six
NO BRAIN IS AN ISLAND
“LET US MAKE MANKIND IN OUR IMAGE, AFTER OUR likeness,” says the God of Genesis, “and let them have dominion over the fish of the sea, and over the fowl of the air, and over the cattle, and over all the earth, and over every creeping thing that creepeth upon the earth” (Genesis 1:26). Except for the Lord himself, humanity reigns supreme over the planet, destined to shape rather than be shaped, command rather than be commanded, and consume rather than be consumed. A similar belief in the hegemony of Homo sapiens is shared today by most people of the world, regardless of their religion or lifestyle. Even committed vegans or Buddhists, who ostensibly reject the subjugation of animals, participate in a global civilization that progressively subdues the planet through urbanization, transportation, agriculture, and industry. We are, as many now say, living in the age of the Anthropocene.
It is an understatement to assert that the triumph of humankind over nature could not have been possible without our brains. We each seem to control our immediate environments through our ability to perform voluntary actions, a capability once associated with the metaphysical soul or self but more recently swallowed up by the central nervous system. “Increasingly it is recognized that the self does not need to be outside the body,” wrote renowned neuroscientist Peter Milner in his 1999 book The Autonomous Brain. “It may more conveniently be thought of as a complex neural mechanism, often referred to as the brain’s executive system.” University College London cognitive scientist Patrick Haggard observes likewise that “modern neuroscience is shifting towards a view of voluntary action being based on specific brain processes.”
Part of the blame for this shift goes to the famous experiments of people like Benjamin Libet, who showed in the 1980s that electrical signals from the brain can be used to predict a person’s apparently voluntary movements even before the person has made a conscious decision to act. In effect, the brain “knows” what we will do before we ourselves can catch on. This suggests that the brain is boss, potentially determining our actions and having our ideas more or less on its own. As neuroscientist David Eagleman puts it, the brain is “the mission control center that drives the whole operation, gathering dispatches through small portals in the armored bunker of the skull.”
But the brain-in-a-bunker metaphor presents a paradoxical view of the interaction between our brains and their environments. In addition to understating the connections between brain and body, this picture presents the wider environment as little more than a passive contributor to brain function. The environment supplies information to the bastioned brain, which then reviews the data and decides how to respond. The brain is the commander, and the originator of all autonomous action. The thickness of the bunker’s armor and the smallness of its portals imply that the commanding officer is well insulated from the action in the field, left to ponder intelligence and plan strategy far away from the actual fighting. But in the stark distinction between what goes on outside versus inside the brain, and in the unbalanced power relationship between them, we can see another example of the scientific dualism we have considered in previous chapters of this book.
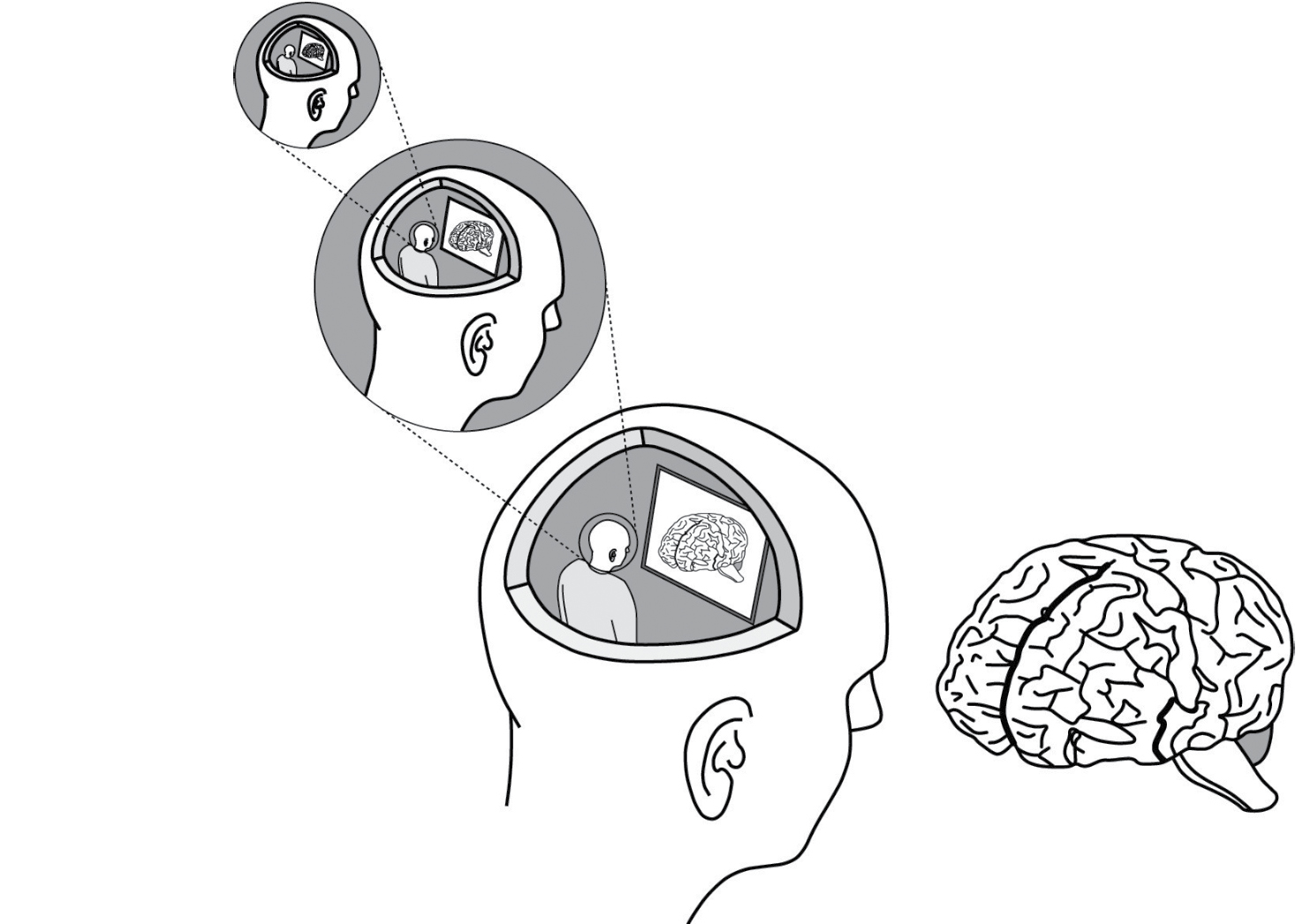
The paradox here lies in the difficulty of identifying a place where dispatches from the outside turn into decisions. Can we conceive of a handoff point—either a distinct locus or perhaps a more distributed “complex neural mechanism”—where the deterministic response to environmental input ends and the brain’s cognitive control kicks in? Some philosophers have argued that having such a place is equivalent to supposing that a little person—a homunculus—sits in the brain, receiving all of the input and figuring out how to respond. The 2015 Disney hit Inside Out spins this cartoonish scenario into a literal cartoon, in which five personified emotions named Joy, Sadness, Fear, Anger, and Disgust vie to influence a young girl’s actions by twiddling knobs and levers on a control panel inside her head. But how do Joy and her colleagues accomplish this? Doesn’t there need to be another set of homunculi inside each of them, converting their input to output? This notion applies recursively, like the never-ending cascade of reflections you see when you stand between two opposing mirrors in a dressing room (see Figure 10). The resulting contradiction is known as “Ryle’s regress,” after the philosopher Gilbert Ryle, who discussed the problem in his 1949 magnum opus, The Concept of Mind.
An obvious way to avoid Ryle’s regress is to give up on the idea that the brain can ever operate independently of the world around us. In this view, the decisive influence of the environment reaches much more deeply into the brain, all the way to our decisions and actions themselves. When an apple drops from a tree, when the winter snows melt into the rivulets of spring, or when a motorcycle skids off the highway into a ditch, it is the laws of physics and the contours of the surroundings that determine their paths. Perhaps the brain is like a falling apple, governed by the forces of nature even as it strikes Isaac Newton on the head. In his famous essay On the Freedom of the Will, the early nineteenth-century philosopher Arthur Schopenhauer insisted that “like the actions of every being in nature, [human doings] must be subject to the law of causality in all its strictness.” If this were true, the human brain would simply be an element in the causal chain, a bead vibrating passively on the string of life, rather than a hand that shakes the string itself. Nature would have dominion over the brain, rather than the other way around.
A difference between our time and Schopenhauer’s is that today we have access to a wealth of experimental data about the long arm of the environment at work in our brains and in our behavior. In this chapter, we will survey some of this evidence and see that the causal role of the environment is more than just a theoretical abstraction. The relationship between brain and environment goes much further than the truisms that people are products of their time and place, that both nature and nurture are important, or that learning and memory arise from experience. When the boundary between the brain and environment is blurred, every thought and action, even at the moment it occurs, becomes a consequence of influences in the wider world. By examining these connections, we will again combat the mystique of the brain as controller and appreciate the extent to which our brains are natural entities subject to the universal laws of cause and effect.
A famous allegory of the interaction between the environment and the mind is the statue of the three wise monkeys. The oldest example is said to be a seventeenth-century carving over a doorway at the Tosho-gu shrine, near the burial place of the great shogun Tokugawa Ieyasu in Nikko, Japan. One monkey covers his eyes with his hands, another clutches his ears, and the third muzzles his mouth, representing the timeless injunction to “see no evil, hear no evil, and speak no evil.” Monkeys are famous for getting around, and these three are no exception. Figurines of the monkeys are now among the world’s most globalized forms of kitsch, sold across six continents, with a small but dedicated society of collectors that runs a website and meets annually. A statuette of the monkeys was one of Mahatma Gandhi’s few possessions, symbolizing his strict moral code. Elements of the Italian mafia took the monkeys to represent their own code, the omerta or vow of silence. Some depictions humorously promote chastity by adding a fourth “do no evil” monkey who guards his private parts. The animals’ input and output are both covered in this way. By juxtaposing seeing and hearing with saying and doing, the monkeys thus teach the lesson that behavior is inseparable from external influences that begin with the senses.
Our principal sensory systems—sight, hearing, touch, taste, and smell—offer the clearest routes by which the environment around us influences our thoughts and actions. It is telling that another Eastern allegory, an ancient Hindu version of Plato’s famous mind-chariot analogy, presents the senses as five horses pulling a vehicle symbolizing the body. Almost all of the things we learn are ingested via our sensory organs, but the senses provide much more than fodder for our education. Sensory systems allow environmental stimuli to shape our thoughts and actions in far more immediate ways. On a continuous basis, we are inundated by the steady stream of input from our sensory organs to the brain. Like a drink from the proverbial fire hose, the force of this information flow is both overwhelming and unstoppable. Our senses remain active even in sleep and under anesthesia, relaying signals into the brain whether we are consciously aware of them or not. By delving into sensory biology, we can begin to appreciate how difficult it is for our brains to resist the influence of external stimuli.
The best studied and probably most influential of our senses is sight. Vision researchers have spent many decades studying how light striking the photosensitive part of the eye, the retina, is detected by the so-called rod and cone photoreceptor cells and subsequently processed into neural impulses (action potentials) that zip toward the brain via the optic nerve. By measuring electrical signals from the retinal output neurons called ganglion cells, neurophysiologist Horace Barlow discovered in the 1970s that each individual quantum of light—each photon—can give rise to an average of one to three action potentials. Even in complete darkness, Barlow found that some ganglion cells can fire as many as twenty of these spikes per second; this activity is essentially noise in the system, but it still floods the brain’s inbox.
Caltech professor Markus Meister devised a way to analyze the first steps in vision by isolating live animal retinae and spreading them like tiny blankets onto beds of recording electrodes, permitting measurements from dozens of ganglion cells simultaneously. Using this technique, Meister and other neuroscientists have observed how retinas adapt rapidly to huge changes in image intensity and contrast, ensuring that the deluge of visual information to the brain is never stanched. In one study, researchers estimated that total data transmission from the human eye to the brain is roughly equivalent to a computer’s internet connection, moving about a megabyte of visual input (four million spikes) each second over neural wires formed by the axons of a million ganglion cells per retina.
The nonvisual senses are also prolific sources of input to our brains. The organ of Corti in the inner ear converts sound waves into neural impulses—it is the ear’s equivalent to the retina. Most of the auditory neurons that emerge from the organ of Corti fire action potentials at a rate of more than fifty spikes per second, even for low sound levels. With about thirty thousand auditory neurons per ear in humans, the total number of action potentials reaching the brain from the ears each second extends well into the millions. An enormous amount of input also comes from the body’s largest sensory organ, the skin. Normal skin contains four types of touch or pressure-sensitive receptor cells, two types of temperature-sensitive receptors, and two types of pain receptors. Most of these receptors connect directly to the spinal cord and synapse with neurons that project from there into the brain. Some touch receptors reach densities of over two thousand per square centimeter, and there are seventeen thousand of these cells in the hand alone. Two specialized types of skin are the surfaces of the tongue and the inner lining of the nose, which contain receptors for taste and smell, respectively. The olfactory receptor neurons are by far the more numerous; over ten million of them connect the nose directly to the brain. This means that despite low average firing rates of about three spikes per second, these cells still collectively transmit more electrical impulses to the brain than either the eyes or the ears do.
We see that the volume of sensory information converging on the brain comprises tens of millions of action potentials every second, reflecting the permanent and powerful connection our brains have to our surroundings. To appreciate the magnitude of this quantity, consider again that the input from a single eye to the brain is comparable to the data transmitted over an active internet connection. If that is so, then the contributions from all our senses combined is probably worth more than ten standard internet connections, bringing in around ten megabytes of data per second over millions of neural fibers. This much data directed at a typical home computer system today could easily be enough to saturate it; hackers sometimes use exactly this approach to target and overwhelm internet sites in what is known as a denial of service attack. By analogy, our sensory environments seem to be waging a continuous denial of service attack on our brains.
Interestingly, as assessed in terms of the sheer number of action potentials, the amount of sensory input coming into the brain is also comparable to the brain’s total output—the constant signaling that passes down from the brain to the rest of the body, initiating movement and regulating muscle tone. Most of the brain’s motor output is relayed by the so-called pyramidal tracts, which consist of over one million axons firing at average frequencies of around ten to twenty spikes per second, again resulting in tens of millions of spikes per second in aggregate. From an outside observer’s perspective, the brain could be seen as a somewhat convoluted mechanism for converting tens of millions of input signals per second into a roughly equal number of output signals, like a television set that converts input from your cable or antenna into moving pictures you can watch.
How do all of the incoming spikes affect the brain itself? Because the brain has evolved to accept this input, the sensory blitz does not really amount to an attack. The brain is not incapacitated—but it is changed. For sensory systems other than smell, the brain’s port of entry is a structure called the thalamus; olfactory signals proceed through a region called the olfactory bulb instead. These areas in turn connect to areas of the cerebral cortex such as the primary visual cortex (abbreviated V1), in the brain’s occipital lobe, or the primary auditory cortex, in the temporal lobe (see Figure 7). The influence of sensory input is felt well beyond these parts, however. More than 40 percent of the cortex is thought to be dedicated to sensory processing. In the visual system, which is the most extensive sensory modality in humans, information spreads from V1 into two sets of brain regions that each pick out different features of every stimulus. In the so-called dorsal stream, areas running along the top of the occipital and parietal lobes distinguish visual stimuli on the basis of gross properties such as their location or motion in space; the ventral stream, which runs along the base of the brain’s occipital and temporal lobes, specializes in more fine-grained analysis, such as the recognition of specific objects or faces. Similar streams of hierarchical processing regions handle sounds, smells, tastes, and touch.
Like gossip spreading among far-flung friends or family, the brain’s incoming sensory signals eventually reach almost everywhere. Even the sophisticated high-order sensory regions maintain responses to extremely simple stimuli. Images of flashing lines, for instance, produce neural responses throughout the entire dorsal and ventral visual processing streams. More surprisingly, like the medieval barbers who also pulled teeth, brain regions that specialize in processing one type of stimulus can actually respond to other stimuli as well. For example, researchers have shown that neural signals in the visual cortex can also indicate auditory stimuli; other studies have shown responses to both visual and tactile stimuli in the auditory cortex. Brain areas known for nonsensory roles also respond to simple sensory stimuli. Parts of the frontal cortex, seat of the brain’s “executive functions,” are among areas activated by basic visual or auditory input. Visual responses in frontal regions can be observed even under anesthesia, showing that sensory stimuli can reach far into our brains even if we are not aware of them. A remarkable phenomenon first noted by neuroscientist Mark Raichle in the early 2000s is that a constellation of disparate brain regions is also consistently deactivated by many stimuli—in other words, sensory input appears to decrease the level of neural activity in these regions. The deactivated areas cover a sizeable fraction of the cerebral cortex and consist mostly of territory outside the commonly recognized sensory or motor processing systems. These regions have been termed the brain’s default mode network because they appear to be most active when nothing noteworthy is going on.
Even subtle environmental influences can significantly affect what goes on in the brain. Most neurobiological studies of sensory responses are anything but subtle; they are performed using strong, short-lived stimuli designed to evoke brief episodes of brain activation. For instance, a visual response might be examined by measuring neural spiking or fMRI changes while a computer screen alternates every couple of seconds between a boring gray field and a bright red-and-green checkerboard. To study brain dynamics without such obtrusive stimuli, researchers perform a very different kind of experiment, in which they monitor a subject’s brain under constant conditions, over many minutes, while he or she is simply lying passively in a scanner (and encouraged not to fall asleep!). The resulting resting state data generally display very slight fluctuations in the neuroimaging signals at each point in the brain. If you watch a crowd at a sporting event, you can probably figure out which groups of people are rooting for the same team based on who cheers or boos together, even if you have no idea what is going on in the game itself. Using similar logic, researchers try to discern which brain regions “go together” by looking for pixels that increase or decrease in intensity at the same time. Such correlations are thought to reflect neural activity in distinct brain networks and are referred to as resting state functional connectivity.
Resting state imaging studies show that changes in brain activity occur during continuous visual stimulation of the sort that impinges on us all the time. Neurologist Maurizio Corbetta and his colleagues collected fMRI and MEG data from subjects who passively watched either a blank video screen or clips from a spaghetti western meant to simulate naturalistic sensory experiences. Analysis showed that functional connectivity patterns differed significantly between the two conditions. During the movie segments, the correlations among neuroimaging signals were reduced across multiple brain-wide networks, revealing how ongoing sensory experiences perturb dynamics over disparate brain areas. Importantly, stimuli much less exciting than Clint Eastwood films also alter brain activity. Tamara Vanderwal of Yale Medical School presented subjects with a video featuring abstract, continuously changing shapes—images you might see on a computer screen saver—and tested its influence on resting state fMRI fluctuations. Despite their thoroughly bland content, Vanderwal’s videos nevertheless perturbed functional connectivity in brain networks associated with vision, attention, and motor control. Another study, led by researchers at MIT, showed that meaningless acoustic noise can also affect functional connectivity patterns. The background noise affected regions in Raichle’s default mode network, demonstrating again that seemingly trivial sensory factors can perturb gross signatures of brain function.
The science of sensory processing thus helps us appreciate why the three wise monkeys might be worried about what comes in through their eyes and ears. The sensory organs conduct a ceaseless torrent of millions of neural impulses into our brains each second. The brain has no barrier against this flood of environmental input. Even the most banal sensory signals seem to worm their way into the most exclusive tiers of the cerebral cortex—regions such as the frontal lobe, which evolved in comparatively recent history and distinguishes both monkeys and humans from most other mammals. This does not yet prove, however, that signals from the environment exert control over us. When we see sensory influences permeating the deepest reaches of the brain, we may simply be witnessing how information reaches the proverbial homunculus. Perhaps a homunculus-like neural mechanism in each of our brains preserves our autonomy even in the face of the sensory onslaught. To address this possibility, we must examine the extent to which our behavior itself is determined by stimuli from the surroundings.
The writer Albert Camus saw that people are more slaves than masters of their environment. In his novel The Stranger, Mother Nature drives Camus’s antihero Meursault so far as to commit murder. In the book’s climactic scene, Meursault encounters an adversary on a beach in Algiers and shoots him dead. “All I could feel were the cymbals of sunlight crashing on my forehead and, indistinctly, the dazzling spear flying up from the knife in front of me,” the protagonist narrates. “The scorching blade slashed at my eyelashes and stabbed at my stinging eyes. That’s when everything began to reel. The sea carried up a thick, fiery breath. It seemed to me as if the sky split open from one end to the other to rain down fire. My whole being tensed and I squeezed my hand around the revolver.” Meursault comes to the fight armed with a gun and primed for violence by earlier events in the book. But at the moment of truth, it is the blazing sun and roaring waves that stimulate him to pull the trigger, rather than hot blood or malicious forethought originating in his brain. Through Camus’s prose, we witness a man whose mind is not his own. “Meursault’s crime seems utterly will-less, undetermined, and unfree,” writes Camus scholar Matthew Bowker.
One might say that the heat made Meursault do it. In this respect, the fictional Frenchman in Algiers has something in common with real-life police officers in Amsterdam. In a 1994 study, a group of psychologists monitored Dutch police as they performed a training exercise in a room with variable temperature. The officers demonstrated increasing signs of hostility and belligerence when the room got hotter. Most strikingly, they showed a 50 percent greater tendency to shoot at mock offenders when the temperature was 81 degrees Fahrenheit than when it was 70 degrees. This was not a fluke finding. In an ambitious survey of sixty different studies linking climate to various forms of conflict, researchers led by Solomon Hsiang at Princeton presented evidence that higher temperature leads to increased hostility and violence across a huge range of geographic settings and time scales. Ten cases revealed correlations between temperature and violent crime or domestic disputes in particular. In one instance, the number of assaults in the city of Minneapolis was found to vary on an hourly basis with ambient temperature, even when time of day and other potential biases were taken into account. This points toward a biological basis for some of the temperature dependence, rather than to societal factors like the density of people on the street or fluctuations in the local economy. Backing this up, brain imaging studies have shown temperature-dependent changes in functional connectivity that could relate to effects on behavior. “Although the physiological mechanism linking temperature to aggression remains unknown,” wrote Hsiang and his coauthors, “the causal association appears robust across a variety of contexts.”
The relationship between temperature and aggression has two features that illustrate the directness with which the sensory environment can govern behavior. First, it is a relationship few of us are consciously aware of and that we certainly do not have a conscious say in. This places the effect of temperature on aggression firmly outside our control, and limits the extent to which cognitive processes can intervene. Second, unlike responses to many artificial environmental stimuli, such as traffic lights or television programs, the behavioral effects of temperature cannot easily be explained by learning. Even untrained laboratory mice show increased aggression as a function of temperature between about 65 and 90 degrees Fahrenheit, as indicated by their tendency to bite each other when caged together in thermostat-regulated rooms. This suggests that temperature-dependent aggression is more or less hardwired, and it emphasizes our lack of freedom to acquire or shed this environmental sensitivity. One can imagine a chain of events in which temperature changes drive receptors in our skin to stimulate alterations in brain activity and neurochemistry, resulting in increased probability of hostile or violent actions, all without any semblance of control by us or our brains.
Another environmental factor that is hardwired to human behavior is light. Appreciation of this phenomenon owes much to the work of a psychiatrist named Norman Rosenthal. Rosenthal moved to the United States from South Africa in 1976 to continue his medical studies. Migrating from the agreeable climate of Johannesburg, he found the more rugged weather of New York hard to adjust to; the long nights of winter were a particular drag. Each time the season hit, Rosenthal felt his energy sapped and his productivity declining. “My wife had it much worse than me,” he recalled. “She was virtually bed-ridden for some of that time.” Unlike other victims of the “winter blues,” however, Rosenthal’s medical background put him in a position to do something about it. He became interested in circadian rhythms, the cycles of fatigue, hunger, and other biological processes that wax and wane each day, and he joined a research group studying them at the US National Institutes of Health. One day Rosenthal and his colleagues came across a manic-depressive patient who had carefully documented his mood swings and seemed convinced that they were connected to seasonal fluctuations in the amount of light available each day. “Let’s give him more light,” suggested Rosenthal’s colleague Alfred Lewy. Sure enough, supplementing the patient’s daylight hours with exposure to bright fluorescent lighting reversed the depressions he experienced during the winter months. This finding was repeated in several larger studies and led to the identification of a medical condition called seasonal affective disorder (SAD), a pathology which affects millions of people worldwide and is often treated using light therapy.
Ambient light levels control both mood and circadian rhythms using a dedicated visual sensory path that again evades the potential for conscious control. A special set of retinal ganglion cells in the eye responds directly to blue light, partially bypassing the normal pathway for light sensation via rod and cone photoreceptors. The special ganglion cells are connected to a brain region called the suprachiasmatic nucleus (SCN), so named because it sits over the spot where the optic nerves from the left and right eyes cross in the shape of a Greek letter chi (χ), at the base of the brain. The retinal input influences genes in the SCN to turn on and off in regular fashion throughout the day. This genetic circadian rhythm influences neural signaling from the SCN to another brain structure called the pineal gland. When darkness falls, the SCN spurs the pineal to release melatonin into the bloodstream. Melatonin is a hormone that acts broadly on many of the body’s physiological systems—among other things, promoting sleep. There are competing theories about how this process relates to depression during periods of low light. In one view, too much melatonin itself is a downer, and in another, it is the timing of melatonin release that becomes problematic when days are too short. Winter months marked by excess melatonin also correspond to decreased levels of a closely related neurochemical called serotonin. Low serotonin levels are themselves associated with depression, and antidepressant drugs like Prozac and Celexa work specifically by boosting serotonin levels in the brain.
Along with heat and light, the colors in our environment can also act through our sensory systems to influence our behavior. The painter Kandinsky once declared, “Colour is a means of exercising direct influence upon the soul,” and there have been centuries of debate about his claim. Biological studies of the effects of color are sometimes traced back to a Civil War general named Augustus Pleasonton, who introduced a pseudoscientific medical technique called chromotherapy. Pleasonton’s method centered on the idea that sky-blue light promotes healing. Chromotherapy is now a component of New Age practices but is discounted by most doctors. In psychology the influence of colors is better substantiated than in medicine, however. A notable example is the phenomenon of Baker-Miller pink, a particular shade of light magenta that appears to soothe the savage beast in people. The calming effect was discovered by Alexander Schauss in the 1960s. Schauss showed that exposure to the pink hue caused a reduction in heart rate and breathing following exercise. He convinced a local prison to paint its cells with the color, and amazingly the jail reported a subsequent sharp drop in hostility among the inmates; the prison directors Gene Baker and Ron Miller became the color’s namesakes. It is difficult to rule out the contribution of cultural biases to the effects of Baker-Miller pink. The strong association of pink with femininity in American society could influence its reception in the United States, for instance. The fact that further experiments with the color have produced conflicting results supports the notion that different populations may react disparately to the stimulus.
In a rigorous study of the effects of colors on mental function, UCLA psychologist Albert Mehrabian and his student Patricia Valdez showed seventy-six different color samples of varying hue, brightness, and saturation to 250 undergraduates and asked the subjects to report their emotional responses. They found strong effects of color saturation on arousal—more saturated colors, particularly in the blue-green-yellow range, were considered to be the most arousing. In addition, specific colors differed greatly in how pleasing they were found to be; participants rated blue to purple hues significantly higher than yellows or greens. These relatively generic findings have been extended to demonstrations of the effects of colors on a variety of cognitive tasks. In one example, researchers at the Universities of Munich and Rochester showed that subjects performed significantly more poorly on IQ tests labeled with red pen than on tests marked with green or shades of gray. Although the color stimuli were too subtle for subjects to remember them, scalp electrode recordings showed that slight changes in brain activity correlated with the color-dependent behavioral effects. A 2009 paper published in Science magazine by researchers at the University of British Columbia provided a possible explanation for this result by showing that the color red tends to induce subconscious avoidance behavior. The experiments also showed conversely that the color blue tended to attract participants in both word game and product choice tests, and that blue stimuli also enhanced performance in a creativity task. Although these results certainly do not imply anything medicinal about the benefits of blue sky, they are oddly resonant with the beliefs of Pleasanton and the chromotherapists.
So people are like plants, it seems, blossoming or fading with the weather—or in some cases at the whim of interior decorators. Brighter days lead to brighter moods, hotter days lead to hotter tempers, and clearer days may foster clearer thinking. The environmental influences we have considered certainly act through our brains, but they are not governed by our brains. Placed in an ambience of fluctuating conditions, our brains absorb and reflect their surroundings, channeling external influences seamlessly into changing emotional states and behaviors. Environmentally driven periods of vigor and listlessness can have deep consequences in our lives. We pursue different goals when we are aroused, equanimous, ebullient, or depressed. Others then see us differently, affecting our careers and our relationships, and perhaps even influencing the fate of our genes. On an instantaneous basis, our emotional state helps determine how we react to specific contexts or stimuli, setting the tone for momentous decisions made in the blink of an eye—like whether to accept a marriage proposal, take a job offer, or jump off a bridge. At the same time, the environmental factors that mold our emotional makeup act on a scale that is slow compared with much of human thought and action. They also make use of only a tiny fraction of the sensory input our brains receive. How does the remaining input affect us?
The bulk of the sensory bombardment alters our behavior on a much shorter time scale than ambient light and heat. As with environmental factors that affect our emotions, fast-acting stimuli carry a force of their own—they are not merely dispatches to be received by our cognitive central processor. One way to see this is to examine how stimuli interfere with each other in ways that no central controller could possible invite. If you are like me, you experience this whenever you try to concentrate on work in the presence of background noise. The resulting conflict is a well-studied phenomenon because of its importance in educational settings. Psychology researchers have defined something called the irrelevant sound effect, which describes the disruption of short-term visual memory (critical to reading) in the presence of background acoustic stimuli. In one example of the effect, psychologist Emily Elliott administered memory tests to cohorts of different age groups while forcing them to listen to a set of irrelevant words played over and over again. She found that adult subjects performed about 10 percent worse in the presence of the irrelevant speech than in silence, while second graders performed close to 40 percent worse. Nonspeech background noises such as tones and music also compromise cognitive task performance.
Our perceptual abilities depend on interactions between stimuli, as if different inputs fight for dominance over our brains. If you are a concertgoer, you might have noticed that some people seem to appreciate music best with their eyes closed. You have probably also noticed that people almost always close their eyes when kissing. These phenomena may be due in part to the fact that shutting off visual input can heighten other perceptual abilities. Researchers at the University of London asked a group of subjects to remember letters from a visual display while at the same time testing their ability to detect a touch stimulus. The subjects detected the touch much better when the visual task was easier, showing that the visual task interfered with detection of the tactile input. In another study, scientists at the University of Jena in Germany showed that closing the eyes in either light or darkness helps improve sensitivity to tactile stimuli. One of the most bizarre illustrations of the influence of visual input over other types of stimulation is the so-called McGurk effect, an amazing phenomenon in which observed movements of a speaker’s mouth seem to override perception of speech sounds. You can listen to someone imitating a sheep—bah, bah, bah—but when this soundtrack is paired with a video of the speaker mouthing fa, fa, fa, then it is the latter sound you perceive, with an F rather than a B. As soon as you close your eyes, the sound goes back to the bleating bah, bah, bah. In the multisensory environment, what you see is what you get, even if you are supposed to be hearing something different.
The controlling influence of the environment is thrown into sharp relief through the phenomenon of attention. Attention is a metaphorical spotlight that shines on the thing or things that interest us at a given moment, and it is one of the most critical cognitive capabilities we have. The attentional spotlight determines which stimuli we most readily process, remember, and respond to. Where the light shines from is a controversial matter, however. We refer to our attention as being given and paid, but also attracted and caught, reflecting a dichotomy between forms of attention in which we ourselves are more active than passive participants. Neuroscientists convey this distinction by speaking of top-down mechanisms, apparently controlled by each individual, as opposed to bottom-up mechanisms, guided by the stimuli themselves. The great William James wrote in 1890 that “the turnings of our attention form the nucleus of our inner self,” and that the act of “voluntarily bringing back a wandering attention… is the very root of judgment, character, and will.” James even went so far as to declare that “volition is nothing but attention,” arguing that to do anything autonomously or freely in practice boils down to paying attention to it.
Bottom-up attention by nature puts the environment rather than our own brains in the driver’s seat. Walking through any metropolis, we are constantly at the beck and call of external stimuli. Our heads turn instinctively toward the honk of a horn or the nearby screech of tires on asphalt. Even a distant siren places us involuntarily on alert. The smells snaking out of a local pizzeria or Chinese restaurant also call to us, perhaps clawing at our stomachs and activating hunger for a meal. At night, the winking neon signs we pass and the flash of headlights seem to draw our gaze as if they were magnets for our eyes. These are survival reflexes, and there are good reasons for them to be deterministically written into our brains. One can imagine that the same brain mechanisms were at work when the primordial human dodged a pouncing lion or tumbling boulder.
In bottom-up attention, the biology that links sensory input to behavioral responses rests not only on the sensory systems themselves but also on the brain pathways involved in flagging stimuli as important or salient. These pathways automatically give salient stimuli more influence, while others pass unnoticed. The most salient stimuli are often those that suggest particular benefits or threats; this explains why our attention is more easily captured by the scent of pizza than the smell of car exhaust, or by a sudden crash from a construction site rather than the slow rumble of a train. Some apparently trivial sensory stimuli gain their force by association with a salient stimulus. Ivan Pavlov’s dogs learned to associate the ring of a bell with the impending arrival of food—the bell became salient, and the hounds salivated whenever they heard it. Many neuroscientists believe that salience is signaled in the brain by pulses of particular neurotransmitters. The best-studied effects arise from rewarding stimuli such as food and sex, which induce dopamine release in regions of the brain involved in motor function. Alarming stimuli by contrast seem to cause release of the neurotransmitter norepinephrine. The involvement of distinct neurochemical processes in bottom-up attentional mechanisms emphasizes again the extent that evolution has virtually hardwired behavioral responses to large classes of environmental stimuli.
In contrast to the involuntary responses evoked by salient stimuli, top-down attention is engaged by goals you set for yourself, placing it at least nominally under internal control. But even in this context, the environment you operate in plays a strong part in your behavior, influencing what you do even on a short-term basis. How this happens is brought colorfully to life in Martin Handford’s Where’s Waldo? Each page features a tiny drawing of an elflike character named Waldo, wearing a red-striped shirt and stocking cap, embedded in a huge mob of other brightly colored individuals of about the same size and shape. Our task as readers is to find Waldo, and it is not easy. Research by attention expert Robert Desimone and his colleagues shows that our visual system processes each picture in two ways. It takes in the whole scene, searching for preferred features—a blotch of red, the peak of a hat—that invite a closer look. At the same time, it zooms in on small regions at the center of the visual field, examining them to see if they directly match the impish fellow. In Handford’s complicated scenes, a thousand features beckon to our glance at once, causing our eyes to dart reflexively around the page. Our eye motions are surprisingly systematic, however; both the time we spend at each spot our eyes land on and the length of jumps our eyes make between spots in the picture are predictable, driven by characteristics of the visual scene rather than any internal decision making. A similar thing happens when we look at faces. Our gaze jumps around in a pattern that systematically takes in the eyes, nose, and mouth of the subject. Again, the details of the stimulus confronting us largely determine our brain activity and behavior.
Even at a coarser level, top-down attention is not all that top-down. It is obvious, for a start, that instructions about what to pay attention to are often derived from the environment. In laboratory experiments, top-down attention is guided by explicit visual cues like flashing signs or arrows, or by instructions delivered by researchers. In ordinary, unscripted life, shifts in top-down attention and the sometimes complex sequences of actions that follow are also instigated by trivial figments of the world around us. The taste of a cookie crumb sends Marcel Proust’s narrator into a seven-volume rhapsody about life, the universe, and everything, while a visit to a once-familiar old house inspires Evelyn Waugh’s Charles Ryder to find religion in Brideshead Revisited. The ancient historian Suetonius reported that the music of a mysterious piper drew Caesar’s troops to the banks of the Rubicon in 49 BCE, and that the minstrel’s blast upon a trumpet then triggered the general’s fateful determination to cross into Rome, beginning the civil war that led to Caesar’s dictatorship and the eventual end of the Roman Republic. Deep decisions are prompted by shallow sensory experiences.
Top-down attention is highly susceptible to the encroachment of such influences. To the dismay of classroom instructors worldwide, studies of attention spans show that most people can only pay attention to a given activity for a matter of minutes before distractions win out. Neuroscientist John Medina defines a “ten minute rule” based on the principle that audiences stop paying attention to lectures after about ten minutes. To keep people engaged, Medina proposes offering emotionally salient anecdotes or stimuli at regular intervals, in effect exploiting bottom-up attentional mechanisms to discipline listeners who are supposed to be doing better on their own. When digital devices are brought into the picture, reported attention spans become even more pathetic. A widely reported study of online habits published by Microsoft in 2015 claimed that average attention spans are as short as eight seconds, largely as the result of the prevalence of all of the electronic distractions in our twenty-first-century milieu. No matter how long it takes us to lose interest in the thing we’re paying attention to at a given moment, there is always another digital stimulus there for us to—wait, let me just read this message! It is no accident that some of the activities people seem to find most engrossing—playing video games, watching TV, surfing the web—are constantly in flux, throwing new stimuli at us all the time. When faced with such barrages, our attention is held not because our brains want it to be, but because the external world is so effective at keeping our brains in its sway.
Among the stimuli we humans receive from the world, those we get from each other pack a particular punch. We are all aware of powerful influences from other people, but the extent to which these influences degrade our own capacity for self-determination is dramatic. In 1951, a young psychologist named Solomon Asch conducted a classic experiment that demonstrates the potent effects of stimuli from peers. Asch arranged subjects into groups and asked them to take a perceptual judgment task. They were shown a card with a line drawn on it. Then they were given a second card depicting three lines, and asked to vote publicly on which line matched the line on the first card. Unbeknownst to each subject, the other members of the group were decoys—actors embedded in the experiment and instructed how to vote in the task. In some cases, the decoys were all told to vote incorrectly, even though the correct answer was always obvious. In these cases, the befuddled subjects were forced to choose between concrete visual data and the apparently unanimous but misguided opinion of others in the room. Astonishingly, most of the participants rejected the evidence of their eyes in favor of peer pressure at least some of the time. Some of the subjects went with the majority no matter what, and only about a quarter of the subjects stuck to their guns on a uniform basis.
“That we have found the tendency to conformity in our society so strong that reasonably intelligent and well-meaning young people are willing to call white black is a matter of concern,” wrote Asch in summarizing his experiment. But the conformists in his trials may have been no more free to act independently than a single fish is to swim against the school, or a single wildebeest is to deviate when the herd bolts at the approach of a predator. Just as inanimate stimuli like tones and images affect the brain in distinctive ways, so do social stimuli like human forms and voices. Modern neuroscience has revealed a constellation of brain areas that are apparently specialized for socially important functions such as recognition of human anatomy and processing of human language, some of which we saw in Chapter 4. These areas are somewhat akin to sensory systems for more basic functions like vision and hearing. They are complemented by potent mechanisms for forming and solidifying interpersonal relationships, ensuring that the signals we receive from other people are salient elements of our environment.
At first sight, there appears to be a world of difference between the way low-level input like a loud noise affects our behavior and the way the actions of another person influence us. In the latter case, we generally consider a response to some extent; for instance, the subjects in Asch’s conformity experiments considered their own level of confidence, the reliability of others in the room, and the properties of the stimuli before making a judgment. But social stimuli are not necessarily more sophisticated than loud noises. Think about how we instinctively react to a human scream, the cry of a baby, a smile on someone’s face, or an expression of fear. Functional brain imaging studies have shown that viewing emotional facial expressions in particular produces immediate consequences to brain activity that partially mimic what would be expected if the viewer herself was experiencing those emotions.
Nonemotional social stimuli can also produce virtually automatic behavioral responses. A famous example is contagious yawning, which serves no known function but has been demonstrated in both humans and chimpanzees. Another example is the phenomenon of subliminal speech priming, in which a rapidly spoken word alters the listener’s answers to subsequent questions even if the word is not consciously perceived. For instance, the word “cow” inserted imperceptibly into a stream of incomprehensible babble would make you more likely to recognize the word “cow” when later spoken more clearly. Speech priming effects show that our responses to human language can be every bit as reflexive as our involuntary responses to less animate stimuli.
Powerful evidence for the importance of social stimuli in our environment also comes from considering what happens when these stimuli are eliminated. Today, tens of thousands of Americans are subjects in a perverse experiment to measure the effects of such social deprivation: they are prisoners in solitary confinement. Prisoners in solitary confinement are kept in spartan jail cells no more than eighty square feet in size, with meals delivered through a slot in the door and at most an hour a day outside the cell for exercise. The only human contact they have is with prison guards, during delivery of meals or escort to the exercise yard. According to the advocacy group Solitary Watch, prisoners in solitary confinement report a litany of psychological difficulties, including “hypersensitivity to external stimuli, hallucinations, panic attacks, cognitive deficits, obsessive thinking, paranoia, and impulse control problems.” A prisoner interviewed by journalist Shruti Ravindran described a particular form of breakdown that takes place in solitary. “All day, every day in the Box people go off. They yell, they scream, they talk to they-self.… At two or three in the morning, somebody starts screaming ‘Aaaaaa!’… you shake your head sayin’, ‘Another one.’” A 1972 study looked at neuroelectrical signals from prisoners and found that inmates housed alone over multiple days showed a general slowing of brain waves. Backing up behavioral findings of stimulus hypersensitivity, the recordings also showed faster brain responses to flashes of light than observed in nonconfined prisoners. Clearly the brain of an isolated person is a different species from the brain in its natural, socially enriched environment.
A slippery slope leads from the momentary stimuli that affect individuals to global cultural factors that can shape entire populations. Social stimuli on a grand scale include the effects of everything from wars, famines, and mass migrations to divorce rates, education, and the internet. These factors set a stage on which we are merely players. You may have read the “xeroxlore” that heaven is where the police are British, the lovers French, the mechanics German, the chefs Italian, and it is all organized by the Swiss, while hell is where the police are German, the lovers Swiss, the mechanics French, the chefs British, and it is all organized by the Italians. While anyone with an extended set of European friends might rebel against the crude stereotypes, the possibility of culturally conditioned behavioral patterns is very real. For instance, French people turn out to be significantly more tolerant of extramarital affairs than other national groups, according to a 2013 Pew survey of thirty-nine countries. It is extremely unlikely that this means anything about the intrinsic nature of the French brain—genetically, French citizens are actually closer to the highly monogamous Swiss than to any other European nationality. Instead, cultural traits like attitudes toward marriage are complex social stimuli that influence our brains via the environments we live in.
Neuroscientist Michael Gazzaniga argues that “the space between brains that are interacting with each other” may be part of what “holds the answer to our quest for understanding mind/brain relationships.” Properties such as our perceived autonomy and free will, which some have struggled to associate with the activity of individual brains, emerge instead from what Gazzaniga describes as multilayered, distributed interactions involving many people. It is perhaps in a similar spirit that the metaphysical poet John Donne expressed his famous thesis, “No man is an island, entire of itself.” In Donne’s timeless words, “Any man’s death diminishes me, because I am involved in mankind.” If minds arise from social interactions among brains, then the alteration or death of any person’s brain presents an unmistakable insult to the collective experience.
In this chapter, I have argued that brains must be considered not only in relation to each other but in relation to the entire environment. Our brains are buffeted constantly by stimuli that impinge on us from all directions. The external influences range from subtle changes in our sensory milieu to sharper impulses from both animate and inanimate sources. These influences are not merely informational dispatches to a command center in our heads; they are causative forces that slice to the deepest levels of our brains and minds. The nervous system translates environmental input into behavioral output in much the same way that the physical structure of a tree determines how sun, wind, and rain guide its growth and movements. A tree opens its leaves toward the light and sways to avoid damage from powerful gusts, but only in a twisted sense could one say that the tree controls its actions. Instead, trees and brains are both fundamentally responsive to the world around them. Neither one presents a firewall against the environment, a place where the environment’s role changes from active to passive. Neither one can be understood apart from the forest of influences that envelops it.
The dichotomy between internal and environmental drivers of brain function discussed here parallels other dichotomies we have covered in Part 1 of this book: between brain-specific and body-wide aspects of mental function in Chapter 5, between localized and diffuse brain processing in Chapter 4, between complexity and tractability of neural systems in Chapter 3, and between inorganic and organic views of brain physiology in Chapter 2. In each case, the former view emphasizes ways in which the brain is different or dissociated from the rest of the natural world, leading to popular mythologies about what the brain is and how it works. Depictions of the brain as inorganic, hypercomplex, functionally self-contained, and autonomously powerful present the brain as a surrogate for the disembodied soul and feed the attitude that I have termed scientific dualism. The cerebral mystique rests on this attitude, which is pervasive even among people like myself, who believe in a material basis for the human mind. Rejecting it requires accepting the biological basis of the mind on its own terms and witnessing how the brain, body, and environment work together to shape us. Achieving this was the objective of Part 1. In Part 2, we will turn from science to society by examining why abandoning the cerebral mystique and embracing the biological mind should matter to us both as individuals and as a global civilization.