently produced innovations will survive and become more systematically used by speakers of the language.
What’s adapting to what?
The perspective of linguistic evolution offers a dramatic reversal of the traditional nativist stance on language, as noted by Morten Christiansen and Nick Chater (2008). Nativists who argue that children are innately outfitted with a universal grammar take the view that the human mind has changed over time so as to adapt to the forms and structures that occur in human languages. But the alternative perspective I’ve just sketched out—let’s call it the cultural transmission view of language change—says that it’s languages that have changed over time so as to adapt to the human mind, with all the constraints, limitations, and abilities that human minds bring to the task of learning or using language. Each of these two accounts offers an explanation for why it is that human languages share a number of similarities with each other, but the explanations look strikingly different. Under the nativist view, commonalities across languages reflect hardwired biases to learn certain forms of language. These biases allow children to quickly zoom in on the correct structures of their languages rather than floundering around considering all the logical possibilities. Under the cultural transmission view, languages are similar to each other because they’ve all had to adapt to the human brain, and presumably, the capabilities and limitations of the human brain are similar across many cultures.
The cultural transmission view has some fascinating implications to explore. For example, if the cultural analogues of “natural selection” include both a language’s usefulness and its learnability, then perhaps a language might be coaxed into different shapes depending on the makeup of the population that uses and learns it. A close parallel from biology would be the different selectional pressures on species depending on their particular habitats. That is, just as a species occupies a specific ecological niche, a language occupies a linguistic niche that is particular to that language. It might be very interesting, for example, to compare the development of NSL with another emerging sign language that has recently caught the attention of language scientists, Al-Sayyid Bedouin Sign Language, or ABSL (see Figure 2.7). First described by Wendy Sandler and colleagues in 2005, ABSL has emerged in a small Bedouin community in Israel. This community is unusual in that a great many of its members have an inherited form of deafness because of the high rates of intermarriage within the small population. Far from posing a problem for the community, this has led to a new signed language, created by the people and used by deaf and hearing individuals alike. A deaf child grows up fully integrated within the society, and usually marries a hearing person, which means that families continue to be a mixture of hearing and non-hearing individuals, which in turn motivates hearing members of the family to learn ABSL. This emerging language is now three generations old. Its earliest adopters are now dead, so it’s not possible to know exactly how this language first came to be, and whether it was at the initiative of deaf or hearing people or the result of a joint effort. What is known is that even the basic word order is different from any of the spoken languages in the region, so from the beginning ABSL seems to have sprung up with its own structures rather than simply importing structures from the nearest spoken language at hand.

Figure 2.7 Two speakers of Al-Sayyid Bedouin Sign Language, spoken by about 150 deaf and hearing speakers of the Al-Sayyid Bedouin community in the Negev desert in southern Israel. (From Senghas, 2005, Curr. Biol. 15, R463; photographs by Shai Davidi, University of Haifa.)
There are a few striking differences between the groups of people who collectively created NSL and ABSL and, as pointed out by Ann Senghas (2005), these differences might turn out to be relevant for how the languages take shape. First of all, the speakers of ABSL all come from a single, very tight-knit community, whereas the speakers of NSL were bused in from across a large region, coming from many families and villages that have little connection to one another. Why might this matter? Remember that many of the innovations that were brought into NSL involved making the linguistic code more precise and unambiguous. This reduced people’s need to rely on shared contextual knowledge in order to recover the subtleties of the intended meaning. The earliest speakers of NSL, gathered together from diverse backgrounds, would likely have had much less shared knowledge than the earliest speakers of ABSL. Because of this, the communicative pressures within the two communities might have been quite different, with possibly more pressure on the NSL community to pack a lot of information into its linguistic code very early on.
A second important difference between the two communities speaks to the issue of learnability. New learners of ABSL are exposed to the language from birth within their own families. But deaf children in Nicaragua typically don’t start to learn NSL until they’re old enough to be sent to school. This means that the cognitive makeup of the new learners is quite different between the two groups in a way that may be very relevant to language development—if learners serve as “filters,” influencing the shape of the languages they are learning, this might result in different kinds of information being filtered out of the language system by the two groups.
There’s a good deal of research that shows that older learners, particularly adults, don’t learn language in the same way (or as successfully) as younger learners. (You’ll get a closer view of some specific differences in later chapters.) You don’t have to look to signed languages to find evidence of languages that have a heavy proportion of older learners. This can easily happen when a large number of the speakers of a language learn it as a second language, typically in high school or adulthood. This happens to be true of English speakers worldwide, 64% of whom are second-language learners, and of speakers of Swahili, a whopping 90% of whom learn it as a second language (Lupyan & Dale, 2016). In contrast, very few people learn Czech as a non-native language—for the most part, you’re either born into the language or don’t know it at all, and in my experience, native Czech speakers react with a mixture of horror and amusement upon hearing their language coming out of the mouth of a non-native speaker. Such differences might affect how languages evolve over time. For example, some researchers have found that languages with a high rate of second-language learners tend to rely on word order to signal the difference between subjects, direct and indirect objects, and so on, rather than using word endings called case markers (Bentz & Winter, 2013), and generally load less information into single words compared with languages with lower rates of second-language learners (Bentz et al., 2015).
Theories of cultural transmission provide an alternative to the notion of universal grammar when it comes to thinking about how language and the human mind fit together. But the two general approaches need not be incompatible. It could well turn out that we have a core set of innate predispositions that come from our being genetically adapted for language. But it may also be true that not all of the universal properties of languages have come out of these predispositions—some of them may have arisen as adaptations of languages to us.
What we still don’t know
While language science isn’t yet in a state where we can confidently choose among any of the competing views of language evolution that we’ve described in this chapter, it is in a state where it makes sense to spend a lot of time discussing them. The various theories can now serve as an engine to drive the more detailed questions that researchers need to resolve before being able to answer the big questions of where language came from and why we have it. Much of the detailed groundwork will have to do with questions like these:
■ What do our language-related abilities look like, and how specialized are they—that is, how similar or different are language abilities from the cognitive abilities we use for purposes other than language?
■ What are the structural properties of language, and what are the optional versus obligatory aspects of language?
■ How are the various components of language learned, and why do children seem to learn them better than adults?
■ How do we produce and understand language, and under what conditions do we do these things smoothly or bumpily?
■ How do speakers and hearers negotiate how much information needs to be put into the linguistic code, and how much can be left to be figured out from the context?
The upcoming chapters will deal with these questions. Here’s an interesting exercise: After you’ve worked your way through this book, come back and reread this chapter. You’ll likely find that with the knowledge you’ve gained, the big themes that have been sketched out in this chapter will feel more “alive” for you. You may find yourself developing strong opinions about competing ideas about language evolution, spinning off new questions, generating new hypotheses, and even thinking of ingenious ways to test some of those new ideas.
GO TO