FOURTEEN

Basal Ornithischia
Ornithischia is a long-established and well-corroborated clade of dinosaurs that share a considerable number of anatomical characters. In recent years it has become apparent that a number of taxa of stratigraphically early and anatomically conservative ornithischians can be recognized. These are referred to in this chapter as basal ornithischians and, in the absence of further data, occupy a position on the ornithischian cladogram on the stem prior to Genasauria.
The phylogenetic positions of Lesothosaurus, Pisanosaurus, Technosaurus, and several other taxa, notably Fabrosaurus, that are based on nondiagnostic remains (table 14.1) have proved controversial. Recent cladistic analyses of their relationships to other ornithischians have demonstrated, however, that they include the most primitive members of this large and diverse dinosaurian group (Gauthier 1986; Sereno 1986, 1991b). The most basal ornithischians known to date are small (1–2 m long), obligate bipeds that exhibit anatomical features (tooth and jaw shape, as well as pelvic morphology) normally linked with a primarily herbivorous diet.
Definition and Diagnosis
Ornithischia is a stem-based taxon defined as all dinosaurs closer to Iguanodon than to Cetiosaurus. It can be diagnosed on the basis of the following characters: a rostral premaxillary border with a rugose edentulous gap in the midline; an edentulous predentary bone capping the dentary symphysis; the coronoid process of the dentary overlapping the surangular; median premaxillary rugosities above the edentulous oral margin; the palpebral, a rodlike bone attached to the rostral orbital margin and curving across the orbit; six slightly bulbous, conically pointed premaxillary teeth; maxillary and dentary teeth with cylindrical roots separated from the crown by a constriction; the base of the crown expanded to form a cingulum; the maxillary and dentary crowns laterally compressed and broadly triangular in lateral profile, with a thick central portion, the mesial and distal edges of the crown bearing a fringe of five to nine simple, pointed denticles; the crowns of the teeth wider at the base than at the roots and consequently arranged in an en echelon fashion; the long axis of the crown offset from the long axis of the root in mesial or distal view; dentary teeth graded in size, the largest crowns being found more distally within the dentition; the sacrum comprising four sacrals carrying sacral ribs and one sacrodorsal; a long, slender, rod-shaped pubis oriented caudoventrally, lying parallel to the ischium, and terminating distally in a short, median pubic symphysis that lies adjacent to the distal end of the ischium; a deep, tab-shaped prepubic process; a long, transversely compressed, tapering preacetabular process projecting from the dorsal edge of the ilium beyond the prepubic process; prominent lateral expansion of the ischial peduncle of the ilium; a prominent and narrow, pendant fourth trochanter on the femur; and ossified tendons that are longitudinal elements arranged epaxial to the dorsal, sacral, and caudal vertebrae.
Anatomy
Lesothosaurus diagnosticus is well known from skull and postcranial material (figs. 14.1, 14.4; Thulborn 1970a, 1972; Santa Luca 1984; Weishampel 1984a; Crompton and Attridge 1986; Sereno 1991b). Additional postcranial material thought to pertain to this taxon (Santa Luca 1984) is also discussed here. Pisanosaurus mertii was described by Casamiquela (1967b) and Bonaparte (1976) and partially reillustrated by Sereno (1991b). The material, which includes portions of the skull, the axial skeleton, and the hindlimb, with a few fragments of the shoulder and pelvic girdles, is incomplete and poorly preserved. Size differences among these specimens suggest that the association is partially incorrect. When originally described, Technosaurus smalli was based on a small number of cranial and postcranial elements pertaining to at least two individuals (Chatterjee 1984). Sereno (1991b) reviewed this material and concluded that the lower jaw with teeth (fig. 14.2C) belonged to an ornithischian but that some of the material (the premaxilla and the caudal end of the mandible) probably belonged to a smaller prosauropod, while the remainder proved to be indeterminable.
Skull and Mandible
The caudal half of the skull of Lesothosaurus (fig. 14.1) is box-shaped, while rostral to the orbital region the snout is elongate and tapered. The external naris is small, there is a large, shallow, triangular external antorbital fenestra, and the orbit is large and partially traversed by an elongate palpebral bone; the infratemporal fenestra is oblong with an oblique axis, while the supratemporal fenestra is oval in outline. The lower jaw has a large external mandibular fenestra bordered by the dentary, surangular and angular.
TABLE 14.1
Basal Ornithischia
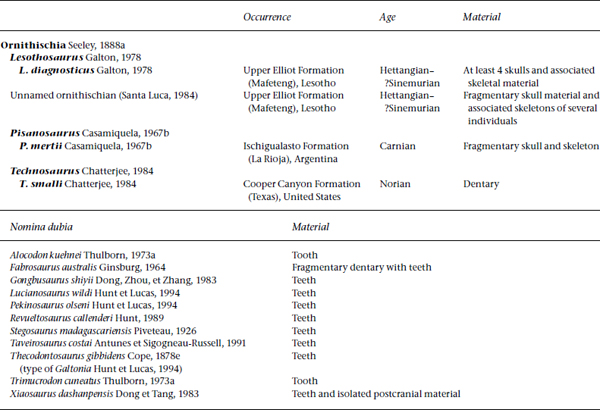
The premaxilla of Lesothosaurus forms the lower half of the opening for the naris, and a well-marked narial fossa extends to just above the labial margin. The dorsal premaxillary process meets its counterpart as a distinct butt joint and contacts the nasal in a scarf joint rostral to the naris. A tapering, lateral process wedges between the maxilla and the nasal, completely excluding the maxilla from the margin of the naris. Foramina pierce the premaxillary body immediately rostral to the naris and close to the midline (Crompton and Attridge 1986); adjacent to this edentulous area the tip of the premaxilla is roughened (Sereno 1991b:fig. 6c) as if for attachment of a keratinous rhamphotheca. There are six teeth in the premaxilla in Lesothosaurus (Sereno 1991b). The ventral part of the palatal premaxillary surface is excavated to receive the rostral process of the maxilla.
The nasal in Lesothosaurus forms the entire dorsal margin of the external naris and roofs the nasal cavity. Ventrally the nasal contacts the maxilla and the prefrontal. Medial to the nasal-prefrontal joint the nasal overlaps the frontal.
The maxilla in Lesothosaurus surrounds most of the external antorbital fenestra, so that only the lacrimal and a narrow rostral process of the jugal bound its caudal margin. The external antorbital fenestra delimits an extensive but shallow antorbital fossa in the dorsolateral wall of the maxilla. The medial wall of the fossa, formed by the maxilla and the lacrimal, is penetrated by a small, internal antorbital fenestra connecting the nasal cavity and the antorbital fossa. The dorsal part of the fossa is formed by a simple beveling of the maxillary wall and deepens ventrally to form a distinct trough delimited by a lateral lip, the supralveolar lamina (Witmer 1997a). The region just dorsal to the teeth presents a concentration of external neurovascular foramina, most of which converge on a slitlike aperture within the floor of the antorbital cavity; these foramina conveyed branches of the maxillary neurovascular bundle (Witmer 1997a). Rostrally the upper edge of the maxilla forms a scarf-edged suture with the lateral process of the premaxilla, and a medially directed rostral process contacts the premaxilla near the midline; unfortunately, this part of the skull is not sufficiently well preserved for us to determine whether the vomer and the maxilla lock into the back of the premaxilla. Beneath the orbit, the upper surface of the maxilla forms an elongate, narrow facet for attachment of the jugal. Immediately beneath the lower edge of the external antorbital fenestra the body of the maxilla is slightly deflected medially. Pisanosaurus exhibits a deep, more extensive lateral buccal recess on its maxilla. There are 15–16 tooth positions in the maxilla in Lesothosaurus (Sereno 1991b) and at least 11 in the partial maxilla of Pisanosaurus (fig. 14.2A); Bonaparte (1976) estimated a full maxillary complement of 16 to 18.
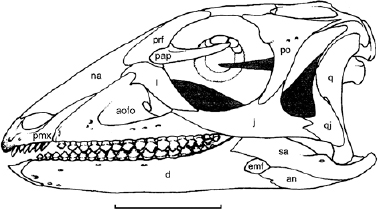
FIGURE 14.1. Lesothosaurus diagnosticus, skull in left lateral view. Shading indicates the rostral ramus of the quadrate and palatine in the floor of the orbit, the styloid parasphenoid at the rear of the orbit, and the pterygoid-quadrate plate through the infratemporal fenestra. Scale = 2 cm.
FIGURE 14.2. A, B, Pisanosaurus mertii: A, maxillary fragment in lateral view; B, lower jaw in lateral view. C, Technosaurus smalli, lower jaw in lateral view. Scale = 1 cm. (A, B, after Bonaparte 1976; C from Chatterjee 1984.)
The lacrimal in Lesothosaurus is a curved, strutlike element that interlocks against a fingerlike process of the maxilla around the external antorbital fenestra (Sereno 1991b). In doing so the lacrimal separates the antorbital fossa and the orbit in a structural arrangement that is similar to that seen in the ornithopod Hypsilophodon (Galton 1974a) and Dryosaurus (Janensch 1955). The lacrimal articulates dorsally with the prefrontal and ventrally with the jugal along short scarf joints. The orbital surface of the lacrimal is pierced by the nasolacrimal canal. Although the canal cannot be directly traced further rostrally, it clearly did not pierce the ventral ramus and thus must have taken a more dorsal course through the rostral ramus, as seen in Hypsilophodon and the basal sauropodomorph Plateosaurus (Witmer 1997a, 1997b).
The prefrontal has a teardrop outline. It contacts the nasal rostrally, the lacrimal ventrally, and the frontal medially; its lower orbital edge is notched to support the base of the supraorbital, and it forms the rostrodorsal margin of the orbit. The frontal forms the broad rostral portion of the skull roof and contributes extensively to the dorsal margin of the orbit. A transverse and moderately interdigitate suture is made with the parietal. The interfrontal joint is straight throughout its length. The fused, broad parietals form both the caudal portion of the skull roof and a ledge that slightly overhangs the occiput. There is a low but distinct sagittal crest. Caudally the parietal is drawn out into laterally projecting flanges that contact the squamosals along the crest of the paroccipital processes.
The short medial process of the postorbital contacts the frontal and the parietal along a slightly interdigitate suture. The ventral process reaches nearly to the body of the jugal to form virtually the entire postorbital bar but articulates principally along the rostrolateral edge of the dorsal process of the jugal. The postorbital makes an extensive scarf joint with the squamosal as a long, overlapping, tapering fingerlike process; together these bones form the supratemporal arch.
The jugal forms the lateral wall of the adductor chamber, covering the coronoid process of the mandible. It makes a scarf articulation with the postorbital and extensively overlaps the quadratojugal; rostrally it contacts the lacrimal and maxilla. The quadratojugal is large and V-shaped. Dorsally it forms a tapering process that runs up the leading edge of the quadrate, terminating where it contacts the prequadratic process of the squamosal, excluding the quadrate from the margin of the infratemporal fenestra.
The area immediately above the prequadratic process of the squamosal forms a ledge that probably represents the area for attachment of M. adductor mandibulae externus superficialis (cf. Galton 1974a). The head of the quadrate fits into a deep pocket on the ventral surface of the squamosal. Directly behind this pocket the postquadratic process overlaps the lateral aspect of the paroccipital process.
Broad and flat dorsally, the quadrate head is buttressed caudally by a small protuberance on the upper part of the quadrate shaft that contacts the postquadratic process of the squamosal. The shaft of the quadrate is shallowly concave caudally, while rostrally it is subdivided vertically into pterygoid and jugal rami. The jugal ramus is notched for a small paraquadratic foramen located between the jugal and the quadratojugal and mostly hidden in lateral view. The deep pterygoid ramus forms an extensive overlapping suture with the pterygoid that is visible through the infratemporal fenestra. The distal condyle of the quadrate is transversely broad and forms a bicondylar jaw joint.
Sereno (1991b) described the palate and braincase of Lesothosaurus. The supraoccipital is subtriangular and transversely arched, and there is a midline ridge. It forms the entire dorsal margin of the foramen magnum and laterally rests on the medial portions of the paroccipitals. Laterally and dorsally the supraoccipital is extensively fused to the parietals, while rostrally it reportedly has a lateral exposure on the braincase, considered an epiotic by Sereno (1991b). The exoccipitals and opisthotics are indistinguishably fused to form the paroccipital processes; a delicate footplate and ossified proximal section of the stapes has also been illustrated (Sereno 1991b). The paroccipital processes project more or less horizontally from the lateral margins of the foramen magnum and widen slightly distally; they are slightly pendant when viewed laterally. The prootic forms a large, thick triangular plate that makes up much of the dorsal part of the lateral wall of the braincase. Rostrally it forms a butt joint with the laterosphenoid, which tapers and swings laterally to underlie the postorbital and form the rostral wall of the supratemporal fenestra. The basioccipital and basisphenoid form robust hemicylindrical elements on the floor of the braincase. The crescentic occipital condyle extends to the base of the exoccipitals, which form small condylids on either side of the foramen magnum. Neither the structure of the basioccipital-basisphenoid suture nor those of the basal tubera are known. The prominent basipterygoid processes extend rostrolaterally and slightly ventrally to terminate in bluntly rounded articular surfaces that contact the central body of the pterygoid. Between the basipterygoid processes is a long, styloid parasphenoid that projects rostrally and essentially horizontally; its dorsal surface bears a sulcus for the cartilaginous interorbital septum.
The vomers are long, thin triangular plates that lie on the midline of the palate. Each vomer has been reported as being fused to its neighbor along the ventral margin, forming a median keel. The palatine is a transversely broad plate that is firmly sutured to the maxilla. Its dorsal surface bears two excavations separated by a strong ridge; the caudal excavation grades into the pterygoid bone and represents a fossa for the dorsal pterygoideus muscle, while the rostral fossa is associated with the nasal or, more likely, the antorbital cavity and thus may be a pneumatic structure (Witmer 1997a). The palatine articulates with the pterygoid medially, and the latter two bones are locked against the maxilla by a U-shaped and straplike ectopterygoid bounding a small suborbital fenestra. The pterygoid is not well known, although the quadrate ramus is thin and deep, contracting toward the central plate and the basal articular region. Short, stout ventral pterygoid flanges are present, and the rostrodorsal process projects forward and between the palatines as flat plates.
A small, arrowhead-shaped predentary caps the rostral tip of each dentary. The short, lateral processes of the predentary wrap around the tip of each dentary and dorsally along the edentulous oral margin. A median ventral process fits wedgelike under the full length of the dentary symphysis. Large vascular foramina are found in the predentary body at the base of each lateral process and on adjacent areas of the dentaries.
The dorsal and ventral margins of the dentary in Lesothosaurus, Pisanosaurus, and Technosaurus are subparallel for most of its length; these edges diverge caudally to accommodate the postdentary bones and to support a coronoid process that lies on the rostral margin of a dorsal expansion of the surangular (fig. 14.1). Neurovascular foramina are found scattered along the lateral face of the dentary and also concentrated close to the symphysis. The dentaries meet rostrally at a narrow but spout-shaped edentulous symphysis that is deflected ventrally (to accommodate the predentary), shallow, and ovoid in shape. The rugose symphyseal surface is set at an angle to the horizontal. In Lesothosaurus the dentary accommodates as many as 20 tooth positions (Gow 1981); at least 15 positions are recorded in the damaged and incomplete lower jaw of Pisanosaurus; Technosaurus is too fragmentary to offer meaningful information. Whether there is a slight rostral diastema between the first tooth and the caudal margin of the keratinous predentary beak (Crompton and Attridge 1986; Weishampel and Witmer 1990a; contra Sereno 1991b) is unclear. The distal end of the tooth row is slightly inset medially from the side of the dentary. This condition is also seen in Technosaurus (fig. 14.2C) and Fabrosaurus australis (Thulborn 1992). The dentary teeth of Pisanosaurus (Bonaparte 1976) are clearly inset along most of the length of the dentary.
Lesothosaurus has a well-developed external mandibular fenestra. Casamiquela (1967b) described a large opening at the junction of the surangular, dentary, and angular in Pisanosaurus, which he and subsequently Bonaparte (1976) considered to be a preservational artifact. However, this is the approximate position for an external mandibular fenestra, and Sereno (1991b) claimed to have seen some bony edges supporting such an interpretation. On the medial wall of the dentary (and maxilla) in Lesothosaurus a row of alveolar foramina (one per tooth position) represents the position of access for neurovascular supply associated with the dental lamina.
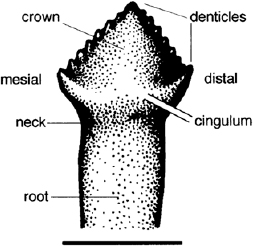
FIGURE 14.3. Dentary tooth of Lesothosaurus diagnosticus in labial view. Scale = 1 cm.
The caudodorsal edge of the dentary swings dorsally and lateral to the dentition to form a tongue-shaped process that overlaps the surangular and forms the elevated coronoid process of more derived ornithischians. The extent of development of the coronoid region in Pisanosaurus cannot be established with confidence because this area of the lower jaw is clearly broken.
The surangular forms the major portion of the coronoid process, the dorsal margin of the external mandibular foramen, the inner wall of the adductor fossa, the lateral wall of the glenoid, and the lateral aspect of the retroarticular process. Laterally the rim of the jaw joint is buttressed by a liplike thickening of the surangular, beneath which is found a small surangular foramen. The angular forms the caudoventral margin of the lower jaw, wrapping around to the medial side beneath the articular. It also forms a caudal portion of the external mandibular fenestra. The articular forms the medial portion of the jaw joint and is lodged between the surangular, the angular, and the prearticular. The prearticular is an elongate strap of bone lying medial to the articular, ventral to the adductor fossa and (in all probability) overlapping the medial wall of the dentary. The rostral part of the prearticular was overlapped by the platelike splenial. The long, deep splenial covers much of the medial surface of the dentary and the channel forming the mandibular canal.
The dentition of Lesothosaurus has been described by Thulborn (1970a, 1971a), Galton (1978), Weishampel (1984a), Crompton and Attridge (1986), and Sereno (1991b). Premaxillary teeth are procumbent, narrow, conical, moderately recurved, and slightly spoon-shaped toward the apex (fig. 14.1). A neck and slight swelling separates the base of the crown from the long, subcylindrical roots. On the lingual surface of the more mesial premaxillary teeth a vertical furrow and an adjacent sharp ridge extend toward the mesial edge of the crown. More distally along the series the last two crowns acquire mesial and distal denticles. Both the maxillary and the dentary tooth crowns are buccolingually compressed and mesiodistally expanded to assume their characteristic subtriangular shape (fig. 14.3), and both are entirely coated by an even layer of prismatic enamel. From five to nine denticles are found on the mesial and distal edges of the maxillary and dentary teeth. With slight distal displacement of the crown apex, there are often slightly higher numbers of denticles on the mesial edges. The crown meets the root at a distinct neck, above which the base of the crown is swollen to form a cingulum (fig. 14.3) that extends mesially and distally to form the first denticle at the base of each edge. In mesial and distal view the long axis of the crowns is offset against the long axis of the root. Teeth vary in size along the tooth row, increasing in size distally along the series and reaching a maximum toward the rear half of the series.
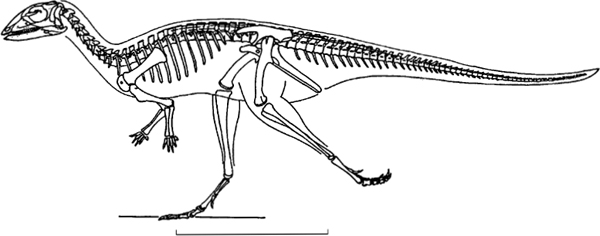
FIGURE 14.4. Lesothosaurus diagnosticus, skeletal reconstruction. Scale = 20 cm.
The roots of the maxillary and dentary teeth are closely packed; consequently adjacent crowns are imbricated and develop interdental pressure facets where they rub against each other during occlusion. Wear facets usually develop on the mesial and/or distal halves of the crown (on the lingual side for maxillary teeth, in the buccal for dentary teeth), although occasionally crowns show apical truncation and abrasion. Most adjacent facets are set at obtuse angles, but such facets are irregularly and unevenly scattered along the dentition. Individual wear facets are oriented nearly vertically (subtending a low angle to the longitudinal axis of the tooth).
While the above description is broadly applicable to the little that is known of Technosaurus, the dental morphology in Pisanosaurus differs significantly. In Pisanosaurus the maxillary and dentary teeth are closely packed and apparently lack a cingulum. Occlusal surfaces are continuous between adjacent teeth. For the maxillary teeth, which curve lingually from their base, the occlusal plane is inclined at approximately 45° to the vertical. The dentary teeth vary in size, from small mesially to large distally. Unlike the maxillary crowns, the dentary crowns are straight and vertical. The slightly buccally concave wear surfaces are inclined 60° to 70°.
Postcranial Skeleton
The postcranial description relies principally on the work of Thulborn (1972) on Lesothosaurus (fig. 14.4), with some additional observations by Santa Luca (1984) and Sereno (1991b).
AXIAL SKELETON
To date, the vertebral column of Lesothosaurus has not been sufficiently well preserved to allow a full description. The proatlases are small, curved, straplike elements that articulate with the prezygapophyses of the atlas arches and link the atlas to the occiput. The neural arch of the atlas is a paired structure, each part comprising a pedicle that is expanded and articulated with the margins of the intercentrum; it also forms part of the facet for the occipital condyle. Above this base the arch contracts and curves dorsomedially to approach its neighbor in the midline, forming an enclosure of the nerve cord. The upper part of the neural arch sends out a prezygapophysis that faces slightly dorsolaterally and forms a platform for the proatlas. Caudally the ventral surface forms a zygapophyseal facet for articulation with the axis and a laterally projecting stylelike process, or epipophysis (Sereno 1991b). The atlas intercentrum and odontoid are not known to date. The axis preserves neither the intercentrum nor an odontoid but comprises a spool-shaped centrum with amphiplatyan articular ends; dorsally a high, curved neural spine extends caudodorsally to overhang the succeeding vertebra. The prezygapophyses face dorsolaterally, while the postzygapophyses face ventrolaterally and are supported on short, stout processes that project from the base of the neural spine. There is no indication of discrete articulation facets for an axis rib on either the centrum or the neural arch. The third cervical has a dorsoventrally flattened, spool-shaped centrum with a broad ventral keel and amphiplatyan articular ends. There are facets for both the capitulum and the tuberculum on the lateral wall of the centrum (a parapophysis on the lower rostral rim of the centrum, a diapophysis on the neurocentral suture, there being no discrete transverse process) as first described by Santa Luca (1984). The neural spine is short and hooked. The pre- and postzygapophyses are prominent and well separated from the midline and have facets that are angled at approximately 45° to the vertical. Sereno (1991b) noted the presence of flanges (epipophyses) above the postzygapophyses in this vertebra (similar structures are found only on the fourth cervical of Heterodontosaurus [Santa Luca 1980:fig. 5A]). More caudal cervicals assume a slightly more usual form (Santa Luca 1984) in the sense that they develop a horizontal shelf that connects the pre- and postzygapophyses and from which develops a discrete, robust transverse process bearing the diapophysis for the tuberculum of its rib. The centra are spool-like but compressed rostroventrally to produce a narrow keel that broadens caudally. The articular faces are amphiplatyan. The parapophysis migrates dorsally from its position on the rostral margin of the centrum until it crosses the neurocentral suture in caudal members of the series. The neural spine is short and hooklike along the entire cervical series, and the pre- and postzygapophyses are prominent and widely spaced.
Three of the vertebrae referred to Pisanosaurus were originally thought to be caudals by Casamiquela (1967b) but have been interpreted by Bonaparte (1976) as cervicals; however, they and other assorted postcranial remains are too small to pertain to the same individual to which the lower jaw belongs.
The dorsal centra of Lesothosaurus are spool-shaped, and rostral members of the series are more laterally compressed and retain a narrow (cervical-like) ventral keel that is lost in more caudal centra. The centra are amphiplatyan, even though larger individuals have been reported to have generally amphicoelous centra (Santa Luca 1984). The transverse processes are stout in Lesothosaurus and range from dorsolateral to horizontal, especially caudally. Parapophyses are located dorsal to the neurocentral suture and before the base of the transverse process; they migrate dorsally toward the diapophysis in the caudal portion of the series. The neural spines are short and roughly rectangular (Thulborn 1972), although Santa Luca (1984) noted that the edges of the spine diverge in some individuals, creating a trapezoidal profile.
The sacrum of Lesothosaurus is composed of five sacrals bearing specialized sacral ribs and a fused sacrodorsal bearing a normal, albeit short, rib. The broad sacral centra are spool-shaped but dorsoventrally compressed and have reniform articular surfaces. A narrow, median keel is variably developed. The broad, horizontal transverse processes undoubtedly were extensively ligamentously bound to the sacral ribs. The neural spines are transitional in shape between the dorsals and caudals, ranging from trapezoidal cranially to rectangular caudally (Santa Luca 1984).
The number of caudal vertebrae cannot be estimated at present. The spool-shaped caudal centra become lower and more cylindrical in the distal part of the series. The articular surfaces become more circular distally. The proximal caudals bear ventral chevron facets on their distal surfaces, but hemal arches are absent in the distal portion of the series. The proximal caudal neural arches exhibit robust, rectangular spines and long, horizontal transverse processes. All of these structures become progressively reduced along the series, so that the spines become low ridges (mirroring the loss of the chevrons) and the transverse processes are lost. The prezygapophyses overlap and clasp the postzygapophyses; as a result they appear longer than the postzygapophyses simply because the latter lie alongside the reclined neural spines.
Ribs are two-headed except at the extreme ends of the dorsal series. Changes in the position of diapophyses and parapophyses reflect the structure and position of the ribs: the tuberculum and capitulum approach each other in successively more caudal portions of the column.
Ossified tendons (fig. 14.4), described in Lesothosaurus by Thulborn (1972), are found epaxially on the dorsal and caudal portions of the column. Tendons vary in structure from flattened and splay-ended to rodlike. Although subject to postmortem displacement, they are arranged loosely in bundles on either side of the neural spines of the caudal dorsal, sacral, and rostral caudal vertebrae.
APPENDICULAR SKELETON
The scapula is elongate (10%–15% longer than the humerus) and bowed around the rib cage, with a strongly expanded dorsal edge, a narrower shaft, and an expanded base where it is sutured to the coracoid and supports the humeral glenoid. The distal edge of the scapula is rugose and pitted, indicating the likely presence of a cartilaginous suprascapula. The proximal portion is expanded to form a prominent acromion, a broad sutural area for the coracoid, a notch leading to the coracoid foramen, and the hooklike buttress for the humeral glenoid. The coracoid is a thick, subrectangular plate that forms the lower half of the glenoid; it is pierced laterally by a discrete coracoid foramen.
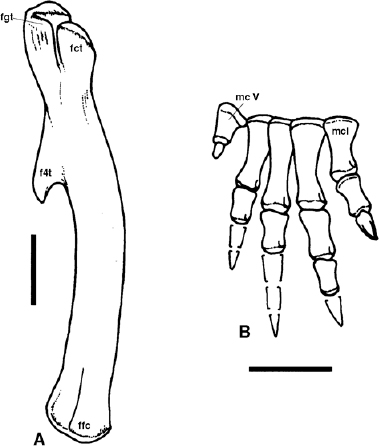
FIGURE 14.5. Lesothosaurus diagnosticus: A, femur; B, manus in dorsal view. Scale = 2 cm.
The humerus is a simple bowed bone (fig. 14.4). The humeral head is bulbous, centrally placed, and flanked by narrower lateral and medial shoulders. The deltopectoral crest is proximally positioned and projects prominently from the lateral margin of the shaft as an extension beneath the lateral shoulder. The distal end of the humerus comprises rounded lateral and medial condyles that form a trochlear structure (Santa Luca 1984).
The radius, which is about 70% as long as the humerus, is a straight bone with an oval cross section and expanded articular ends. Its proximal articular surface is slightly depressed and subcircular. Its carpal articular surface is also subcircular and slightly warped. The ulna is slightly longer and more robust than the radius, and there is no olecranon process.
Thulborn (1972) identified two carpal bones in the forelimb of Lesothosaurus. In addition, he described and reconstructed a partial associated manus (Figure 14.5B). Further preparation of this material has revealed additional bones, notably metacarpal V, an associated small, nubbinlike phalanx, and the remainder of digit I, all of which confirms Thulborn's original assessment. The phalangeal formula of the manus has been reconstructed as 2-3-4-3-1 (Sereno 1991b:fig. 5B). The manus is short and broad, with the metacarpals forming a slightly divergent array; the bases are twisted and imbricate such that the medial elements overlap the more lateral elements. Metacarpal I is shorter and stouter than metacarpals II and III; the latter two are equal in length, while metacarpal IV is intermediate in length between metacarpal I and metacarpal III, and metacarpal V is short and blocklike. The distal articular surface of metacarpal I is trochlear and asymmetrical, with the lateral condyle more prominent than the medial; as a result the two phalanges of digit I diverge slightly from the remainder of the hand and can be flexed obliquely across the palm. In similar fashion digit V, even though foreshortened, is also twisted away from digits II–IV. The ungual of the first digit is pointed and flattened rather than strongly decurved and talonlike as in Heterodontosaurus (Santa Luca 1980).
The hindlimb is gracile and much longer than the forelimb. The ilium is long and deep (fig. 14.6). The preacetabular process is laterally compressed, tapers cranially, and varies from straight to slightly decurved (Santa Luca 1984). The postacetabular process is short and deep. The dorsal margin is essentially horizontal and slightly thickened, ending at an obliquely truncated edge. Beneath this area the blade of the ilium sweeps cranioventrally to the ischial peduncle. The ventral surface of the postacetabular blade is dominated by a well-developed brevis shelf; this shelf is horizontal and undercuts the postacetabular process but is walled off medially by a thin curtain of bone that is supported medially by the sacral yoke. The ischial peduncle is flattened ventrally but forms a discrete ovoid swelling that projects laterally and forms a buttress at the back of the acetabulum. The acetabulum is deep and overhung by a pronounced, oblique supracetabular crest. The medial wall of the acetabulum extends ventrally as a thin sheet, so that the perforate acetabulum, which is characteristic of dinosaurs, is almost occluded. The pubic peduncle is triangular in section and unexpectedly large and robust, bearing much of the supracetabular crest that projects obliquely cranioventrally, and is bluntly truncated distally. (Whether this process can be said to taper or expand distally in lateral profile is dependent solely on the degree of preservation of the acetabular margin.) Medially there are four to five facets for the sacral ribs; the precise positions of the more caudal facets are unclear probably because the sacral ribs fuse distally to form a sacrocostal yoke.
In Lesothosaurus the long, rodlike pubis meets its counterpart in a distal symphysis. The short, deep prepubic process (fig. 14.6) consists of an oblique, laterally concave plate of bone rostral to the obturator foramen. The pubic contribution to the acetabulum is a thickened lip immediately above the obturator foramen; a ventral process from the acetabular margin and a dorsal process from the pubis enclose the latter. Opisthopuby in Pisanosaurus cannot be confirmed (contra Bonaparte 1976) due to the poorly preserved impression of the pelvis.
The ischium is a long, curved bone lying parallel to the pubis (fig. 14.6). Its proximal end is expanded to form the caudoventral rim of the acetabulum. The iliac peduncle is short and blunt; the pubic peduncle is longer but equally blunt. The shaft of the ischium exhibits some torsion and is bowed cranially. The caudal edge bears a distinctive longitudinal groove. Thulborn (1972:41) identified a proximally situated obturator process that he illustrated and described as “a thin and sheet-like extension of the rostral margin.” Sereno (1986:247) denied its presence in the same material but referred to a form “of possible close affinity to Lesothosaurus” that does possess an obturator process. Sereno (1991b) reillustrated and described the ischium of Lesothosaurus, almost entirely confirming the original description by Thulborn but repeating the claimed that an obturator process was absent. However, no mention was made of other lesothosaur-like material (see also Thulborn 1992). No consideration was given to the ischium described by Santa Luca (1984), whose material was considered to be “referred correctly to Lesothosaurus diagnosticus” (Sereno 1991b:171), which exhibits a prominently crested, proximally positioned obturator process (Santa Luca 1984:fig. 18).
In Lesothosaurus there is a prominent thin and sheetlike extension to the craniomedial margin of the ischial shaft, rather than the discrete tab-shaped obturator process seen in derived ornithopods. In both Lesothosaurus and nonheterodontosaurid ornithopods the pubis lies lateral to the obturator process and presumably gained a measure of mechanical support. Although debatable, Thulborn's sheetlike obturator process in Lesothosaurus may be a homologue of the tab-shaped obturator process in more derived ornithopods.
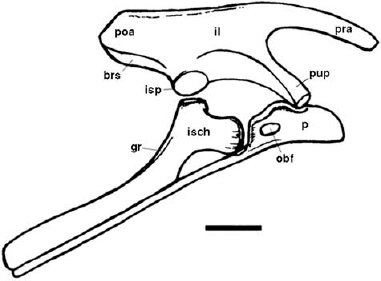
FIGURE 14.6. Lesothosaurus diagnosticus, pelvis in right lateral view. Scale = 5 cm.
The femur (fig. 14.5A) is elongate and has a bowed shaft. Proximally the articular surface that formed the femoral head is twisted dorsomedially rather than set off from the shaft by a distinct neck. A warped, tapering shoulder connects the region of the femoral head with a small, rostrally curved ridge that forms the greater trochanter; the latter is separated by a deep cleft from the laterally flattened and vertically oriented cranial trochanter. On the caudomedial edge of the femoral shaft, just below the in-turned head, is a well-developed, pendant fourth trochanter. There is no extensor groove between the distal femoral condyles, but there is a broad, deep caudal intercondylar fossa. The medial condyle is slightly larger than the lateral.
The tibia in Lesothosaurus is about 25% longer than the femur. The laterally compressed, robust cnemial crest intervenes between the convex medial surface and the concave lateral surface. A thickened crest runs distally from the lateral condyle. The distal tibia is laterally expanded, and the articular surfaces for the proximal tarsals are characteristically step-shaped. A medial transverse, raised ridge marks the dorsal extent of the astragalar articulation. Laterally a distinct ramp forms a raised step that separates the raised (cranially offset) lateral articular area from the astragalar facet. The lateral articular area has a craniolateral facet for the fibula and distally forms an attachment area for the calcaneum.
The fibula in Lesothosaurus is slender and approximately equal in length to the tibia. The laterally compressed proximal end of the fibula expanded into pronounced cranial and caudal processes. The distal end of the shaft is flattened for attachment to the tibia, and its articular end abuts the calcaneum.
In Lesothosaurus the tarsus consists of the astragalus, the calcaneum, and two disclike distal tarsals (Santa Luca 1984). The astragalus is wrapped around the transversely cylindrical distal surface of the medial articular surface. This contact is reinforced by a short, subtriangular, robust ascending process of the astragalus that projects proximally onto the rostral face of the tibia and rises obliquely toward the intercondylar sulcus. The calcaneum is described as crescentic (Santa Luca 1984) but presumably caps the lateral articular area of the tibia and forms a footplate for the distal end of the fibula.
The tibia and fibula assigned to Pisanosaurus are elongate. The proximal end of the tibia is expanded rostrocaudally, there are two distinct caudal condylids and a prominent rostral cnemial crest, and the shaft is twisted axially 90° (Bonaparte 1976). Distally the medial and lateral malleoli are distinct; however, the astragalar articular surface is recessed laterally to accommodate the stout, short ascending process, and the lateral articular area is narrow and caudally offset. The fibula is much more slender than the tibia and moderately expanded proximally but less so distally. The distal end of the tibia is unusually narrow rather than transversely flared as in all other ornithischians. The ankle joint is mesotarsal (contra Bonaparte 1976), and the astragalus has a short, blunt ascending process that is lodged in a recess on the rostrolateral surface of the medial articular surface. The calcaneum is represented by a nubbin of bone that lacks anatomical information.
The pes is about as long as the tibia in Lesothosaurus and about 60% as long in Pisanosaurus. The pedal phalangeal formula was probably 2-3-4-5-0 (Thulborn 1972). In Lesothosaurus metatarsal I is unusual in that it is small, thin, and splintlike; its well-formed distal condyles are set at an angle to the remaining metatarsals, and it bears a slender proximal phalanx and a slightly decurved, pointed ungual. The other three metatarsals are elongate, with metatarsal III being longest (as in Pisanosaurus), while II and IV are subequal. In Lesothosaurus the proximal articular surfaces are flat and the phalangeal articulations are weakly bicondylar in metatarsal III but simply convex in metatarsals II and IV. Metatarsals II and III are laterally compressed proximally and closely appressed throughout most of their length. Metatarsal IV is transversely expanded proximally, and its shaft diverges from the other metatarsals. The metatarsus exhibits a weak transverse arch in which the plantar surface is concave. In Pisanosaurus the base of metatarsal III overlaps that of metatarsal IV. The slender, elongate phalanges have well-developed collateral ligament pits. The unguals are slightly decurved but essentially conical with a flat plantar surface and display clear ungual grooves laterally and medially.
Systematics and Evolution
Lesothosaurus was once regarded as one of the earliest representatives of Ornithopoda and therefore a member of the so-called primitive stock from which all ornithischians could have been derived (Thulborn 1971b; Galton 1972, 1978; Chatterjee 1984). In this context other forms—Fabrosaurus, Scutellosaurus, Tawasaurus, Alocodon, Trimucrodon, Echinodon, Technosaurus, Nanosaurus, and Xiaosaurus—were grouped with Lesothosaurus as Fabrosauridae (Galton 1978 et seq.). This latter position has also been advocated by Peng (1997) following the description of Agilisaurus (Peng 1990, 1992), an ornithopod from the Middle Jurassic of China; however, the characters listed by Peng (1997) to diagnose Fabrosauridae are ornithischian symplesiomorphies, except for the groove found on the dorsal edge of the ischium. (The latter taxon is considered further in chapter 18.)
With the first cladistic analyses of these forms (Gauthier 1986; Sereno 1986, 1991b), Fabrosauridae was disbanded as a monophyletic taxon and Lesothosaurus was identified, not as an ornithopod, but as the most basal of all known ornithischians. Other taxa, such as Scutellosaurus (chapter 15; Gauthier 1986; Sereno 1986) and Echinodon (chapter 18; Norman and Barrett 2003), have been referred to higher ornithischian taxa. The majority of the remaining putative primitive taxa can be classified solely as Ornithischia incertae sedis or relegated to the status of nomina dubia (table 14.1). The two most important taxa that remain to be considered after this process of reassignment are Lesothosaurus diagnosticus and Pisanosaurus mertii.
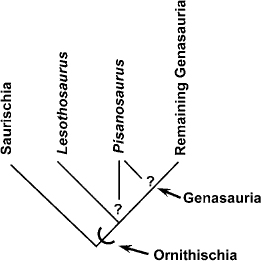
FIGURE 14.7. Cladogram of Ornithischia, emphasizing the relationships of Lesothosaurus diagnosticus and Pisanosaurus mertii.
In order to evaluate the relationships of Lesothosaurus and Pisanosaurus within Ornithischia, outgroup comparisons were made successively with Genasauria and Saurischia. The possibility that the sister taxon of Ornithischia is Prosauropoda or Segnosauria and that in combination they constitute Phytodinosauria, although repeatedly discussed (Bakker and Galton 1974; Bonaparte 1976; Cooper 1981a, 1985; Sereno 1984; Paul 1984a, 1984b; Bakker 1986), has been little supported. As a result, Saurischia is by consensus regarded as monophyletic (Gauthier 1986; Benton 1990a; Sereno 1997).
Ornithischia is a well-supported monophyletic taxon, diagnosed using the characters listed in the “Definition and Diagnosis” section above. Among taxa recognized as basal ornithischians, Lesothosaurus diagnosticus is excluded from a higher position within Ornithischia because it retains a large external mandibular fenestra. It is regarded as the sister taxon to genasaurian ornithischians (fig. 14.7; Gauthier 1986; Sereno 1999a). Lesothosaurus has been diagnosed (Sereno 1991b) using the following apomorphies: a rostral premaxillary foramen, a fingerlike process on the maxilla that overlaps the rostral lacrimal process, a longitudinal groove on the dorsal surface of the ischial shaft, broad lateral exposure of the brevis shelf on the ilium, and pedal digit I supported by a splintlike metatarsal. Lesothosaurus also possesses a prominent crested obturator process on the ischium, and metatarsal V either was absent or strongly reduced.
The position of Pisanosaurus mertii is extremely problematic. Originally regarded as an ornithopod, or more specifically as a heterodontosaurid (Casamiquela 1967b; Thulborn 1971a; Galton 1972, 1986a; Charig and Crompton 1974; Bonaparte 1976; Cooper 1981a; Weishampel and Weishampel 1983; Weishampel 1984c; Crompton and Attridge 1986), more recently it has been considered the most basal of all known ornithischians (Weishampel and Witmer 1990a; Sereno 1991b). The main problems with Pisanosaurus are its poor preservation and the loss of information as to how the original material was associated. That it is now represented by at least two individuals of different size and perhaps two different taxa (Sereno 1991b) makes the need for careful interpretation of the association of Pisanosaurus material obvious.
Several ornithischian synapomorphies are found in the jaws of Pisanosaurus: the separation of the crown and root of the teeth by a neck, maximal tooth size near the middle of the tooth row, and the rostral portion of the coronoid process formed by dentary.
Other features—pronounced buccal emargination of the maxillary and dentary tooth rows and the development of extensive, apparently confluent wear facets between blades formed by the maxillary and dentary teeth—are seen in cerapodans. On the basis of the few characteristics seen in the cranial material of Pisanosaurus, this taxon may well be referable to Genasauria. The associated pelvic impression is difficult to interpret, as indicated by the subtly different interpretations of Bonaparte (1976) and Sereno (1991b); we regard this material as moot on the matter of whether it indicates a propubic or an opisthopubic condition. Thus, the sine qua non of Ornithischia is not known to be present with certainty in Pisanosaurus. Furthermore, Sereno (1991b) indicated that only two of the vertebral impressions, rather than five as originally thought (Casamiquela 1967b; Bonaparte 1976), are sacrals, which renders this feature irrelevant as to ornithischian affinities. The lack of distal flaring of the associated tibia is also plesiomorphic with respect to Ornithischia (and indeed Dinosauria). Likewise, the apex of the ascending process of the astragalus is laterally situated near the articulation with the calcaneum in Lesothosaurus, basal thyreophorans, prosauropods, basal theropods, and basal dinosauromorphs. Autapomorphies of Pisanosaurus include the recessing of the tibia to receive the ascending process of the astragalus.
Given these features, we suggest the following. If the jaws, the ambiguous pelvic impression, the tibia, and the proximal tarsals are considered to belong to one individual, then the most parsimonious character distribution places it as a genasaur. The narrowness of the distal tibia and the position of the ascending process of the astragalus would thus represent apomorphies of this taxon. An ornithopod position is consistent with earlier claims that Pisanosaurus is a heterodontosaurid. If, however, we treat the elements individually, the jaw material can be referred without question to Cerapoda. On the other hand, the hindlimb elements suggest an animal of nondinosaurian affinity. Clearly, we need better-preserved material referable to P. mertii before a resolution of its systematic position can be achieved.
Chatterjee (1984) referred Technosaurus smalli to Fabrosauridae, a taxon that has now been abandoned as paraphyletic. Technosaurus is a chimaera of prosauropod and ornithischian material (Sereno 1991b). A section of dentary (fig. 14.2C) bears emarginated and triangular teeth covered with an even layer of enamel; crowns imbricate, tooth size increases toward the caudal of the row, and the crowns are separated from the roots by a distinct neck. These are plesiomorphic ornithischian characters. Based on these characters, the dentary of Technosaurus is ornithischian, but its position within the clade cannot be resolved.
Several other taxa thought to be primitive ornithischians have also been erected. One of these, Fabrosaurus australis, is based on a dentary with three in situ teeth (Ginsburg 1964). Thulborn (1970a, 1972) referred additional material to F. australis, but Galton (1978) noted that these specimens were distinct from Ginsburg's original dentary and erected Lesothosaurus diagnosticus on their basis. In his study of Lesothosaurus Sereno (1991b) concluded that Fabrosaurus has no autapomorphies and should be regarded as Ornithischia indet. and a nomen dubium (see also Charig and Crompton 1974). In striking contrast, Thulborn (1992) argued that F. australis possesses a number of unique features (all relating to dental morphology) and that material referred to Lesothosaurus diagnosticus must be relegated in nomenclature as a junior subjective synonym of the former. However, the features used to diagnose F. australis are not particularly compelling. Consequently, it is here considered to be a nomen dubium.
A large number of taxa thought to be basal ornithischians have been established on the basis of isolated teeth. These include Alocodon kuehnei, Trimucrodon cuneatus, Nanosaurus agilis, Xiaosaurus dashanpensis, Tawasaurus minor, Revueltosaurus callenderi, Galtonia gibbidens, Pekinosaurus olseni, Tecovasaurus murryi, and Lucianosaurus wildi (Thulborn 1973a; Galton 1978; Young 1982a; Dong and Tang 1983; Hunt 1989; Hunt and Lucas 1994). Some of these species have one or a few ornithischian apomorphies, thus bringing them into the clade, but otherwise lack additional features that would place them more precisely within the clade. Some teeth have sufficient peculiarity to their crown shape to offer potential autapomorphic features (A. kuehnei, T. cuneatus, possibly N. agilis), but reevaluation of their status must wait for new and better-preserved material. Others, such as those belonging to X. dashanpensis and R. callenderi, do not. The former have been considered Ornithischia incertae sedis and the latter Ornithischia indet. as nomina dubia (Weishampel and Witmer 1990a; Sereno 1991b). G. gibbidens is diagnosed with reference to F. australis, but the former is based on a premaxillary tooth, and premaxillary teeth are unknown in the latter. All that can be said at this point is that Galtonia is Ornithischia indet. as a nomen dubium. Similarly, P. olseni, T. murryi, and L. wildi are considered Ornithischia indet. as nomina dubia because they possess ornithischian synapomorphies but lack individual autapomorphies. Finally, T. minor is not an ornithischian at all but referable to Prosauropoda indet. (Sereno 1991b).
Paleoecology and Biology
The fossil record of Lesothosaurus diagnosticus is limited to the Early Jurassic of South Africa. If we regard Pisanosaurus mertii and Technosaurus smalli as basal ornithischians, then the distribution of these forms extends from the Late Triassic through the Early Jurassic and includes not only southern Africa but also southern South America and western North America. Unfortunately, it is difficult to assess the probable source area for Ornithischia on the basis of these data. Even in comparison with the primitive distribution of Thyreophora (unresolved), Cerapoda (possibly Asia), and Saurischia (South America? Africa?), resolution of all of these geographic distributions does not yield a single, unambiguous region of the world as an ornithischian source area, even at the continental level.
Because of the rarity of fossils and the often imperfect nature of the material referred to Lesothosaurus, Pisanosaurus, and Technosaurus, little can be said about their paleoecology. Local environments from which these animals derive represent chiefly semiarid habitats. As herbivores, Lesothosaurus, Pisanosaurus, and Technosaurus may have been active foragers of ground cover and shrubby vegetation.
A keratinous rhamphotheca was present in Lesothosaurus and probably covered the entire surface of the predentary. Its occlusal margin is pointed at the tip and sharp-edged distally, not unlike the stocky lower beak of a nut-eating bird (seen in some species of finch). By contrast, the premaxilla is largely dentigerous and has a restricted keratinous beak on the tip of the upper jaw (fig. 14.1). It is unclear whether the upper rhamphotheca extended distally so as to envelope some or all of the premaxillary teeth or whether the keratinous and dentigerous portions were functionally separated. There is no modern functional analogue for the composite keratinous, tooth-lined beak, and thus its operation is enigmatic. No wear facets have been identified on lesothosaur premaxillary teeth; it may well be that the premaxillary teeth were embedded in, and provided mechanical support for, the keratinous sheath of the upper beak.
Norman and Weishampel (1991) suggested that Lesothosaurus may have used its narrow, pointed beak (fig. 14.1) to feed selectively on soft, less fibrous plant material (fruiting bodies, shoots), which was more easily processed and assimilated than the more heavily lignified parts of plants. Barrett (2000a) has argued that some dinosaurian herbivores (basal ornithischians and sauropodomorphs) exhibit dental morphologies that strongly resemble those seen in living reptiles (e.g., iguanines) that are omnivorous rather than exclusively herbivorous. An omnivorous diet, which offers greater feeding flexibility, may well have been appropriate for small, highly energetic (active, fast-running) forms such as Lesothosaurus.
Thulborn (1971a) and Weishampel (1984a) have suggested that Lesothosaurus, like other primitive ornithischians, relied on simple adduction of the lower jaws to produce a vertical or near-vertical tooth-tooth shearing motion between bilaterally occluding maxillary and dentary teeth. In contrast, Crompton and Attridge (1986) and Sereno (1991b, 1997, 1999a) have described longitudinal lower-jaw rotation (controlled passively by the predentary-dentary articulation) as an important component of lesothosaur jaw action. This alternative mechanism is not corroborated by the structure of wear facets in the dentition, which are uniformly high-angle and planar (Barrett 1998).
Based on limb and trunk proportions and the general gracile nature of their skeleton (fig. 14.4), lesothosaurs (and possibly the other taxa under consideration here) were probably agile, cursorial bipeds (Coombs 1978a; Thulborn 1982). A quadrupedal stance was probably only relevant during foraging or when standing still. In Lesothosaurus the forelimbs are short and the distal portions of the forelimb are reduced. It is clear from its general structure (fig. 14.6B) that the manus was multifunctional: the bases of the metacarpals imbricate and could therefore be collapsed to allow the digits to be bunched together for use in simple scooping or grasping movements. Such functions may have been aided by the fact that the first digit may have been capable of being partially opposed to the palm. Alternatively the digits could be spread widely if the manus were placed palm outward or downward (on the ground) for temporary support. The reduction of the fifth digit to form a short, oblique, perhaps prop-like digit may well indicate that hand-mediated body support was an important component of the lifestyle (possibly associated with a specialist browsing feeding regime hinted at above).
The femur (fig. 14.6A) was probably held in a slightly abducted position to facilitate a parasagittal gait despite the presence of a bulky ventral, midline gut (Norman and Weishampel 1991). Lever-arm mechanics suggest that femoral retraction, mediated by iliac and caudal musculature, was both powerful and fast. The attitude of the dorsal, sacral, and proximal caudal axial skeleton was largely horizontal, dorsally arched, and tensioned by the epaxial ossified tendons. In contrast, the cervical vertebrae were more flexibly articulated and capable of adopting a sinuous curve; the distal caudals may also have been highly flexible to assist balance.