EIGHTEEN

Basal Ornithopoda
Ornithopoda (table 18.1) is the name first used by Marsh (1881b) to designate bipedal, unarmored, herbivorous dinosaurs, some of which (hadrosaurids) had complex dentitions. Ornithopods have a stratigraphic range spanning the Lower Jurassic–Upper Cretaceous. The systematic position of the clade has been clarified and subsequently refined within a phylogenetic framework (Sereno 1986, 1998; Weishampel 1990a; Weishampel and Heinrich 1992). Use of the terms Ornithopoda and Euornithopoda follows that established by Weishampel (1990a).
The ornithopod clade includes the small heterodontosaurids from the Early Jurassic and euornithopods, which range widely in body size and are known from the Middle Jurassic to the end of the Cretaceous. Monophyly of Euornithopoda is reasonably well supported and was at one time considered by some to consist of two monophyletic taxa: Hypsilophodontidae and Iguanodontia (Milner and Norman 1984; Sereno 1984, 1986, 1998; Sues and Norman 1990; Weishampel and Heinrich 1992). The former clade, however, has been the subject of reanalysis in recent years following the discovery of new taxa (Scheetz 1998, 1999; Winkler et al. 1998). These latter contributions suggest that Hypsilophodontidae sensu Galton (1974a et seq.) may be paraphyletic.
This chapter examines the anatomy, phylogeny, and paleobiology of noniguanodontian Ornithopoda (from here on called basal ornithopods). Among these is Gasparinisaura cincosaltensis Coria and Salgado, 1996b. Although originally described as a basal iguanodontian from the Late Cretaceous of Argentina, its anatomy is more consistent with that of the basal ornithopods considered in this chapter, and this is borne out in the systematic analysis. Basal ornithopod taxa can be arranged in successively closer relationships to Iguanodontia through the serial acquisition of characters.
Many of the ornithopods described here, particularly heterodontosaurids, are among the smallest known ornithischians (adult length of 1–2 m, although there are reports of larger individuals) and the stratigraphically oldest (Hettangian–Sinemurian). Basal members of Euornithopoda have a more extensive record through the remainder of the Mesozoic (Bathonian–Maastrichtian). All well-preserved heterodontosaurid material comes from South Africa, although potentially additional material has been reported from Arizona, United States; Dorset, England; and Yunnan, China. Basal euornithopods have been collected from Asia, Australia, Europe, South America, North America, and Antarctica. Several new forms, from Arizona and Texas, United States (Crompton and Attridge 1986; Winkler et al. 1988, 1998), and the Antarctica Peninsula (Hooker et al. 1991) have yet to be fully described.
The skulls of these dinosaurs are modified principally in association with the species' presumed herbivorous habits (depressed jaw joint, robust and closely packed teeth, extensive tooth wear). Locomotion is typically bipedal, and hindlimb anatomy and proportions indicate that many of these basal forms were cursorial (Coombs 1978a). These ornithopods have also proved to be the source of much new and intriguing information on growth, life histories, and soft anatomy. For example, Horner and Weishampel (1988, 1996), Long and McNamara (1995), and Scheetz (1999) have used growth-series information from several ornithopod species to examine the impact of heterochrony on the dynamics of dinosaurian evolution. Much more controversially, Fisher et al. (2000) claim putative cardiac remains within the articulated skeleton that they referred to Thescelosaurus.
Definition and Diagnosis
Ornithopoda is a stem-based taxon defined as all cerapodans closer to Edmontosaurus than to Triceratops. It is diagnosed by the following features: ventral offset of the premaxillary occlusal margin relative to the maxillary tooth row, a crescentic paroccipital process, jaw articulation offset ventral to the maxillary tooth row, premaxilla contact with the lacrimal on the external surface of the snout.
Anatomy
Skull and Mandible
Many skulls of basal ornithopods have been discovered. Among heterodontosaurids, two complete skulls and isolated cranial elements have been found for Heterodontosaurus tucki (figs. 18.1A, B, 18.2D; Crompton and Charig 1962), and somewhat less complete material has been referred to Abrictosaurus consors (Thulborn 1974; see also Charig and Crompton 1974; Hopson 1980b; Crompton and Attridge 1986). Partial maxillae and a fragmentary dentary, all bearing teeth, are known for Lycorhinus angustidens (fig. 18.2A, C; Haughton 1924; see also Gow 1975a, 1990b; Hopson 1975a). Among basal euornithopods, the skull is probably best known from several individuals of Hypsilophodon foxii at different ontogenetic stages (fig. 18.3A; Huxley 1869b; Galton 1974a), although several other taxa are represented by either complete or at least partial skulls. Those taxa for which relatively complete skulls have been found include Agilisaurus louderbacki (fig. 18.4A; He and Cai 1983; Peng 1990, 1992, 1997), Bugenasaura infernalis (fig. 18.4D; Morris 1976; Fisher et al. 2000), Thescelosaurus neglectus (fig. 18.4C; Gilmore 1915b; Galton 1995), Orodromeus makelai (fig. 18.4B; Horner and Weishampel 1988; Scheetz 1999), and Gasparinisaura cincosaltensis (fig. 18.5A; Coria and Salgado 1996b). In addition, a relatively complete, but highly deformed skull is known for Parksosaurus warreni (fig. 18.5C; Parks 1926b; Galton 1973c, 1995). Othnielia rex is known from incomplete skull remains, but a nearly complete skull is reported to be in private hands (Galton 1983c). Other basal euornithopods are known with incomplete skull material, including Leaellynasaura amicagraphica and Atlascopcosaurus loadsi, from Australia (Rich and Rich 1989; Rich and Vickers-Rich 1999), Zephyrosaurus schaffi (fig. 18.5B; Sues 1980b) and Drinker nisti from North America (Bakker et al. 1990), and “Gongbusaurus” wucaiwanensis, and Yandusaurus hongheensis from Asia (He 1979; Dong and Tang 1983; Dong 1989).
TABLE 18.1
Basal Ornithopoda

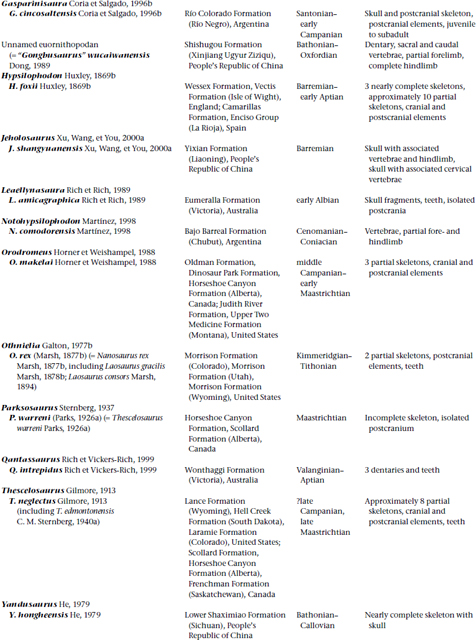
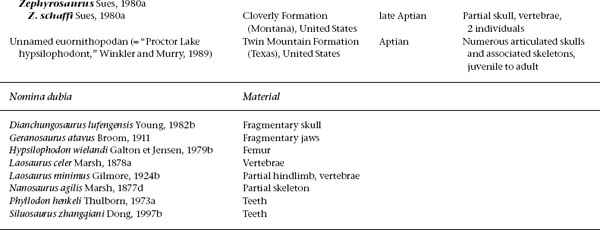
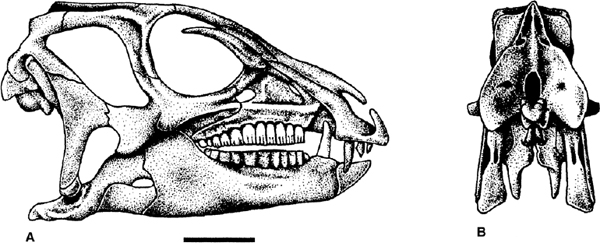
FIGURE 18.1. A, Heterodontosaurus tucki, skull in right lateral view; B, occipital view of the same skull. Scale = 2 cm. (From Weishampel and Witmer 1990b.)
In all cases, the skulls of these taxa are comparatively small and the facial skeletons moderately short and high. Where known, the skull is not much modified from the cranial proportions observed in juveniles. In immature skulls, however, the snout is truncated with an abrupt downward slope (viz. the juvenile material of Abrictosaurus consors and Agilisaurus louderbacki). In some of the geologically youngest taxa, Parksosaurus warreni, Thescelosaurus neglectus, and Bugenasaura infernalis among them, the facial skeleton is long, forming a rather low, rostrally extended profile.
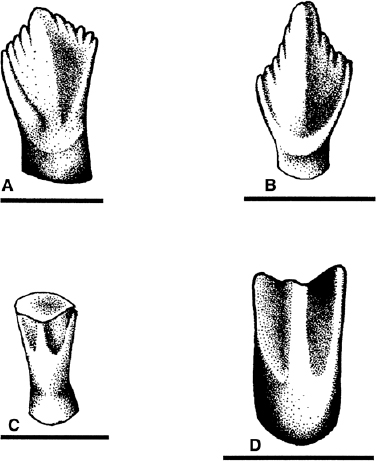
FIGURE 18.2. A, Maxillary tooth of Lycorhinus angustidens; B, dentary tooth of Abrictosaurus consors; C, dentary tooth of Lycorhinus angustidens; D, worn dentary tooth of Heterodontosaurus tucki. Scale = 5 mm. (A after Gow 1975a; B after Thulborn 1974; C after Hopson 1975a; D from Weishampel and Witmer 1990b.)
The orbit, infratemporal fenestra, and antorbital fossa are large in proportion to the overall size of the skull (figs. 18.1, 18.3–18.5) but the external naris is small, never more than 15% of the skull length. The orbit is either circular or slightly oval and encloses a ring of sclerotic ossicles. The infratemporal fenestra ranges from high and narrow to obliquely oval in outline and, in Thescelosaurus neglectus, Hypsilophodon foxii, Orodromeus makelai, and Agilisaurus louderbacki (possibly also Parksosaurus warreni), it is bordered by the squamosal and postorbital dorsally and the jugal, quadratojugal, and quadrate ventrally. Gasparinisaura cincosaltensis has approximately the same condition as the aforementioned species, but the quadrate is reported (Coria and Salgado 1996b) to be excluded from the caudal margin by the conjoined quadratojugal and squamosal. In Heterodontosaurus tucki (fig. 18.1A) and Agilisaurus louderbacki (fig. 18.4A), however, the infratemporal fenestra is wide and high (rhomboid in H. tucki), bordered by the postorbital and squamosal dorsally and the quadratojugal and jugal ventrally.
The antorbital fossa forms a pocketlike recess in the lateral wall of the maxilla. Its rostral margin has the form of a rounded ridge that runs parallel to the suture between the premaxilla and maxilla. The dorsal and caudal margins of the fossa are formed by the lacrimal and these are sharp edged where this margin overhangs the recess formed by the fossa. In Heterodontosaurus the caudal margin of the fossa extends as a notch onto the maxillary process of the jugal (fig. 18.1A). Largest in H. tucki and smallest in Parksosaurus warreni, the fenestra that bounds the antorbital fossa is subtriangular in H. tucki, Hypsilophodon foxii, “Agilisaurus” multidens, and A. louderbacki; subcircular in Orodromeus makelai, Gasparinisaura cincosaltensis, Thescelosaurus neglectus, and Bugenasaura infernalis; and crescentic in P. warreni. Bounded dorsally and caudally by the lacrimal, the inner wall of the antorbital fossa of Heterodontosaurus and H. foxii displays two subsidiary foramina: the principal opening is a teardrop-shaped foramen at the back of the fossa adjacent to the contact between the maxilla and jugal and a smaller rostral opening lying beneath the contact of the maxilla with the lateral process of the premaxilla.
The rostral region of the premaxilla is edentulous for a distance amounting to at least the dimension of one tooth position. Its outer surface is rugose and probably supported a small, keratinous rhamphotheca in life. In Zephyrosaurus schaffi, the premaxillae are fused with each other, but in other taxa they remain separate, although obviously tightly connected with strong sutural ligaments (Sues 1980b; Weishampel 1984c). Directly behind the beak margin the premaxilla of adult Abrictosaurus consors and Heterodontosaurus tucki accommodates three teeth, the last of which is enlarged as a caniniform. In the juvenile specimen of A. consors there are two premaxillary teeth, but no indication of a caniniform. In addition, in these two taxa, the caudal edge of the premaxilla bears a pocketlike recess for the caniniform tooth of the dentary. In Agilisaurus louderbacki, Hypsilophodon foxii, Zephyrosaurus schaffi, and Thescelosaurus neglectus, Orodromeus makelai, Bugenasaura infernalis, there are five premaxillary teeth. Premaxillary teeth have also been reported to be in cranial material attributed to Othnielia rex. However, none are caniniform and there is no diastema or pocketlike recess to accommodate a caniniform dentary tooth. Premaxillary teeth are reported in Drinker nisti (Bakker et al. 1990).
Primitively in ornithopods the ventral margin of the premaxilla is offset to a level below that of the maxilla (Sereno 1986). The dorsal process of the premaxilla, where it forms the upper margin of the external naris, is slender and smoothly arched. Several small foramina are visible within the narial fossa, along the rostral margin of the external naris; these are seen particularly well in Zephyrosaurus schaffi (Sues 1980b) and Hypsilophodon foxii (Galton 1974a). One foramen communicates with the palatal surface of the premaxilla rostral to the first tooth position and, in H. foxii at least (Galton 1974a), opens into a cavity within the body of the premaxilla. The caudal end of the body of the premaxilla is jointed against the front edge of the maxilla; this joint is especially distinctive in Zephyrosaurus schaffi (fig. 18.5; Sues 1980b) and Orodromeus makelai (fig. 18.4B; Scheetz 1999), and this may reflect the initial development of a secondary (i.e., rostrolateral) premaxillary-maxillary articulation. The narrow, elongate lateral process of the premaxilla of Heterodontosaurus tucki extends to the junction between the prefrontal and lacrimal (fig. 18.1A), removing external contact between the nasal and maxilla (unfortunately this region is not known in other heterodontosaurids). In contrast, the lateral process of the premaxilla is short and does not completely separate the maxilla and nasal on the side of the face in most basal euornithopods: Parksosaurus warreni, Thescelosaurus neglectus, Hypsilophodon foxii, and Agilisaurus louderbacki. The sagittal premaxillary sutural surface includes a distinct slot to accommodate the rostral processes of the maxillae, as well as the rostral process of the fused vomers (cf. Hypsilophodon, Galton 1974a).
The nasal, in turn, roofs the nasal cavity and forms the dorsal margin of the external naris in all the taxa under consideration here. A generally narrow element widening caudally as it approaches the skull roof, each nasal articulates with the rostral end of the lacrimal via a ridge-and-groove arrangement; its caudolateral edge bears a groove for reception of the prefrontal and it overlaps the frontal caudally.
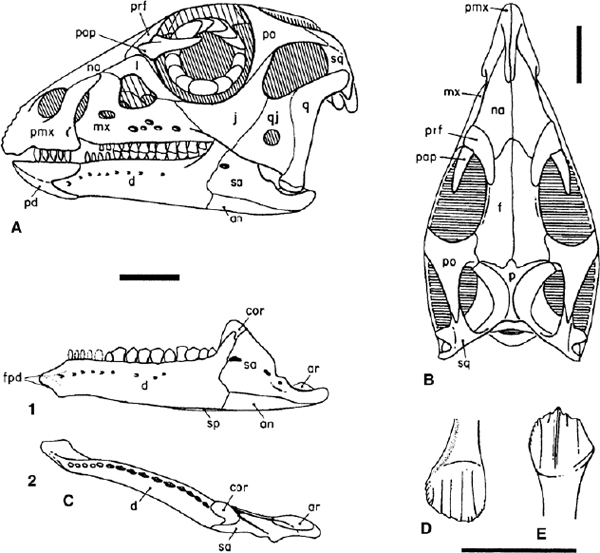
FIGURE 18.3. Skull and teeth of Hypsilophodon foxii: A, B, skull in left lateral and dorsal views; C, left lower jaw (without predentary) in lateral, 1, and dorsal, 2, views; D, maxillary crown in buccal view; E, dentary crown in buccal view. Scale = 2 cm (A, B, C), 1 cm (D, E). (After Galton 1974a.)
In ornithopods the rostrodorsal margin of the maxilla contacts the lateral process of the premaxilla along a shallow, convex articular surface. At its most rostral end the maxilla also backs the steplike gap between the premaxillary and maxillary dentitions. This structure is exaggerated in Heterodontosaurus tucki (fig. 18.1A) and Abrictosaurus consors as a result of the presence of the recess for the enlarged dentary caniniform.
Dorsally, the maxilla has a short contact with the nasal (except in Heterodontosaurus tucki) and overlaps the lacrimal both medially (where it contributes to the inner wall of the antorbital fossa) and laterally. Caudodorsally, the lateral maxillary surface forms a prominent combined suture for the lacrimal and jugal. Ventral to this position, the tooth row is inset from the side of the skull and creates an overhang; the tooth row of the dentary is similarly inset. The longitudinal facial recess formed by the inset maxillary and dentary dentition strongly hints at the presence of some form of fleshy cheek. A number of variably sized neurovascular foramina open onto the lower lateral portion of the maxillary recess, at least in Lycorhinus angustidens, Abrictosaurus consors, Zephyrosaurus schaffi, Hypsilophodon foxii, Gasparinisaura cincosaltensis, Atlascopcosaurus loadsi, “Agilisaurus” multidens, A. louderbacki, Thescelosaurus neglectus, and possibly in Parksosaurus warreni. The medial and dorsal surface of the caudal end of the maxilla shows a prominently scarred sutural surface for the palatine.
In heterodontosaurids, 11maxillary tooth positions are known in Heterodontosaurus tucki, 12 in Abrictosaurus consors, and at least 12 in Lycorhinus angustidens. Among basal euornithopods, Hypsilophodon foxii has 11or 12maxillary tooth positions, which is similar to the condition in Atlascopcosaurus loadsi. There are 12 or 13 positions in Leaellynasaura amicagraphica and Gasparinisaura cincosaltensis, 14 or 15 in Zephyrosaurus schaffi and Agilisaurus louderbacki, perhaps 18 in Parksosaurus warreni, and as many as 20in “A.” multidens, Thescelosaurus neglectus, and Bugenasaura infernalis. In Orodromeus makelai, the tooth number is thought to range ontogenetically from 10 to 13 (Scheetz 1999). In all cases, the tooth row is either straight or slightly laterally concave in ventral view.
The jugal of both heterodontosaurids and basal euornithopods is straplike and links the maxilla to the suspensorium, closing off the orbit and infratemporal fenestra ventrally and enclosing the adductor chamber. Rostrally, the body of the jugal forms a long, overlapping suture with the dorsolateral surface of the maxilla. In Heterodontosaurus (fig. 18.1A) the external surface of this process is excavated by a caudal extension of the antorbital fossa; the antorbital fossa does not extend on to the jugal in other taxa. A stout jugal boss projects laterally from the external surface of the jugal in Heterodontosaurus tucki, Zephyrosaurus schaffi, and Orodromeus makelai (figs 18.1A, 18.4B, 18.5B). In Heterodontosaurus tucki, the caudal process of the jugal forms an interdigitate suture with the quadratojugal, whereas in euornithopods the joint is an apparently simple, overlapping arrangement. The caudal process is especially deep and does not contact the quadrate in basal euornithopods. The ventral margin and sides of this caudal process are generally striated, presumably reflecting the attachment of connective tissues associated with the cheeks. Ventral to the jugal-quadratojugal articulation, a prominent ventral extension of the jugal descends across the surangular portion of the coronoid process in H. tucki, the only species for which this condition is known.
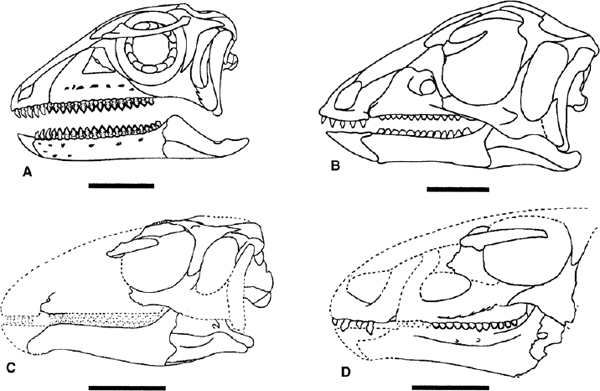
FIGURE 18.4. Skulls of various basal ornithopods: A, Agilisaurus louderbacki; B, Orodromeus makelai; C, Thescelosaurus neglectus; D, Bugenasaura infernalis. Scale = 20 mm (A, B), 5 cm (C, D). (A after Peng 1997; B after Scheetz 1999, copyright R. Scheetz; C, D after Galton 1995.)
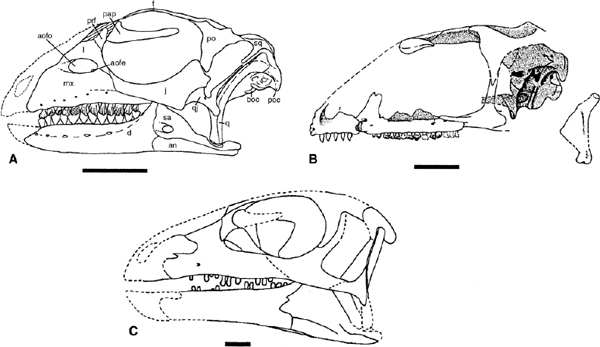
FIGURE 18.5. Skulls of various basal ornithopods: A, Gasparinisaura cincosaltensis; B, Zephyrosaurus schaffi; C, Parksosaurus warreni. Scale bar = 2 cm. (A after Coria and Salgado 1996b; B after Sues 1980b; C after Galton 1995.)
In Heterodontosaurus tucki and perhaps Abrictosaurus consors, the quadratojugal is large, practically coextensive with, and lying lateral to, the quadrate. The dorsal process is sutured along a short, interdigitate contact, with the prequadratic process of the squamosal. In virtually all taxa, the quadratojugal forms the caudal half of the ventral margin of the infratemporal fenestra (it is excluded from the caudal margin of the fenestra by the extensive caudal process of the jugal in Agilisaurus louderbacki). It is pierced by a prominent foramen in Hypsilophodon foxii, similar to the condition in the basal iguanodontian Tenontosaurus tilletti. The same feature may also apply to Gasparinisaura cincosaltensis (Coria and Salgado 1996b; D. Suzuki, pers. comm.), although the quadratojugal is highly eroded in this region.
The lacrimal forms the caudal and dorsal margins to the antorbital fenestra. The thickened ventral margin rests on the dorsal end of the jugal in a short, simple articulation and the orbital margin bears the opening for the nasolacrimal canal.
In all cases for which the palpebral is known, it tapers as it curves across the dorsal region of the orbit. It is short (less than 70% of the orbit diameter) in Hypsilophodon foxii, Zephyrosaurus schaffi, Thescelosaurus neglectus, Orodromeus makelai, and Agilisaurus louderbacki. The palpebral is much longer, often reaching the orbital margin of the postorbital, in Bugenasaura infernalis, Agilisaurus louderbacki, Gasparinisaura cincosaltensis, and Heterodontosaurus tucki. The base of the palpebral contacts the orbital margin at the prefrontal-lacrimal suture in Heterodontosaurus tucki, Bugenasaura infernalis, Agilisaurus louderbacki, Orodromeus makelai, and possibly Thescelosaurus neglectus, and Parksosaurus warreni. Alternatively, it contacts the prefrontal alone (in Gasparinisaura cincosaltensis, Hypsilophodon foxii, and possibly in Zephyrosaurus schaffi and “Agilisaurus” multidens).
The prefrontal forms a jointed suture with the lacrimal, while medially, it contacts both the nasal and frontal. Each frontal has the approximate shape of a somewhat compressed pentangle in dorsal view, indented laterally where it forms the concave, thin, and rugose margin of the orbit. Laterally, it contributes to the dorsal margin of the orbit, and caudally, it forms a moderately interdigitate transverse suture with the parietal. The interfrontal suture runs straight along the midline of the skull. Laterally, the frontal forms an undulating suture with the postorbital.
The short medial process of the postorbital meets the frontal and a small portion of the parietal along a coarsely interdigitate suture. The caudal process overlaps, and interfingers with, the squamosal, forming the upper temporal bar. The central region of the postorbital adjacent to the orbital margin is often expanded and rugose, possibly reflecting a cartilaginous or connective tissue attachment area for the palpebral. In Heterodontosaurus tucki, the ventral process of the postorbital meets the jugal along an unusually stable, interdigitate suture to form the postorbital bar. In contrast, all other taxa maintain an oblique, and apparently weak, scarf joint between these two bones. The lateral surface of the postorbital in H. tucki is marked by a prominent dorsoventrally oriented ridge that may reflect the attachment of an unusually well-developed M. adductor mandibulae externus pars superficialis; in all others, the external surface of the postorbital is unsculpted.
The body of the squamosal is dominated by a caudolaterally and slightly ventrally facing recess that receives the quadrate head. In all taxa except Heterodontosaurus tucki, the lateral face of the squamosal, directly above and rostral to the quadrate cotylus, bears the scar for M. adductor mandibulae superficialis pars externus. The short and robust rostral process of the squamosal contacts the postorbital in an interdigitate joint in H. tucki and an intricate scarf joint in other taxa. The prequadratic process is robust and meets the dorsal process of the quadratojugal to exclude the quadrate from the margin of the infratemporal fenestra in H. tucki, “Agilisaurus” multidens, Gasparinisaura cincosaltensis, and possibly Abrictosaurus consors. In A. louderbacki, Thescelosaurus neglectus, Orodromeus makelai, and Hypsilophodon foxii, however, the prequadratic process is shorter, terminating well above the dorsal tip of the quadratojugal. Caudally, the squamosal contributes only a short postquadratic process to the lateral aspect of the paroccipital process.
From its articulation with the squamosal, the quadrate arches forward and downward to a transversely expanded mandibular condyle. The dorsal head is small and ovoid in shape, often supported caudally by a buttress that rests against the adjacent paroccipital process. The medial side of the quadrate shaft bears a thin, vertical sheet of bone that forms an extensive overlap with the equally winglike quadrate ramus of the pterygoid. The lateral wall of the quadrate shaft is excavated to form an extensive sutural surface for the quadratojugal. In Heterodontosaurus tucki, a small paraquadratic foramen is found on the caudal surface of the midsection of the quadrate shaft. The mandibular condyle is, to a variable degree, divided into a prominent lateral and smaller medial portion.
Returning to the top of the skull, the fused parietals form a saddle-shaped plate covering the central portion of the skull roof and divided in the midline by a sagittal crest. In dorsal view, a median process may be visible (fig. 18.3B; Hypsilophodon, Galton 1974a) fitting between frontals, while lateral spurs contact the postorbitals. The caudal margin is incised but covers much of the supraoccipital.
The braincase is known in Heterodontosaurus tucki, Hypsilophodon foxii (Galton 1974a), Zephyrosaurus schaffi (Sues 1980b), Orodromeus makelai (Scheetz, 1999), and Thescelosaurus neglectus (Galton 1997b); many other taxa offer additional information. Showing a general sauropsid configuration, the prootic, opisthotic, and exoccipital form the lateral walls, the basioccipital, basisphenoid, and parasphenoid make up the braincase floor, and the parietals and supraoccipital form the roof. The rostrolateral extremity of the laterosphenoid is expanded into a transverse condyle that fits into a well-delimited depression on the ventral aspect of the frontal and postorbital. The parasphenoid and orbitosphenoid are found across the back of the orbit. The foramen for n. trigeminus (c.n. V) is surrounded by a fossa positioned topographically at the junction of the laterosphenoid, prootic, and basisphenoid. More caudally, the prootic laterally overlaps the opisthotic to enclose the otic capsule. The crista prootica crosses the external surface of these two elements and merges with the base of the paroccipital process. Running vertically through the prootic-opisthotic complex from the parietal incisure is a foramen for the vena cerebralis media. Immediately caudal to the prominent trigeminal fossa on the side of the braincase is a longitudinal row of foramina. The foramen for n. facialis (c.n. VII) is found as a discrete opening at the top of a dorsoventral groove that transmitted the palatine branch of this nerve to the opening of the internal carotid canal and then rostrally, entering the pituitary fossa within the basisphenoid. More caudally there is an auditory recess containing the fenestra ovalis for the footplate of the stapes and subsidiary foramina accommodating n. glossopharyngeus (c.n. IX) and v. jugulare; beyond this region there are foramina for nn. vagus and accessorius (c.nn. X, XI), and a foramen for n. hypoglossus (c.n. XII)—the degree to which these are preserved varies among taxa.
Projecting rostroventrally from the base of the basisphenoid, the basipterygoid process articulates with a reciprocal surface on the central plate of the pterygoid. A swollen basal tuber is located caudal to the basipterygoid process and marks the area of the basioccipital suture. The basioccipital has the approximate form of a hemicylinder and contributes to the median floor of the endocranial cavity. The lateral and ventral walls of the basioccipital are frequently marked by muscle scars associated with anchorage of the median neck musculature. The caudal end of the basioccipital is expanded to form the bulk of the occipital condyle, which ranges in form from crescentic to subspherical in caudal aspect.
The caudal region of the braincase is dominated by the opisthotic that flares obliquely laterally to form a stout, somewhat barlike paroccipital process. In occipital view the supraoccipital contributes much of the dorsal rim of the foramen magnum (fig. 18.1B). In Heterodontosaurus tucki, Zephyrosaurus schaffi, and Hypsilophodon foxii, a circular vascular foramen is found in the caudal face of each paroccipital process near its dorsal edge; this probably represents a remnant of the posttemporal fenestra and transmitted vena capitis dorsalis. Small exoccipitals form the ventrolateral margins of the foramen magnum and contribute as condylids to the cranioatlantal joint; the remaining, larger part of the occipital condyle is formed by the basioccipital.
The palate where known is moderately vaulted (viz. Hypsilophodon foxii, Galton 1974a). Rostrally, the fused vomers form a narrow median spine that deepens and subdivides caudally, forming plates of bone that curve upward and increase in depth. Each vomer has a loose ventral contact with the palatine and the palatine ramus of the pterygoid. The palatine is fan-shaped and dished dorsally where it floors the orbit and sutures firmly with the caudal process of the maxilla along a ridged surface. The palatine extends caudally and contacts the pterygoid along the rostral margin of the latter element. The triradiate pterygoid forms much of the suspensorium that links the braincase, maxilla, and rostral portion of the palate by means of an array of thin rami. The central part of the pterygoid (from which all the rami radiate) articulates with the basipterygoid processes of the basisphenoid. The quadrate ramus consists of two parts: a more ventral horizontal region that is short and stout and a more dorsal contact that is high, thin, and oblique. The rostral edge of the ventral part of the ramus contacts the ectopterygoid laterally, and then projects ventrally nearly to the level of the mandibular condyle. In all cases, the sharply curved ectopterygoid forms a stout, straplike element that helps to lock the maxilla and pterygoid together.
The predentary is scoop-shaped and has paired lateral processes that cap the rostral end of each dentary and a short median ventral process that underlies the dentary symphysis. The oral margin is sharp and unserrated. The body of the predentary of Abrictosaurus has a large foramen on its lateral aspect (Thulborn 1974). In both Abrictosaurus and Heterodontosaurus the predentary fits against the slightly inflated blunt rostral surface of each dentary, creating a spheroid joint between them (Weishampel 1984c); in basal euornithopods this articulation is more constrained.
Caudally, the dentary becomes thicker, deeper, and divergent, especially where it forms an elevated coronoid process. In Abrictosaurus consors (Thulborn 1974) three foramina are found at the rostral end of the dentary near its contact with the predentary; such foramina may be lacking in Heterodontosaurus tucki. Otherwise, a number of prominent neurovascular foramina are found along the body of the dentary along the approximate margin of the floor of the buccal recess. A discrete coronoid process (rather than a rounded eminence) is developed and rises above the ventral edge of the jugal. In all taxa the jaw articulation is set below the occlusal plane of the dentary dentition and the latter is inset from the side of the dentary, producing a pronounced buccal recess (figs. 18.1A, 18.3A). Each mandibular ramus has a sinuous outline in occlusal view, and the tooth row extends along the medial edge of the dentary, terminating at the base of the coronoid process (fig. 18.3C).
The number of tooth positions in the dentary dentition is variable. Thirteen tooth positions are preserved in Heterodontosaurus tucki (the first is occupied by a large, strongly caniniform tooth and accounts for the rostral expansion of the dentary), 13or 14are recorded in Abrictosaurus consors, Hypsilophodon foxii, Othnielia rex, Orodromeus makelai, and possibly “Gongbusaurus” wucaiwanensis, and 18 or more in Thescelosaurus neglectus, Agilisaurus louderbacki, “A.” multidens, and Parksosaurus warreni.
The surangular forms a caudal buttress for the coronoid process, the lateral wall of the mandibular fossa, the outer edge of the glenoid (including a lip or buttress somewhat rostral to the articulation), and the lateral wall of the retroarticular process. Contact with the dentary (sometimes at the junction with the angular) is often marked by a small foramen that probably represents the remnant of the external mandibular fenestra (seen in Heterodontosaurus tucki, Hypsilophodon foxii, Thescelosaurus neglectus, Bugenasaura infernalis, and Gasparinisaura cincosaltensis).
The coronoid, known in Hypsilophodon foxii, Orodromeus makelai, and Thescelosaurus neglectus, fits against the medial surface of the coronoid process of the dentary and projects slightly above it; its rostral extent is unclear. The prearticular forms the medioventral wall of the adductor fossa and medially contacts the dentary, surangular, and articular. The articular forms the medial portion of the jaw joint as well as the medial surface of the retroarticular process. In lateral view, the angular overlaps the external surface of the surangular, but has little medial exposure, such that it contributes only a wedge to the caudoventral border of the mandible. The large splenial makes up most of the medial aspect of the mandible at its midsection, where it covers the mandibular canal.
In Agilisaurus louderbacki, Hypsilophodon foxii, Zephyrosaurus schaffi, Orodromeus makelai, Thescelosaurus neglectus, and Bugenasaura infernalis, the premaxillary teeth are small, peglike, and slightly recurved. The crowns of all premaxillary teeth are completely covered by enamel. In Heterodontosaurus tucki, the first two premaxillary teeth are roughly the same, while the third (and last) is caniniform with fine serrations on both mesial and distal margins. The absence of a caniniform tooth led Thulborn (1974) to suggest that the individual attributed to Abrictosaurus (= Lycorhinus) consors was female; however, this absence may equally well represent a juvenile condition or a stage in the tooth replacement cycle. Vertical tooth wear on the inner premaxillary crown surfaces no doubt results from occlusion with the predentary rhamphotheca.
Among heterodontosaurids, the maxillary dentition is well known in Abrictosaurus consors, Lycorhinus angustidens, and Heterodontosaurus tucki (figs. 18.1, 18.2). The crowns are chisel-shaped, with denticles restricted to the apical third. The mesial and distal margins of successive crowns are well separated in A. consors, moderately overlapping in L. angustidens (based only on the lower dentition), and in close contact in H. tucki. Tooth replacement was continuous but slowed down in old age (Hopson 1980b). Spacing between Zahnreihen varies around 3.0. In L. angustidens, the teeth within each tooth row are arranged in offset triplets that reflect emplacement of teeth of roughly the same age. Enamel is continuous on all surfaces of the tooth crowns in L. angustidens and A. consors, but the buccal surface of maxillary tooth crowns and the lingual surface of dentary tooth crowns are more heavily enameled. In contrast, in H. tucki, enamel is restricted largely to the buccal and lingual surfaces of maxillary and dentary crowns, respectively. The maxillary teeth of L. angustidens (Gow 1990b) have three to seven marginal denticles on each mesial or distal edge. The most distal denticle is the largest, separated from the remainder of the crown on both the buccal and the lingual surfaces by a deep groove, and merges with the basal cingulum. There is the slight suggestion of a broad central eminence on the buccal surface of the crown, but there are no secondary ridges. The crowns of the maxillary dentition in A. consors vary from slightly higher than wide to twice as high as wide. The apex of the crown is either symmetric or slightly displaced distally. Four to seven denticles are situated toward the apex of the crown and are supported by weak, more or less vertical ridges on the buccal surface. The most distal ridge merges with the basal cingulum. Ornamentation is not differentiated into primary and secondary ridges. In H. tucki, the maxillary teeth have high crowns and are more robust than in A. consors. A large primary ridge is symmetrically placed on the buccal surface of the crown. The most mesial and distal ridges are slightly less robust than the primary ridge; these outer ridges converge at the base of the crown. A single secondary ridge is developed mesially and distally midway between the primary ridge and the outer ridges. There is no cingulum, so that the crown merges gradually with the columnar root.
Extensive tooth wear is found in all heterodontosaurids. In Lycorhinus angustidens, the lingual surface is planed off nearly to the roots, thereby truncating virtually all marginal features with the exception of the most distal denticle (fig. 18.2C). The resulting mesial and distal facets are of slightly different orientation but create an overall occlusal surface that is highly inclined (75°–80° below the horizontal). In Abrictosaurus consors, there is a large facet; in Heterodontosaurus tucki, there are two facets set off at a slight angle and each is continuous on adjacent teeth. The wear facets are much less steeply inclined than in Lycorhinus ranging from 30° to 40° below the horizontal.
Among basal euornithopods (fig. 18.6), the maxillary dentition generally consists of mesiodistally broad shield-shaped crowns with denticulate margins and a swollen base that forms an incipient cingulum. In Thescelosaurus neglectus, Bugenasaura infernalis, Agilisaurus louderbacki, “A.” multidens, Drinker nisti, Othnielia rex, and Orodromeus makelai, the crowns are triangular, with a centrally placed apex. In contrast, in Zephyrosaurus schaffi, Yandusaurus hongheensis, Gasparinisaura cincosaltensis, Atlascopcosaurus loadsi, and Hypsilophodon foxii, the crown is distinctly asymmetric in buccolingual view, with the apex offset distally. Enamel is deposited on both sides of the crown in Leaellynasaura amicagraphica, Yandusaurus hongheensis, Parksosaurus warreni, Drinker nisti, and “Agilisaurus” multidens, whereas in Thescelosaurus neglectus, Othnielia rex, Hypsilophodon foxii, Gasparinisaura cincosaltensis, and Zephyrosaurus schaffi, it is thicker on the buccal surface. The buccal surface often bears a fanlike array of ribs or subvertical ridges in the enamel that terminate in a marginal denticle; however, not all marginal denticles are necessarily supported by ridges (viz. Hypsilophodon foxii). The total number of marginal denticles varies: Agilisaurus louderbacki, Atlascopcosaurus loadsi, Orodromeus makelai (fig. 18.6A–C), and Zephyrosaurus schaffi show a total of 10 or 11 denticles along the mesial and distal margins; there are 8 or fewer in “Agilisaurus” multidens, Gasparinisaura cincosaltensis, Bugenasaura infernalis, Thescelosaurus neglectus, Yandusaurus hongheensis, Atlascopcosaurus loadsi, and Parksosaurus warreni, and more than 15 in Drinker nisti (which also has subsidiary denticles on each denticle as well as a sharp, denticulate, partial cingulum; fig. 18.6E, F), Othnielia rex (fig. 18.6D), and Hypsilophodon foxii (fig. 18.3D, E). There is no differentiation of the vertical ridges, except in Gasparinisaura cincosaltensis and Atlascopcosaurus loadsi, in which the primary ridge reaching the apical denticle is more prominent than the others. No supporting ridges are found in Orodromeus makelai, Agilisaurus louderbacki, and “A.” multidens. In Othnielia rex, Zephyrosaurus schaffi, Leaellynasaura amicagraphica, and Thescelosaurus neglectus, ridges extend on to the lingual side of the crown.
Tooth replacement among basal euornithopods is polyphyodont, with two generations of teeth per tooth position. The spacing between Zahnreihen ranges from 2.2 to2.3 in Parksosaurus warreni and Hypsilophodon foxii, to 3.0 in Zephyrosaurus schaffi.
The occlusal surface of functional teeth typically shows substantial wear in the form of oblique single or double facets depending on the state of wear. In either case, wear facets are inclined lingually, shallowly concave(50°–65° from horizontal). Together with their partially interlocking arrangement, tooth wear across the entire maxillary row produces a continuous if uneven cutting edge.
The dentary dentition of basal ornithopods is similar to that of the maxilla, but with some significant differences. In heterodontosaurids, the first dentary tooth is enlarged to become caniniform, is thinly enameled, and fits into the maxillary diastema. In material referred to Abrictosaurus consors (Charig and Crompton 1974; Hopson 1975a), only the sharp mesial edge of the caniniform tooth crown bears fine serrations, whereas in Lycorhinus angustidens and Heterodontosaurus tucki, both the mesial and distal margins are serrated. A small diastema separates the caniniform tooth from the remaining lower cheek teeth. The second tooth is small, conical, and recurved, at least in L. angustidens and referred material of Abrictosaurus consors. The only known material without a caniniform dentary tooth is the type specimen of A. consors (Thulborn 1974)—in this instance the first dentary tooth is small, broad, and recurved and the mesial and distal margins lack denticles. The second and third crowns are similar in structure; the third, however, is denticulate along its distal edge and transitional in form to those in the remainder of the series.
The more distal tooth crowns in heterodontosaurids are spatulate and approximately twice as high as wide. In Abrictosaurus consors, the middle of both lingual and buccal aspects of the crown is enlarged to form a broad central eminence. In Heterodontosaurus tucki and possibly Lycorhinus angustidens, there is clear differentiation of a strong central primary ridge. In all heterodontosaurids, the apex of the crown tends to be symmetrical, supported by a strong primary ridge. Where they are asymmetrical with respect to the lingual surface of the tooth, the apex and primary ridge are offset somewhat mesially. The apex is surrounded by three to five denticles on each mesial and distal margin in A. consors, and these denticles are supported by short secondary ridges. In H. tucki and L. angustidens, the number and configuration of secondary ridges are often difficult to determine owing to the extreme tooth wear, but those that support the most mesial and distal denticles merge with the base of the crown. A weakly developed cingulum is found in both A. consors and L. angustidens but is absent in H. tucki. Dentary tooth wear in heterodontosaurids produces either a single facet confluent mesially and distally with adjacent teeth (A. consors) or a bifaceted surface, each facet of which is continuous with a facet on neighboring crowns (L. angustidens and H. tucki). The angle of these wear facets increases with tooth age, ranging from 45° to 65° below the horizontal.
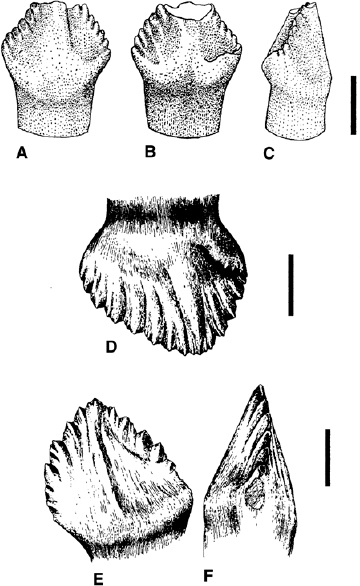
FIGURE 18.6. Basal euornithopod teeth. A–C, Orodromeus makelai dentary tooth in A, lingual, B, buccal, C, distal views. D, Othnielia rex maxillary tooth, lingual view. E, F, Drinker nisti dentary tooth in E, buccal, F, distal views. Scale = 1 mm. (A–C from Galton 1995; D–F from Bakker et al. 1990.)
The dentary dentition of basal euornithopods exhibits the same patterns of wear and replacement as the maxillary teeth. In large part, the morphology of the dentary crowns is also similar: buccolingually compressed with denticulate margins. In Hypsilophodon foxii, the mesialmost three or four dentary teeth are simple and more conical (fig. 18.3). Enamel is found on both lingual and buccal sides in “Gongbusaurus” wucaiwanensis, Orodromeus makelai, and Parksosaurus warreni, but it is considerably reduced or absent on the buccal surface in other taxa. In all basal euornithopods, the dentary crown is either triangular (Agilisaurus louderbacki, “A.” multidens, Orodromeus makelai, Othnielia rex, Thescelosaurus neglectus, Zephyrosaurus schaffi, “Gongbusaurus” wucaiwansensis) or spatulate (Hypsilophodon foxii, Atlascopcosaurus loadsi) and symmetrical, with a centrally placed apical denticle. Denticles are continuous with ridges on the thickly enameled lingual face of the dentary teeth in Atlascopcosaurus loadsi, Zephyrosaurus schaffi, Thescelosaurus neglectus, and “Agilisaurus” multidens; similar ridges are abbreviated or absent in Orodromeus makelai, Othnielia rex, and Drinker nisti. In taxa that exhibit lingual ridging, differentiation into primary and secondary ridges is known in Hypsilophodon foxii. These ridges are contiguous with the marginal denticles, 5 or 6 in Gasparinisaura cincosaltensis, 10 or 11 in Othnielia rex, “Agilisaurus” multidens, A. louderbacki, Parksosaurus warreni, Drinker nisti, Orodromeus makelai, and Thescelosaurus neglectus, and as many as 14 in Atlascopcosaurus loadsi. In Hypsilophodon foxii, there is a single primary ridge and three secondary ridges, but up to 14 marginal denticles (fig. 18.3E). As with the maxillary dentition, the marginal denticles are trifid in Drinker nisti. Numerous vertical ridges are also preserved on the buccal surface of unworn teeth in Zephyrosaurus schaffi, Thescelosaurus neglectus, Bugenasaura infernalis, Agilisaurus louderbacki, and “A.” multidens. At its base, a modest cingulum often separates the crown from the root.
The sclerotic ring (known in Abrictosaurus consors, Parksosaurus warreni, Agilisaurus louderbacki, and Hypsilophodon foxii) is composed of as many as 15 platelike ossicles that are distributed over quadrants of unequal size. The only hyoid elements known to date for these same basal ornithopods are rodlike elements presumed to be the first ceratobranchials.
Postcranial Skeleton
The following discussion of heterodontosaurid postcrania is based almost entirely upon the one well-preserved skeleton of Heterodontosaurus tucki (fig. 18.7; Santa Luca et al. 1976; Santa Luca 1980; see also Bakker and Galton 1974; Norman 1985; Bakker 1986). That of basal euornithopods (fig. 18.8) comes principally from Hypsilophodon foxii (Galton 1974a), Othnielia rex (Galton and Jensen 1973a), Orodromeus makelai (Scheetz 1999), Thescelosaurus neglectus (C. M. Sternberg 1940a; Galton 1974a), Parksosaurus warreni (Parks 1926b), Yandusaurus hongheensis (He and Cai 1984), “Agilisaurus” multidens (He and Cai 1983, 1984), A. louderbacki (Peng 1992, 1997), and Gasparinisaura cincosaltensis (Coria and Salgado 1996b; Salgado et al. 1997c).
AXIAL SKELETON
Nine vertebrae primitively make up the cervical series in Ornithopoda. Parts of the proatlas are known in Hypsilophodon foxii, Agilisaurus louderbacki, “A.” multidens, and Orodromeus makelai. As described by Galton (1974a) the proatlas is a curved element that links the atlas neural arch to the occiput. The atlas is unfused, comprising an intercentrum, a well-developed pleurocentrum (odontoid process) sutured to the axis, and separate neural arches. The prezygapophyses of the axis face laterally to receive the atlas. The axis is cylindrical, with a prominent bladelike neural spine. The naturally sigmoid curvature of the neck is demonstrated by the lateral profiles of the remaining cervical centra: parallelogram-like cranially, rectangular in the middle of the series, and trapezoidal caudally. The centra are longer in the cranial portion of the series, but the height remains constant. All centra are spool-shaped with a broad median ventral keel. The centra of the third to seventh cervical vertebrae are mildly opisthocoelous, whereas those of the eighth and ninth are amphiplatyan. There is a lateral concavity in the centra that becomes deeper caudally, especially just caudoventral to the parapophysis. The parapophyses are situated on or just below the neurocentral suture. The diapophysis is near the parapophysis cranially but migrates caudodorsally later in the series. The neural spines form a ridge at the confluence of the postzygapophyses for much of the series, but start to develop a slight elevation and hooked profile as they approach the dorsal series. The cranial cervical zygapophyses behind the axis are nearly horizontal, whereas the middle ones are inclined and the caudal ones become more horizontal again. With the exception of those of the atlas and axis, all the cervical ribs are double-headed, with the capitulum and tuberculum close together; caudally, they become more widely separated and the tubercular process becomes shorter.
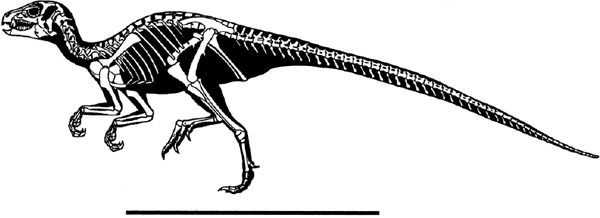
FIGURE 18.7. Skeletal reconstruction of Heterodontosaurus tucki. Scale = 50 cm. (Modified from Paul in Brett-Surman 1997.)
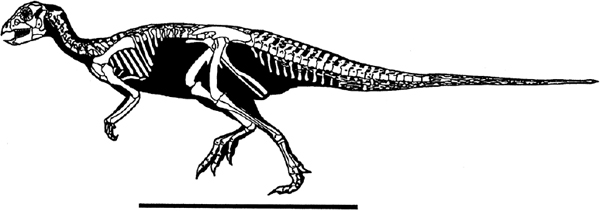
FIGURE 18.8. Skeletal reconstruction of Hypsilophodon foxii. Scale = 50 cm. (Copyright G. S. Paul.)
Dorsal vertebrae probably numbered either 15 or 16, depending on the incorporation of a dorsal into the sacrum. All dorsal vertebrae have amphiplatyan centra, the lengths of which increase slightly caudally. The centra are spool-shaped and rectangular in lateral view. The transverse processes are horizontal, directed caudolaterally, and are situated at the level of the zygapophyses. The parapophysis moves on to the transverse process near the cranial end of the series. In the middle portion of the series, the parapophysis and diapophysis are found on the cranial and caudal portions of the transverse process, respectively, but merge on the transverse process of the last dorsals. The neural spines are stout, rectangular, and nearly as long as the centra. The cranial dorsal zygapophyses are horizontal and become inclined to about 45° caudally.
The dorsal ribs, with the exception of the last one or two, are double-headed. The capitulum and tuberculum are close together and the tuberculum becomes reduced caudally. The first six or seven ribs are curved, especially near their proximal ends, and have broad and flattened distal shafts. Ribs caudal to the seventh dorsal vertebra become progressively shorter and straighter. In Hypsilophodon foxii, Parksosaurus warreni, Othnielia rex, and Thescelosaurus neglectus, the sternal segments of the first six or so ribs tend to be at least mineralized if not ossified.
There are five co-ossified sacral vertebrae in Agilisaurus louderbacki and “A.” multidens, whereas Heterodontosaurus tucki, Orodromeus makelai, Parksosaurus warreni, Thescelosaurus neglectus, Othnielia rex, and Gasparinisaura cincosaltensis have six sacrals. Galton (1974a) reported dimorphism in the structure of the sacrum in Hypsilophodon foxii, in which some specimens have five sacral vertebrae, with the centrum of the last dorsal expanded caudally and forming extensive sutural contacts with the first sacral rib. By contrast, other individuals have six sacral vertebrae; the first vertebra is a sacral because its ribs are sutured to both the centrum and the neural arch and contact the pubic peduncles of the ilia. In addition, Thulborn (1974) reported that Abrictosaurus consors has four unfused sacrals; this may be evidence of juvenility, consistent with the absence of caniniform teeth in the same individual.
A reasonably complete caudal series is available from Heterodontosaurus tucki (34–37 vertebrae), Parksosaurus warreni (42 vertebrae), Agilisaurus louderbacki (44 vertebrae), and Hypsilophodon foxii (45–50 vertebrae). All of the vertebrae are amphicoelous and become progressively lower and more slender toward the distal end of the tail. The proximal caudal centra are more concave ventrally because the facets for articulation of the chevrons are more pronounced; on the distal caudals, the articulation is reduced and the ventral border is flatter. The transverse processes are horizontal, ventral to the neurocentral suture, and directed laterodistally. The proximal transverse processes are strong, but more distal ones are weaker and the distalmost are absent. The neural spines are vertical and tall proximally but become inclined and shorter distally. Neural spines are absent distal to the tenth caudal. The zygapophyses are elevated about 45° proximally but become more nearly vertical distally. Chevrons are found throughout the caudal series. Proximally, they are robust and become progressively more slender distally; those from the middle of the series are slightly expanded distally. The chevrons of Gasparinisaura cincosaltensis differ from other ornithopods in being platelike and subtriangular, with an asymmetrical outline.
In nearly all taxa, ossified tendons are found along the epaxial region of the dorsal, sacral, and caudal portions of the vertebral column. These tendons extend in parallel longitudinal rows along the sides of the neural spines. Both epaxial and hypaxial regions of the tail are sheathed in a large number of ossified tendons in Hypsilophodon foxii, Thescelosaurus neglectus, Gasparinisaura cincosaltensis, and a variety of other basal euornithopods.
Small elements of axial dermal armor near the skull have been reported in Hypsilophodon (Hulke 1874b; Nopcsa 1905c; disputed by Galton 1974a). Dermal ossifications have been illustrated in Bugenasaura (Morris 1976). More extensive body armor associated with the ribcage has also been reported in articulated skeletal material referred to Thescelosaurus (Fisher et al. 2000).
APPENDICULAR SKELETON
In all basal ornithopods, the scapula is the same length as, or slightly longer than, the humerus. The scapular shaft is bowed along its length to fit around the rib cage; it has a rounded transverse midsection but expands into a flattened blade distally. In Heterodontosaurus tucki, the scapular shaft is narrow and rather straplike although it is expanded distally (Santa Luca 1980). In basal euornithopod taxa it is wide (especially so in Thescelosaurus neglectus). The convex distal end of the scapular blade is pitted, indicating the presence of a cartilaginous suprascapula (partially ossified in Parksosaurus warreni). The shaft is slightly concave internally to accommodate the rib cage. The ventral end is expanded, into a flangelike acromion that projects cranially (large in Hypsilophodon foxii, Thescelosaurus neglectus, Orodromeus makelai, Othnielia rex, “Agilisaurus” multidens, and A. louderbacki and less pronounced in Parksosaurus warreni, Gasparinisaura cincosaltensis, and Yandusaurus hongheensis). The glenoid fossa of the scapula faces caudolaterally. In Heterodontosaurus the triceps tubercle is separated from the dorsal lip of the glenoid.
The subrectangular coracoid has flat proximal and distal ends. Its distal portion curves strongly medially and the sternal process is especially prominent in Hypsilophodon foxii, Parksosaurus warreni, Orodromeus makelai, Othnielia rex, and Gasparinisaura cincosaltensis. Together with that contributed by the scapula, the glenoid fossa of the coracoid faces caudolaterally. In external view, the coracoid foramen is not contiguous with or close to the scapulocoracoid suture. There is a tubercle for muscle attachment at the ventral corner of the coracoid.
Santa Luca (1980) reported sternal plates in Heterodontosaurus tucki; however, details of both structure and attachments cannot be determined. In Hypsilophodon foxii, Parksosaurus warreni, and probably in Orodromeus makelai and Othnielia rex, the flat sternals are reniform in ventral view. The sagittal rim is thickened where each element meets its partner. The caudolateral margin is also thickened where it articulates with the sternal segments of the ribs.
In Heterodontosaurus tucki, Orodromeus makelai, Yandusaurus hongheensis, Thescelosaurus neglectus, and Hypsilophodon foxii, the humerus is robust, whereas in Notohypsilophodon comodorensis, Gasparinisaura cincosaltensis, Othnielia rex, “Agilisaurus” multidens, and A. louderbacki it is considerably more gracile. In all taxa, the shaft is slightly twisted and the proximal end of the humerus is broader than the distal end. Adjacent to the humeral head is a large medial tuberosity. In Heterodontosaurus tucki, Hypsilophodon foxii, Yandusaurus hongheensis, and Orodromeus makelai, the deltopectoral crest is limited to the proximal half of the humerus. In all these species except “Agilisaurus” multidens, Othnielia rex, and Thescelosaurus neglectus (where it is low), the deltopectoral crest is angular and directed craniolaterally, extending for about 40% of the length of the humerus. In contrast, the deltopectoral crest of Notohypsilophodon comodorensis, Agilisaurus louderbacki, Gasparinisaura cincosaltensis, Bugenasaura sp., and perhaps Parksosaurus warreni is restricted to the proximal half of the humerus, is low, and bears a less distinct angular margin or none at all. In all of these species, the humerus bears well-developed distal condyles; the articular surfaces generally do not extend caudally, and there is no more than a modest olecranon fossa in some species. The ulnar condyle is rounded and projects distally beyond the radial condyle. In Heterodontosaurus tucki, a prominent medial epicondyle projects medially from the shaft above the ulnar condyle. The radial condyle is less rounded and is inclined somewhat laterally. A lateral supracondylar ridge extends proximally from the radial condyle in H. tucki.
In all basal ornithopods, the length of the radius is about 70% that of the humerus. The radial shaft is somewhat twisted. Proximally, the shaft is circular in cross section and becomes more triangular or rectangular distally. The humeral articulation of the radius forms a semicircular sulcus; a process extends proximomedially from the caudal surface and contacts the ulna. The distal articulation has planar contacts for the ulnare as well as the radiale.
The length of the ulna is about 80% that of the humerus. The most striking feature of the ulna in Heterodontosaurus tucki is the prominent olecranon that bears rugosities for the insertion of M. triceps. By contrast, in Hypsilophodon foxii and Parksosaurus warreni, the ulna bears only a moderately developed olecranon process, whereas in Notohypsilophodon comodorensis this process is small. The olecranon process is notably well developed in Heterodontosaurus tucki compared to all other basal ornithopods. The humeral articulation is somewhat trochlear. The shaft narrows in the middle and expands distally.
The carpus is poorly known among basal ornithopods. That of Heterodontosaurus tucki (fig. 18.9A; Santa Luca 1980) comprises nine bones, a proximal row of the radiale, ulnare, and pisiform, a middle row (intermedium, contra Santa Luca 1980), and a distal row (distal carpals 1–5). In contrast, the carpus of Hypsilophodon foxii (fig. 18.9A; Galton 1974a) is formed only of the ulnare, intermedium, and a single distal carpal, whereas in Orodromeus makelai, there is an ulnare, intermedium, and one distal carpal (Scheetz 1999). Finally, “Agilisaurus” multidens bears an ulnare, intermedium, and radiale. The wrist is best documented in Heterodontosaurus tucki, in which the radiale forms broad contacts with the radius and distal carpal 1 and more restricted contacts with adjacent carpals. The ulnare bears a concave articulation for the ulna and a planar one for the radius; it articulates broadly with the centrale and distal carpal 4 and narrowly with distal carpal 5 and the pisiform. The intermedium intervenes between proximal and distal carpals. Distal carpals 1–3 are large; distal carpals 4 and 5 are abducted.
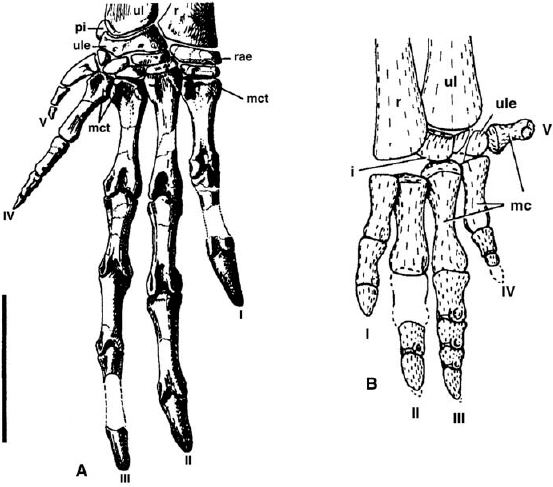
FIGURE 18.9. A, Right manus of Heterodontosaurus tucki in dorsal view; B, left manus of Hypsilophodon foxii in dorsal view. Scale = 2.5 cm. (A after Santa Luca 1980; B after Galton 1974a.)
In Hypsilophodon foxii, the ulnare articulates proximally with the medial margin of the ulna, medially with metacarpal V, and distally with metacarpal IV; otherwise its morphology is not determinable. The intermedium, a convexoconcave nubbin of bone, articulates proximally with the greater part of the distal ulna and the medial margin of the distal radius. Smaller cuboid and lenticular elements correspond to the distal carpals. The condition in Orodromeus makelai and “Agilisaurus” multidens is thought to be most similar to that of H. foxii.
In all basal ornithopods, the manus is small relative to the pes. Although Thulborn (1974) described the manus of A. consors as diminutive, the basis for this is unclear; it is large in the related form Heterodontosaurus, longer than the radius or ulna and almost as long as the humerus. Metacarpal II is the longest, followed by metacarpals III, IV, and V. The carpal articulation of metacarpals I and II is carried onto the dorsal surface, indicating that these carpometacarpal joints could have been extended farther than the others. The dorsal surfaces of the bases of metacarpals I–IV bear strong lateral and medial tubercles for extensor tendons. As a result of the tight planar articulations between adjacent metacarpals and the carpals, the bases of metacarpals II–V are almost cuboid. Metacarpal I is unusual in that it is asymmetric, with a longer lateral border. Its metacarpophalangeal joint is also asymmetric: the medial condyle is aligned with the long axis of the shaft, whereas the lateral condyle is perpendicular to it. This asymmetry would cause the digit to deviate medially on extension and laterally on flexion. Metacarpophalangeal joints II and III are symmetrical and restrict movement to simple flexion and extension. The cranial surfaces of the metacarpal heads of digits I–III bear deep pits that received processes from the proximal phalanges; this suggests that digits I–III could be hyperextended at the metacarpophalangeal joints. Metacarpal IV is reduced distal to its proximal cuboid portion, and metacarpal V is even more reduced.
Among basal euornithopods, in Hypsilophodon foxii, metacarpal V, the smallest, is distinctly set off from the other metacarpals and metacarpals I–IV are closely packed, running parallel with each other. In contrast, in Orodromeus makelai and “Agilisaurus” multidens, metacarpal V is much less medially angled and the remaining metacarpus is less closely packed. In all taxa, metacarpal III is the longest, followed by metacarpals II, IV, and I. The distal condyles exhibit a modest extensor groove and there is no dorsal intercondylar groove.
The phalangeal formula of the manus of Heterodontosaurus tucki is 2-3-4-3-2. Digits IV and V are reduced, whereas digits I–III are large (fig. 18.9A) and complex and exhibit elements of asymmetry. The first phalanx of digit II and the first and second of digit III are characterized by asymmetric distal articulations. The shafts are twisted such that the heads are medially rotated. Likewise, the lateral margins are longer, imparting a medial curve to the shafts. Finally, the medial condyle is larger and oriented differently than the lateral condyle. These articulations would cause lateral deviation during flexion and medial deviation during extension. The penultimate phalanges of digits I–III are symmetrical and permitted simple extension and flexion. The unguals of digits I–III formed large claws; the flexor tubercles are unusually large for an ornithischian. The phalangeal formula of Abrictosaurus consors is reported to be 2-3-4-2-1. Among basal euornithopods, the phalangeal formula in Hypsilophodon foxii is 2-3-4-3?-? (fig. 18.9B), in “Agilisaurus” multidens it is 2-3-4-2?-2, and in Orodromeus makelai it is ?-3-4-3-?. Unlike the highly modified manus of Heterodontosaurus tucki, the metacarpophalangeal and interphalangeal articulations are more symmetrical, but less well developed. However, the unguals are similarly clawlike. In other taxa—Thescelosaurus neglectus and Yandusaurus hongheensis, among them—the manus is poorly known, while in the remainder it has yet to be discovered.
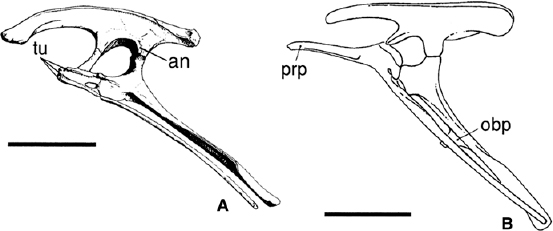
FIGURE 18.10. A, Pelvis of Heterodontosaurus tucki in left lateral view; B, pelvis of Hypsilophodon foxii in left lateral view. Scale = 5 cm (A), 10 cm (B). (A after Santa Luca 1980; B after Galton 1974a.)
The profile of the ilium is primitively low in Ornithopoda (fig. 18.10). In Heterodontosaurus tucki (fig. 18.10A), Othnielia rex, Orodromeus makelai, Agilisaurus louderbacki, “A.” multidens, Parksosaurus warreni, and Hypsilophodon foxii (fig. 18.10B), the dorsal margin is broadly convex, while in Gasparinisaura cincosaltensis and Thescelosaurus neglectus, it is sigmoidal. A supracetabular process (“antitrochanter”) is developed as a narrow, but prominent, vertical flange on the caudal margin of the acetabulum above the ischial peduncle in Heterodontosaurus tucki (fig. 18.10A) and this feature is variably developed in other species (e.g., Hypsilophodon has a bulbous lateral expansion above the ischial peduncle). The slightly twisted preacetabular process curves outward (bearing a longitudinal ridge along its medial surface) and is about as long as the postacetabular process. The postacetabular is thick and shallow in Heterodontosaurus, but in euornithopods the process is laterally compressed and the brevis shelf that underlies it is horizontal. This latter shelf is shallow in Heterodontosaurus tucki and possibly Parksosaurus warreni, but deep in Agilisaurus louderbacki, “A.” multidens, Orodromeus makelai, Gasparinisaura cincosaltensis, Hypsilophodon foxii, Parksosaurus warreni, and Othnielia rex. The pubic peduncle is slender and projects cranioventrally at an angle of about 20° to the horizontal; it is especially long in Heterodontosaurus tucki and more perpendicular to the long axis of the ilium. The ischial peduncle is shorter and more robust than the pubic peduncle. The supraacetabular rim is modestly well developed.
In Heterodontosaurus tucki, the pubis has a short prepubic process that is stout, is squared-off cranially, and bears three small tubercles (fig. 18.10A). In contrast, that of Hypsilophodon foxii (fig. 18.10B) Thescelosaurus neglectus, Othnielia rex, Gasparinisaura cincosaltensis, Orodromeus makelai, “Agilisaurus” multidens, and A. louderbacki is long, has a flat striated external surface, and its ventral edge is grooved. The pubis itself is a simple long rod that is usually straight but sometimes slightly dorsally concave in all basal ornithopods, except for Parksosaurus warreni in which it is short. The obturator foramen is fully enclosed by the pubis in H. tucki, G. cincosaltensis, O. makelai, “Agilisaurus” multidens, A. louderbacki, and H. foxii, but in O. rex it is open, forming a notch. Both conditions are known in T. neglectus.
The iliac peduncle of the ischium is rather long and rectangular, whereas the pubic peduncle is broad and spatulate (fig. 18.10A). The expanded proximal end of the ischium is separated from the large, bladelike distal portion by a constricted shaft. The obturator process is either absent or represented by a flangelike ledge (fig. 18.10A) in Heterodontosaurus tucki. In contrast, in Parksosaurus warreni (where the ischium is unusually short) and Hypsilophodon foxii (fig. 18.10B) it is ventrally situated at about midlength on the shaft of the ischium, while in Thescelosaurus neglectus, Othnielia rex, “Agilisaurus” multidens, A. louderbacki, Gasparinisaura cincosaltensis, and Orodromeus makelai, it is positioned on the proximal third of the bone.
The long bones have gracile proportions, although they are much more robust than the forelimb in nearly all basal ornithopods. The femur of all basal ornithopods is straight in cranial view and only slightly bowed in lateral view. The femoral head is oriented at about 90° to the shaft, except in Orodromeus makelai and Notohypsilophodon comodorensis, where it makes a 110° angle. The greater trochanter is not well separated from the head and is on about the same level in Heterodontosaurus tucki, Leaellynasaura amicagraphica, Agilisaurus louderbacki, “A.” multidens, Hypsilophodon foxii, and Yandusaurus hongheensis. The cranial trochanter in H. tucki, Gasparinisaura cincosaltensis, O. makelai, L. amicagraphica, and Parksosaurus warreni is not separated from the greater trochanter by a cleft. In contrast, the cranial trochanter is separated from the greater trochanter by a shallow cleft in Othnielia rex, Y. hongheensis, N. comodorensis, A. louderbacki, “A.” multidens, Fulgurotherium australe, and H. foxii. The fourth trochanter is pendent and located on the proximal half of the femur. In H. tucki there are neither extensor nor flexor intercondylar grooves (Santa Luca 1980), but in all basal euornithopods, the extensor groove is weakly developed and on the flexor aspect, the subequal lateral and medial condyles form a deep caudal intercondylar groove. The lateral condyle extends more distally than the medial condyle, suggesting that the femur angled slightly away from the sagittal plane.
The tibia is longer than the femur by as much as 30%. Proximally, it is only moderately expanded. The medial tibial condyle is larger than the lateral and is mediolaterally compressed. The cnemial crest is well developed in Heterodontosaurus tucki, Parksosaurus warreni, and possibly “Gongbusaurus” wucaiwanensis, and bears an elongate fossa on its lateral surface, while the cnemial crest is somewhat smaller in Notohypsilophodon comodorensis, Thescelosaurus neglectus, Orodromeus makelai, “Agilisaurus” multidens, and A. louderbacki. In all species, the tibial shaft is straight and transversely expanded distally. H. tucki is unusual in having the distal tibia fused to both the adjacent fibula and the proximal tarsals, producing a rather birdlike tibiotarsus. In all basal ornithopods, the fibular head is laterally compressed and the shaft attenuates distally.
In Heterodontosaurus tucki, the astragalus and calcaneum are fused to each other and the fused distal tibia and fibula (Santa Luca 1980), while in all other forms they remain distinct. The astragalus caps the distal articular end of the tibia and wraps around onto the extensor face of that bone to form a modest ascending process. The blocklike calcaneum caps the distal end of the fibula and abuts the lateral aspect of the astragalus. The number of distal tarsals is variable among basal ornithopods. Four are known in “Gongbusaurus” wucaiwanensis (Dong, 1989). The first is ovoid and articulates with metatarsal II; distal tarsal 2 is intermediate between the proximal surfaces of metatarsals II and III; distal tarsal 3 (the largest) lies on metatarsal III; and the fourth element articulates with metatarsals IV and V. Fewer distal tarsals are found in other basal ornithopods. In Heterodontosaurus tucki, three elements are fused with each other and with the metatarsals, producing a tarsometatarsus. Distal tarsal 1 is applied to metatarsals I and II, and distal tarsals 2 and 3 contact metatarsals III and IV, respectively. The caudal surface of distal tarsal 1 is expanded into a vertical flange functionally similar to the hypotarsal ridges of birds. The cranial surface of distal tarsal 3 is pierced by a foramen (Santa Luca 1980). In Hypsilophodon foxii, Othnielia rex, Agilisaurus louderbacki, “A.” multidens, Parksosaurus warreni, and Orodromeus makelai, there are two ovoid distal tarsals, while in Gasparinisaura cincosaltensis (Coria and Salgado 1996b; Salgado et al. 1997c) this number is further reduced to one, although the single element may be formed by the fusion of two original elements.
Primitively within Ornithopoda, metatarsal III is the longest (more than half the length of the femur) and most robust; metatarsal V is a short splint and metatarsal I is about half as long as metatarsal III. The phalangeal formula of the pes in all species except Gasparinisaura cincosaltensis and Orodromeus makelai is 2-3-4-5-0; in the former, all of the phalanges are eliminated from digit I (Coria and Salgado 1996b). The foot was digitigrade (although digit I was too short to reach the ground during normal locomotion) and the unguals are narrow and clawlike, yet flat ventrally. The hallux in Hypsilophodon foxii was not opposable, contrary to earlier assertions (Galton 1971d, 1971e).
Systematics and Evolution
Formerly these animals, particularly those referred to as hypsilophodontids (viz. Sues and Norman 1990), were depicted as an implicitly paraphyletic assemblage from which all other groups of ornithischians evolved (Romer 1966; Galton 1972, 1974a, 1974b, 1980d). Heterodontosauridae (Kuhn 1966) was put forward as a monophyletic group (Galton 1972; Thulborn 1974). This latter hypothesis together with a basal position among ornithopods was supported by later research (Sereno 1986, 1991b, 1997). In addition, Milner and Norman (1984), Norman (1984a, 1984b, 1998), Sereno (1984, 1986, 1997, 1999a), Cooper (1985), and Weishampel and Heinrich (1992) advocated a more restricted, monophyletic Hypsilophodontidae. Hypsilophodontid monophyly has been challenged recently following brief reports of more comprehensive cladistic analyses (Scheetz 1998, 1999; Winkler et al. 1998).
In order to assess the topology of the evolutionary tree of basal ornithopods, a character-taxon matrix was constructed and a cladistic analysis was carried out using ten of the better-represented species in the group (PAUP*4.0b10, Swofford 1999). Based on 54 characters, outgroup comparisons were made using Marginocephalia and the theoretical ancestor outgroup. The branch-and-bound analysis yielded 28 equally most parsimonious trees. This analysis was repeated after character reweighting (using maximum values of the rescaled consistency indexes). The second run of the data generated three equally most parsimonious trees, whose topology is represented in fig. 18.11A–C. For this tree L = 44.6 steps, CI = 0.82, RI = 0.83, and RC = 0.67. The data were further analyzed by restoring the original weightings and then removing three of the least stable taxa from the matrix and rerunning the analysis using the exhaustive analysis under both the ACCTRAN and DELTRAN options. Removal of Othnielia, Zephyrosaurus and “A.” multidens and running under either option resulted in two equally most parsimonious trees represented in fig. 18.12A, B. For this tree L = 77 steps, CI = 0.68, RI = 0.60, and RC = 0.41. There is a reasonably robust core topology to these cladograms, and one that confirms the challenge to the monophyly of Hypsilophodontidae proposed by Scheetz (1998) and Winkler et al. (1998).
The analysis suggests that Ornithopoda and Euornithopoda are robust clades (figs. 18.11, 18.12A), although the reduced-taxon analysis (fig. 18.12B) implies ambiguity in placing heterodontosaurids as a sister taxon to marginocephalians. A poorly resolved Ornithopoda (sensu Weishampel 1990a) is, at least historically, unexpected given its constancy as a hypothesis of relationships since at least 1984. However, it also highlights uncertainty concerning the affinities of Heterodontosauridae, which has led a number of workers to suggest that this clade may have other affinities. For example, the differing views of Santa Luca (1980), Cooper (1985), and Marya ska and Osmólska (1985), and the labile position adopted by Norman (1984a) are reflected in the patterns seen in some of the trees obtained from our analysis. The phylogenetic nexus of basal ornithopods (especially with respect to Heterodontosauridae and Marginocephalia) may have to be revisited in the near future.
ska and Osmólska (1985), and the labile position adopted by Norman (1984a) are reflected in the patterns seen in some of the trees obtained from our analysis. The phylogenetic nexus of basal ornithopods (especially with respect to Heterodontosauridae and Marginocephalia) may have to be revisited in the near future.
Heterodontosauridae share several unique features relative to euornithopods. The paroccipital process is unusually deep, the quadrate is tall (making the occipital profile taller than wide and depressing the jaw joint far below the level of the tooth row); the caudolateral process of the premaxilla is elongate (contacting the lacrimal and covering the contact of the nasal and maxilla); the cheek teeth are high crowned and, when abraded, are chisel-shaped with the denticles on the mesial and distal margins being restricted to the apical third of the crowns. The monophyly of this taxon is further supported by the caniniform tooth in both the premaxilla and dentary and perhaps by the low-angle tooth wear on the dentary teeth. Reduction or absence of a cingulum between the root and the crown may also characterize this node. Postcranially, the humerus is shorter than the scapula (a feature shared with Iguanodontia), the olecranon is prominent (also found in Rhabdodon priscus and Zalmoxes robustus), manus digit V is offset (found in Hypsilophodon foxii), the ilium has a long preacetabular process, and the distal tibia and fibula fuse not only with each other, but with the proximal tarsals as well (Santa Luca 1980).
Within-clade relationships of Heterodontosauridae are unclear. Of the three apparently valid heterodontosaurid taxa (Abrictosaurus consors, Heterodontosaurus tucki, Lycorhinus angustidens, table 18.1), only H. tucki is at all well known. We have assessed the position of the remaining heterodontosaurids in post hoc fashion. Within the clade, Lycorhinus and Heterodontosaurus may be sister taxa based on the shared presence of both mesial and distal serrations on the dentary caniniform tooth (Charig and Crompton 1974) and the broad contact between mesial and distal surfaces of the tooth crowns (may also be seen in Pisanosaurus, see chapter 14). Owing to the absence of these features, Abrictosaurus consors is excluded from this higher-level clade.
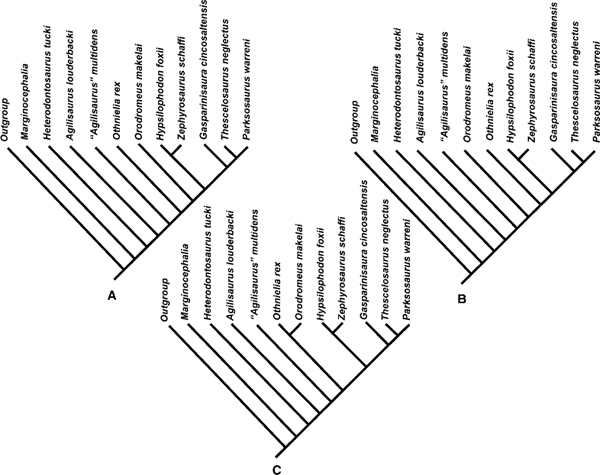
FIGURE 18.11. Cladogram of basal Ornithopoda. First run as a branch-and-bound analysis, then using the reweighting to generate trees A, B, and C.
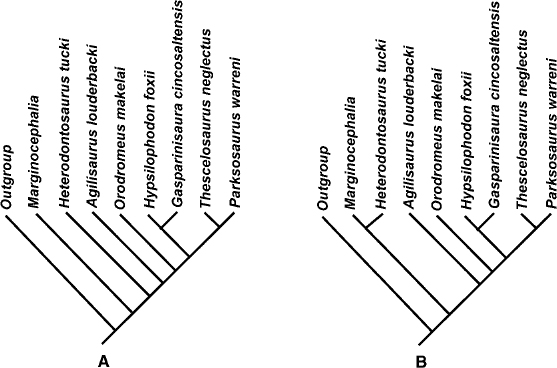
FIGURE 18.12. Cladogram of basal Ornithopoda, with the removal of Othnielia, “Agilisaurus” multidens, and Zephyrosaurus. Analysis run using Exhaustive search option, which produced two equally most parsimonious trees (A, B). L = 77 steps, CI = 0.68, RI = 0.60, RC = 0.41, Min = 52, Max = 115.
Euornithopoda can be diagnosed on the basis of the following: a scarflike suture between the postorbital and jugal, an inflated edge on the orbital margin of the postorbital, a deep postacetabular blade to the ilium, a well-developed brevis shelf, a laterally swollen ischial peduncle, an elongate and narrow prepubic process, a tablike obturator process on the ischial shaft, and a deep pit on the femoral shaft adjacent to the fourth trochanter. Other, less consistent, features include the angle subtended by the floor of the braincase and the ventral edges of the occipital condyle and basipterygoid processes, the jugal participating in the antorbital fenestra, and the position of the paraquadrate foramen.
Within Euornithopoda, this analysis does not support a monophyletic Hypsilophodontidae, which was only ever united on a few synapomorphies (Milner and Norman 1984; Sereno 1986, 1997; Weishampel and Heinrich 1992). Instead, what had formerly been at least eight species making up a monophyletic Hypsilophodontidae now form a paraphyletic sequence of taxa ever-more closely related to Iguanodontia and thereby supporting the preliminary assertions of Scheetz (1998) and Winkler et al. (1998). The most basal euornithopod recognized in this analysis is Agilisaurus louderbacki from the Middle Jurassic Lower Shaximiao Formation of Sichuan, People's Republic of China. The next less inclusive clade includes “Agilisaurus” multidens (also from the Middle Jurassic of Sichuan Province) as its basal member. Initially called Yandusaurus multidens, Peng (1992) referred this species to Agilisaurus (as A. multidens), but Barrett (pers. comm.) indicated that there are significant differences in skull structure between A. louderbacki and A. multidens sufficient to merit separate generic status. This coincides with a number of anatomical changes, including the following: reduction in the size of the external antorbital fenestra, loss of contact between the lacrimal and premaxilla, reduction in the length of the palpebral, development of a distinct cingulum on dentary crowns, and an angular deltopectoral crest. Consequently, this taxon is henceforth referred to as “A.” multidens.
Above “A.” multidens resolution at the base of the next clade is less clear. Orodromeus makelai from the Late Cretaceous and Othnielia rex from the Late Jurassic, both of the western United States, represent taxa that are closely related and less derived than the two successive taxa, Hypsilophodon foxii from the Early Cretaceous of England and Zephyrosaurus schaffi from the Early Cretaceous of the western United States. All of these taxa share a subcircular external antorbital fenestra, a distal offset to the apex of maxillary crowns, a tendency to shift the palpebral articulation to the prefrontal bone alone, a strongly constricted neck to the scapular blade, ossification of the sternal ribs, hypaxial ossified tendons in the tail, and an obturator process that is positioned near the midshaft of the ischium. The sequence of acquisition of characters is complicated by the fact that the monographic description of Orodromeus (Scheetz 1999) is not published, the partial skeleton of Othnielia (Galton 1983c) lacks any skull bones, and Zephyrosaurus (Sues 1980b) is only known from a fragmentary skull, whereas Hypsilophodon is one of the most complete and best-described ornithopods known to date (Galton 1974a).
The next most derived clade comprises Gasparinisaura cincosaltensis from the Late Cretaceous of Río Negro Province, Argentina, the more derived Thescelosaurus neglectus from the Late Cretaceous of the western United States and western Canada, and Parksosaurus warreni from the Late Cretaceous of Alberta, Canada. These are distinguished from more basal taxa by the restriction of the quadratojugal so that it has migrated away from the lower jaw articulation and the sinuous curvature to the dorsal margin of the ilium. The positioning of G. cincosaltensis differs from that suggested by Coria and Salgado (1996b) and Coria (1999), in which it has been identified as an iguanodontian with a closer relationship to Dryomorpha than to Tenontosaurus. This shift in position is due to a combination of factors, notably the loss of a monophyletic Hypsilophodontidae as well as the selection and rescoring of characters among a paraphyletic sequence of taxa.
Parksosaurus and Thescelosaurus represent the most derived of Euornithopoda considered in this chapter. They show an elongate skull form (reflected in a flattening of the frontals to form flattened rather than arching plates—provided that the reconstruction of Parksosaurus developed by Galton (1973c) is confirmed. These forms also show an elevation in the maxillary and dentary tooth counts and a considerably more robust hindlimb, reflected in the structure of the femur.
Several poorly known, but probably legitimate, species of basal Euornithopoda were not analyzed here. These include Bugenasaura infernalis, Atlascopcosaurus loadsi, Drinker nisti, Fulgurotherium australe, Leaellynasaura amicagraphica, Qantassaurus intrepidus, and Notohypsilophodon comodorensis. To the extent that they provide phylogenetically significant information, we have tried to fit them onto the cladogram in a post hoc fashion.
Bugenasaura infernalis from the Late Cretaceous of South Dakota, United States (Galton 1995, 1997b), based on a skull originally identified as ?Thescelosaurus sp. by Morris (1976) and by material not yet described (Horner, pers. comm.). On the basis of its low, elongate skull and high dental count, B. infernalis should be placed in an unresolved relationship with Thescelosaurus and Parksosaurus.
Atlascopcosaurus loadsi, from the Early Cretaceous of Victoria, Australia (Rich and Rich 1989; Rich and Vickers-Rich 1999), is a basal euornithopod, perhaps at the level of Gasparinisaura, to judge by the presence of a prominent primary ridge on the buccal and lingual surfaces of maxillary and dentary teeth, respectively; the well-developed subsidiary ridges that also traverse the crown surfaces; the apparently low tooth count; and the gracile form of associated femora.
Little can be said about the relationships of Drinker nisti, Fulgurotherium australe, and Leaellynasaura amicagraphica with other basal euornithopods. While D. nisti, from the Late Jurassic of Wyoming, United States, can be diagnosed (Bakker et al. 1990), published information on the teeth and a few postcranial elements do not provide any character information that would give it anything other than an unresolved relationship near the base of Euornithopoda.
Fulgurotherium australe, from the Early Cretaceous of Australia (New South Wales, Victoria), similarly is difficult to place among basal euornithopods. Originally known from a femur thought to belong a small theropod dinosaur (Huene 1932) but now including additional femora (Molnar and Galton 1986; Rich and Rich 1989; Rich and Vickers-Rich 1999), this species at best has an unresolved position at the base of Euornithopoda, although no synapomorphies at this level are recognized in existing material. It can also be excluded from Iguanodontia because there is no extensor groove on the femur and the medial condyle does not overlap the distal flexor fossa.
Like the other Australian taxa, Leaellynasaura amicagraphica, from the Early Cretaceous of Victoria, may be diagnostic (Rich and Vickers-Rich 1999), but lacks characters that would fit it into the cladogram. Those that have been discussed (among them, low tooth count, emarginated maxillary dentition, expansion of the greater trochanter) are plesiomorphic for Ornithopoda. Qantassaurus intrepidus (Rich and Vickers-Rich 1999) is known from short and robust dentaries with teeth of variable character (some of which bear a prominent primary ridge, as well as a fan-shaped array of subsidiary ridges). This taxon is currently too poorly known to allow specific placement in this systematic context, but bears a superficial similarity to material referred to Zalmoxes robustus (see chapter 19).
Finally, Notohypsilophodon comodorensis from the Late Cretaceous of Chubut Province, Argentina (Martinez 1998), may be a basal euornithopod. Unfortunately it consists of a fragmentary postcranial skeleton that lacks the appropriate features that would commit it to a specific level within the clade.
Biogeography
Skeletal remains referable to basal Ornithopoda are currently known from the Early Jurassic to the Late Cretaceous throughout Laurasia and Gondwana. Among them, heterodontosaurids are known best from the Early Jurassic of southern Africa, although they are also reported from the southwestern United States and possibly from the Early Cretaceous of England (Norman and Barrett 2003). Noniguanodontian euornithopods, by contrast, are known from the Middle Jurassic to the end of the Cretaceous of North America, Europe, Asia, South America, Antarctica (Hooker et al. 1991), and Australia.
Looked at in detail from a phylogenetic perspective (optimization of geographic distributional data onto the cladogram and calibrating for ghost lineages), the common ancestor of the entire ornithopod clade (including Heterodontosauridae) had an Asian distribution, particularly as Marginocephalia and Thyreophora also had an Asian origin. This common ancestor must have had an Early Jurassic or older age (the age of Heterodontosaurus). Heterodontosauridae, one of the basal lineages, thereafter dispersed to southern Africa. Euornithopoda, the immediate result of the other basal split, also has an origin in Asia prior to the Middle Jurassic (the age of material attributed to Yandusaurus hongheensis). A major dispersal to North America took place prior to the latter part of the Jurassic (the age of Othnielia rex). There were two subsequent dispersal events from North America. One was to Europe, consisting of the lineage leading to Hypsilophodon foxii, which took place sometime during or prior to the Early Cretaceous, while the other, comprising the lineage leading to Gasparinisaura, was to South America and probably took place (at the latest) during the early Late Cretaceous. Whether another dispersal event to South America is needed to account for the evolution of Notohypsilophodon, or whether it evolved in the context of the dispersal leading to Gasparinisaura, is not yet known.
Undoubtedly this biogeographic pattern, and its allied stratigraphic calibration, is heavily dependent upon extensive ghost lineage durations during critical parts of the Jurassic. This geographic interpretation may be subject to revision if the pattern of basal relationships between the Ornithopoda and Marginocephalia, as constructed, undergoes some alteration in future analyses. Discovery of new material of all of these taxa or of new ones will also produce concomitant refinements to the overall topology of current cladograms and is also likely to have a material impact on such biogeographic hypotheses.
Paleoecology and Behavior
Because the geographic and temporal distribution of the best-known heterodontosaurids is limited, it is relatively easy (perhaps deceptively so) to characterize the paleoecology of the group. The semiarid depositional environment of the Stormberg Series (Upper Elliot and Clarens formations) of southern Africa has yielded the best-known material referable to heterodontosaurids. These sedimentary rocks represent mixed flood fan and eolian dune facies (Kitching and Raath 1984; Smith 1990; Visser 1991). The siltstones and mudstones are well oxidized, so little organic matter is preserved. Flood-deposited sandstones interspersed with eolian deposits indicate that water was abundantly available only sporadically. Consequently, preservation of heterodontosaurid skeletal remains is rather poor. Isolated bones are most common, although at least one fully articulated skeleton is known. In addition, most species are not known from more than three individuals, and the majority is based on single specimens. Of these, jaw elements are the most common.
Altogether, the Upper Elliot and Clarens paleoenvironments were extremely inhospitable. It is perhaps surprising then that the associated terrestrial vertebrate assemblage is extremely diverse and often rich, including early mammals, derived nonmammalian cynodonts, and the exceptionally abundant prosauropod Massospondylus, as well as terrestrial crocodylo-morphs. No fully aquatic members of the fauna are known. It is tempting to think that at least some of these remains were transported into the catchment area by floods from more benign and luxuriant environments, but no evidence to date, either taphonomic or sedimentological, supports such a contention.
Paleoecological inferences for basal euornithopods are much more difficult to obtain, owing to their ubiquitous occurrence in Jurassic and Cretaceous terrestrial communities. All are known from terrestrial depositional settings, ranging from the delta-plain channels of the Horseshoe Canyon Formation of Alberta, to the broad meandering rivers of the Morrison and Cloverly formations of the Western Interior of the United States, to the upland meandering streams of the Two Medicine Formation of Montana (Dodson 1971; Dodson et al. 1980a, 1980b; Eberth 1987). Often found in association with diverse terrestrial, semi-aquatic, and aquatic faunal elements, these basal euornithopods also lived in a variety of climatic conditions, from well-watered semitropical to seasonally semiaridity (Allen 1998). No obvious underlying environmental restrictions or preferences are associated with these dinosaurs, or at least not with their depositional environments.
In terms of feeding, all basal ornithopods were probably active herbivores, mainly foraging on ground cover and shrubby vegetation. Browsing was concentrated within the first meter or two above the ground. In all cases, the absence of teeth along the rostralmost part of the premaxilla, the development of a keratinous beak (rhamphotheca), and a narrow oral margin suggest selective cropping (Norman and Weishampel 1991). Wear on the premaxillary teeth, produced by occlusion with the outer margins of the predentary, indicates not only piercing prehension of food but also some degree of shearing. Studies of mastication in heterodontosaurids by Weishampel (1984c) and Crompton and Attridge (1986) indicate that, like all other ornithischians, these animals chewed bilaterally. To combine this isognathic condition with a low-angle transverse power stroke (as indicated by dentary tooth morphology) all heterodontosaurids combined vertical mandibular movement with a slight degree of medial rotation or deflection of the mandible about their long axes (Weishampel 1984c; Crompton and Attridge 1986). The rotational mobility is facilitated by the restructuring of the symphysis and dentary-predentary contact into a spheroid articulation (Weishampel 1984c). Otherwise, the skull was exceptionally rigid to resist deformation during chewing.
Euornithopods also chewed using transverse jaw motion, but in a decidedly different manner. In order to achieve this transverse power stroke for effective oral processing of plant food, the skull was modified by a distinctive pattern of intracranial mobility (Norman 1984b; Weishampel 1984c; Norman and Weishampel 1985). Pleurokinesis (Norman 1984b) involved rotation of a functional unit comprising the maxilla, jugal, lacrimal, palatine, pterygoid, and ectopterygoid relative to the muzzle and skull roof by means of a diagonal hinge between the premaxilla and maxilla. When the upper and lower teeth were brought bilaterally into occlusion, opposing surfaces sheared obliquely past one another, and, as a consequence, the maxillae rotated laterally and the dentaries medially. Pleurokinesis reduced cranial induced torsional stresses generated during bilateral occlusion in the absence of an ossified secondary palate. At the same time, it permitted transverse excursion of the upper teeth relative to the lower within a primitively isognathous masticatory apparatus. Resistance to maxillary movement, necessary for shearing between occluding teeth, was presumably provided by the constrictor dorsalis musculature (or a ligamentous remnant thereof) connecting the palate (and hence the upper jaw) and braincase (Norman 1984b). In this way effective shearing of the food between the teeth could be achieved. As in the case of heterodontosaurids, the narrow rostral end of the snout in these ornithopods suggests that selective cropping of the plant parts was possible (Norman and Weishampel 1991), in contrast to the condition seen in more derived iguanodontians, which are characterized by a transversely rostrum.
Among these taxa, Hypsilophodon was long regarded as a rock-hopping or arboreal dinosaur (Hulke 1882b; Abel 1912), but this was subsequently modified. Heilmann (1927) demonstrated that the arboreality was secondary (as in flightless birds) and that this animal was originally a ground dweller. Galton (1971d, 1971e, 1974a) reviewed this topic and demonstrated that Hypsilophodon lacked the structural adaptations—in particular, the opposability of the hallux—necessary for arboreality. Rather, on the basis of limb and trunk proportions and the general gracile nature of their skeletons, ornithopods have been interpreted as agile, cursorial bipeds (Galton 1971d, 1971e; Thulborn 1972). A quadrupedal stance was probably used during foraging or when standing still. The femur was held in a slightly abducted position to facilitate a parasagittal gait and horizontal knee joints. Lever-arm mechanics suggest that femoral retraction was powerful (Galton 1969). The noncervical portion of the vertebral column was arched horizontally.
In Heterodontosaurus and Hypsilophodon, the cranial series of dorsal vertebrae turn downward (Galton 1974a; Santa Luca 1980), whereas the cervical vertebrae are dorsiflexed. Ossified epaxial tendons over the dorsals strengthened this portion of the vertebral column, but their apparent absence over the caudal series in Heterodontosaurus left the tail to move about more or less freely. The hindlimbs were highly adapted for rapid progression, displaying greatly elongate distal limb segments and fusion of elements within segments.
The forelimbs of Heterodontosaurus are also specialized, but their purported function is controversial. On the basis of the large medial epicondyle, large olecranon, and ungual flexor tubercles, Santa Luca (1980) suggested that forelimb function encompassed powerful extension and antebrachial and manual flexion. He further regarded quadrupedal locomotion as important only at slow speeds. In contrast, Paul (1987b) considered Heterodontosaurus to be a quadrupedal galloper. Although Paul (1987b) did not cite any features in support of this view, the same attributes noted by Santa Luca are equally applicable in quadrupedal animals; to this list can also be added the modified wrist and hand joints that suggest hyperextension of the carpometacarpal and metacarpophalangeal joints. However, neither of these ideas about forelimb function take into account the structure of the claws and the relations of the scapula, coracoid, and sternum to limit scapular rotation, both of which argue against rapid quadrupedal locomotion. We suggest, alternatively, that all of these forelimb features are consistent with a digging and tearing habit in the manner of mammals such as pangolins, aardvarks, and certain edentates. Heterodontosaurus may have used its large claws and powerful forelimbs for digging up roots and tubers.
Thulborn (1978) presented evidence that heterodontosaurids may have aestivated during times of environmental stress, based on the purported lack of tooth replacement and propalinal jaw action in these animals. In his rejoinder to Thulborn, Hopson (1980b) noted that heterodontosaurids had a transverse power stroke and that tooth replacement in these animals was reduced, but not lost. Thus, there is no compelling reason to believe that heterodontosaurids engaged in aestivation.
The caniniform teeth in heterodontosaurids may have been employed for intraspecific display and combat (Steel 1969; Thulborn 1974). Since caniniform teeth are thought to be present only in mature males (Thulborn 1974), they would likely have been used not only for gender recognition but also for intraspecific combat, ritualized display, social ranking, and possibly courtship (much as in extant suids and tragulids, Molnar 1977). Similarly, the development of the jugal boss (also in Orodromeus makelai and Zephyrosaurus schaffi) may be similarly interpreted.