Hematopoietic stem cells, which are found in the bone marrow of adults, give rise to all cells involved in an immune response throughout the lifetime of an organism (see Chapter 2). Myeloid cells, part of the innate immune system, include antigen-presenting cells (dendritic cells and macrophages) and granulocytes (neutrophils, basophils, and eosinophils). Once mature, these cells exit the bone marrow. Some continuously circulate through the body, while others take up long-term residence in our many tissues and organs. Lymphoid cells, part of the adaptive immune system, also begin their development in the bone marrow, but complete it in distinct niches. B cells mature among the osteoblasts of the bone marrow, and some complete maturation in the spleen. T-cell precursors leave the bone marrow quite early and mature in a distinct organ, the thymus. During maturation, B and T cells rearrange genes that encode the B-cell receptor and T-cell receptor, and each generates a unique BCR or TCR (see Chapter 6). Immature T cells (thymocytes) and B cells undergo negative selection to rid their antigen receptor repertoires of autoreactive cells. Thymocytes also undergo positive selection for T-cell receptor specificities that recognize self-MHC (major histocompatibility complex)–peptide complexes with some affinity. The few immature lymphocytes that survive these selection events exit from the thymus and bone marrow as mature, naïve T lymphocytes and naïve B lymphocytes (see Chapters 8 and 9).
Naïve Lymphocytes Circulate between Secondary and Tertiary Lymphoid Tissues
About 30 minutes after entering the bloodstream, nearly half of all mature, naïve lymphocytes produced by the thymus and bone marrow travel directly to the spleen, where they browse for approximately 5 hours (Overview Figure 14-1). Most of the remaining lymphocytes enter various peripheral lymph nodes, where they spend 12 to 18 hours scanning cellular networks for antigen. A small number of lymphocytes (about 10%) migrate to barrier immune tissues, including the skin and gastrointestinal, pulmonary, and genitourinary mucosa, where they are in close contact with the external environment (see Chapter 13).
How do the millions of newly generated naïve lymphocytes find their antigenic match, if there is one? The odds that the tiny percentage of lymphocytes capable of interacting with an antigen (one in 105) actually makes contact with that particular antigen are improved considerably by a variety of strategies. First, lymphocytes circulate through secondary lymphoid tissues continuously. An individual lymphocyte may make a complete circuit from blood to tissues to lymph and back again once or twice per day, probing earnestly for antigen in secondary lymphoid tissues. Second, as we will see shortly, the organization of lymphocytes within secondary lymphoid tissues profoundly increases the probability that a lymphocyte will make contact with “its” antigen (Overview Figure 14-2).
Naïve lymphocytes destined for lymph nodes exit the blood at the high-endothelial venules (HEVs) in the cortex of the lymph nodes (Figure 14-3a). These specialized postcapillary venules are lined with distinct endothelial cells that have a plump, cuboidal (“high”) shape (see Figure 14-3b). This contrasts sharply in appearance with the flattened endothelial cells that line the rest of the blood vessel. As many as 3 × 104 lymphocytes move through a single lymph node’s HEVs every second.
FIGURE 14-3 Lymphocyte migration through HEVs. (a) An HEV in a lymph node as visualized by two-photon microscopy. Blood vessels are stained red and the HEV appears orange. (b) Schematic cross-sectional diagram of a lymph-node high endothelial venule (HEV). Lymphocytes (blue) are shown in various stages of attachment to the HEV and in migration through the endothelial cells (tan) into the cortex of the lymph node in the direction of the arrow. (c) Micrograph of frozen sections of lymphoid tissue. Some 85% of the lymphocytes (darkly stained) are bound to HEVs (in cross-section), and constitute only 1% to 2% of the total area of the tissue section.
Interestingly, HEVs fail to develop in animals raised in a germ-free environment. Investigators tested the possibility that HEV formation was dependent on antigen by surgically blocking the afferent lymphatic vessels, where antigen typically enters. Within a surprisingly short period, HEVs stopped functioning as access points for lymphocytes and in days the endothelial cells reverted to a more flattened morphology. These results show that endothelial cells receive and respond to signals from the lymph node.
How do naïve cells know that HEVs are the right site for entry into lymph nodes? The answer illustrates a common theme in immune cell migration: where cells go is determined by the array of surface protein receptors they express and the set of ligands expressed by cells they browse. Several different sets of interactions regulate lymphocyte homing to specific tissues before, during, and after an immune response. These include chemokines and their corresponding chemokine receptors, selectins, integrins, and adhesion molecules. We will consider some of these below and describe them more thoroughly in Advances Box 14-1.
HEVs express a specific combination of ligands, collectively known as an addressin, that is recognized by CD62L, a selectin that is expressed on the surface of newly generated naïve T and B cells (see Advances Box 14-1, Figure 1). Endothelial cells associated with other tissues, such as the intestinal mucosa, skin, or brain, express distinct addressins that allow entry of different subsets of white blood cells, including activated lymphocytes.
When naïve lymphocytes engage HEV addressins, they slow down and begin to roll along the blood vessel wall. This initiates a sequence of events that ultimately induce cells to leave the blood vessel by squeezing between endothelial cells, a process called extravasation.
Extravasation Is Driven by Sequential Activation of Surface Molecules
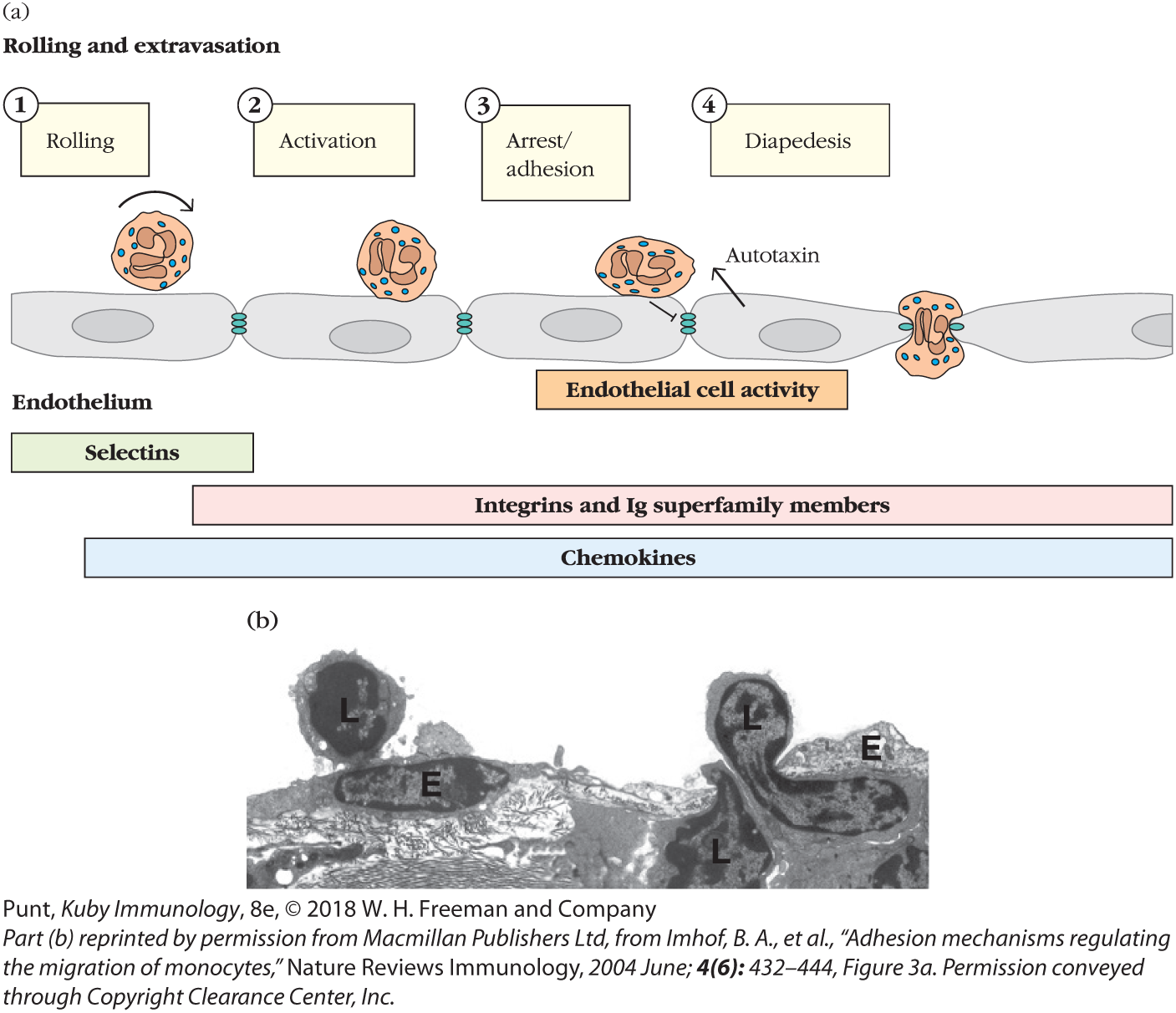
Extravasation is divided into four steps, each of which is regulated by distinct families of molecules: (1) rolling, mediated by selectins; (2) activation by chemokines; (3) arrest and adhesion, mediated by integrin interaction with Ig family members; and, finally, (4) diapedesis, transendothelial migration (Figure 14-4).
FIGURE 14-4 The steps of leukocyte extravasation. (a) Schematic of the major events regulating extravasation. A neutrophil is depicted, but the events depicted are applicable to all leukocytes. Tethering and rolling are mediated by binding of selectin molecules to sialylated carbohydrate moieties on mucin-like CAMs. Chemokines then bind to G protein–linked receptors on the leukocyte, triggering an activating signal. This signal, combined with the shear force produced by blood flow, induces a conformational change in the integrin molecules, enabling them to adhere firmly to Ig superfamily molecules on the endothelium. The arrest of a leukocyte also generates signals within endothelial cells that generate factors (autotaxin) that enhance leukocyte motility, loosen adhesive connections between cells, and induce extension of processes that help draw the leukocyte across. Ultimately, leukocytes crawl between endothelial cells into the underlying tissue (transmigration). (b) Transmission electron micrograph capturing lymphocytes (L) migrating through the endothelial cell (E) layer.
As naïve T and B cells approach the HEV in a lymph node, their surface CD62L latches on specifically to the adhesion molecule GlyCAM, which is expressed by endothelial cells. Because this interaction is relatively weak, the cell doesn’t adhere tightly to the blood vessel wall, but rather tumbles and rolls as the blood flows by. This slows the cell down long enough to allow new interactions to form. Chemokines decorating the surface of the endothelial cells interact with the chemokine receptor CCR7, which is expressed by T and B cells. Chemokine receptor signals induce a change in conformation of the lymphocytes’ integrins (e.g., LFA-1) that enhance their ability to bind tightly to ICAMs on the endothelial cell. Even the shear forces generated by blood flow contribute to enhanced binding and lymphocyte arrest. The naïve lymphocyte is now prepared to crawl between endothelial cells, driven in addition by attraction to higher concentrations of chemokines inside the lymph node.
Lymphocyte motility is enhanced by the activity of an enzyme, autotaxin, released by HEV cells themselves. In fact, endothelial cells are very active participants. They recognize the binding of an arrested lymphocyte and send signals that loosen adhesions with neighboring endothelial cells. They also reorganize their cytoskeleton and extend processes that help draw the lymphocyte between. All in all, it takes about 2 to 3 minutes for lymphocytes to pass between endothelial cells.
Naïve T and B cells are not the only cells that extravasate, of course. Any circulating leukocyte that recognize addressins expressed by particular endothelial cells exit the blood and enter the neighboring tissues. Only cells that express the right set of receptors for the addressins are allowed to cross. Effector cells, for instance, recognize addressins induced by inflammatory responses at the site of infection. However, they will pass by healthy tissues, ignoring endothelial cells that do not express appropriate addressins.
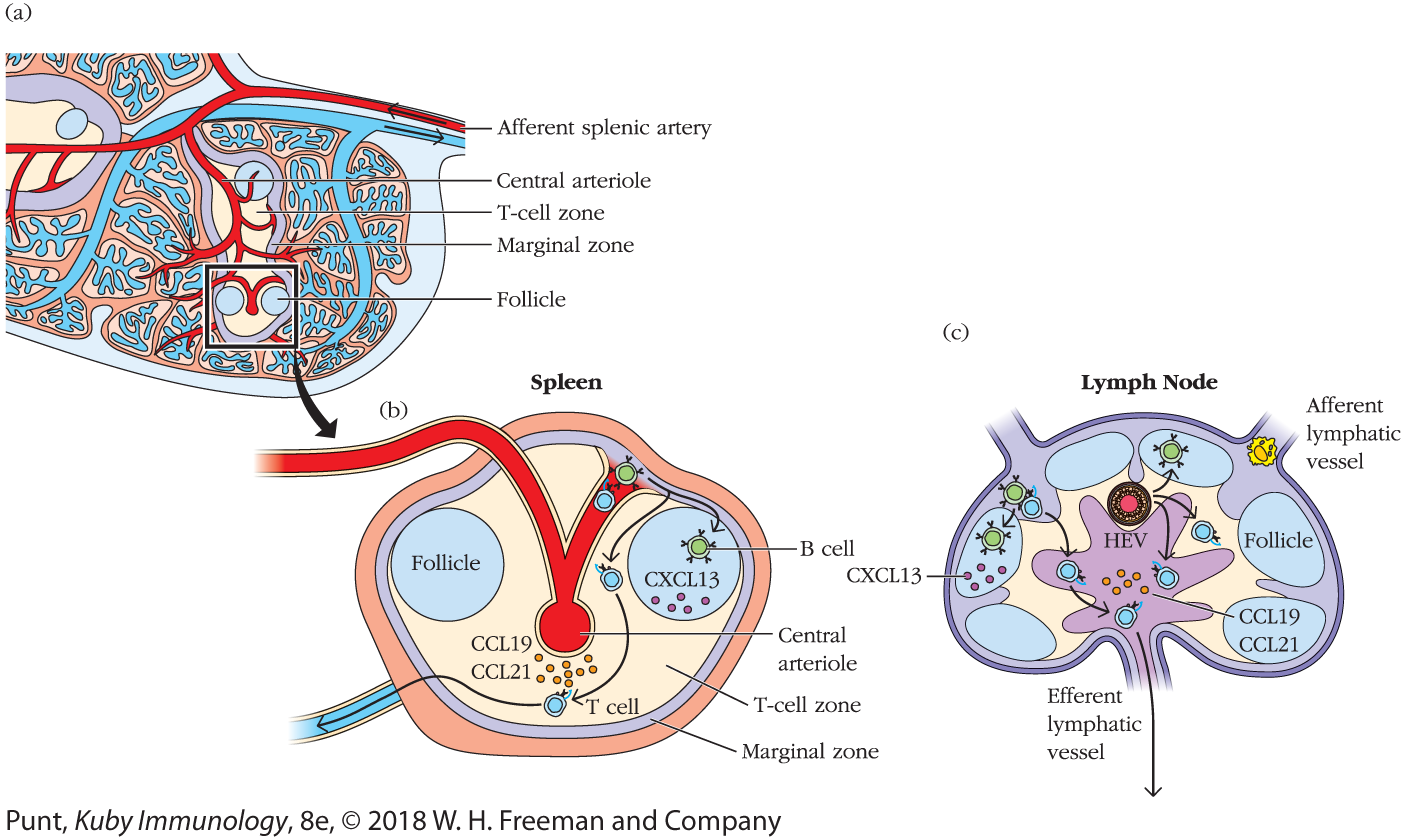
Interestingly, the spleen appears to have no HEVs. Arterioles release immune cells directly into the marginal sinus. (See Figure 14-5 for a comparison of approaches taken by spleen and lymph node.) Furthermore, the cues lymphocytes use to home to the white pulp of the spleen differ from those they use to enter the cortex of the lymph node and are not yet fully understood. Entry appears to require the coordinated activity of chemokines, integrins, and autotaxin, but is independent of selectins such as CD62L, which are important for lymph-node entry.
FIGURE 14-5 Lymphocyte migration in the spleen. (a) Schematic of spleen microanatomy showing the central arteriole, which releases circulating lymphocytes into the marginal zone, a microenvironment that separates blood from the white pulp. (b) T cells then migrate into the T-cell zone (PALS) and B cells migrate into follicles. Cells that do not meet their antigen usually exit via the splenic vein. In the lymph node (c), these migrations are regulated by chemokine receptor–chemokine interactions and guided by FRC and FDC networks, which are described in more detail in the text.
Naïve Lymphocytes Browse for Antigen along the Reticular Network of Secondary Lymphoid Organs
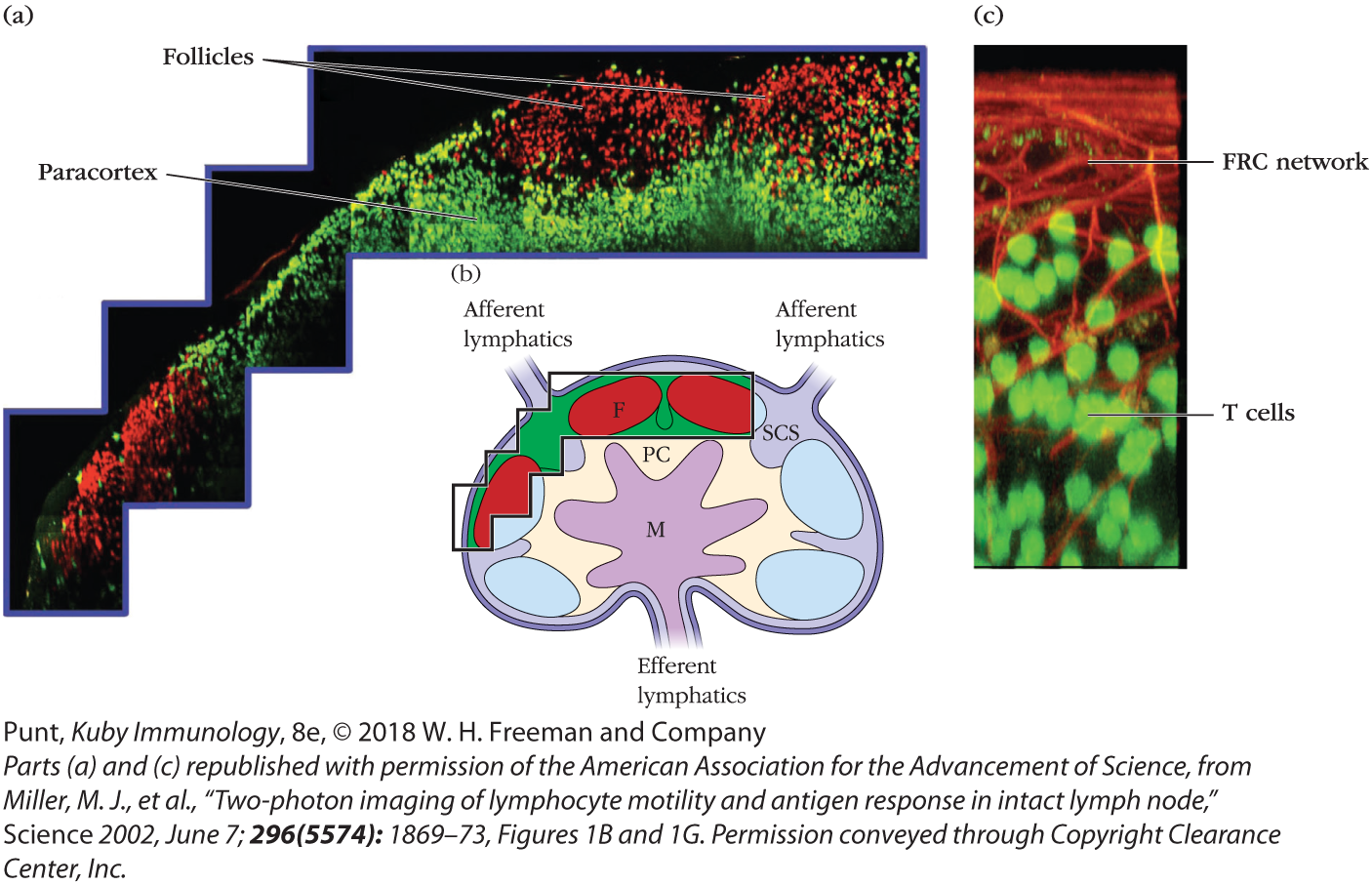
After squeezing between HEV cells, naïve B and T lymphocytes enter the cortex of the lymph node (see Chapter 2), where they are guided by different chemokine interactions to distinct microenvironments. The movements of naïve lymphocytes that enter and scan the lymph node are remarkable to watch. When these were first visualized, the idea that lymphocytes were rather dull and inert (an impression based on cells fixed on microscope slides) was immediately reversed (see Figure 14-6; see also Video 14-6v for an example; direct links to original sources for all videos can be found at the end of this chapter, on p. 545). These initial videos also inspired the discovery of the fibroblastic reticular cell network. Investigators recognized that fluorescently tagged lymphocytes did not move freely but were influenced by “invisible” structures. These were ultimately identified as the fibroblastic reticular cell (FRC) network of conduits and cells that provide roadways for naïve lymphocytes and antigen-presenting cells (see Figure 2-14).
FIGURE 14-6 Two-photon imaging of live T and B lymphocytes within a mouse lymph node. Fluorescently labeled T lymphocytes (green) and B lymphocytes (red) were injected into a mouse and visualized by two-photon microscopy after they homed to the inguinal lymph node. (a and b) T and B cells localize to distinct regions of the lymph node: T cells in the paracortex (PC) and B cells in the follicles (F). Antigen and APCs enter the subcapsular sinus (SCS). Cells leave via efferent lymphatics from the medulla (M). (c) A magnified image of T cells (fluorescing green) interacting with the fibroblastic reticular cell (FRC) network (stained red).
Naïve B and T cells spend many hours probing for antigen: B cells in B-cell follicles and T cells in T-cell zones (also called the paracortex) (see Figure 14-5). Naïve T lymphocytes, which express the chemokine receptor CCR7, dive in and out of the lymph-node parenchyma, using FRC tracts (see chapter opening image–associated Video 14-Ov). They are specifically attracted to this network because it is decorated with the CCR7 ligands CCL21 and CCL19. Naïve B cells that transit across HEVs into the lymph-node cortex also begin their travels along FRC fibers. However, because they express different chemokine receptors, including CXCR5, they soon change allegiance to the fibers established by the follicular DCs, which are decorated with the corresponding chemokine CXCL13.
The movements of naïve T and B cells in the spleen are guided by the same chemotactic cues and conduit network (see Figure 14-5). Once they find their way into the white pulp, naïve B cells are attracted to CXCL13 in the follicle and T cells are attracted to CCL19 and CCL21 in the periarteriolar lymphoid sheath (PALS).
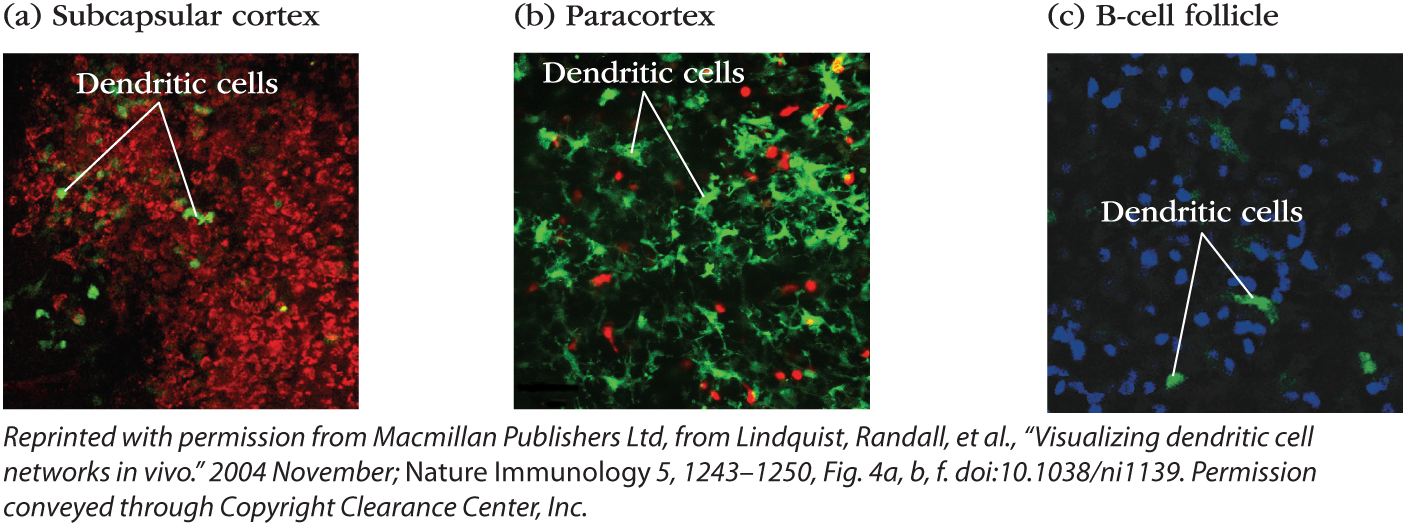
Antigen-presenting cells are found in every secondary lymphoid tissue and every microenvironment (Figure 14-7). Some are long-term residents and others actively migrate within and between tissues. Figure 14-7 shows dendritic cells in several different areas of a lymph node. Video 14-7v shows the activity of DCs (green) traveling among a bed of more sessile, long-term resident macrophages (red) in the subcapsular sinus of a lymph node. A population of migratory DCs also enters the deeper T-cell zone of the lymph node, where they crawl less vigorously and become part of the FRC network (see Figure 14-7b). They extend their long processes along the conduits to allow naïve T cells to scan their surfaces during their travels. A few DCs are even found in B-cell follicles (see Figure 14-7c), where they also present antigen.
FIGURE 14-7 Antigen-presenting cells are present in all lymph-node microenvironments. Intravital microscopy of inguinal lymph nodes of anesthetized mice, all of whose dendritic cells fluoresce green. Several areas of the lymph node are visualized (a-c). In the subcapsular sinus (a), green dendritic cells are surrounded by macrophages, which have taken up a red fluorescent dye. In the T-cell zone or paracortex (b), dendritic cells mingle with T cells (red). In the follicle (c), dendritic cells are not as numerous but can be found among the B cells (blue).
Early estimates using static imaging techniques suggested that up to 500 T cells probe the surface of a single dendritic cell (DC) per hour. However, more accurate dynamic imaging data reveal that one DC can be surveyed by more than 5,000 T cells per hour! Suddenly it is much easier to imagine that one in 100,000 antigen-specific T cells can find the MHC-peptide complex to which it can bind.
Naïve B cells spend a similar amount of time scanning for antigen. They travel on distinct cellular pathways, specifically the follicular DC networks in the follicles (Figure 14-8). They probe for antigen on the surface of the follicular DCs themselves, and can also bind soluble antigen that has entered follicles from draining afferent lymphatic vessels.
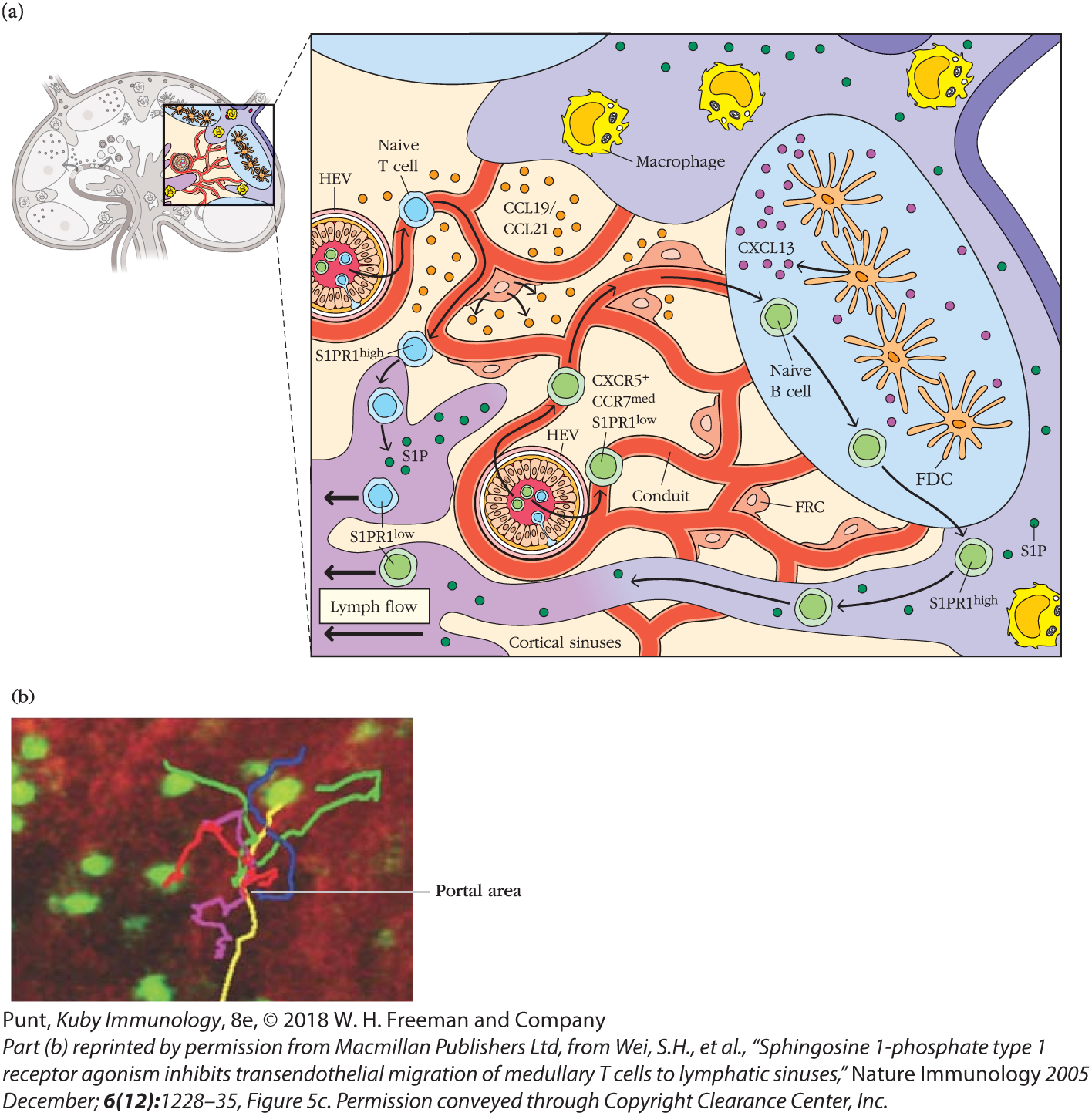
FIGURE 14-8 Lymphocytes exit the lymph node through portals in the cortical and medullary sinuses. (a) Schematic of lymphocyte traffic in and egress from a lymph node. T cells and B cells follow chemokine cues as they probe the FRC and FDC networks, respectively. If naïve lymphocytes do not find antigen within a certain time period they up-regulate S1PR1 and enter efferent lymphatic vessels from the cortex (B cells) or the medulla (T cells). (b) The movements of fluorescently labeled T cells (green) were visualized by two-photon microscopy in a lymph node where the medullary sinus stains red. The movements of several T cells are traced with colored lines and indicate that they leave at discrete sites called portals. See Video 14-8v.
If they do not find and bind antigen on their travels, naïve B and T cells up-regulate the sphingosine 1-phosphate (S1P) receptor, S1PR1, which allows them to leave (egress) secondary lymphoid tissues (see Figure 14-8a). Naïve lymphocytes circulating through lymph nodes leave via efferent lymphatics in both the cortical and medullary sinuses (see Overview Figure 14-2). Cells lining these sinuses express S1P, the ligand for S1PR1. S1PR1-S1P interactions induce naïve lymphocyte migration via specific portals (see Figure 14-8b and Video 14-8v). Once they have left the node, they return to the blood via the thoracic duct and resume their search for antigen in other lymph nodes. Interestingly, S1PR1 mediates lymphocyte egress from a wide range of other tissues, including the bone marrow and thymus.
Although the details of egress from the spleen are still being worked out, the red pulp is rich in S1P and clearly plays a role in regulating the exit of naïve B and T cells. However, most lymphocytes circulating through the spleen appear to exit into the bloodstream directly from the white pulp or marginal zone.

 Video 14-6v for an example; direct links to original sources for all videos can be found at the end of this chapter, on p. 545). These initial videos also inspired the discovery of the fibroblastic reticular cell network. Investigators recognized that fluorescently tagged lymphocytes did not move freely but were influenced by “invisible” structures. These were ultimately identified as the fibroblastic reticular cell (FRC) network of conduits and cells that provide roadways for naïve lymphocytes and antigen-presenting cells (see Figure 2-14).
Video 14-6v for an example; direct links to original sources for all videos can be found at the end of this chapter, on p. 545). These initial videos also inspired the discovery of the fibroblastic reticular cell network. Investigators recognized that fluorescently tagged lymphocytes did not move freely but were influenced by “invisible” structures. These were ultimately identified as the fibroblastic reticular cell (FRC) network of conduits and cells that provide roadways for naïve lymphocytes and antigen-presenting cells (see Figure 2-14).