CHAPTER TWO
In Your Next Life, You Should Be a Parasite
PARASITISM IS ONE OF THE MOST INTERESTING AND IMPORTANT topics I have ever studied. Parasitism evolved very early in the history of life—maybe even at the very beginning. The volcanic hydrothermal vent hypothesis proposes that life on this planet began in the vicinity of deep-ocean volcanic vents. The temperature there, approximately 100°C, allowed complex molecules to form from such simple building blocks as carbon, nitrogen, sulfur, phosphorous, water (H2O), and an abundance of micronutrients (iron is of particular interest in this regard). The volcanic hydrothermal vent hypothesis also proposes that noncellular life evolved at almost the same time as cellular life. Another name for noncellular life is “virus.” Viruses contain nucleic acids that allow them to direct the synthesis of additional copies of their own genetic codes and protective protein-based shells inside a cellular host. As viruses must use cellular life to replicate, they are the quintessential parasite. Parasites are life-forms that derive their capacity to replicate by using the resources of another life-form. They differ from predators, which usually consume their prey all at once. The relationship between parasite and host can last seconds (as in bacteriophages attacking bacteria) or decades (as in parasitic worms infesting large-bodied animals). Parasitism occurs within the genome of living things (e.g., transposable genetic elements) or within their cells (e.g., bacteriophages and viruses) or within their organ systems (e.g., malaria, schistosome, and nematode parasites). From all we can tell, there are more parasitic forms of life than there are free-living organisms—proof positive that this mechanism has had great success in the history of life.
It is entirely possible that the unexplained extinctions of large-bodied organisms that we observe in the history of life were driven by their microscopic parasites. We would have no direct way of knowing, but given what we do know concerning the evolutionary success of these organisms, I would bet on the parasites. I first got interested in the question of parasites and their significance to human life in 1978 while I was a graduate student in Lowell. Shortly after I arrived there to start my master’s thesis work, I met with my new adviser, Jack Mallet, who had decided to take me on as a master’s student because of my training in limnology at Oberlin. He was involved in research into the distribution of schistosomiasis in the Nile Basin. Schistosomiasis is a disease caused by infection of flatworms from the genus Schistosoma and is still a major public health concern.1 In 2019 it was estimated that there were 230 million infections (in Africa, the Middle East, East Asia, and Latin America), accounting for at least two hundred thousand deaths worldwide. The disease results from the massive number of eggs laid by the female worms. An infected individual can go into immune shock, causing the spleen to rupture, and death follows. Those who do not die suffer from a variety of conditions, such as anemia, lethargy, and lowered intelligence.
Mallet’s work was funded by the US Agency for International Development (USAID). The overall project at Lowell was under the direction of John I. Bruce, head of the Center for Tropical Disease. Bruce was the first person of African descent I had ever encountered who was involved in scientific research, let alone leading a research center. His position and prestige influenced me, specifically helping me visualize a real future in science, but I had little direct contact with him.
In our first meeting, Mallet (whom I later called Jack) outlined for me a scientific problem that he wanted tackled as the core of my master’s thesis research. In 1978, Lake Nasser was still expanding and filling after the building of the Aswan High Dam. The dam had been completed by the Soviet Union, but it became of interest to the United States after the new Egyptian president, Anwar Sadat, broke ties with the Russians.2 Lake Nasser was expanding more in area than in depth, making it the perfect habitat for the snails that are the intermediate hosts of schistosomiasis (Bulinus spp. and Biomphalaria spp.).3 The US government had Landsat pictures of the region that could be used to estimate the amount of chlorophyll in the potential snail habitats. Chlorophyll is the primary pigment used by algae (diatoms, green algae, and blue-green bacteria) to conduct photosynthesis. Measure chlorophyll levels, and you can estimate how much food there will be for the snails. Knowing food levels, you can predict the abundance of snails, and this in turn tells you how much transmission of the disease you should expect in the region. However, to make that estimate, we had to find a way to provide what is called ground truth—that is, observed or measured information—for the satellite pictures. That was the project Jack assigned me: to figure out how to measure the amount of chlorophyll from the algae encrusting the rocks in such a way that we could correlate it with pictures and thus measure chlorophyll from space.
In graduate school all the things that had been weaknesses in my undergraduate years soon turned into strengths. One of the problems that contributed to my lack of success as an undergraduate was my capacity to see through the holes in existing scientific knowledge. This meant that I often overthought my responses on exams. The creativity I developed by critiquing the weaknesses of what we were being taught was now deployed to attempt to solve new problems and generate new knowledge. I found out that I was very good at this. In addition, my graduate appointment required me to teach biology to undergraduates, which meant I had to spend more time learning the things I should have learned when I was an undergraduate, so that I could teach them. I was also spending less time on organized sports. I still played pickup basketball, with various volleyball clubs, and in some matches with the Mystic River Rugby Club in Boston. However, I wasn’t going to many practices or spending as much effort conditioning, greatly reducing the amount of time involved in sport.
Jack taught the limnology course, but Harold Asch (last living student of Clark P. Read) taught parasitology. Read was one of the pioneers of American parasitology. Legend has it that to study the Asian liver fluke (Clonorchis sinensis), Read actually infected himself to get the organisms back to America alive, although, as the story was related to me, he almost died from the experience. I don’t put much stock in that story. As an undergraduate, most of my organismal training had been devoted to plants. However, we never really spent any time on their diseases. The only one I can remember is mistletoe (Viscum album, European species), a semiparasitic species that grows on various tree species. The practice of kissing under the mistletoe during the Yule season is actually derived from the plant’s history as a pagan symbol of fertility.4
The world of parasitology opened a whole new understanding of how biology worked, as well as of its significance to human affairs. Later on, when I was tasked to teach Diversity of Life at the University of California, Irvine, my lectures were driven by what I had learned in parasitology. All living things are parasitized by something (even parasites are parasitized by other parasites). Parasitic disease had played a crucial role in the development of human societies. Notably, the Greeks moved their cities from the marshy coastal lowlands to the high altitudes to reduce the transmission of malaria.5 Indeed, as I now understand, it is precisely the things that we are best at (evolutionarily) that are making us vulnerable to parasitic disease.
In addition, evolutionary forces work in favor of the parasites. The high mortality that humans face because of parasites results from our much longer generation time compared to theirs. Our active cellular immunity allows us to produce a bewildering amount of antibodies and immune cells against pathogens, but over time the pathogens’ rapid generation times enable them to avoid this system by evolving ways to avoid immune detection. In addition, new parasites, such as SARS-CoV-2, evolved the capacity to cross over into our species, and because our active immunity had never seen them before, we had no means to fight them. Indeed, I doubt that when our species goes extinct, it will have been destroyed by nuclear war; rather, I would bet on viruses to do the job (although asteroid impact and volcanic eruption may contend for the honor). For those of you who are thinking I forgot about climate change, I have not. The most likely final blow resulting from climate change will be the spread of infectious disease. With rising global mean temperatures, the insect vectors of tropical disease will be able to complete their life cycles at higher latitudes. The last thirty years has already seen an expansion of the transmission of the insect-borne arboviruses (which cause, among others, dengue, chikungunya, and Zika fever).6 This should place us on high alert concerning the number of viruses that are lining up to become the next global pandemic.

SO WHY ARE THERE SO MANY PARASITIC LIFE-FORMS? THE ANSWER is simple: if you are not an autotroph (an organism that can generate its own energy from sunlight or a chemical means), then you must get the energy to replicate your genome from another organism. Parasitism is in many ways more advantageous than predation for small-bodied organisms in that it maintains the organism’s food source for a longer period of time. Parasitism is found in virtually every taxon of cellular life (bacteria, archaea, and eucarya). In addition, acellular life (viruses) by definition must be parasitic because they rely on the cellular machinery of other organisms to complete their life cycle. It is also important to remember that the vast majority of living things on this planet are unicellular (one-celled). We are most familiar with multicellular life—plants, fungi, and animals—but these are only a small proportion of living things. The larger-bodied forms of these groups represent a virtual smorgasbord of delights for parasitic organisms.
Some organisms are free-living and parasitic depending upon the environmental circumstances in which they find themselves. Escherichia coli, a normal component of your gut microbiome (microbial organisms living in the gut), can become a deadly parasite should it enter a cut. This is why thorough hand washing after using the bathroom is a good idea, and why the Vietcong used to foul punji sticks with human feces to ensure wounded soldiers would come down with vicious infections.7 Some organisms shift from free-living to parasitic at different life stages (facultative parasitism). This capacity undoubtedly played a role in the evolution of organisms that are always (and cannot survive without being) parasitic (obligate parasites). Some organisms shifted their mode of living to parasitism when a suitable opportunity arose. Hitching a ride on bigger organisms is a commonly observed behavior for dispersal. Many people know that remoras hitch rides on sharks (and feed off the scraps of the shark’s meal). However, such hitching has also been shown to be a potential route to parasitism for mites (arthropods) and nematodes.8 The bubonic plague caused one of the most devastating disasters in history. It was facilitated by fleas (arthropods) that might once just have been hitchhikers but that later came to feed on human blood; their bites transmitted the disease-causing bacterium Yersinia pestis. Another incentive for organisms to become parasites is that their large-bodied hosts represent environmental stability. Organisms like ourselves expend a great deal of energy to maintain our internal temperature, pH (acid level), and ionic composition (together these are called homeostasis). What better place for an organism to live (and get fed at the same time!)? And finally, turnabout is fair play. Sometimes organisms survive being eaten and then manage to evolve parasitism inside the bodies of their would-be predators. The ciliate (single-celled eukaryote) Lambornella clarkii survives being ingested by the larva of the mosquito Aedes sierrensis and then becomes a parasite of the adult.9 This is a probable scenario for the way the kinetoplastid genus Falciparum found its way to becoming one of the most lethal of all human parasitic infections: malaria.
Parasite transmission can occur by four basic means: from host to host; up the food chain (parasites in an organism that is eaten by another mature in the eater); through a vector (parasites live in a vector host that transmits them to the final hosts); and by horizontal transmission of the parasites to others within the population or through vertical transmission in the germ line (cells that produce gamete cells) to their offspring. We are becoming refamiliarized with the host-to-host mode of transmission, as COVID-19 has demonstrated how easily it can be transmitted from person to person. The classic case of humans being parasitized via the food chain are the tapeworms (Cestoda)—which is why I am not a fan of rare steaks or sushi. Diseases transmitted by vectors were mentioned earlier (bubonic plague by fleas from rats; malaria by mosquitoes). The relationship between horizontal and vertical transmission takes a little more scaffolding to explain. The best examples are given by viruses (obligate intracellular parasites). Viruses such as Epstein-Barr (EBV) can be transmitted from person to person, or they can be transmitted vertically through the germ line to one’s offspring. EBV has been implicated as a causal agent in Burkitt’s lymphoma, nasopharyngeal cancer, post-transplant lymphoma, and Hodgkin’s disease.10 Of these forms of transmission, parasites that require intermediate hosts tend to be the most complex.
As life cycles go, that of schistosomes isn’t particularly complex. The adults live in the veins of the adult mammalian host: Schistosoma haematobium (Africa, Middle East) live in the veins of the urinary bladder. S. mansoni prefer the portal veins draining the large intestine. And S. japonicum live in veins draining the small intestine. Within the host’s bladder or intestines, the female worm lives in a specialized canal of the larger male worm. Their sole behavior is copulation and egg laying. The eggs work their way through the tissues of the bladder or intestines to be released into the environment. The eggs hatch into a stage called the miracidium, a multicellular ciliated organism that lives just long enough to find an intermediate host (the snail Bulinus truncatus for S. haematobium). Once inside the snail, the miracidium develops into what is called the mother sporocyst stage. After several more generations of sporocysts, the cercariae (the infectious stage for mammals) are produced. These are shed from the snail into the water, where they wait for a suitable host (Figure 2.1).
The life cycle of the schistosomes seems relatively simple, and as a master’s student I did not realize that they represent incredibly complex problems that now form the core of modern genomic research. The National Science Foundation (NSF) lists some of these problems under their 10 Big Ideas, specifically under Understanding the Rules of Life.11 For example, the stages of the parasite’s life cycle must display dramatically different patterns of gene expression (transcriptomes). The miracidium stage is tasked solely with finding snails to infect. Most schistosomes began their parasitic career with snails, as both flatworms and mollusks existed long before primates (emerging approximately 540 million years ago and approximately 50 million years ago, respectively). This suggests that the capacity to successfully infect primates resulted from an evolutionary reorganization of or addition to its genome. The capacity to add to the genome of eukaryotes itself results from genomic parasites (transposable genetic elements) that have sequentially added copies of their own code to genomes. This additional genetic material allowed evolution to produce genes with new functions. The sporocyst stage is devoted to asexual reproduction, allowing the schistosomes to rapidly increase the number of infective units to find a primate host. Gene expression here would be dominated by the genes involved in cellular division and growth. The cercaria stage would be dominated by the expression of genes allowing them to find a human host, swimming to the host, and penetrating the host’s skin. Finally, adult worms would have gene-expression profiles associated with metamorphosis to the adult body form, uptake of food and oxygen from the human bloodstream, avoidance of the host’s immune response, and the production of gametes. Modern studies of the transcriptome (pattern of genes expressed) of schistosome parasites show that the genes expressed in the miracidium stage versus the adult stage are quite different, as expected by the different functions of these life stages.12
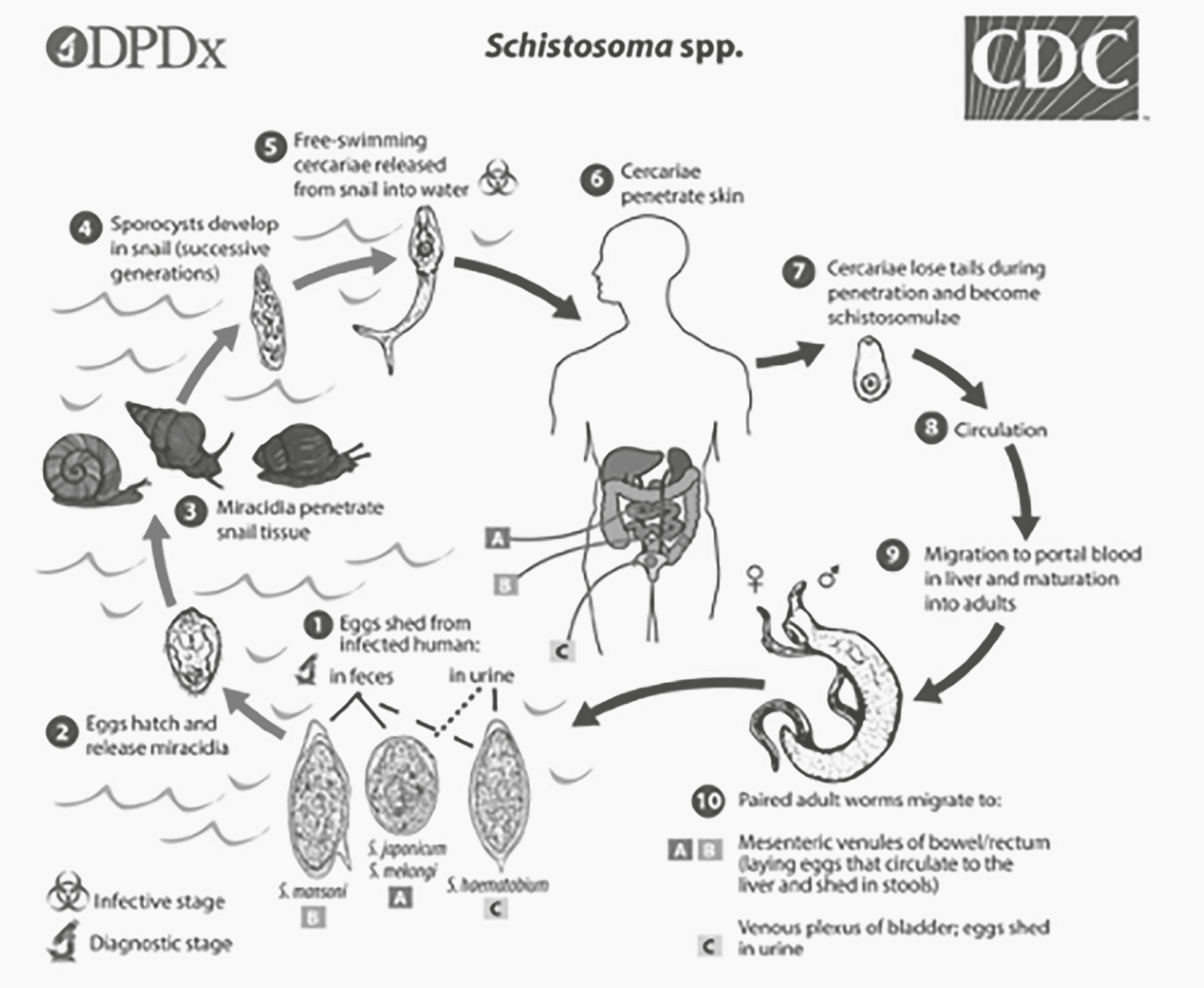
Figure 2.1. The life cycle of Schistosoma spp. Eggs hatch and miracidia locate snails (intermediate hosts). Several generations of spore-producing units (sporocysts) reproduce asexually to increase the number of cercariae that infect human hosts. Cercariae lose the ability to swim and are circulated by blood flow. This stage matures into male and female adult worms that begin the cycle again.
THE TOOLS TO INVESTIGATE THESE SORTS OF ISSUES SIMPLY DID not exist in the late 1970s. My research was tasked with learning about the factors determining snail abundance in Lake Nasser. Bulinus truncatus, the host of S. heamatobium in Africa, are herbivores that live off the microbiome (algae and blue-green bacteria) that encrusts rocks or other surfaces in lakes and rivers. My first task was separating the encrusting algae from the rocks. I collected some rocks from a nearby pond (despite common names, it is depth, not surface area, that distinguishes ponds from lakes). My long struggle with dental health suggested that a toothbrush would be the most effective way to separate the algae from the rock surface. Microscopic examination before and after the brushing showed that it worked amazingly well. I would brush, then swirl the biofilm (I didn’t know that word back then) in a beaker of methanol. One form of chlorophyll, chlorophyll a, absorbs light most intensely in the range 450–475 nanometers (10-8 m). I measured absorption between those wavelengths, which allowed me to determine the relative amount of chlorophyll (using the standard curve method). When I began to compare my field result measurements with those coming from satellite images, I observed amazingly high correlations (r > 0.90; r-value explains how well two different data sets correlate with each other). I couldn’t understand it; I felt I must be doing something wrong. I agonized over the results, as I had a meeting coming up in which I had to present my results to Jack. That night I had an extremely vivid dream. I was sitting in my linear algebra class from sophomore year at Oberlin College. Maria Zumoff (one of my favorite professors) was writing a theorem on the board. In the dream she turned and looked at me, and said, “Joe, you know why your results are so good; if vector A is orthogonal to vector B, and vector B is orthogonal to C, then it must be true that vectors A and C are orthogonal to each other!” I sat up in bed and said, “Yes, eureka, I’ve got it!” Zumoff had explained it to me via a rather obvious theorem akin to the commutative property in algebra. My results had to correlate: both methods were measuring the same thing, and thus they had to correlate.
My success with the ground truth method contributed to my being chosen to go to the Nile to collect samples of snails and determine the composition of the biotic community associated with their abundance. Community composition is a topic within ecology that is focused on understanding the relationships between plants and animals that influence the numbers of the various species within a given habitat. For example, an overabundance of predators could drastically reduce the number of snails in a given habitat, or the absence of suitable floating plant substrates might lower snail abundance.
We left for Egypt in January 1978. We spent about four weeks there traveling from the Nile Delta to Aswan collecting samples at various localities. The trip wasn’t all fieldwork with snails. It was my first opportunity to directly observe the legacies of colonialism. The most egregious thing I experienced was the way Egyptians were treated by Europeans. Indeed, this issue became a major point of contention between Jack and me. It took on dimensions reminiscent of the disagreement between Robert FitzRoy and Charles Darwin, and almost led to a fistfight between us. One evening we were drinking pretty heavily, and the conversation veered into all the good the United States was doing for the Egyptian people. I pointed out to him that I wasn’t observing that good, and Jack retreated to the “America’s the greatest country in the world” trope. I reminded him of the subordinated state that all the Black graduate students at Lowell felt and the number of times I had been called “nigger” since coming to Massachusetts. The third person with us (Jim Bailey) recognized that the tenor of our conversation was also being noticed by the other patrons of the bar, a place that felt like it came from Raiders of the Lost Ark. From the corner of my eye I could see various factions in the room reaching for blades. I think that Jack noticed the same thing, so we both realized it was better to deescalate the conversation.
The most memorable part of the trip was our return to the United States. First, the power in our hotel went out. We were on the eighth floor and had to carry all our samples down the stairs. By the time we got everything into the taxis, we had about thirty minutes to make Cairo International Airport for an international flight. This was impossible—until it started raining on our way to the airport. You may not think that rain is such a big thing, except that it rains in Egypt maybe once every hundred years! By our standards, the rain was at best a shower, but it was enough to stop departures out of the airport, so we made the flight.
We arrived at Logan International Airport in Boston on the morning of February 5. Snow was just beginning to fall. Jim and I got back to Lowell before the roads became impassible. Jack needed to call for help from an armored personnel carrier to get him back to the laboratory with our samples. In the next twenty-four hours, fifty-six inches of snow fell. The snowdrifts were so high that once we dug ourselves out, getting around was like walking through World War I trenches. It was an amazing contrast: two days earlier I was sweltering in the heat of the desert, then experienced the one rainfall of a century in Egypt, to get home just in time for the blizzard of the century in New England. Couldn’t help but feel that someone was trying to tell me something.
ON MY RETURN TO THE UNITED STATES I HAD A LOT OF TIME TO think about the implications of endemic parasitic disease in the tropics. Who gets sick? How sick? And how many survive these brutal infections? It was clear to me that the exposure to parasitic disease has never been evenly distributed in modern human societies. The poor and socially subordinated have always shouldered a disproportionate burden of infectious disease. Consider the findings of John Snow, the founder of the modern science of epidemiology. His study of the 1849 cholera outbreak in London that killed more than five hundred people showed that most of the cholera victims drew their water from the Broad Street pump. The pump had been contaminated with raw sewage—not something that happened very often in the neighborhoods of the wealthy.13 Parasitic disease has had important impacts on American society as well. The roundworm Necator americanus (nicknamed “the American killer”) evolved in Africa but was brought to the Americas with the transatlantic slave trade.14 It was first discovered in Brazil (which received the largest number of enslaved Africans) and was later found in Texas. Modern phylogenetic analysis has confirmed that Brazilian N. americanus is derived from Africa.15 To take another example: Sex workers, particularly women, have shouldered much of the burden of venereal diseases. Even today, across the world many female sex workers are enslaved by violent males. Models of HIV transmission show that reducing sexual violence against these women would reduce transmission of HIV by 17 percent and 20 percent in Kenya and Canada, respectively.16 Thus, human social factors that determine wealth and power also determine who is likely to live in the most dangerous environments and therefore likely to contract infectious disease.
If generations of individuals live with exposure to a specific infectious disease, protections against it may evolve. The classic examples of this are the antimalarial genetic variants such as sickle cell hemoglobins, thalassemias, and the Duffy allele found in various humans around the world. Adaptation only occurs if the genetic variant involved confers greater fitness (the product of age-specific survivorship and reproduction) to individuals that carry it. There is ample reason for parasitized populations to evolve resistance to these organisms. There has been a great deal of research to determine what the genomic basis of resistance to schistosome infection is for the snails that vector it to humans. A genomic region, PTC2, has been discovered in the snail Biomphalaria glabrata (vector of S. mansoni) where variants can cause as much as a fifteenfold difference in a snail’s susceptibility to being parasitized.17 Unfortunately, there are no loci discovered in humans that have such major effects. The genomic region chromosome 5q31–33 has been shown to be involved in human resistance to S. mansoni, but no major effects result from variations in this locus. Also, variants have been found that affect susceptibility to the roundworm Ascaris lumbricoides (chromosomes 1 and 13), but again, no major effects result.18 Resistance to these worms clearly has a genetic basis: heritability, or the degree that offspring resemble parents, is h2 = 0.17–0.44—that is, 17 to 44 percent of the resistance results from genes, and 56 to 83 percent results from environmental factors. So, unlike resistance to malaria, which often results from a few genetic loci of major effect (such as HbS—sickle cell trait—or G6PD deficiency), worm resistance results from many genetic loci of small effect.
Another thing that differentiates worm resistance from malaria resistance is that a small worm burden may actually help infected individuals. Thus, for most of the history of our species, humans maintained populations of flatworms and roundworms in their guts. Disease would only occur if the balance of the gut microbiome became dysregulated. These worm species would have been commensal in the human gut (as the bacterium E. coli is). These organisms were like old friends, and their dysregulation by modern hygienic practices has led to the increasing prevalence of allergic and autoimmune diseases such as asthma, psoriasis, rheumatoid arthritis, and inflammatory bowel disease in industrialized nations.19
My time in Egypt convinced me that pharmacological approaches to reducing the prevalence of schistosomiasis were doomed to fail, because the parasites themselves would readily evolve resistance to any drug deployed against them since the people would have to be continually treated for reinfection. Attempts to use chemical methods to reduce snail vector populations were doomed to fail for the same reason. Thus, the only methods that would work were the same public health measures that were used to reduce the transmission of parasitic disease in the industrialized countries: providing people with plumbing to supply clean water for drinking and bathing and instituting a public health education program to teach people about the danger of the disease.
Of course, there is a reason why this has never occurred in the underdeveloped world and why schistosomiasis is still considered a neglected tropical disease. This tragedy continues in a region of the world that, despite the poverty of its people, is one of the richest in natural resources. We are all at risk if such social injustices continue, as US history should demonstrate. Yellow fever, caused by viruses of the Flaviviridae, was brought to the Americas via the slave trade. It is mosquito-borne, and epidemics of yellow fever were particularly destructive, killing almost twenty people per day in Philadelphia in 1793. It killed indiscriminately, as persons of African descent had no greater resistance to the virus than Europeans.20 Abigail Adams wrote of the epidemic in her letters. In the summer of 1793, the federal government shut down and moved out of the city. By October the death rate was more than one hundred people per day. Yellow fever was finally controlled in the United States after Walter Reed revealed in 1901 that the virus was transmitted by mosquitoes.
MY ROOMMATE PLAYED THE ROLLING STONES ALBUM STICKY Fingers incessantly during the summer of 1978 while I was attending the Experimental Marine Botany course held at the Marine Biological Laboratory (MBL) in Woods Hole, Massachusetts. Every time I think of that summer, that music is the first thing that comes into my head. Don’t get me wrong, I always dug the Stones. My favorite of their songs will always be “Gimme Shelter,” but he almost ruined my enjoyment of them that summer.
Woods Hole is one of the most revered places in the history of American biology. It is interesting to note that the location was selected for the Marine Biological Laboratory as a gathering place for biologists partly because it promised an escape from parasitic diseases in the summertime. Another appeal of Woods Hole was that it was a place to get away from the poor people who were becoming more and more prevalent in the overcrowded cities of late nineteenth-century America.21 Biology in the United States began as a discipline in places like Johns Hopkins University, Columbia University, the University of Chicago, and the University of Pennsylvania. Of course, notably absent from this list is any place where an African American was likely to get an education, such as Howard University. In 1883 the American Society of Naturalists was formed. It limited its membership to those participating in academic research (effectively eliminating women and racially subordinated people as potential members).
Also, places like Woods Hole allowed for the study of animals and plants in their “natural” environments. The seashore had already been established as a place of such study; the most prominent American biologist of the mid-nineteenth century, Louis Agassiz, was primarily an invertebrate zoologist (despite his false ruminations on the human species and its supposed races).22 He founded the first American institute for marine zoology in 1873 on a small, desolate island in Buzzards Bay off the coast of Cape Cod. Agassiz died that same year, and the institute’s location guaranteed it wouldn’t last. On the other hand, the village of Woods Hole offered a pleasant summer climate and was only two hours from Boston by train. After prominent American biologists such as Charles Otis Whitman (and private donors) were brought together, the potential of Woods Hole as a summer home for biologists was clear. Its connection with local and national universities led to the number of biological scientists summering there growing from a few to over two hundred between 1888 and 1928. The MBL had a transformative impact on American biology. Researchers there included Jacques Loeb (artificial parthenogenesis), Frank Lillie (sexual reproduction), E. B. Wilson (geneticist), and Nettie Stevens (whose collaboration with T. H. Morgan influenced his Nobel Prize–winning work in the 1930s). The biologists who frequented the MBL became the first Americans to gain international reputations. Their work reversed the flow of American graduate students going to Europe and led to increasing numbers of European students coming to America.23 Of course, none of those students were of African descent.
I knew nothing about this history at the time. I was ecstatic when I received a letter from the MBL telling me I had been accepted for the Experimental Marine Botany course and awarded a fellowship from the Josiah P. Macy foundation to attend cost-free in the summer of 1978. The feel of the MBL in that summer was very much what its founders had intended. An international community of scientists, their families, and students occupied the houses and dormitories. I was particularly popular with the young children of the MBL, who thought I was “bionic” (after the Six Million Dollar Man TV show that was still running on NBC). I would pull off tricks like hopping over three-foot-tall fences from a standstill or leaping over the hoods of Volkswagen Beetles with a little bit of running start. I always prefaced these tricks by reminding the kids that I was bionic, so they shouldn’t try these things themselves.
My class included two other African Americans, both women from Texas (Figure 2.2). We were clearly among the first African American students to be part of the MBL summer courses in some time, as I noted on a tour of the summer-class pictures that hung in one of the main campus buildings (I remember it being the Lillie Building). I could detect no one in those pictures who appeared to be of African descent until I ran into a dark-skinned individual in the 1920s, whom I suspect was Ernest Everett Just. Just was twenty-six years old when he started out there as a research assistant to Frank Lillie (in about 1909). The story of Just’s time at MBL is well told in Kenneth Manning’s Black Apollo of Science.24 My copy was given to me by one of the most important mentors in my life, Beverly Rathcke. When I read his biography for the first time, I had no idea how closely my life in science would follow his. Just was one of the greatest scientists of his age, yet eugenicist Charles Davenport would describe him as being a man of “[not] any exceptional brilliancy or initiative.”25

Figure 2.2. Students enrolled in Experimental Marine Botany, 1978. Asterisks indicate those students who completed a postcourse research program.
Source: Trustees, The Marine Biological Laboratory eighty-first report, for the year 1978: Ninety-first year, Biological Bulletin 157, no. 1 (1979): 1–103.
Experimental Marine Botany was primarily taught by Jerome Schiff (Brandeis University) and Harvard Lyman (Stony Brook University), and fieldwork was taught by James Fiore (Suffolk University). This course cemented my love for and dedication to research. One of our first laboratory experiments involved isolating and purifying a chlorophyll-binding protein from the red alga Chondrus crispus. Chlorophyll-binding proteins had been isolated from red algae before, but no one had ever done so from this particular species. The laboratory ended at 5:00 p.m., and everyone in the course except me stored their materials in the refrigerators and adjourned for dinner. The next morning, when Lyman came in to set up the laboratory, he found me running my sample of the extracted chlorophyll-binding protein through the spectrophotometer. He asked me if I had been there all night, and of course I had. To say the least, he was impressed.
I was in my element at the MBL. The laboratories and library stayed open 24/7. I had the most fun (and encountered the most danger) with Jim Fiore. Marine algae did not just walk into the laboratory to be analyzed; they had to be collected. We collected them by skin diving (for me, snorkel and fins; I never managed scuba certification). I was not a strong swimmer, but with the buoyancy of salt water and a snorkel I was okay. Jim, I, and a few other students did the bulk of the collecting for the course. In retrospect, we did quite a few dangerous things collecting materials. The most memorable was on our journey to Black Rock in Buzzards Bay. This “island” is located at the point where the gently sloping continental shelf gives way to the open ocean. On the continental shelf side, the currents were such that you could easily swim back to the rock. On the ocean side, it was quite a different matter. Jim and I maneuvered around to the ocean side, pulling ourselves along on the seaweeds anchored to the rock. If we had let go, we would have been swept out to the open ocean; swimming back would have been impossible, and we would have had to be rescued by boat. Needless to say, we held on tight, collected the samples, and made it back. However, the most dangerous thing I ever did with Jim was to have drinks together in Little Italy in the North End of Boston. This was two years after the busing riots in Boston, and racial tensions in the city were still at a fever pitch. The entire time I was at Lowell, the only African American radio station in Boston went off the air at sundown to protest the ongoing racism in the city.
I first met Harvard’s Stephen Jay Gould at MBL, on August 4, 1978. He was the guest lecturer, and he spoke on his recently published Ontogeny and Phylogeny.26 The core of this book is to show why German biologist Ernst Haeckel’s recapitulation theory, “ontogeny recapitulates phylogeny,” is wrong. Haeckel thought that the developmental stages of organisms revealed their immediate evolutionary history, in the sense of the progressive icon of evolution. This commonly held but mistaken view sees evolution as proceeding in a linear progression, rather than in the branching pattern of common ancestry. I was one of the graduate students who had the opportunity to have lunch with Gould. In that conversation, I told him of my MS thesis research, and I am pretty sure that he was the one who informed me that Richard Lewontin also had an interest in the evolution of the snails responsible for vectoring schistosomiasis in Africa. I can’t be sure if it was his idea or mine to contact Lewontin about my potentially applying to Harvard to complete a PhD in evolutionary biology.
Gould had major impacts on my career and thinking beyond this meeting at MBL and my decision to apply to Harvard. Not the least was his The Mismeasure of Man, published in 1981.27 This book was one of the first that I read to lay out a case for the significance of structural racism influencing the way people thought in science. Over the years, I would periodically run into Gould at scientific meetings. He and I were not friends; he seemed to remember having met me but could never quite place my name.
During the spring of 1979 I applied for a National Science Foundation Graduate Research Fellowship (GRF). The GRF required that you identify potential PhD mentors. As I was living in Massachusetts, I wanted to attend Harvard. I was already spending a lot of time on the campus at the Museum of Comparative Zoology (MCZ), which housed the library I was using for my research and the offices of the Department of Integrative Biology. I made an appointment to see Richard Lewontin concerning my interest in attending Harvard University and working on the evolution of the genus Bulinus in Africa. These snails are interesting for more than their role in disease transmission. They are part of a polyploid series. B. tropicus was diploid (2N)—that is, each individual had two copies of each of its genes, due to biparental inheritance (the same as we do). B. truncatus was tetraploid (4N).
I only applied to two PhD programs: Harvard and Michigan, not bad choices at the time. My interest in Michigan stemmed from the fact that researchers there were collaborators with Mallett and Bruce on their schistosomiasis research in Egypt. Michigan invited me out for an interview. As soon as I set foot on the campus, I was already leaning toward Michigan. In particular, a Big 10 campus was in stark contrast to the gray, overcast, and aged features of my campus in Lowell. But as before, fate made my decision for me.
First, I was informed by the NSF that I had been awarded a GRF fellowship, one of the most prestigious awards a graduate student can earn. In 2016 I was cited as one of the “Sensational Sixty” commemorating sixty years of the NSF GRF. That meant the NSF chose me to represent the GRF for the year in which it was awarded to me, 1979. Universities gobble up NSF GRF winners, not just because they acquire the winner of a prestigious award, but also because they are getting a free student (the NSF paid for tuition, research assistantship, and cost of living). Thus, I suspect I am the only person in the history of the GRF to be rejected for admission to a graduate program in the same year the award was made.
Shortly after receiving my GRF award letter, I got a very strange letter from Harvard University. I am only paraphrasing it here because I no longer have the letter, but as I recall, it said, “Dear Mr. Graves, We find that your credentials are entirely consistent with admission to the Department of Integrative Biology for the PhD degree. However, we can find no faculty member who is willing to take you on as a graduate student. Therefore, we are denying your admission.” I was overwhelmed with the strangeness of this response. I am absolutely sure that in my personal statement I mentioned my interest of working with Lewontin, as every graduate applicant is required to identify possible PhD mentors.
I cannot know what happened in the admissions committee, but I can say what didn’t happen. Whoever was on that committee had no idea about or was simply unconcerned about the absence of African Americans in careers related to evolutionary biology. One would think that a person who knew something about or cared about the issue would have tried to find someone willing to advise the only African American student entering the program. Certainly, had there been any such students in the department, I would have run into them, given the amount of time I spent in that building. I do remember European American students coming back and locking their offices or removing valuables from sight when I walked through the common area, so my guess is they were not used to Black faces being around.
At the end of the day, the Harvard rejection didn’t matter, for I was already decided upon going to Michigan. In terms of my subsequent intellectual development, it was the best decision I ever made.