
The cat family
With a global population that may exceed a billion, there are perhaps 300,000 housecats for every Tiger left on earth. The most generous population estimates for all wild cat species combined might reach one per cent of housecat numbers; 10 million wild cats, most of them small-bodied, wide-ranging generalists such as Bobcats and Leopard Cats.
The domestic cat is one of the most successful mammals on earth and its most successful carnivore1. Resident cat populations occur on every major continent except Antarctica and on most of the world’s offshore islands. Cats can survive in virtually any habitat from the Sahara Desert to sub-Antarctic islands, whether they are cared for by people or not. At least half a billion cats are kept as pets around the world, and there are many hundreds of millions more that live as strays loosely associated with humans or completely feral with no reliance on people at all.
The cat’s success embodies the evolutionary triumph of the Family Felidae. Felids have walked the earth for around 30 million years and prior to very recent anthropogenic impacts, have been extremely successful. Felid evolution began in Eurasia where the Family’s first unambiguous representative – sufficiently different from earlier fossil carnivores to be considered a true cat – is Proailurus lemanensis. The oldest Proailurus fossils are 25–30 million years old from what was then a vast subtropical forested landscape and is now Saint-Gérand-le-Puy in France. Proailurus lemanensis is the likely progenitor of all cat species, living and extinct, that have ever lived. By approximately 18–20 million years ago, Proailurus had diverged into two distinct genera that seeded the two main branches of cat evolution. One of these, the genus Pseudaelurus included cats which for the first time in felid evolution had reached the size of the modern Leopard. Their skulls and teeth also carried incipient sabretooth features such that Pseudaelurus is now considered ancestral to the felid subfamily Machairodontinae, the sabretooths. This spectacular experiment in felid evolution produced many dozens of species with famously elongated canine teeth and a raft of other modifications in the skull and skeleton that differentiates them from other felids. The sabretooth cats prospered in Eurasia, Africa and the Americas until very recently. The best-known genus Smilodon lived until 10,000 years ago in North and South America, and included some of the most extraordinary, largest felids to have ever evolved. Smilodon fatalis – the celebrated Californian sabretooth known from over 1,200 specimens in the Rancho la Brea tarpits – was as tall as the modern Tiger but was more heavily built and weighed more, while the massive South American Smilodon populator far out-weighed any living cat at close to 400 kilograms. Both species lived alongside humans.
SABERTOOTH FAMILY TREE
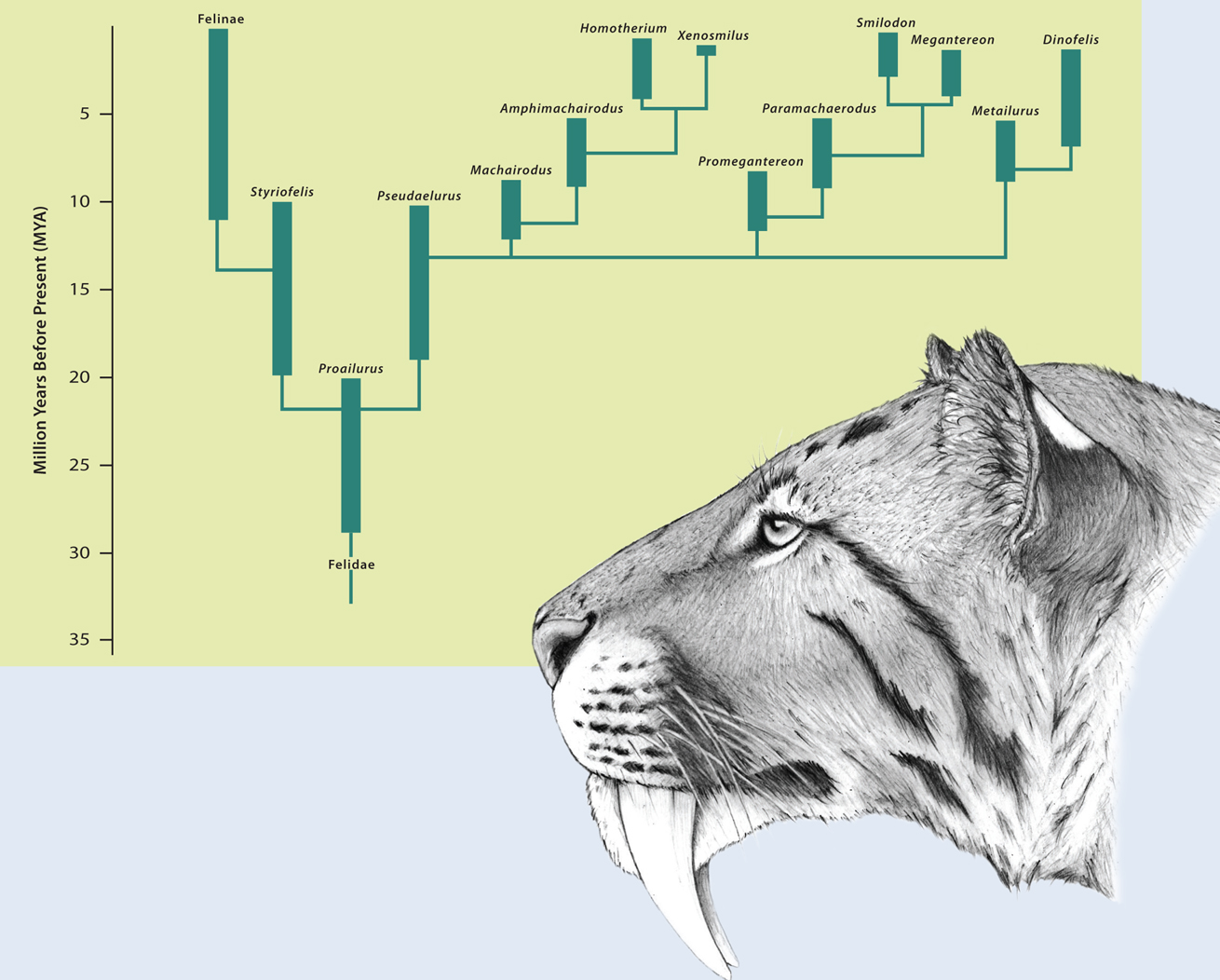
The Machairodontinae and Felinae are two separate branches on the felid family tree that diverged early in cat evolution. All are true cats (Family Felidae) but all living felids are more closely related to each other than to the sabertooth cats. The misnomer ‘sabertoothed tiger’ (typically used for Smilodon, pictured) is particularly erroneous; the Tiger is more closely related to the housecat than it is to any sabertooth. (Chart redrawn from Anton, M. 2013 Sabertooth Indiana University Press.)
Conversions
Throughout this book measurements, weights and areas have been provided using the metric system, however, those more used to the Imperial system may find the table here useful.
In parallel to the great proliferation of sabretooths, the second major branch of felid evolution arising from Proailurus took shape as the subfamily Felinae, the conical-toothed cats. The Felinae began with Styriofelis2, a genus of relatively small species around the size of modern Wildcats to Lynxes. Just as Pseudaelurus was the progenitor of all sabretooths, Styriofelis led to all cat species living today (and many conical-toothed species which are now extinct). They evolved alongside the sabretooths, often with many members of both subfamilies occupying the same environment and presumably with the same complex interrelationships we see among modern cat species today. Nine million years ago, the landscape that today surrounds modern Madrid had at least four species of now-extinct cats representing both subfamilies. Two species of Felinae, one Wildcat-sized (Styriofelis vallesiensis) and one Serval-sized (Pristifelis attica) must have occasionally fallen prey to two sabertooth species which were the size of a small Leopard (Promegantereon ogygia) and a Lion (Machairodus aphanistus). Fast forward to the early Pleistocene of East Africa and the Felinae branch had proliferated dramatically out of the shadows of their sabretoothed cousins. Around a million years ago, the Cheetah, Leopard and Lion shared the African landscape with at least three species of large sabretooth cats. Sadly, we will never know how six large cats from the two great felid subfamilies interacted but their relationships must have been intriguing.
FELIDAE FAMILY TREE
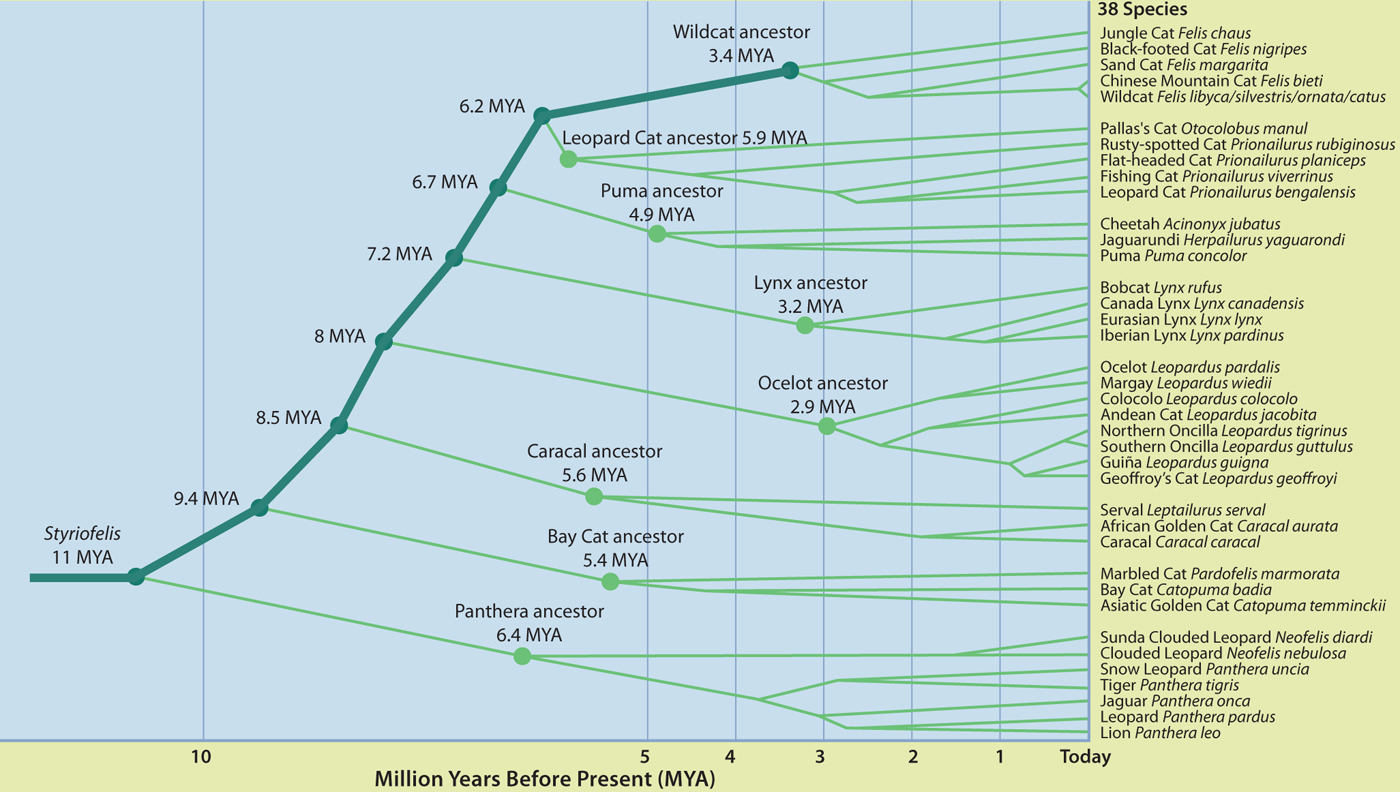
Comparing the DNA sequences of specific genes in every living cat shows their inter relatedness; the more similar the genes, the closer the relationship. By applying a known mutation rate for key genes and comparing the differences between species, it is possible to estimate approximately when lineages emerged and when individual species arose. (Redrawn from O’Brien & Johnson, 2007 ‘The evolution of cats’ Scientific American.)
Approximately 11 million years ago, the genus Styriofelis began a rapid evolutionary radiation in Eurasia that would ultimately produce all lineages of living cats. By combining genetic analyses of living felids with the felid fossil record, the evolutionary relationships of modern cats and thus the composition of each lineage is now fairly well delineated. The actual number of cat species however is still surprisingly fluid. This book recognises 38 living felids, following the latest taxonomy adopted by the IUCN Red List (see here) though taxonomy is not static and ever more sophisticated molecular analyses are likely to produce further revisions. Since 2006, genetic analyses have revealed the existence of ‘cryptic species’ of Oncillas and Clouded Leopards. In both cases, populations across the range were traditionally regarded as a single species based chiefly on very similar morphology until genetic analysis revealed two very similar but genetically distinct species hiding in plain sight.
1 This book uses the term carnivore as used in scientific nomenclature, i.e. only to indicate species of the Order Carnivora.
2 Styriofelis and Pseudaelurus are presumably closely related genera. Earlier classifications recognise only Pseudaelurus and assume that various Pseudaelurus species gave rise to the two major felid evolutionary branches.
THE FELID LINEAGES
There are eight widely accepted lineages of modern cats which together make up the subfamily Felinae. As the most ancient and distinct lineage, Panthera is sometimes treated as its own subfamily Pantherinae but it is more correctly treated as a lineage with the Felinae.
Felis Lineage
The last lineage to diverge and therefore the youngest branch. Five closely related species in one genus, Felis with an African and Eurasian distribution. The taxonomy of the Wildcat and closely related Chinese Mountain Cat is unresolved. Some authorities classify the Chinese Mountain Cat as a Wildcat subspecies Felis silvestris bieti based on very limited genetic data which is disputed. There is some genetic and morphological evidence that the European Wildcat should be classified as a distinct species from African-Asiatic Wildcats. Finally, the domestic cat is occasionally elevated as a full species Felis catus although this does not have widespread support.
Prionailurus Lineage
Five species in two genera Prionailurus and Otocolobus, with a tropical and temperate Asian distribution. The evolutionary relationships of Pallas’s Cat are poorly known, with evidence that it is intermediate to both Prionailurus and Felis but it is normally classified within the Leopard Cat lineage. There is some evidence for separating the Leopard Cat into two separate species where the dividing line is the narrow Kra Isthmus on the Malay Peninsula.
Puma Lineage
Three species in three genera which likely arose in North America and now have an Afro-Asian and American distribution. The Puma and Cheetah are not closely related to other large cats: they are essentially evolutionarily over-sized small cats that evolved larger bodies to occupy similar ecological niches to the Panthera genus.
Lynx Lineage
Four species in one genus with a temperate Eurasian and North American distribution. Lynx species are morphologically similar and distinct in all having bob-tails and ear tufts; it is unlikely that either feature has a strong selective advantage. This is among the most prey specialized of all felid lineages, particularly the Iberian Lynx and Canada Lynx.
Leopardus Lineage
Eights species in one genus with a Latin American (and barely USA) distribution. The eight members of this lineage diverged recently and are therefore closely related; hybridisation in the wild occurs between Southern Oncilla and Geoffroy’s Cat and possibly between Northern Oncilla and Colocolo (see here). The Oncilla is now classified as two species and there is some evidence to separate out the isolated Central American population as a third species. The Leopardus lineage differs from all other cats in having 36 chromosomes rather than 38.
Caracal Lineage
Three medium-sized species in two genera with an African and Asian distribution. The Caracal is not closely related to the genus Lynx despite a similar appearance, suggesting perhaps a distant, common ancestor carried Lynx-like features or the incipient genetic variation for them.
Pardofelis Lineage
The second cat lineage to diverge following the Panthera branch and therefore one of the oldest. Three species in two genera with a Southeast Asian distribution. There is some evidence that the Marbled Cat should be reclassified as two separate species either side of the Kra Isthmus.
Panthera Lineage
The ‘big cats’. The first lineage to diverge and therefore the oldest branch. Seven species in two genera, Panthera (the roaring cats) and Neofelis the Clouded Leopards. Most of the Panthera cats share the ability to roar, thought to be the result of unique modifications in the larynx and an elastic hyoid structure which supports it; and they cannot purr continually which is a feature of all other lineages. Snow Leopards and Clouded Leopards do not roar and both can purr.
THE SOLITARY CAT
Early in felid evolution, the Felidae evidently settled on an essentially solitary lifestyle which, as far as the fossil record tells, has worked successfully for most species that have ever lived. It likely arose with the cat’s ability to kill large prey without help. The feline template combines acute senses, hair-trigger reflexes, explosive muscular strength and a supple skeleton that promotes solo hunting; protractile claws and elastic wrists give tremendous control for grasping and handling large prey, while truncated, powerful jaws deliver a precise killing bite. Social carnivores like canids and hyaenas have more robust, less flexible bodies built for stamina to tire prey over long distances but which lack the cat’s solitary killing prowess. A lone Puma is able to take down an adult Elk but it takes a few Wolves to do likewise.
Prey is also the main factor in the spacing patterns of cats. All cats attempt to secure sufficient resources for two fundamental requirements, survival and reproduction. Cats need water as well as suitable habitat for hunting, to avoid danger and to have cubs but these are usually secondary to their prey requirements. Female cats are contractionists. They occupy the minimum area required to provide for themselves and their cubs, which is dictated largely by their prey; its size, distribution in the landscape and how frequently it is replenished. Females can occupy small home ranges where prey is abundant, stable and uniformly distributed. Small ranges are more easily defended from competitors so they also promote increased territorial defence and little overlap between adjacent female ranges. Where prey is scarce, fluctuates dramatically or migrates, female ranges are larger, overlap is greater and territorial defence is reduced, often to a small, exclusive core area (or areas).
Most felids show variation between these extremes according to the prevailing ecological conditions. A female Leopard living in very productive, prey-rich African woodland savannah can live her entire life in 10km2 compared to a Kalahari Desert female that will have a home range 50 times larger. Equally, some species always tend towards one extreme. Snow Leopards only inhabit areas with dispersed prey at low densities so they always live in large home ranges at low densities. No female Snow Leopard lives in any habitat where she can meet her needs in a home range of 10km2.
Male cats have ranges that are typically larger than predicted by food requirements alone. Males compete with other males for access to breeding females and they typically range over large areas in order to maximise the opportunities to encounter females; males are expansionists. They attempt to defend a territory from other males and monopolise females, so male ranges are usually larger than those of females. Where females maintain small, closely clustered ranges, a male can superimpose his territory over numerous female ranges with little overlap among males. Where females live at low densities in large home ranges, male ranges are typically larger and less exclusive with greater overlap between males.
The adults of most cat species undertake daily life alone (or are likely to, for poorly known species) but they are not asocial. Adults in the same area inhabit a complex social community of constant communication by scent-marks and vocalizations that allow familiar individuals to meet and rivals to avoid each other. Males and females come together to mate, and mothers are accompanied by the cubs of successive litters for most of their adult life but even among ‘classically solitary’ cats, adults often interact far more frequently and richly than is widely assumed. Unrelated adult Pumas in the Greater Yellowstone Ecosystem sometimes share large kills, perhaps because a large carcass can provide for many mouths and the costs of fighting over it are potentially too high. Similarly, male cats that apparently do little to provide for their cubs are still good fathers. Male Leopards and Tigers often spend time with their females and cubs, interacting amicably for extended periods including sharing kills.
Often dismissed as free agents that do little to raise cubs, male cats actually play a vital role. They patrol and defend the territory from immigrant males that would kill unrelated cubs if given the chance. Infanticide hastens the onset of oestrous in bereaved females, providing the new male with an accelerated window to produce his own offspring. Resident males repel such intrusions and provide mothers with the vital cloak of security that allows them to raise a litter to independence. In such a system, it is not so surprising that tolerance between familiar adults is manifested in regular social contact. No doubt, the same pattern – essentially solitary but with tolerant, sometimes enduring social relationships – occurs in other cat species awaiting further study.
In a few felids, sociality is constant and complex. The Lion’s extended families are built around a matriline of related adult females and their cubs that share a communal territory. Just as with solitary cats, Lioness range size is determined largely by prey requirements but rather than filling the needs of one mother and her litter, the range must meet the collective requirements of the entire female pride and its cubs. Male Lions live in coalitions that attempt to control as many female prides as possible which they defend from other coalitions – again, a scaled-up, highly social version of the basic felid socio-spatial pattern.
The Lion is the only cat that has developed the pride system. The reason lies, in part, again with prey. With both a rich diversity and high density of large herbivore species, African savannahs enable the formation of groups in large cats; simply put, there is enough to eat for big groups of big cats. Yet this does not automatically make pride-living the best strategy. In fact, the collective demands of feeding multiple mouths quickly overshadow the potential benefits of cooperative hunting. If food intake was the sole criteria, Lionesses would actually be better off alone or in pairs, particularly given that a Lioness is capable of killing all but the very largest prey species on her own. Indeed, it is that characteristic feline ability which probably fostered group living in Lionesses. In open savannah habitats, a large kill is a liability; it cannot be eaten quickly, is difficult to hide and is vulnerable to competitors. Lions evolved among all living large carnivores as well as three large sabretooth cats, and at least two extinct large hyaena species, any of which may have been able to dominate a single Lioness defending her kill. In such a highly competitive environment, it is better to share the kill with relatives who will help to defend it.

So-called solitary cats are far more socially elastic than often portrayed. Here, two adult male Leopards consort with a female in oestrous, Sabi Sand Game Reserve, South Africa. The two males are almost certainly territorial neighbours and ‘dear enemies’-rivals who know each other and who chose to be tolerant when the costs of fighting are high for both.
Ironically, group defence of kills would have created a further challenge for the ancestral Lioness. Female groups represent an extremely attractive resource to males and, just as with large kills, are likely to attract unwanted attention in open habitat. As incipient sociality emerged in Lions, so too did the risk of infanticide. Banding together would also have better enabled females to defend their cubs from foreign males. The evolution of the pride appears to be the Lion’s response to acute competition over their cubs and kills. It did not arise in other felids presumably because the same combination of selective pressure and ecological opportunity did not exist. An individual Tiger, Leopard or Puma might gain a modest advantage from teaming up with conspecifics but that only makes evolutionary sense when that benefit outweighs the costs of group living. Presumably, none had the Lioness’ persistent problem of defending very large, very obvious carcasses (or a similar ecological pressure), so remaining solitary is still the prevailing strategy for the cat family. Aside from Lions, the only wild cat to form enduring social groups is the Cheetah, in which males may form coalitions for similar reasons to male Lions (though female Cheetahs are solitary; see here).
THE SCIENCE OF STUDYING CATS
Wild felids are extremely challenging to study. Cats are generally rare, shy of people, and they often inhabit remote or inhospitable habitat. As a family, the Felidae does not lend itself easily to being observed, captured or monitored and many cat species are still very poorly known. The information in this book draws on thousands of scientific papers, reports and books written by researchers and naturalists spending months and sometimes years collecting data in the field. But how was that information collected? This section covers some of the main techniques we use to study wild cats.
Telemetry
Radio-telemetry has been a mainstay of wildlife research since the early 1970s. Until recently it relied largely on VHF (very high frequency) signals in which researchers use a receiver and directional antenna to detect a transmitter’s signal and thus locate the animal. VHF radio-tracking relies on a clear line-of-sight between the transmitter (on the animal) and the receiver (with the researcher). Distance, dense habitat, mountainous terrain and even large electrical storms affect reception while species with large daily movements are often temporarily lost and large parts of their range might be entirely inaccessible, for example during the wet season. All impact the likelihood of finding a collared cat and therefore the quality of the data.
VHF radio-tracking is being replaced by collars with a GPS module, using the same technology as in car navigation systems. A GPS-collar automatically logs its own position as often as the researcher wants, provided the collar can communicate with the GPS satellite system. GPS collars store locations for later retrieval or they can be relayed remotely to the researcher, via satellites or cellular phone networks (provided the collar is within range). GPS telemetry has major advantages over VHF telemetry in being able to automatically gather hundreds or thousands of accurate locations per collar and send them from the field to a laptop or cell phone anywhere in the world. Because GPS telemetry collects so much data, its results are more meaningful than similar efforts using VHF telemetry (see here for an example with Snow Leopards) and GPS collars are now sufficiently small to be useful for research of all cat species. Its main drawback is expense – a GPS collar costs 5–10 times as much as an equivalent VHF collar, and inexplicably high failure rates of the technology in the field. Every researcher who has deployed GPS collars on cats has experienced the terrific disappointment of collars failing to work as intended.

A radio-collared Andean Cat in the Argentinean Andes, one of a handful of this species that has ever been collared. Provided that collars are small, lightweight and fitted carefully, cats completely ignore them.
Telemetry is the mainstay of understanding felid spatial ecology. Locational data is used to calculate how much area a wild cat requires for its ecological needs – the home range (or territory, in case of those species which actively defend the range from conspecifics), as well as how it uses the area, for example, whether certain habitats or features in the landscape are preferred for certain activities such as raising cubs. If sufficient individuals (a large sample size) are telemetred, it also provides an insight into social and population ecology – how cats share the landscape with other members of their species, and how a population of cats behaves. Telemetry also provide an enormous amount of ancillary data on other aspects of felid behaviour including feeding and reproductive ecology, either by direct observation or gathering evidence after the event. Where direct observation is possible, think of radio-collared, vehicle-friendly Cheetahs in the Serengeti, researchers can view first-hand what cats hunt and kill, where they den their cubs and so on. GPS telemetry furnishes the same information even if the collared cat is never seen; location clusters direct researchers to possible kills or den sites that can be searched once the cat has moved off.
The greatest constraint of telemetry is the need to capture cats which is highly specialised, expensive and carries some risk to the animal. There is sometimes also concern about whether collars cause distress to cats, usually when collared animals are viewed by tourists in parks though this is largely misplaced. Long-term monitoring shows that radio-collars do not influence survival, behaviour or reproduction, provided that collars are correctly fitted (which, most importantly, entails minimizing their weight). Similarly, the use of compact ‘drop-off’ devices automatically removes a collar without the need to re-capture the cat to recover it. Radio-collaring will continue to be an essential tool for wild cat research, especially for those species which have never been systematically studied. Nonetheless, it must be recognised that telemetry is not always the most suitable method for the question, and that advances in less invasive techniques provide extremely useful alternatives. These techniques are discussed next.
Camera-trapping
The use of camera-traps is now the most common research technique used by field researchers working on wild cats (and many other species). Camera-traps use a motion-sensitive sensor to trigger the camera, automatically taking a photo of whatever passes by. It is an enormously useful technique that overcomes many of the challenges of observing or capturing elusive species. Camera-trapping provides a wide variety of data types. A simple inventory helps to refine the distribution and status of cats; camera-trapping is the source of many recent new range records as well as, regrettably, losses in range. Repeated camera-trap surveys in areas of historic range without any evidence of the species probably indicates local extinction; the loss of Tigers from Cambodia, Lao PDR and Vietnam is a particularly unfortunate example.
Surveys that use many dozens or hundreds of camera-traps placed in a specific array permit scientists also to estimate the number of cats in a population. The process relies on recognising individual cats from photos; fortunately, spot, blotch or stripe patterns are unique to individuals, like a fingerprint. With enough photographs, capture-recapture statistical models use the relationship between the number of unique individuals photographed (or ‘captured’ by the camera) and how often each individual is photographed (‘re-captures’) to estimate the density of cats in an area. The technique has some constraints, for example, the survey must cover a sufficiently large area and continue for a long enough to sample a representative fraction of the population, but not all individuals in the area need to be photographed for an accurate result. Recently developed analytical models show promise for estimating population density even for cats which lack a unique pattern of spots or stripes such as Lions and Pumas. Most of the density estimates given in this book are calculated from data gathered during camera-trapping surveys and many of the photographs would not have been possible without camera-traps.
Repeated camera-trapping at the same site is very useful for monitoring changes in the status of cat populations either by detecting changes in the estimated density or by another analytical technique called occupancy modeling. Occupancy analysis uses camera-trap photos (or any evidence of a species including observations and tracks) to estimate the proportion of large survey areas in which the species is present and absent. Occupancy models employ powerful statistics to compensate for failing to detect the species when it actually is present but not observed during the survey, and they do not rely on being able to recognise individuals. Just as for changes in density, repeated camera-trap surveys at the same site can detect changes in occupancy; a decline in occupancy may signal a population under increased threat that warrants increased conservation effort.
Cats at the molecular level
Biologists working on cats have always collected their scats (field-worker shorthand for faeces), mainly to understand what they eat. Cat scat contains undigested signatures of their prey, hair, feathers, scales and claws, which, by comparing to a reference collection under a microscope, can often be identified to species. Much of the information on feeding ecology in this book comes from studying scats, especially of the lesser-known, poorly studied species where scats might be the closest a researcher ever comes to an encounter.
Until fairly recently, data on diet was as much as a researcher might hope to extract from old pooh but advances in molecular technology have opened extraordinary new opportunities for research. Every scat contains DNA of its owner in the naturally shed cells of the intestinal lining. Provided the scat sample is sufficiently fresh or well-preserved (for example, by sun-drying in very dry habitats), the process of isolating the DNA and identifying its origin is now routine. Molecular scatology can identify which cat species are present in the sampled area, how many individuals and their sex. Applying the same capture-recapture analyses as used with camera-trap data even allows researchers to estimate population size and density; each scat left by each individual substitutes seamlessly into the analysis instead of photos. Just as for DNA isolated from tissue, fur or blood, faecal DNA can used to analyse how populations are related and connected to each other, and their phylogenetic relationships; for example, a 2013 analysis of 601 scat samples from across the Puma’s range shows three distinct groupings in North America, Central America and South America.
The sophistication and power of molecular analysis is now reaching the point that knowing which species and individual left a scat is just the beginning. Prey remains in the scat can be identified by the same process and the days of comparing hair samples under microscopes are beginning to wane. Researchers have even successfully isolated the DNA of internal parasites from scats. In the near future, molecular scatology will be able to furnish a complete genetic profile of the entire organism; the species, sex, individual, what it ate, whether it is carrying any parasites and even which viruses or bacteria it has recently encountered.
Using scats to learn about cats is particularly appealing because it does not require handling the animal (of course, samples of tissue and blood from captured or killed animals are routinely used for the same analyses). Hair holds some of the same advantages, at least in being able to identify species and individuals. Hair-traps – a sticky plate, barbed wire or wire brush – snag fur as the cat rubs against them although enticing felids to brush against them is a serious challenge. Ideally, DNA can be collected where cats chose to leave it; recently, researchers working in Sumatra demonstrated that it is possible to isolate Tiger DNA from urine-sprayed bushes used by the cats to demarcate territory.

Researcher David Mills sets a camera-trap during a study of the little-known African Golden Cat in Kibale National Park, Uganda. Effective camera-trapping relies on anticipating where cats will move; placing cameras indiscriminately in the landscape will produce few photos.