Figure 1.1. 1999 Geologic Time Scale. Reprinted by permission of the Geological Society of America. In order to read this chart in stratigraphic order, read the columns from bottom to top, starting at the bottom of the Precambrian column, adding the bottom of the Paleozoic column to the top of the Precambrian column, the bottom of the Mesozoic column to the top of the Paleozoic column, and the bottom of the Cenozoic column to the top of the Mesozoic column.
The vicinity of Cincinnati, in the Ohio River Valley of southwestern Ohio, including adjacent northern Kentucky and southeastern Indiana, is among the most fossil-rich regions in North America, if not the entire world. The profusion of fossils in the local limestone and shale attracted many pioneering geologists and paleontologists of the nineteenth century, and much fundamental work in American paleontology and stratigraphy was accomplished here. Hundreds of fossil species were first discovered and named from these rocks. Early geologists gave the entire series of strata exposed here the name “Cincinnatian,” and this name was applied to strata of similar age throughout North America. Cincinnatian fossils are displayed in museums all over the world. Researchers, students, and amateur fossil collectors regularly visit the Cincinnati region to collect fossils. Many of those who have grown up in the region are aware of the abundance of fossils, yet few appreciate the uniqueness of this richness and its broader significance to our understanding of the Earth’s past. The purpose of this book is to explore the richness of Cincinnatian fossils and the stories they tell about life over 450 million years ago, when shallow seas inundated North America and the site of Cincinnati was in the Southern Hemisphere.
Why are fossils so abundant in the rocks of Cincinnati’s hills? Beyond sheer abundance, what is their significance for our knowledge of the history of life, evolution, and ancient environments? There is no single answer to these questions, but rather several answers can be given which collectively reveal the significance of Cincinnatian fossils. These answers can be found under four categories: organic evolution, environment, preservation, and history.
Of the many prolific collecting grounds in the continental interior, none excels the Ohio river bluffs at Cincinnati, Ohio. Here the Upper Ordovician rocks are almost literally made of fossils; many are as perfectly preserved as fossils can be. The river banks, road cuts, and even the soil in the gardens are replete with fossils more common than pebbles. Almost every museum in the world has specimens from this locality.
William Lee Stokes 1960, 188–189
Organic Evolution
[The Ordovician radiation] represents one of the largest major turnovers in the history of life and marks the appearance of groups that came to dominate marine ecosystems for the next 250 million years.
Droser, Fortey, and Li 1996, 122
Fossils found in Cincinnati’s limestones and shales are the remains of animals that lived during an interval of Earth history called the Ordovician Period. The Ordovician is the second oldest period of the larger time interval known as the Paleozoic Era (Figure 1.1). The beginning of the Paleozoic Era (meaning “time of ancient animals”) is marked by the oldest rocks containing abundant fossils of multi-celled animals (metazoans). Radiometric dating of volcanic ash beds interbedded with these fossiliferous rocks places the beginning of the Paleozoic at about 543 million years ago. Similar methods date the beginning of the Ordovician Period at about 490 million years ago and its end at about 443 million years ago. The span of Ordovician time represented by the Cincinnatian strata amounted to less than 10 million years, and fell approximately during the latter part of the Ordovician, termed the Late Ordovician. In the Cincinnati region, a total thickness of over 250 meters (820 feet) of interbedded limestone and shale was deposited during the Late Ordovician, constituting the Cincinnatian and containing fossils throughout. Further discussion of the nature and subdivisions of Cincinnatian rocks, and estimates of their age, are the subject of chapter 4.
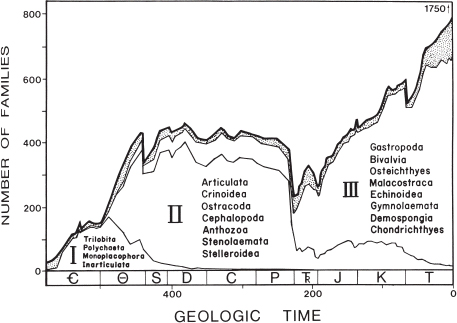
Figure 1.2. Diversity of marine fossil metazoan families through the Phanerozoic. The heavy uppermost curve depicts the sum of the three “evolutionary faunas,” each shaded differently, while the stippled portion below the total curve represents residual diversity not accounted for by the three component faunas. Taxa listed for each evolutionary fauna are those taxa that contribute most heavily to the diversity of that fauna. I = Cambrian Fauna, II = Paleozoic Fauna, and III = Modern Fauna. From Sepkoski (1981) and reprinted by permission of The Paleontological Society.
Professor Stig M. Bergström of the Ohio State University is among the world’s most knowledgeable and widely-traveled specialists on Ordovician fossils and stratigraphy. He indicated to us that “there is nothing that can be compared elsewhere in the world” to the diversity of shelly fossils in the Cincinnatian (Bergström, pers. comm.). Metazoan marine life first began to diversify during the so-called Cambrian explosion that marked the onset of the Paleozoic, but accelerated during the Cambrian and Ordovician Periods to reach a peak late in the Ordovician when the Cincinnatian strata were deposited. In fact the Ordovician Period is recognized as a unique time of evolutionary diversification, termed the Ordovician Radiation (Droser et al. 1996) or the Ordovician Biodiversification Event (Webby, Paris, Droser, and Percival 2004). The Ordovician marked a convergence of what Sepkoski (1981) called three “evolutionary faunas”: metazoan groups that first appeared during the Cambrian but persisted into the Ordovician (“Cambrian Fauna”), groups that began to diversify during the Ordovician (“Paleozoic Fauna”), and groups that first appeared in the Ordovician that diversified after the end of the Paleozoic (“Modern Fauna”) (Figure 1.2). At the end of the Ordovician there occurred a global mass extinction that eliminated species on a large scale. Thus the Cincinnatian time was significant in the history of life as a Golden Age of evolutionary diversification just before a major crisis of mass extinction. In many ways the Late Ordovician is comparable to the Late Cretaceous Period, another Golden Age preceding a crisis (Figure 1.2; Seilacher 1998). Few if any fossil species found in the Cincinnatian strata survived into the succeeding Silurian Period. Chapters 5–14 introduce each of the major groups of organisms found as fossils in the Cincinnatian.
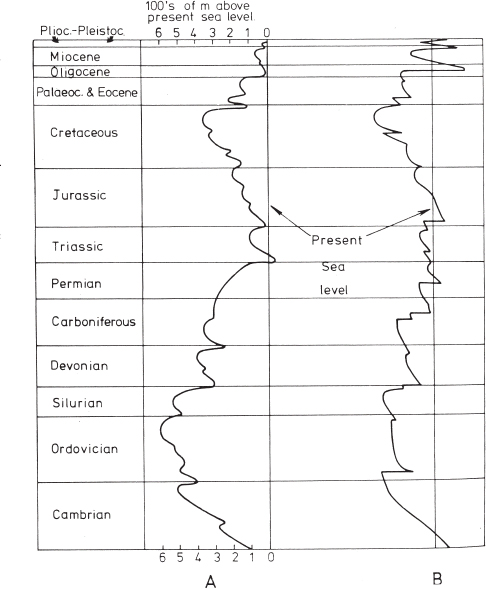
The environment of Late Ordovician time in the Cincinnati region contributed to the abundance and richness of fossils in several fundamental ways. Cincinnatian fossils and rocks bear profound testimony to the existence of widespread shallow seas (called epicontinental or epeiric seas) over most of the North American continent at this time (Plate 1). Using many sources of evidence, geologists have compiled a record of the rise and fall of sea level during the past half billion years of Earth history (Figure 1.3). The Late Ordovician was one of the times of maximum rise of sea level over the entire globe, rivaled only by the Late Cretaceous (according to the reconstruction by Hallam [1984]). The cause of this flooding has been attributed to high rates of sea floor spreading which swelled the mid-ocean ridges, displacing immense volumes of seawater from the deep ocean basins onto the continental plates. The Atlantic Ocean as we know it did not exist, but instead, a narrower ocean called the Iapetus Ocean separated North America from continental plates later to constitute Europe and Africa (Plate 1). The nearest landmasses to the Cincinnati region were the rising Appalachian mountain chain, about 300 miles to the east, and the low-lying Canadian Shield to the north. Just before and during the Late Ordovician, a phase of major tectonic (mountain-building) activity, the Taconic Orogeny, resulted in severe crustal deformation and uplift along the region bordering New York and New England. Islands were raised high above sea level as lofty and jagged mountain chains resembling the modern Alps or Himalayas. Weathering and erosion attacked these ranges, and rivers carried huge loads of fresh water, sediments, and nutrients into the shallow sea.
Great volumes of sediment, consisting of coarse gravels, sands, silts, and muds (termed siliciclastics) were deposited as river deltas and redistributed by oceanic currents near the coastline in the Appalachian Basin. The total thickness of the Late Ordovician strata in the Appalachian Basin in Virginia reaches about 1000 meters (over 3000 feet) whereas the same time interval is represented in the Cincinnati region by strata less than 300 meters (less than 1000 feet) thick (Figure 1.4; Kay 1951). Offshore, only the muddy components of this heavy sediment input remained suspended as clay particles, and were carried by currents to reach the Cincinnati area. These muds were thus imports to the region that eventually lithified (turned to stone) to form shales. In the Cincinnati area, shales are interbedded with limestones, which are composed of calcareous shells and skeletons of “native” marine invertebrates. In the western United States and Canada, the Late Ordovician contains mostly limestones secondarily converted to dolomites. Thus, the Cincinnati region represents an intermediate zone of mixed shales and limestones between the great thickness of siliciclastics to the east and pure limestones farther west. Both sediments intermingled in the Cincinnati region, producing a varied and patchy sea floor that was muddy in places and shelly in others. Such a variegated bottom environment offered more potential types of living spaces for bottom-dwelling organisms (the benthos), and provides a further reason why high diversity developed in the region. Because there was very little vegetation on land during the Late Ordovician, erosion may have carried a heavier load of dissolved inorganic nutrients into the sea. These nutrients may have acted as a fertilizer to stimulate the production of benthic biomass. In addition, climate, oceanographic conditions, and available food supply must have been crucial to support prolific marine life in the Cincinnatian sea; these factors are explored in detail in chapter 15.
Environment
We are accustomed to thinking of North America as terra firma, one of the large high and dry segments of the earth’s crust, and it is difficult for us to imagine a time in the past when our continent was so submerged beneath the sea that fish could have swum directly from the Atlantic Ocean to the Pacific Ocean, from Hudson Bay to the Gulf of Mexico. Yet such a time did exist 450 million years ago when the epeiric sea spread from Arctic to Gulf, from Atlantic to Pacific.*
Clark and Stearn 1960, 68
*Of course, true bony fish had not yet evolved in Late Ordovician time, and as we will see, Cincinnatian rocks contain no fossil evidence of the early, jawless fish that are known from the Late Ordovician elsewhere.
Figure 1.3. Global sea level curves for the Phanerozoic. A. Hallam curve, B. Vail et al. curve (1977). From Hallam (1984) and reprinted by permission of Annual Reviews. According to more recent studies (Miller et al. 2006), maximum rise of sea level in the Cretaceous was lower than these estimates, reaching 100 m ± 50 m above present sea level, but this does not contradict the evidence that Ordovician sea level was also very high and extensive over North America.
Preservation
When we look at rock layers as crowded with well-preserved fossils as those of the Cincinnatian, we tend to think we are looking at a complete picture of life on the Ordovician sea floor—a snapshot—in terms of both the diversity of species present and their abundance. Unfortunately, the correspondence between this fossil assemblage and the original living community from which it was derived is rarely that simple and direct. The fossil record provides a mere glimpse of ancient life, one that is heavily biased by many factors. In order to assess the impact of these factors on the quality of the fossil sample, paleontologists have devoted an entire subdiscipline, called taphonomy, to the investigation of processes affecting organic remains from death to ultimate fossilization. Taphonomy emphasizes the wide variation in the preservation potential of organisms. An appreciation of the significance of variable preservation can be gained by considering aspects of life, death, and post-mortem history that entered into the complex equation that determined the ultimate fossil record of the Ordovician sea.
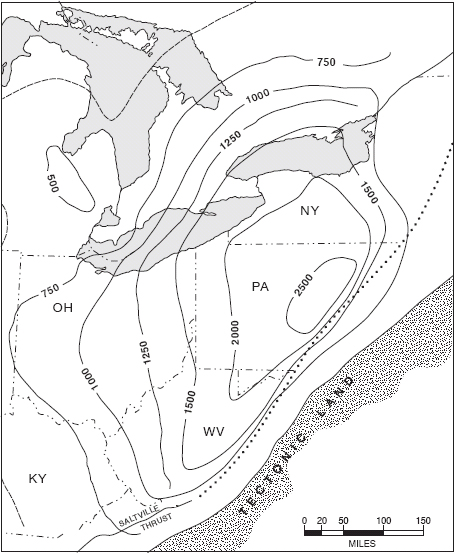
Figure 1.4. Thickness of Upper Ordovician strata in relation to the ancestral Appalachian Mountains (tectonic land) that was uplifted during the Late Ordovician Taconic Orogeny. Contours are lines of equal rock thickness (isopachs). From Kay (1951, figure 4) and reprinted by permission of the Geological Society of America.
The organic remains here are remarkably well preserved for so ancient a rock, especially those occurring in a compact argillaceous blue limestone, not unlike the lias of Europe. Its deposition appears to have gone on very tranquilly, as the Lingula has been met with in its natural and erect position, as if enclosed in mud when alive, or still standing on its peduncle.
Charles Lyell 1845, 49
Biological factors affecting preservation potential include presence of “hard parts,” their chemistry, mineralogy, and construction, and the mode of life of the organism. By far the most important requirement for fossilization is possession of mineralized hard parts such as shells or skeletons. Soft body parts including skin, muscle, hair, and internal organs almost always decay rapidly following death. Many common marine invertebrates like worms lack hard parts altogether or have only hardened jaw structures. In some marine environments, animal communities are dominated in numbers of species or individuals by such soft-bodied species with little or no fossilization potential. One of the best-known exceptions to the dominant preservation of hard parts is the Cambrian Burgess Shale of British Columbia, with its amazing wealth of soft-bodied worms, arthropods, and other invertebrates, along with shell-bearing forms (Gould 1989). In the Cincinnatian, there is virtually no preservation of soft-bodied species or soft parts of shell- or skeleton-bearing species. The only records known to us of soft-body preservation in the Cincinnatian are a worm described by Ulrich (1878) and the recent discovery of fossilized “tube feet” in a brittle star (Glass 2006). Our knowledge of the Cincinnatian biota is thus heavily biased in favor of species with hard parts, the shells and skeletons, complete or partial, known as body fossils. Fortunately, this is offset to some degree by evidence of the activity of soft-bodied species from trace fossils (burrows, tracks, and trails—the subject of chapter 14). However, it must be kept in mind that potentially great numbers of species in the biota will never be known because they left no fossil record whatsoever.
Shells and skeletons preserved in Cincinnatian strata are predominantly composed of calcium carbonate (CaCO3) in the mineral form calcite. Some shells of brachiopods (see chapter 8) and the microfossils known as conodonts (see chapter 13) are preserved as calcium phosphate. Despite the abundance of calcium carbonate in Cincinnatian fossils, not all shells having this chemical composition are equally well preserved. The reason for this is that some organisms form calcium carbonate shells or skeletons not as calcite but as a different mineral called aragonite. Aragonite, with a different crystallographic structure than calcite, becomes unstable in seawater after death of the organism and recrystallizes as calcite. In some cases this transformation occurs as a solid-state replacement of aragonite by calcite, altering the microstructure but retaining the macroscopic structure of a shell. Aragonitic shells can also be lost entirely by dissolution even before burial in sediment. In other cases, a shell may become buried, and as the internal soft parts decay, sediment seeps into the shells, replacing the soft parts and forming a perfect mold of the interior. After the aragonitic shell dissolves, the sediment infilling remains and can be lithified by calcitic cement. In this manner an internal mold or steinkern is formed which perfectly preserves the internal spaces of a shell, often molding features of the inner shell surface like muscle scars, even though the actual original aragonitic shell disappears. In other cases the shell may not be infilled, and once the shell dissolves, a void remains as an external mold of the outer surface of the shell, or the external mold can be infilled with sediment to form a cast. These are often the only ways a record of an aragonitic shell is preserved, and we have no way of gauging how many aragonitic shells dissolved leaving no trace whatsoever. Thus it is very difficult to estimate the original abundance of species forming aragonitic shells.
Even among species forming calcitic shells, preservation can be highly selective. Thinner, more delicate shells are more likely to be destroyed before they can be buried. In groups like trilobites (see chapter 11), the exoskeleton is composed of the protein chitin, with varying amounts of calcium carbonate. Juvenile, or newly molted, trilobites had weakly calcified exoskeletons, and were thus less preservable than more heavily calcified individuals. Thus, within a single species, preservational potential is unequal. Species having shells formed of one or two valves (snails, clams, or brachiopods) have a higher preservation potential than species with multi-parted skeletons such as crinoids or trilobites. Multi-parted skeletons are held together with connective tissue, which is susceptible to scavenging and decay, causing the skeleton to become disarticulated and scattered by currents. The consequence of all these variable factors of shell composition and structure is that all organisms producing a calcitic shell capable of preservation do not have an equal potential for actual preservation. Preservation is highly selective even among shells chemically and mineralogically stable enough to survive post mortem.
The mode of life of organisms determines preservation potential even before animals die. For aquatic species, bottom-dwellers (benthos) have a higher likelihood of preservation than swimming (nektonic) or floating (planktonic) species. Among the benthos, species that burrow into the sediment for a living (infauna) obviously have a much higher potential for preservation than do surface dwellers (epifauna). Among the epifauna, species living permanently attached to the bottom often have a higher potential for preservation than free-living, mobile species, simply because they are unable to escape sudden burial by sediment.
Fossilization is a rare event, not a process happening every day. Most animals that survive through old age and die of “natural causes” such as predation or disease will not become fossilized. Unburied carcasses are torn apart by predators and scavengers or destroyed by decay and exposure to the elements. Fossilization very often depends on a rare, catastrophic event that buries an entire assemblage of living organisms, much in way the eruption of Mt. Vesuvius buried Pompeii in ad 79, preserving incredible details of Roman life. Thus, processes of mortality are of fundamental importance in determining how organisms are preserved. When we see a fossil, the first question should be: “What happened?” The answer may tell us more about the nature of rare events, such as storms, earthquakes, or volcanic eruptions than about day-to-day processes.
The paleoecologist must never forget that he is studying not the living inhabitants of the village but only the bodies in the churchyard, and then only after many visits by grave robbers.
Derek V. Ager 1963, 184
In the Cincinnatian, the best-preserved fossils, such as complete trilobites or crinoids, probably resulted from sudden burial of a sea floor population by muddy sediment. Great storms are capable of shifting masses of sediment around on the sea floor or stirring it into suspension, only to settle out as a blanket over the bottom when the storm subsides (see chapter 4). Organisms were smothered by these events and protected from the normal cycle of scavenging, decay, and destruction. These cases offer the best opportunity to see a snapshot of Ordovician marine life. But even here we should be cautious, because such smothering events can preserve not only organisms living at the time, but also remains long dead and accumulated over time. Indeed, many highly fossiliferous limestone beds in the Cincinnatian represent long-term (time-averaged) accumulations of shelly material along with better-preserved specimens that were buried alive in an instantaneous event. With care, these components can be recognized, so that we can assess what species made up the life assemblage. The death assemblage of remains already dead at the time of burial is also informative, because, like a graveyard, it can record multiple generations and occurrence of rare species. Table 1 lists some of the most useful characteristics to look for in distinguishing fossils buried while living from those accumulated gradually as dead remains.
Table 1. Characteristics of Life Assemblages and Death Assemblages
|
Life assemblage |
Death assemblage |
Articulation |
good |
disarticulated |
Breakage |
rare |
common |
Abrasion |
rare |
common |
Preserved in life position |
maybe |
not often |
Size-sorting |
uncommon |
possible |
History
. . . our search for a mechanism forces us to range far beyond the Cincinnati region and consider the geologic history of much of eastern North America, especially the continental collisions . . . referred to as the Taconic, Acadian, and Alleghenian orogenies.
Paul E. Potter 1996, 71
If, in light of the foregoing discussion, the reader is not fully convinced of the extreme rarity of fossilization and the uniqueness of the fossil richness of the Cincinnatian strata, the following section should provide additional food for thought. Subsequent to life and death during the Late Ordovician Period, 450 million years ago, the remains of marine animals were buried in sediment. Worldwide, a great many fossils from very ancient Paleozoic strata are poorly preserved because they have suffered greatly from the ravages of time—chemical and physical modifications occurring during and after burial. These changes, technically known as diagenesis, include dissolution of original shell material, with or without replacement by other minerals, pervasive recrystallization of the rock, with partial or complete obliteration of fossil contents, or crushing and deformation of fossils during compaction of the enclosing sediments. Fossils that survive diagenetic damage at low temperatures may be later destroyed by actual metamorphism, in which deeply buried strata are heated, recrystallized, and deformed to varying degrees. Metamorphic processes transform primary sedimentary rocks such as shales into slates, limestones into marble, and sandstones into quartzite, accompanied by nearly total obliteration of fossils and other primary features of the sediment. Metamorphism is associated with deep burial by overlying strata, or mountain-building processes of tectonics, including folding, faulting, shearing, and volcanic activity.
The pristine quality of many Cincinnatian fossils is clear evidence that they have undergone very little diagenetic alteration and no metamorphic change over their long burial since the Ordovician. How could these fossils have survived with so little alteration over such a vast span of time? The answer is “location-location-location” and the history of the Cincinnati Arch.
Because the Cincinnati region is located inland from the continental margins, it is far distant from regions that have undergone intense deformation and metamorphism over the course of time. The closest deformed strata of the Appalachian tectonic zone lie about 200 miles to the southeast (Pine Mountain, Tennessee), and metamorphosed rocks are even farther (the Great Smokies and Blue Ridge of Tennessee). Furthermore, the Upper Ordovician strata of the Cincinnati region were never buried deeply beneath younger sedimentary strata. At the close of Ordovician time in the region, there is evidence that the seas became very shallow, the sea floor perhaps exposed subaerially (above sea level), producing a gap in the stratal record known as an unconformity. Subsequent sedimentation from the Silurian through Pennsylvanian Periods (a span of 150 million years) was again submarine, but mostly of very shallow water origins. After the Pennsylvanian Period (about 290 million years ago), there is no record of further marine sedimentation in all of Ohio. Altogether, the total thickness of strata deposited over the Ordovician may have been 300–600 meters (1000–2000 feet) at most (Potter 2007). Indications of shallowing, retreating seas suggest that the Cincinnati region was undergoing regional uplift, because marine deposits are thicker to the east and west than those closer to Cincinnati. On the continental scale as well, epicontinental seas retreated by the end of the Paleozoic, only to return during the Mesozoic, but farther west than Ohio.
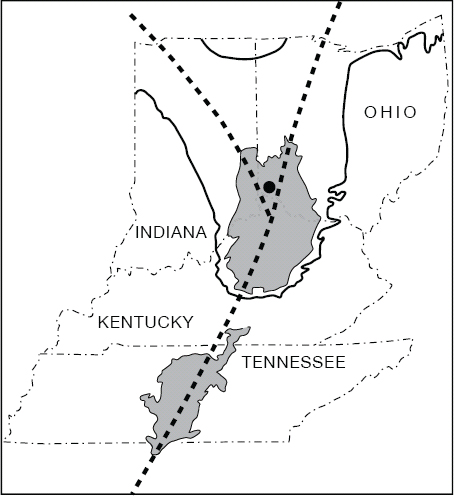
Figure 1.5. Axis of the Cincinnati Arch and its branches, the Findlay Arch (through Ohio) and the Kankakee Arch (through Indiana). Shaded areas depict outcrop of Ordovician bedrock; heavy lines indicate Silurian-Devonian contact that defines the Findlay and Kankakee branches.
The regional uplift that affected the Cincinnati region was part of a broad zone called the Cincinnati Arch, trending north-south and splitting just north of Cincinnati into the northeasterly-trending Findlay Arch and the northwesterly-trending Kankakee Arch (Figure 1.5). The term “arch” implies an upwarping of the Earth’s crust, but anyone who has noticed the strata of limestone and shale exposed in roadcuts around Cincinnati has seen essentially horizontal layers. It is only when we travel eastward or westward from Cincinnati that we encounter geologically younger strata overlying the Ordovician strata along the axis of the Arch, and the character of the Arch as a very gentle, broad upwarping becomes apparent. The tilt or dip of the strata across the Arch is usually less than one degree, or four to seven feet per mile, in Hamilton County (Potter 1996). Over a distance of about 80 km (50 miles) east and west of Cincinnati, a cross-section through the bedrock shows the structure of the Arch clearly (Figure 1.6). Even though the Cincinnati region is distant from the deformed rocks of the Appalachian mountain belt, the Cincinnati Arch may have resulted from the same large-scale tectonic processes that uplifted the Appalachians, beginning in Ordovician time. The reader is referred to Potter (2007) for further discussion of the origin of the Cincinnati Arch.
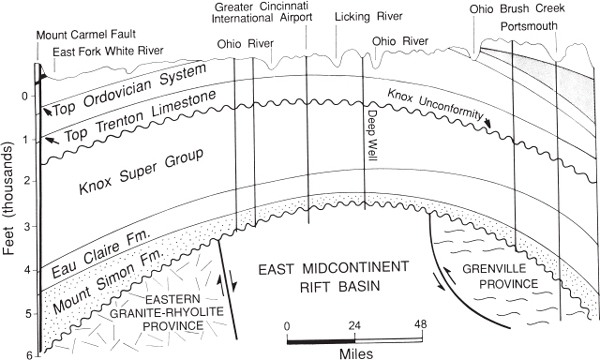
Figure 1.6. Diagrammatic east-west cross section from near Bedford in south-central Indiana across the Cincinnati Arch to near Portsmouth, Ohio. From Potter (1996) and reprinted by permission of the Kentucky Geological Survey.
Because uplift continued along the axis of the Cincinnati Arch, it was continually eroded, stripping away strata lying above the Ordovician, preventing their deep burial. During the past two million years, the glaciers of the Pleistocene Epoch covered most of Ohio and the ice sheet scraped away remaining overburden or washed it away as the ice melted, completing the exposure of Ordovician strata at the surface. By these processes, begun virtually at the same time the Ordovician seas covered the Cincinnati region, sediments and fossils deposited then were never deeply buried and deformed, and became exposed over a broad belt through the region. The significant consequence is that the entire Cincinnati Arch region has one of the most extensive surface exposures of Upper Ordovician strata in North America, if not the entire world. Natural exposures (outcrops) in streambeds and the sides of valleys (Figure 1.7A) and human-made exposures, mainly as roadcuts (Figures 1.7B, 1.8) and quarries, provide access to the Ordovician bedrock throughout the region. In the Cincinnati Arch region we have a truly unique window to the past—easy access to ancient strata and fossils that elsewhere lie buried under thousands of meters of rock.
Figure 1.7. A. Natural exposure of Cincinnatian strata in the bed of Stonelick Creek, Clermont County, Ohio. Streambed outcrops have wide surfaces of fossiliferous beds (bedding planes) that provide information about fossil distribution and orientation. Here, the fossil-rich Bellevue Limestone is exposed and examined by participants in a 1981 Geological Society of America field trip. B. One of the most extensive roadcut exposures of Cincinnatian strata, 1.3 km (0.8 miles) in length and about 75 m (250 ft) high, along Kentucky Route 3071, leading to the Ohio River, west of Maysville, Mason County, Kentucky. The lowest strata exposed are the lower Cincinnatian (Edenian Stage) Kope Formation, and the highest strata are the Bellevue Limestone (Maysvillian Stage). Photo by Paul E. Potter.
Figure 1.8. Trammel Fossil Park, Sharonville, Hamilton County, Ohio. This is a ten acre hillslope where construction exposed four fossiliferous formations: the Fairview Formation, Miamitown Shale, Bellevue Limestone, and Corryville Formation. The developer R. L. Trammel donated the site to the City of Sharonville as an educational, geological park where visitors can learn about the Ordovician geology and paleontology. The park has easy access, parking space, and includes interpretive signage and a protected, in situ fossil shell pavement. Because of the abundance of fossils, surface collecting of small specimens is permitted.
The foregoing overview shows that the abundance of fossils in the Ordovician rocks of the Cincinnati region is the result of many interacting factors. Because of this unique and fortunate combination of factors, the Cincinnati region became one of the earliest centers of intense interest and study of fossils in North America. Scores of Ordovician fossils were first discovered and described from this region, and many practices and concepts of paleontology and geology originated from research on Cincinnatian fossils and rocks. Because the Cincinnati region was one of the birthplaces of modern geological science, we will explore the early history of study of the fossils and rocks here in the following chapter.