2
Evolution, Groups, and Scaffolded Minds
Linnda R. Caporael
Rather than seeking to explain human mind and behavior by appeal to genetic costs and benefits, my aim in this chapter is to sketch an evolutionary–developmental context, hopefully one leading to new directions in empirical research on cognition and culture. The proposed framework responds to the physical, materially driven, embodied, and situated activity of organisms, which ultimately gives rise to, and results from, evolutionary processes and developmental constraints (Cole 1996; Hendriks-Jansen 1996; Ingold 2000; Rogoff 1990, 2003). Although concerns with culture and cognition seem distant from the wet labs of evo–devo biologists studying topics such as gastrulation (Callebaut, Müller, and Newman 2007; Newman, this volume), the approach in this chapter contributes to an early conceptualization of a human evo–devo perspective (see also Wimsatt and Griesemer 2007). I posit a structure of core configurations of face-to-face groups that plausibly has been repeatedly assembled over long spans of evolutionary history. Each generation recreates the dynamics of group structure through the situated activities of life tasks that scaffolds the development of offspring.
I use scaffolding as a shifting and relative term referring to the support for action or concepts using various resources, which may include (1) group structures or dynamics, (2) cultural–historical resources in the setting created by groups, or (3) endogenous resources. An example of all of these is when children (or adults) talk themselves through their own activity as they solve a difficult problem (Vygotsky 1978). A researcher could focus deeply on one scaffolding relation or on the integration of all three. The latter framework would represent biology, psychology, and culture as coevolved and coextensive: one level of function does not evolve and then become the basis for the next “higher”2 level although there can be situations where distinguishing them for analytic purposes may be useful.
The chapter begins with evolution and the role of face-to-face groups in human evolution. The dichotomy between genes and environment, unavoidable in neo-Darwinism, is replaced by attention to recurrence, or repeated assembly (Caporael 1997, 2003). This is followed by a model of core configurations in face-to-face groups, partially inspired by Hull’s (1988) trenchant observations of the demic structure of science. Core configurations are subgroups of face-to-face interactions that are posited to recur in daily life, ontogeny, history, and plausibly, as part of human evolutionary history. The empirical claim of the model is that an evolutionary/cultural history of living in face-to-face groups has shaped human bodies and minds, and that the repeated assembly of such group structures, whole or in part, scaffolds human development, cognition, and culture (Wimsatt and Griesemer 2007) across generations, even in large-scale society.
The second part responds to the question: what are groups for? The short answer is that groups scaffold coordination (Caporael 1997, Sterelny 2012). More specifically, core configurations serve as an interface between individuals and habitats, where epistemic projects are constantly in construction. These projects, always ongoing and under debate, are concerned with attunement, the alignment of situated activity, “factual knowledge,” and large-scale group coordination. The third part, which brings together elements of the earlier sections, describes the forging of links between generations, specifically a tightly coupled system where the infant scaffolds parental performances that eventually scaffold the infant into its human community.
Evolution Materialized
This section describes the repeated assembly of core configurations (Caporael 1997, 2003). It draws on multilevel selection models and developmental systems theory. Repeated assembly extends the central ideas of hierarchical organization in biological structure and parity of a number of developmental resources, rather than waving a hand to genes and environment. It also naturally articulates with Griesemer’s (this volume) conceptions of material overlap and hybridity and with Wimsatt’s (this volume) focus on the variability and generativity of entrenchment. While this evolutionary framework is general enough to support a number of different accounts for human evolution, I focus on one, a model of core configurations that emerge as a product of bodies, tasks, and activities in face-to-face groups (Caporael 1997, 2007; Brewer and Caporael 2006). The background for this model is an empirical observation: the hominin commitment to bipedalism was also a commitment to group-living. A small-brained, solitary biped lacking tools or biological defenses would more likely be dinner than diner. The consequence of those commitments was a species that is obligately social, unable to reproduce and survive without a group.
Repeated Assembly
Multilevel selection theory (Buss 1987; Szathmáry and Smith 1995) has an emerging affinity with a variety of developmental systems perspectives (Oyama 1985; Hendriks-Jansen 1996; Li, this volume). The first recognizes evolutionary processes at multiple levels of organization, including individuals and groups; the second offers a broader notion of causal resources inherited in development than does neo-Darwinism. This expanded evolutionary theory recasts Darwinism in a form that can accommodate the human case without nature–nurture dualism. The key insight is the nested, hierarchical organization of life. Rather than a single level of selection, centered on genes or “gene–environment” interactions (“the environment” referring to the external habitat), the hierarchical view identifies multiple levels of selection. Under this description, “the environment” refers to the context of entities. Thus, the gene–environment interaction occurs between DNA and its cellular machinery, not DNA and the habitat as is usually implied in references to “gene–environment” interaction. Evolutionary history has been the sequential elaboration of levels of selection—macromolecules, cells, individuals, and on occasion, groups (Buss 1987). Evolutionary analysis, distinct from other kinds of biological analysis, focuses on the interface between levels—the conflicts and synergisms between levels of organization.
Biological entities are hierarchically organized (e.g., DNA, cells, tissues, organisms, groups) in nested part/whole relationships, and organisms are the contingently developmental result of various genetic and epigenetic resources (e.g., genes, nutrition, oxygen, artifacts, language environment, social roles). Resources have different temporal scales and cycles of reproduction. Natural selection results from the materially situated activity of embodied entities with environments having—from the entities’ perspectives—reliable and recurrent affordances for activity (Caporael and Baron 1997). Thus, repeated assembly (Caporael 2003) points to recurrences that we can observe—recurrences from generation to generation as well as recurrence across cultures. Not only do organisms repeatedly assemble, but so also do the products of organisms, including human ideas, artifacts, and cultural practices.
Although sketchy, this rendition offers an alternative to nature–nurture dualism in evolutionary thinking. Repeated assemblies are recurrent entity–environment relations composed of hierarchically organized, heterogeneous components (which may themselves be repeated assemblies) having differing temporal scales and cycles of replication. Neither genes nor environment are privileged in this view although the microbiologist would still focus on genes and the educational psychologist might focus on classrooms. They have expert perspectives that enable them to recognize affordances in their respective intellectual habitats.
Here I mention two points especially relevant to this volume. An appeal to recurrence is fairly obvious although we are most interested in a particular kind of recurrence, that which is recursive, where the seeds of the next cycle are part of a current cycle. These repeated assemblies constitute lineages connected by material overlap (Griesemer, this volume). However, there are other kinds of repeated assemblies, such as those that involve copying, which fall under the general term of repeated assembly because we often don’t know whether a repeated assembly is recursive in a genealogical sense or repeats for other reasons (Caporael 2003). For most social scientists that does not matter. What does matter is the observation of recurrence and the possibility of an empirical investigation that is not foreclosed by genetic determinism.
Components are heterogeneous because they recur over different time scales. DNA, which is deeply entrenched, recurs in cycles that extend over macroevolutionary time; at the other extreme, social rituals are far less entrenched and can rise and fall within a lifetime (affecting phenotype but having no systematic selective effects). Resources may change within lifetimes (e.g., a specific cultural item), indicating low reliability of recurrence but little about causal pathways. In short, assemblies may persist on geological scales, on historical scales, in the Smith family, or in Chris Smith. This view of resources parallels Wimsatt’s (this volume) discussion of maintenance entrenchment as well as overlapping and embedded entrenchments in ontogeny.
What is not a repeated assembly? The answer will often depend on level of analysis and perspective. Some repeated assemblies may result from copying or recur because of the dynamic properties of physical systems. Moreover, such properties—temporality, size, and space, for example—can be significant contexts for evolutionary and developmental processes (cf. Newman, this volume). Any specific individual is unique—even one of a monozygotic twin set. No twin has precisely the same experiences as the other. Organisms in general are repeatedly assembled. Often the product of a repeated assembly with specialized functions is unique. For example, the processes involved in the healing of wounds are repeated assemblies in organisms. However, the specific scars that result are the unique results of an individual’s history. At first glance, it seems that errors would not repeatedly assemble. However, as the work of Allen (this volume) and a substantial literature in developmental and cognitive psychology indicates, there are repeatedly assembled errors resulting from specific contingenies in developing systems, or as a product of the researcher–subject collaboration (Griesemer, this volume).
Core Configurations
Among the many items humans and their ancestors have repeatedly assembled are groups. One look at the human body—a long period of immaturity, no claws, pitiful canines, no hidden sacs of toxic sprays, and not even four feet—and it is clear that such specifics of bodily form co-evolved with group living. Groups scaffold individual and habitat (e.g., protection from predators; location of patchy, distributed resources), just as the multicellular organism functions as an interface between cell and habitat. In general, the situated activity within groups bracket developmental trajectories mediating selection. Moreover, too large a group strains the carrying capacity of the environment; too small a group risks perishing. Given such constraints, there would be a limit on group size and one would expect specialized adaptations for the development and maintenance of group living. Depending on morphology and ecology, all biological life must engage in various tasks for self-maintenance (Lyon, this volume), some of which may be related to group structure and size.
Anthropological research on prehistoric remains and modern hunters and gatherers suggest a fairly common and surprisingly consistent group structure around small hunting groups, residential bands, and seasonally meeting macrobands of about 3–5 individuals, 30–50 individuals, and 100–500 individuals, respectively (Birdsell 1972, Hassan 1981). A lack of variation in hunter-gatherer groups puzzles anthropologists. It remains more or less the same on different continents, in groups exposed to different predation risk, or groups operating in different habitat types (Layton and O’Hara 2010). Existing archeological and fossil evidence indicates a similar lack of variance in size and structure, hence the term “magic numbers” (Lee and DeVore 1968). Such consistency suggests that embodiment is more significant than ecology, especially for the smaller group sizes, which are likely to be more deeply entrenched. While details and orientation differ, the core configurations model aligns with work by Gamble and Gittins (2007) and Dunbar (1993), at least with respect to orders of magnitude for group size, to which I turn next.
The hunter-gatherer research plus considerations of bodily form, tasks, and group size give rise to the model of core group configurations (Caporael 1997). Core configurations are subgroups of face-to-face groups posited to repeatedly occur throughout human evolutionary history as a joint function of subgroup size and modal tasks. Properties of the body serve as (maintenance) scaffolds bracketing co-evolved configurations. A simple example can illustrate this point: a dinner of six people can sustain a common conversation around the table. Dinner for twelve (which seems to always happen at a society meeting) breaks up into conversational groups of three to five people. Features of body and task scaffold bounded groups. Similarly, 100,000 years ago, only so many people could huddle around Ms. Flintstone to examine and debate the edibility of a mushroom she has in the palm of her hand. Twenty people would have been too large a group, but just right to huddle around the campfire to hear a story. The general idea is that capacities that have been repeatedly assembled as part of core configurations in human evolutionary history will leave fingerprints on current capacities, and provide traction in areas of research that current neo-Darwinism lacks.
Table 2.1 presents a model of idealized, artificial hunter-gatherer groups. Although it is narrative in form, intellectually it serves the same purposes of understanding discussed by Schank and colleagues (this volume) for agent-based models. (The ongoing comparison is with individual-based models.) It lists four core configurations: dyad, task groups, deme (anthropology’s band), and macroband or macrodeme, with a range of group sizes and likely candidates for modal tasks. Modal tasks are common features among hunter- gatherers; obviously, there are many more tasks that can be described from a number of perspectives and levels of analysis. From an evolutionary perspective, each configuration affords possibilities for selection, suggested in the far right column. Thus, the dyad affords possibilities for microcoordination such as facial imitation in a mother-infant dyad and the automatic coupling of gait that occurs when two people walk together; the workgroup affords possibilities for distributed cognition (discussed later); the face-to-face group affords a shared construction of reality, which at some point also mediates interaction in macrobands and other superordinate group configurations. Macrobands stabilize and standardize language and psychologically “liberate” people from face-to-face groups through an impersonal but collective group identity (Brewer and Gardner 1996).
Table 2.1
Core configurations
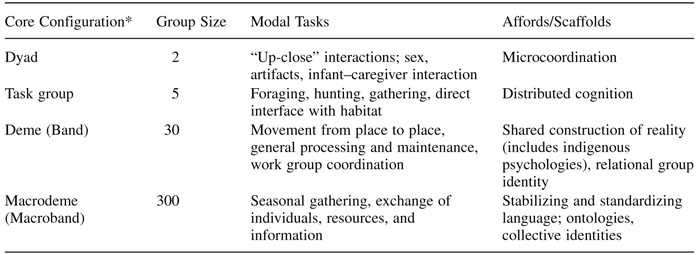
*Core configurations are a joint function of morphology, group size, and task. Except for dyads, the group size numbers should be considered as basins of attraction for group sizes in a range roughly plus or minus a third of this number. In all cases, the names of core configurations are not meant to represent roles or even labels but rather distinctive kinds of situated interactions with humans and nonhumans. The term “bands” is used to refer to (idealized) hunter-gatherers; otherwise the terms associated with “deme” are used to refer to the model.
The order of configurations also roughly suggest depths of entrenchment, from the deep entrenchment of dyads through relatively shallow entrenchment of macrodemes. (There may be multiple specialized affordances, but for clarity and simplicity, one per configuration will do at for this discussion.) The same column can also be read from a developmental perspective as core configurations repeatedly assemble, in evolutionary time, in ontogeny, and in daily life. As infants develop, not only do they become independent, they participate in an increasing range of interdependence. Their widening scope of interaction increases demands for reciprocity, skills, memory, and social judgment.
A few provisos should precede the more detailed report that follows. The group sizes in table 2.1 should be read as basins of attraction rather than absolute numbers, and likewise, the labels attached to groups are meant to be suggestive in describing interactions rather than relationships. Importantly, size alone does not constitute a core configuration. As an illustration, consider five people going to the twenty-fifth floor in an elevator. As they privately ruminate on dinner, what the boss said, who walks the dog tonight, the elevator slams to a stop between floors. All ruminations cease, and the five individuals are transformed into a group, sharing perceptions, memories of previous stoppages, suggestions of what to do next and so on. It has become a group as a function of shared fate. Given the apparent differences between hunter-gatherers—themselves highly diverse—and humans today, capacities evolved in core configurations must have evolved so as to scaffold activity in novel conditions, a topic for later discussion. Note that in addition to core configurations (those with a plausible evolutionary developmental history), non-core and cross-cutting group structures are anticipated.
Finally, human bodies and situated activity constrain, entrain, and scaffold typical boundary conditions for core groups. The available “physics” of body form, energy, time, and space roughly bind face-to-face configurations into their persistent band structures (cf. the parallel to the physics of multicelluarity in Newman, this volume). This does not mean that the “physics” can’t be artificially altered; indeed one of the interesting ways of testing the model would involve whether, and how, core configurations are maintained in the face of novel artifacts that substitute material body, space and time with new media.
Human Demic Structure
The core configuration model was partly inspired by David Hull’s (1935–2010) exemplary description of the organization and practices of working scientists (Hull 1988; all succeeding references are to this work; see also Caporael 1997). In this section, I use his notion of the demic structure of science for three purposes: to further illustrate the core configuration model (and its continuity), to suggest that group activities are also for scaffolding knowledge—what I refer to later as epistemic projects—and to acknowledge the credit he deserves. Hull (1988, 59, 433) seemed to use the terminology of demic structure in two ways; first, as a description of social-structural processes of science, and the other, in a more biologically conventional sense of deme, to mean a breeding population. His conceptual inspiration was Sewall Wright’s shifting balance theory, which includes selection within and between demes, also conceived as subdivisions of a breeding population. The latter use of the term demes is familiar to biologists, but it was the former use that’s put to work here.
Hull’s description of the demic structure of science is surprisingly similar to typical organization among nomadic hunter-gatherer groups above: a nested hierarchy composed of task groups (research groups), microbands (conceptual demes), and seasonal macrobands (invisible colleges). To these configurations, we can add the dyad. In both instances—in science and among hunter-gatherers—group-size numbers at these four levels are fairly constant; dyads, task groups of about 3 to 5 individuals, residential bands or conceptual demes of 30 to 50 individuals, and a macroband or invisible college of 100 to 500 individuals (Birdsell 1972, Hull 1988, 366). In both cases, isolates have reduced viability, and must be part of a subgroup with sufficient “critical mass” to persist. Like hunter-gatherers, individual scientists cannot usually begin a new group as social isolates; they must bring some of the old group members with them or attract new recruits.
In addition to structure, groups scaffolded activity similarly in the demic structures of scientists and hunter-gatherers. Learning to fashion specialized tools, in stone or other materials, or to use specialized laboratory techniques is a hands-on, situated activity (like golf or tennis). Such learning requires feedback from materials, equipment, and an experienced mentor if novices are to develop finely tuned sensory-motor microcoordination. Young scientists learn their specialized tasks and languages in the research team where older pre- and postdoctoral “siblings” and professors scaffold a “zone of proximal development” (Vygotsky 1978). Similarly, learning the complex skills of a specialized task (Micronesian navigations, Hutchins 1995, comes to mind) requires scaffolding from equally expert elders. Small research groups or intensely social small hunter-gatherer groups engage in numerous activities, but two stand out: intense interaction with the habitat and with task group members. A great deal of the daily work in science involves interpreting ambiguous or anomalous data, examining, pointing to, and discussing charts and printouts, much as hunters and foragers work to interpret ambiguous signs of an animal and its condition on a trail (Shaw-Williams 2009). As mentioned earlier, task groups of four to six people appear to distribute cognition; that is, they share tasks such as perception, classification, inference, memory, and contextually-cued responses in skilled practice, interpretation of data, and in interaction with uncertain habitat features.
Hunter-gatherer bands and scientific demes are also loci of practical skills and a clearinghouse for “common knowledge” (Gerson, this volume), some of which may be mythical and derived from macrodemes/society meetings, some of which may be acutely attuned to local conditions, from detailed knowledge of other people to that of the local ecology. In traditional hunter-gatherer groups, the band is the first configuration that can be self-sustaining for survival and child rearing (but not reproduction). The band is the staging ground for domestic life, including task group coordination, and for cooperative alliances, which are the basis for fissioning when the community exceeds resources or is fractured by conflict. Scientific demes are different in some of these respects, illustrating the cross-cutting group structures of modern conditions. For example, departments are typically staging grounds for academic “domestic life,” and anonymous groups (payroll office, the funding agency) control widely accepted, but nevertheless symbolic, tokens of resources for individuals.
For both hunter-gatherers and scientists, seasonal macroband meetings (or yearly conventions) are important for the exchange of myths, gossip, and information about more distant areas and groups (Griffith and Mullins 1972). Macrobands and scientific meetings are also arenas for competitive games as well as the affirmation of common worldviews, the maintenance of languages (hunter-gatherers) and idiolects (scientists), and the exchange of people (mates, new PhDs, or disgruntled members). For science, unlike hunter-gatherer groups, the production of articles in journals may eventually be incorporated as “common knowledge.”
Let me post a warning sign here. The analogies between hunter-gatherer groups and scientific communities partially result from the level of abstraction in the short descriptions of activity. A yearly scientific convention and a yearly meeting of aboriginal bands will have little apparent similarity should you drop in on them. The crucial question, where research is needed, is to “follow the body” in both cases.
David Hull saw himself as providing a “general analysis of selection processes … applicable equally to biological, social, and conceptual change” (284). Evaluating that claim based on his arguments is a daunting task: he was writing at a time when there were too many competing metaphors and analogies, his own among them; the language of biological organizational hierarchy mostly stopped at the organism, and contemporary evolutionary thinking had no language beyond self-interest for theorizing sociality (400). Nevertheless, the brief analysis in this section suggests that his notion of demic structure may have captured something enduring about group structures.
What Groups Are For: Coordination
Earlier in this chapter, I proposed that group living envelops and scaffolds development by mediating interactions with the habitat. Likewise, it imposed other demands on individuals to develop and maintain group membership. In this section, I suggest that groups scaffold different scales of coordination, with face-to-face groups (core and non-core) coordinating people and things. Large-scale coordination as in modern societies depend on symbolic mediation coordinating aggregates of individuals, cross-cutting groups, and bureaucratic organizations.
Epistemic Projects
The comparison of the tasks and activities in demic structures suggest a comparison between Bruner’s (1986) paradigmatic and narrative modes of thought. Paradigmatic modes, of which science is one version, are expected to lead to factual understanding; narrative modes are expected to be lifelike, to provide different views of subjectivity, and often to connote values and moral precepts. These modes of thought appear to serve two different epistemic functions. One attempting to reduce ambiguity and the other attempting to create social coordination, to some extent paralleling the situated activities in task groups and demes. Another epistemic mode, attunement, is tactile–kinesthetic (Sheets-Johnstone 1990), and may be related to the body and to dyadic interactions, as we will see later. Seasonal macrodemes, discussed in the following sections, appear to scaffold a variety of phenomena within and between groups, a discussion in the next section. Although these modes can be loosely associated with different configurations, they are more often interwoven in the production of epistemic projects. We can describe an epistemic project in a general way as a coordinating process producing knowledge of an interface between a group and its habitat. This is not solely a human achievement.
While it may seem odd to think of nonhuman animals as having epistemic projects, animals-moving-in-groups is an epistemic project, achieved through various processes of scaffolding and attunement (Boinski and Garber 2000). Coordination involves which individuals make decisions about when to travel and in what direction; how other members of a group know the decision; the purpose or goal of moving, and if the movement is for prey, the costs and benefits of movement given the expected outcomes. Some social mammals have core configurations as described above: dyads, daytime feeding/hunting groups, and evening sleeping groups. Often some kind of recognition enables subgroups to rejoin larger affinity groups. There is also evidence for “social rallies” that facilitate transition from rest periods to hunting, with synchronization of behavior and division of labor with some individuals adopting specialized roles for the hunt depending on the roles that other group members adopt (Holekamp, Boydston, and Smale 2000). Different species, because they have different morphological and ecological characteristics, will have different coordinating means (e.g., scents, sounds, movements that cue action) for producing an interface between group and environment.
In the human case, initial investigations of epistemic projects are likely to offer more insights in circumstances where the epistemic project is aligned with a core configuration. Consider airport control tower personnel gathering around a radar screen to discuss and interpret signs of possible danger given by ambiguous blips on the screen are examples. There are clear constraints on this kind of distributed cognition. The body itself limits the number of people that can communicate in a meaningful way and the spatial constraints of how many people can simultaneously scrutinize and point to details on a radar screen. Note here that task configurations do not guarantee optimal, or even good enough, data collection or analysis—we have a considerable literature on social loafing, polarization, clique selfishness, and other varieties of group dysfunction (cf. Poole and Hollingshead 2005, Theiner, this volume). Nevertheless, relative to individuals or other core group configurations, the small task group of about five people appears to be the most efficient for serving paradigmatic epistemic functions.
The tower control scenario illustrates dramatic changes in material, temporal, and social organization that have been made over past millennia and especially in the last 300 years. The evolutionary-developmental cognitive processes we hypothesize to have evolved largely in face-to-face interaction must be at a level of analysis, or “grain,” capable of scaffolding activity in novel domains (Caporael 1997; Smith and Semin 2004). (Recall—we don’t really know what these processes are, although there are empirical hints. The goal in this chapter is to sketch a landscape, and given such a perspective, ask how might we recognize, investigate, describe or discover details of social cognition?) Likewise, the worlds of the artificial designed and built by humans must in some way reflect the operation, the scope and scale of human cognitive processes. Conventional thinking seems to draw a line of demarcation between an environment of hunter-gatherers and that of builders, designers, and technologists. However, just as modes of thought overlap, connected by the materiality of the human body and group interdependence, modes of interacting with the habitat should also be expected to overlap (W. Wimsatt, this volume). Decomposing an interaction should provide new material for an evo-devo perspective on human minds that enables an exploration of different scales of repeated assembly and entrenchment.
A quick illustration: airline flight could be a prototype for an “evolutionarily novel” activity, “unnatural” for humans. Nevertheless, consider the pilot and copilot in a plane. Fiddling with their controls engages fine hand–eye coordination, a relatively ancient activity, repeatedly assembled throughout hominine history, deeply entrenched, and scaffolding a huge range of downstream skills. In contrast 400 strangers paying a cost to share the close quarters of the plane is a novel dimension of human experience, one to which most travelers give little thought beyond, perhaps, what book to bring or movie to load on the iPad.
Just as capacities that evolve and develop for one configuration can often be invoked to use in another, technology can also scaffold or bridge situated activities of configurations. A group of 500 people given an order to march on a football field are likely to clump and straggle, but if a rousing marching song is broadcast, they can hardly avoid keeping time. Some institutions have been particularly successful in exploiting core configurations, the military being an important example. It is unlikely that any man has died for his country, but many a poor soul has sacrificed himself in the heat of battle for his comrades-in-arms.
Epistemic Projects for Large-Scale Coordination
Most approaches to the evolution of sociality are overwrought by the problem of defectors, free-riders, or cheats; hence most evolutionary theorists take the evolution of altruism to be the central problem in a view of sociality founded on atomistic individuals. In the repeated assembly of core configurations, sociality is a solution to the problem of defection because it creates some level of interdependence as well as various sanctions that can constrain self-interest, at least until it passes certain group-defined boundaries. Rather than a model of individual agents, large-scale coordination begins with a model of macrodemes in an evolutionary context.
Macrobands, the seasonal meetings of a group of loosely related bands, initially seem to be inert. The situated activities of hunting, foraging, playing, and interacting socially in macrodemes superficially is much the same as that that in demes. Nevertheless, macrobands have been active sites of biocultural evolution for at least two reasons. First, macrodemes recur intermittently. Thus they scaffold the development, stabilization, and standardization of language; the circulation of genetic, artifactual, and organization innovations; and new forms of abstract, collective social identities that operate beyond the level of the relational identities in face-to-face groups. Second, macrodemes have been historically transitional. A key assumption is that macrobands tend to be seasonal in the evolutionary past because of limitations of resources. However, as hunter-gatherers became more sedentary, settled macrobands could become settlements, settlements become villages, and villages become towns and urban centers. Macrodemes (and social identities) are reformed around new kinds of associations. With important exceptions, the roles of macrodemes in modern cultures have changed, largely replaced by institutions (Fiske 1991, Gerson, this volume) and social networks (Dunbar, Gamble, and Gowlett 2010), where individuals are members of multiple cross-cutting groups, and core configurations can persist as the situated activity of unrelated people with a shared epistemic project, much as the jazz musicians described by Gerson (this volume).
There is too much in the sketch above to elaborate here. What it does suggest is that macrodemes can scaffold a large range of coordinating epistemic projects that can cross demic structures and scaffold situated activity in core configurations. For the remainder of this section and in the next, my focus will be on a higher-order epistemic project and a subsidiary project encompassed by it. Face-to-face groups, now and in the past, as previously mentioned, are also increasingly seen as having necessary epistemic functions, particularly because of the distinction between small-group relational identity where knowledge of others is face-to-face versus large-scale collective or group identities where knowledge of others is symbolically mediated by shared origin myths, symbolic artifacts, origin stories, and folk psychologies. Where the main epistemic tasks in small groups might involve personalized trust and honesty, the epistemic issues in macrodemes seem more concerned with good stories and values, obligations, and duties. Stories, parables, norms, laws, circulated images, and urban legends are among the various forms for such abstract knowledge. Humans tend to naturalize many such bodies of knowledge, making them real (i.e., enabling contingent consequences). For example, witchcraft is common across cultures. Although its role in mediating social life is quite variable, where a belief in witches and supernatural forces is legitimized, the evidence supporting witchcraft accusations can still be evaluated systematically at local levels (Caporael 1976; Evans-Pritchard 1937).
The more or less coherent connections, constructions and negotiations of group identity, shared reality, social representations, controlling stereotypes, artifacts, folk psychologies, and cultural narratives such as parables, founding myths and origin stories, constitute social imaginaries (Taylor 2002). Taylor writes of the social imaginary as the way that ordinary people imagine their social surroundings. Imaginaries are collectively shared landscapes that make common practices normative and possible and provide a widely shared sense of legitimacy. His focus is on the superordinate group in the large sense of the “Western social imaginary.” However, the term has been applied to other levels of organization as well. The associations between demic structure, epistemic functions, and identity are rarely mutually exclusive, and one of the remarkable features of modern society is the enormous number of social roles which imaginaries coordinate.
Where do social imaginaries come from? Ultimately (or at least until the Internet), modern social imaginaries must have been constructed by people who belong to multiple, sometimes overlapping, laterally and hierarchically organized groups, and who themselves possess complex social identities (Brewer and Gardner 1996; Brewer and Pierce 2005). How content actually get started and may even come to dominate in a culture or subculture (often because they come to be viewed as natural) is not immediately obvious. It seems most plausible that demes would be a mediating structure because their epistemic function is a local integration between facts and stories and relational and collective identities. A successful deme among macrodemes would presumably garner more influence and resources. Thus, for example, what may have begun as an “upstart religion” (and they all seem to start that way) or as a challenge to an existing power structure (e.g., housewives at Love Canal after the discovery of its use as a toxic waste dump by a chemical company) may have started in a task group that expands into or is absorbed by a functioning deme, a group of “insiders.” From this perspective, “society” is hardly the smooth textured construct that comes to mind in discussions of “individual and society.” Rather, in dense populations with multiple-role cultures, “society” has a lumpy texture, rather than the “smooth texture” of similarly self-interested individuals.
The essential point is that people do dwell within collectively shared landscapes (Ingold 2000). Dog fancier communities, professional communities, and national “communities” are social imaginaries that are held together by a variety of practices, including the telling and retelling of shared narratives that order experiences, beliefs, and social relations. Such uniquely human processes have an evolutionary history that is grounded in repeatedly assembled—and observable—bodies, tasks, and coordinated groups.
Other epistemic projects inhabit social imaginaries (Caporael 2007), including folk psychology, which Bruner (1990) describes as an “instrument of culture.” In his view mind is constituted by culture; it is not a naked product of biology or of physiological maturation as most theorists imply. Intentional states and experience are meaningful only through participation in the institutionalized meanings of culture. Folk psychology, Bruner writes, is “a set of more or less connected, more or less normative descriptions about how human beings ‘tick,’ what our own and other minds are like, what one can expect situated action to be like, what are possible modes of life, how one commits oneself to them, and so on. We learn our culture’s folk psychology early, learn it as we learn to use the very language we acquire and to conduct the interpersonal transactions required in communal life” (Bruner 1990, 35). Parents, friends, and older children scaffold the active developing infant into this domain, starting with the location of birth, who helps in the birth, how the baby is wrapped, when and how it is fed and by whom … and that is just the beginning. (Some years ago, I was once startled to see a group of newborns in a hospital nursery with pink bows taped to their heads. Without guessing or analysis, these were obviously the baby girls, and the pink bows were the maps to their futures.)
Folk psychology is major epistemic project under social imaginaries. It includes expectations about “how people work,” legitimate goals and practices, appropriate behavior for children, the division of labor by gender, and privileges of age. One question we could ask is, How is an epistemic project like folk psychology distributed among core configurations? As a blunt hammer of culture, as in the nursery, folk psychology is an elaboration of epistemic projects of demes or macrodemes. In some cases, modes of attunement seem relevant. The physical form and functioning of the body raises the possibility of sharing or “reading” other minds as well as translation across cultures (although fatal or perfidious misunderstandings are known to occur). The cross-cultural literature on expression and emotions (Ekman and Rosenberg 1997; Sauter et al. 2010) also suggests that shared transactions between bodily experience and psychological experience. Other connections of body and mind to folk psychology have also been illustrated by research on embodied cognition (Lakoff and Johnson 1980). Most striking are the ways that manipulating sensory factors, for example, holding warm coffee (Williams and Bargh 2008) or standing in close proximity (IJzerman and Semin 2010), leads to feelings of interpersonal warmth toward other people.
Other aspects of folk psychology bring task groups and paradigmatic projects to mind. One is simply routinized institutional behavior which has well-known scripts within a culture (Schank and Abelson 1977). In many cultures, including American culture, much of public life (e.g., eating at a restaurant or going to the dentist) is governed by routines that we often begin learning in childhood and, as adults, watch as familiar routines change over time, usually on a plank-by-plank basis rather than as an overhaul of the entire ship of the institution. A second type of relevant experience involves settings, activities and artifacts. If you see your partner standing at the refrigerator door staring at the contents in the middle of the night, you usually safely assume he or she is hungry. Again, epistemic projects concern coordination, not correctness. Psychology has a rich literature, including patterns of errors, on attribution of causes to dispositions rather than to perception–action dynamics in settings and situations that can cause and shape behavior (Dijksterhuis and Bargh 2001).
Scaffolding Generations
Early mother–infant interaction is a rich field for demonstrating reproductive overlap between generations as well as a meeting point between deeply entrenched and shallowly entrenched capacities. Infancy is like a fulcrum between generations, a recursive, evolutionary developmental relationship, well-represented in Shu-Chen Li’s (this volume) figure 15.1 depicting the dynamics of brain and cognitive development over different scales of time. Rather than looking at infant mental capacities as precursors to adulthood, the perspective is one where infants are adapted to the conditions of infancy. Neither blank slates nor adults-in-waiting, infants are born into a “community of other minds” (Nelson 2007) fully attuned, alert and ready to engage in the epistemic project of joining it.
The caregiver-infant dyad is the main setting in the evolution and ontogeny of infancy. It is deeply entrenched because many repeated assemblies “downstream” depend on early development. The microcoordination given as an example of a function of dyads in table 2.1 can refer to a wide range of more specific dyadic interactions where more than just mothers and infants are dynamically coupled in the production of coordination.
The newborn infant bears minimal resemblance to the adult that it will become. Its brain has one-third of the cortical surface area that it will have as an adult (Hill et al. 2010), and its visual acuity, contrast, and field of vision are considerably limited (Farzin, Rivera and Whitney 2010). Objectively, the newborn is strange—its form is out of proportion, it makes funny noises, it has a distinctive smell, and its movements are jerky and uncoordinated. Remarkably, the infant is not seen objectively, and least of all by its parents. This gap between the “objective” (or embodied) infant and the “subjective” (or enactive) infant merits research from embodied and evolutionary perspectives precisely because the embodied infant seems to lie beyond the bounds of common folk psychology. Caregivers attribute a subjective life to infants, simulating desires and intentions on behalf of the newborn without regard for its initial limitations.
Nursing is one of the first activities of the newborn infant. Babies suck in a burst–pause pattern that appears to be unique among mammals which continuously suck without pause (Sroufe and Waters 1976). The pause is something of a mystery: there is no physiological reason for pauses—the infant breaths and swallows as it sucks; there is no difference between bottle-fed and breast-fed infants in pausing, and although bursts of sucking get shorter as the infant gets full, the pause duration remains constant. There is no apparent function for pausing. Kaye and Wells (1980) undertook a naturalistic study of 52 mothers feeding their infants at two days and then two weeks old. Mothers explain the pauses by “mind reading”—infants are initially “involved” with nursing, but then the baby gets “lazy,” “tired,” or “stops paying attention” and stops sucking. So mother begins to jiggle the baby to get his or her attention or to get the baby “back to work.” However, the conditional probabilities calculated in the microanalysis comparing the bursts of sucking and pauses show that jiggling does not serve the mother’s intended goal. The infant does not start a burst of sucking until after the mother stops jiggling. There is no contingency between the initiation of jiggling and the newborn’s next round of sucking; not until the mother stops jiggling does the infant begin sucking again. Yet mothers seem unaware that stopping jiggling leads to the next burst of sucking or that they shorten their jiggling over the next two weeks. Kaye and Wells (1980; Kaye 1982) argue that the endogenous rhythms and reflexes of the infant provide a framework for the temporal structure of turn taking characteristic of dialogue. The mother fits her behavior into the infant’s rhythms.
Hendriks-Jansen (1996) extended this analysis and applies it to other instances where the infant’s rhythms, reflexes, and emerging attention is exquisitely attuned to evoking the parent’s simulation of the future infant mind. Many other animals show abilities for highly structured turn taking as in mating or display behavior, but these are highly fixed and cue bound. Human turn taking requires enormous flexibility over a range of circumstances. Not only language, but music, dance, and nonverbal expression (overt and subtle) are involved in the coordination of meaning and action. As a result of natural selection, the infant scaffolds the parent’s attribution of the baby’s own future intentionality. The burst-pause-mother-jiggle pattern plays no functional role in feeding. Its function is the initiation of a deeply entrenched component that serves as another cycle in the repeated assembly of the next generation of a theory of mind. The mother’s specific interpretation (lazy boy, getting him back on the job, etc.) lies outside the activity patterns of turn taking and constitutes the first steps that will bootstrap the infant into a cultural world with a variety of tools, scripts, and models (Shore 1996) that the infant will make his or her own.
Between six and twelve months, there is considerable mimicry between adults and infants (Jones 2007, 2009)—almost entirely on the part of adults, who are seven to ten times more likely to mimic infants than vice versa. As before, adults respond to the activity of the infant, inserting their own imitative behavior when the opportunity arises. As infants produce higher frequency and variability of behavior, mothers imitate them even more, and such games are often played with squeals of delight, maternal pride and pleasure. Yet as before, mothers believe that their infants are imitating them, even though the probability of matches between infant and mother behavior do not exceed chance expectations until around twelve months of age (Lewis and Rosenblum 1974).
This microanalysis approach to infant development appears to contradict the widespread agreement that infant imitation is innate and observable from birth (e.g., Meltzoff and Moore 1977). A critical review of the literature (Ray and Heyes 2011) indicates that newborns less than six weeks old do match tongue protrusion, but they do not imitate other actions. Moreover, newborns protrude their tongues to other stimuli including flashing lights or dangling toys, suggesting that a range of stimuli elicit tongue protrusion (Jones 2009). Jones (2007) also challenged the widespread view that imitation is innate in a longitudinal experiment with 162 infants tested at two-month intervals between the ages of six and twenty months. In this unique study, which controlled for spontaneous production of gestures, parents were guided in modeling eight behaviors previously claimed to be imitated in early infancy. Jones found that six-month-old infants failed to reproduce any of the gestures at appreciably above chance levels and that four of the eight gestures failed to even be observed at this age. Over successive age levels, infants come to imitate the gestures, beginning with two associated with sound (vocalizing “aah” and tapping the table at eight and twelve months old, respectively) and with the emergence of gestures occurring at different ages but following similar development trends. Notably, infants did not mimic tongue protrusions above spontaneous tongue protrusions until they were eighteen months old, which is consistent with the idea that above-chance matching of newborn tongue protrusion is an evolved activity pattern that scaffolds and stimulates adult attention. It drops out at six weeks and then appears again as “true imitation” at eighteen months.
Imitation does not appear as an innate whole; it is a product of interactive emergence (Hendriks-Jansen 1996). As we would expect from an evo–devo perspective, it seems that infants repeatedly assemble imitation, generation-to-generation, from different kinds of knowledge (e.g., the associations with sound; examination and experimentation with hands and other parts of the body) over a drawn-out course of development prompted, initially, just from endogenous activity such as tongue protrusion, mouth opening, and gaze, and later by smiles, mimicry, and motion, which is in turn elaborated, stimulated, elicited, and guided by adult imitation, attention, praise, and positive affect as adults simulate infant desires, intentions, and (later?) beliefs.
Conclusion
Every human alive today is materially connected to preceding humans through hundreds of millennia. Our children, like ourselves, our parents, and our parent’s parents have been held, nursed, and brought into a human community of rules, practices, and artifacts. Lacking armor, weapons, and means of escape observed in other creatures, humans share a bodily form and developmental trajectory that binds us to coordinated groups. Our bipedal form of human materiality is deeply entrenched, and at the same time, it is highly generative: we continue to hold, to nurse, to teach, and to create whether we make stone tools or space stations.
The “gene's-eye view” for the most part is blind to the organism level, hence, its mathematical generality. What is controversial is how the gaps are filled; sometimes it is impossible to distinguish the scientific and social uses of Darwinism (Maynard Smith 1987; cf. Gieryn 1999). Origin stories, epistemic projects that nicely coincide with our social imaginaries (Caporael 2007), are ubiquitous in human groups. More than one scientific evolutionary scenario can be shown to follow the narrative structure of folktales (Landau 1984; Kuper 1988). Consequently, such a “human evolutionary science” would be inadequate to its own stated aims. What is needed is a “human evolutionary studies” that invites not only a consilience of work in the sciences, arts, and humanities but also a reflexive critique.
In this chapter, I have focused on a conceptual framework, or landscape, based on coordination, recurrence, embodiment, and situated activity rather than on costs and benefits to the genes. Many models of human evolution are explanatory (Smith 2000). A trait such as warfare, the loss of estrous, pair bonding, or cooperation is chosen, its benefits described, and a scenario for its evolution is offered. As Lloyd and Feldman (2002) have pointed out, most evolutionary explanations are unable to shoulder the burdens of proof for a neo-Darwinian account. Accepting that limitation means finding another path. Less exciting, less definitive and strong in its conclusions, more modest in its aims, the repeated assembly of core configurations enables at least a principled conversation about recurrence at different levels of organization. In contrast to explanatory modes, the core configuration model is generative conceptual scaffolding offering a standpoint with some distance from folk psychology. Although space requirements limited the discussion, the core configuration model is constraint based (Caporael 1997; Caporael et al. 1989), requiring attention to research in related disciplines, including social psychology. Obviously, pursuit of this research requires a group. A good-sized macrodeme might be just right.
Notes
References