CHAPTER TWELVE
America’s Biggest Lie
Slight Return
IN 2016 I ONCE AGAIN FOUND MYSELF DRAWN DIRECTLY INTO the debate over genes, race, and intelligence. As in 1994 it began with a phone call, this time from the Campus Debating Society of Williams College. Williams is a private, residential, liberal arts college of about two thousand students located in the Berkshires about 135 miles northwest of Boston. The campus program entitled Uncomfortable Conversations had invited Charles R. Murray (coauthor of The Bell Curve) to speak on campus.1 The Campus Debating Society, associated with the Phi Beta Kappa Honorary Society, had arranged for Murray to debate with me on his ideas concerning the genetic foundations of intelligence and their role in structuring societies. It was already late in the afternoon when I received the call and agreed to take on the debate. However, arrangements were made to get me on a flight the next day (March 9, 2016) to Boston. I rented a car and drove to the campus in time for the debate. When I arrived, I was hosted for dinner by one of the members of the debating society and informed that there would be no debate. It is unclear whether Murray had ever agreed to a debate format or whether I had simply been misled into believing there would be a debate. Instead, I would deliver a lecture just before Murray’s scheduled lecture. Then Murray would take the stage and deliver his lecture. Then I was to be given a privileged position in the question-and-answer period to pose any comments or questions directly to Murray.
By now I had spent considerably more time thinking and writing about the scientific difficulties with race and IQ theories than I had in 1994. The title of my lecture was Race, Genomics, and IQ: Slight Return. The lecture material was drawn from my books (The Emperor’s New Clothes and The Race Myth were now considered classic works of scholarship) as well as from four recent works I had written in response to Nicholas Wade’s 2014 book, A Troublesome Inheritance: Genes, Race and Human History.2 Wade was a former science writer for the New York Times. He once chaired a panel session I participated in at the American Museum of Natural History in New York City (December 2001). When he introduced the members of the panel, he managed to correctly recite the disciplinary credentials of all of the other panel members (who were all white males), but he described me as “a philosopher who had written about race.” That was not the description on the materials provided him to introduce the members of the panel, so when it was my turn to speak I made sure I informed the audience that my PhD was in evolutionary, environmental, and systematic biology, not philosophy. A year or so later, Wade also reported in the New York Times about a very important research paper that examined the apportionment of human genetic diversity.3 He argued that a paper by Noah Rosenberg and colleagues, which used a clustering algorithm entitled STRUCTURE, was modern confirmation of the classification of humans into five continentally based biological races: African, Eurasian, East Asian, Pacific Islander, and Native American.4 There are a lot of problems with the assumptions behind the STRUCTURE algorithm, as well as with the sampling of the individuals used to generate the results of the paper. The most problematic assumption is that there were once five ancestral human races, from which modern humans derive their genetic variability. The characteristics of these ancient races were estimated by using genetic information from modern people (e.g., genetic samples from modern people called Yoruba, or people of European descent from Utah). Thus, the assignment of modern people’s genetic information to ancient parental races, based on modern genetic samples, was blatantly circular logic.5 In addition, sampling people from discrete points of a continuum of genetic diversity cannot help but produce clustered results. It would be like repeatedly sampling the number line between 0 and 100 at discrete points, for example, repeatedly drawing numbers near 10, then repeatedly drawing numbers near 40, and in the vicinity of three other widely dispersed numbers (60, 70, 90), and then claiming that there are only five clusters of numbers on the number line from 0 to one 100!
Also, during the first decade of the twenty-first century new DNA sequencing technology (next-generation sequencing, NGS) made possible much more sequencing of individual genomes. These new data produced genome-wide association studies (GWAS), a new way to examine the relationship between genetic variation and physical traits. NGS revealed that the genomes of any two persons chosen randomly in the human species share about 99.9 percent of their genetic code (nucleotides A, T, C, G). There are 3.3 billion nucleotides in the human genome, so 0.1 percent is enough to make every person unique. Differences in the code appear between any two people about every three hundred to four hundred bases. These differences are called single-nucleotide polymorphisms (SNPs). Other kinds of variants also exist, such as copy-number variants (CNVs). CNVs exist because much of the human genome consists of repeated sequences; as a result of insertion or deletion events, individuals differ in the number of specific repeated segments. Finally, it is important to realize that in eukaryotic organisms (cells with a nucleus and linear chromosomes), like us, the vast majority of the DNA in cells does not code for the proteins that make up the organism. In humans, only about 1.5 percent of our DNA codes for protein, and an additional 5 to 7 percent of our DNA is involved in regulating the expression of our genes.6 These segments are the only ones that face natural selection in ways that we currently understand.
Understanding the relationship between the genome and the physical traits it produces is one of the NSF’s big questions,7 and it is the core research question that has shaped my career from when I came in out of the cold onto the campus of Wayne State University in 1985 until now. Over the thirty years leading up to the quasi debate with Murray at Williams College, I had gotten an intimate understanding of just how complex this relationship is for any organism, let alone humans. As an experimental evolutionist I understood exactly why attempting to find a causal link between genetic variation and racialized differences in intelligence was a fool’s errand. However, calling this project a fool’s errand is wrong, because very intelligent and often evil men were behind it. (Remember, “figures never lie, but often liars figure.”) Such attempts once again remind me of the words of Ian Malcolm in the movie Jurassic Park: “Genetic power being the most awesome force ever unleashed upon the planet, but you wave it around like you found your father’s gun.”
GWAS is one of the most important modern techniques available to study the relationship between genes and the physical traits they produce. The method looks for one of five possible statistical relationships between genetic variants and a physical trait:
1. Direct causation: Variant A causes physical trait B.
2. Epistatic effect: People who display physical trait B are more likely to survive and reproduce if they have variant A.
3. Population subdivision: The population has several subpopulations, and physical trait B and variant A happen to be at high frequency in some of the subpopulations.
4. Type I statistical error: Because GWAS studies test large numbers of genetic markers, by chance alone 5 percent will be significant at p = 0.05 and 1 percent will be significant at p = 0.01.
5. Linkage disequilibrium: Variant A marks a chromosome segment that contains a genetic marker that actually causes physical trait B.
GWAS studies work best for groups that display very large physical differences between them. At the time of the Williams College talk, I had reunited with my postdoctoral mentor Michael Rose to study the genomic basis of long-term differences in life history (reproductive output, life span) in his postponed-aging Drosophila melanogaster stocks.8 We were particularly interested in testing Stephen Jay Gould’s notion of what would happen if “the tape of life” were to be replayed.9 Our question involved repeating some of Michael’s classic experiments and determining whether the same physical traits would evolve and whether parallel genomic changes would underlie them. By the time I headed off to Williams for my lecture, I already knew the answers to these questions: yes, the same traits would evolve, parallel genomic changes did occur, and their genomic foundations were quite complex. Thus, remembering that life span is a complex trait, just as cognitive function is a complex trait, I was prepared to discuss the foolishness behind Murray’s claims about simple relationships between genomic variants and human intelligence (let alone racialized genetic differences in this complex trait).
I began my lecture at Williams by dismantling the idea that our species has biological races. This is a crucial point, as racialist psychometry uses some variant of the claim that there are three or five races. You may remember that in The Bell Curve, Herrnstein and Murray presented a figure showing “Black” and “white” differences in performance on the AFQT. Of course, genetically based differences could exist between groups who are socially defined as Black and white even if they are not biological races. On average, the recent ancestry of Blacks (African Americans) is 84 percent African and 16 percent European, and of whites (European Americans) is 98 percent European and 2 percent African.10 However, most of an individual’s ancestry is determined by genetic variants in the noncoding portion of the genome. With regard to the coding portion of the genome, natural selection has retained a great deal of uniformity in human populations. A recent study of local adaptation in humans shows evidence for differences between human populations for traits such as lactase persistence (the ability to digest lactose sugar into adulthood), malaria and trypanosome resistance, skin color, hair type, and high-altitude adaptation.11 Furthermore, these adaptations are not racialized; lactase persistence is found in northern Europeans and eastern Africans; malaria resistance, which occurs by several different mechanisms, is found in western and central Africa and along the Mediterranean (e.g., in Greece), on the Saudi Peninsula, and in India. Skin color variation is associated with the north/south latitudinal cline of solar intensity (not with the three- or five-race schemes). Furthermore, of all these local adaptations, only one is a polygenic complex trait, height. The evidence for selection for taller height comes from one region of northern Europe.12 So the problem for racial theories of intelligence is not to show that human populations differ in the frequency of various genes (as they certainly do) but to show that they differ in the frequency of genes that matter for creating differences in intelligence.
The next portion of the lecture addressed the definition of intelligence. The concept is problematic, especially from the standpoint of genetics. First, to be quantifiable, a trait must be able to be reliably defined; second, the trait must be measured in an unbiased way. Intelligence fails on both accounts. There is no agreement among scholars on the meaning of intelligence. There are three commonly used definitions:
1. CHC theory suggests the existence of three hierarchically linked strata of intelligence: narrow abilities, broad abilities, and general ability (referred to as g). The second stratum, broad abilities, includes fluid ability (called g-f), which allows one to cope with novelty and to think rapidly and flexibly, and crystallized ability (called g-c), which refers to the general store of knowledge.
2. Multiple intelligence theory suggests seven categories of intelligence: linguistic, mathematical, spatial, musical versus bodily-kinesthetic, naturalist, interpersonal, and intrapersonal.
3. Triarchic theory subdivides intelligence into three areas: creative, analytical, and practical.
The ideas in The Bell Curve were organized around the CHC theory.13
Whatever intelligence is, it must be related to brain function. The function of the prefrontal cortex, the neocortex, and the superior parietal, temporal, and occipital cortexes all contribute to cognition. The subcortical regions of the brain, such as the striatum, and the integration of the parietal and frontal lobes are especially important. The complexity of human cognitive function is shown by the fact that at least 84 percent of the twenty thousand genes of the human exome are expressed in the brain.14 Compare that to the 46 percent of genes that are typically expressed in other tissue types. However, even this is an underestimate of potential genetic contributions to cognitive function, as a wide variety of physiological systems outside the brain contribute to cognitive function. For example, any genetic variant reducing blood flow to the brain would potentially have a negative impact on cognitive function. And all genetic influences on cognitive function are dependent upon their interaction with a large number of environmental factors (e.g., nutrition; medication; chemical exposure; lifestyle practices such as smoking, drinking, or exercise; circadian rhythm, whose dysregulation has powerful consequences for mental function; epigenomic changes; and the individual’s microbiome).
So the notion that we can identify a small number of candidate genes that would capture the majority of the variation in the “normal” range of human intelligence is absurd. Clearly there are a number of mutations that occur in specific genes that reduce intelligence. Well-known cases include the translocation of chromosome 21, which causes Down’s syndrome, or the 47,XXX chromosome condition, which may cause mental disability.15 These are usually mutations of high penetrance with multiple pleiotropic effects. (Penetrance in genetics is the percentage of individuals of a given genotype who will display the phenotype associated with a given mutation. For example, the mutation for polydactyly, having more than five fingers or toes on a hand or foot, does not show up in everyone who inherits the gene.) These conditions are rare (the translocation of chromosome 21 occurs in about one out of 1,200 live births) because of a population-genetic mechanism called mutation/selection balance: their negative effect on survivorship and reproduction causes such variants to be removed from the population until their frequency is approximately equal to the rate at which they reappear by random mutation. Thus, the brain, like any other organ, is relatively easy to mess up but very difficult to improve.
In the next section of the lecture I discussed the established science of GWAS for complex traits, using the example of height. The high heritability of this trait (the degree that offspring resemble parents, h2) is well established. However, GWAS studies conducted during the 2010s could only explain a small fraction of its heritable variation. That fraction resulted from the small effect of individual variants, low frequencies of the various variants within a population, the number of individuals studied, the density of genetic markers along the chromosomes, and the rate at which linkage disequilibrium diminishes with map distance.16 Linkage disequilibrium results when two genes are located close together on a chromosome but natural selection is acting on one of them but not the other. If natural selection increases the frequency of the first gene, it will also increase the frequency of the second because of its proximity to the first. The map distance refers to the distance between the two genes on the chromosome. It is determined by the frequency at which crossing over (during prophase in meiosis) occurs between them. The closer they are to each other, the fewer the number of crossover events between them. Such genes are said to be linked. The last issue is important because some of the SNPs discovered by GWAS techniques are really only marking a portion of the chromosome where the SNP that actually causes the effect is found. Consider a study in which a sample of 180,000 individuals of European descent had an h2 for height of 0.80. There were 180 SNPs identified with p < 5 × 10−8; together, they only accounted for about 10 percent of variation in the trait.17 This means that each significant SNP accounted for 5.5 percent, or 0.00055, of the variation in height explained by these genes. What is most important is that the remaining 90 percent of the variation in the trait was not explained by the 180 SNPs. This general problem is called missing heritability.
In the previous portion of the lecture I had established that for a relatively noncomplex polygenic trait, such as height, the present methods of assigning genetic variation to a physical trait had performed very poorly. Next, I addressed how well GWAS had performed to uncover specific genomic variants associated with variation within the normal range of intelligence. Thus far, GWAS for intelligence has seen little success. One of the first studies conducted a genome-wide analysis of 3,511 unrelated adults of English and/or Scottish descent utilizing 549,692 SNPs.18 These individuals’ intelligence was measured using a variety of tests (such as Raven’s Progressive Matrices, verbal fluency tests, and logical memory tests). The results suggested that 40 percent of the variation in crystallized-type intelligence and 51 percent of the variation in fluid-type intelligence between individuals was explain by linkage disequilibrium between genotyped common SNP markers and unknown causal variants. However, the SNP data only predicted about 1 percent of the variance of crystallized and fluid cognitive intelligence. Furthermore, none of the genotyped SNPs met the standard value used to determine that a SNP has a significant effect on a given physical trait. One gene-based association with fluid intelligence did show a significant effect, but the result did not replicate in another sample, the Norwegian Cognitive Neurogenetics sample. This lack of replication is significant because, given the genetic similarity of English/Scots to Norwegians, if the gene really was playing an important role in determining variation in this trait, it should have done so in both groups. The study concluded that while a substantial proportion of individual differences in human intelligence was due to genetic variation, it was caused by many genes of small effect.
Putting it numerically, about 7.3 × 10–7 (or 0.00073 percent) of the variance in cognitive function was explained per SNP.
A better way to find the genetic variants that are most important in determining an outcome is to examine groups that differ a great deal in the specific trait in question. For example, our ability to identify SNPs associated with life history variation is directly related to the fact that we used experimental evolution techniques to differentiate our groups.19 This idea was deployed in a study that used GWAS to examine exonic variation associated with extremely high intelligence.20 Exons are the portion of genes that code for proteins. Looking for SNPs in these portions of the genome is more likely to produce results that are actually associated with the physical trait under study. The study selected high-IQ individuals from the Duke University Talent Identification Program (TIP), using the top 1 percent of the group, with IQ scores of about 176. Exome array genotyping was done for 1,759 individuals who reported their ethnicity as white. The control group was generated from the Minnesota Twin Family Study, 3,253 individuals, who also identified as white, with IQ scores distributed between 70 and 150. This study too produced underwhelming results. It identified one nonsynonymous SNP in the PLXNB2 locus, whose gene product has been associated with neuronal migration, explaining 0.16 percent of the variance in IQ between the controls and the high-IQ group.
Finally, I discussed another study, which utilized fifty-four thousand adults of European descent, that reported a GWAS accounting for only 1 percent of the variance in intelligence in that cohort.21
After my lecture concluded, Murray delivered his. Much of what he said replicated his ideas in The Bell Curve. Some of it was new material that would contribute to his 2020 book on human diversity.22 As I had been promised, I was allowed to direct questions to him before the rest of the audience did. I reiterated the main points of my own lecture: the fact that human populations are not apportioned into the socially defined races that are the targets of psychometry; the absence of any evidence from modern GWAS for specific genomic variants associated with greater intelligence found in any population, let alone differentiating any populations by intelligence; and finally, the lack of a credible evolutionary scenario by which these genetic differences in intelligence could have come about. Murray is not a geneticist, so some Murray supporters in the audience who were geneticists responded to my technical questions (albeit very inadequately). Murray himself could only argue that within the next five years or so adequate GWAS of intelligence would locate the missing variants. In response to that remark, I offered to debate him on the question of the genetic architecture of intelligence, in any venue of his choosing, when such evidence should emerge. That was over five years ago as of this writing, and he has not yet agreed to that debate.

DURING THE SUMMER OF 2016 I ACCEPTED AN OFFER FROM Harvard University to speak in their lecture series celebrating the 150th anniversary of the founding of the Peabody Museum of Archaeology and Ethnology. At the time I did not realize how great an honor was being bestowed on me. It turned out that my lecture, delivered on September 27, 2016, was the first in the series, which included lectures by many great minds. (It was Harvard; what would you expect?) The museum was founded in 1866 and is one of the world’s oldest museums devoted to anthropology. It is also the resting place for Louis Agassiz’s daguerreotype images of seven enslaved people. Taken in Columbia, South Carolina, in 1850 and rediscovered in 1975, the fifteen detailed images show front and side views of seven largely naked persons, displayed the way a zoologist might show nonhuman animals (Figure 12.1).23
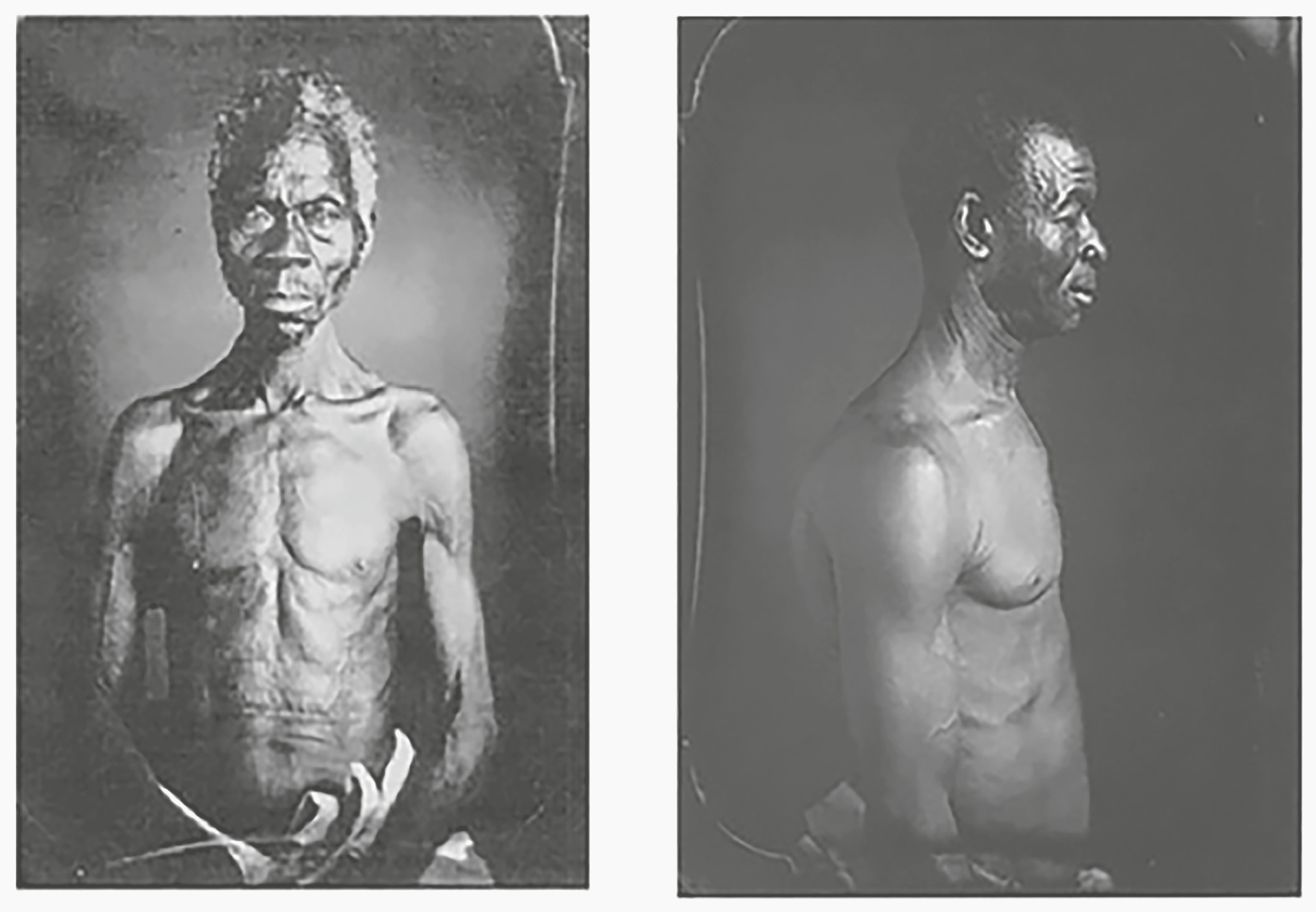
Figure 12.1. Two daguerreotypes of enslaved persons created for Louis Agassiz. They are a slave named Renty (left), a native of the Congo; and Jack (driver) (right), Guinea. The daguerreotypes were taken by J. T. Zealy for Louis Agassiz, March 1850, at the plantation of B. F. Taylor, Esq. of Columbia, South Carolina.
If you have read Gould’s The Mismeasure of Man or my The Emperor’s New Clothes, then you know something about Louis Agassiz’s racial ideology. Agassiz was one of the greatest minds in mid-nineteenth-century biology. He was born into a wealthy Swiss family. His father was a minister and his mother the daughter of a prominent physician. He was a scientific prodigy and became a champion of catastrophist geology. He published his theory of the Ice Ages in 1840.24 Agassiz’s scientific accomplishments, made possible by his considerable intellectual and physical abilities, made him a very desirable commodity in the world of university-associated science. By the mid-1840s he had made up his mind to seek an appointment at a university in the United States. His American colleagues put together a lecture tour for him designed to bring about just such an appointment. He arrived in the United States in 1846 with an itinerary of the nation’s most prominent centers of learning: Boston, New Haven, New York, Albany, Princeton, Philadelphia, and Washington.
Agassiz’s first experience with a person of African descent occurred on his arrival in the United States. Gould recounts Agassiz’s guttural disgust with the persons of African descent he encountered.25 That feeling played a role in his later ideas concerning the separate creation of the human races. Indeed, his decision to become one of the leading theorists of mid-nineteenth century polygeny was both a career move and a political choice. Some polygenists (like Agassiz and Samuel Morton) were special creationists and Christians. Thus, this brand of polygeny adhered to the theory of the pre-Adamite races. Pre-Adamism is the idea that God created several first pairs for the species of humans. The descendants of Adam and Eve gave rise to the Europeans, who were in reality a separate species (not race) from the other inferior species (Africans, Asians, etc.). Polygeny was widely supported by the southern slaveholders as it held that Africans were not members of the same species as Europeans. That is, Agassiz chose to insert himself into the racial crisis of the 1850s just as modern intellectuals choose to insert themselves in issues that curry favor with the nation’s ruling elites.26 As early as December 1847, he discussed race with the members of the Literary and Philosophical Society in Charleston, South Carolina. However, John Bachman (a minister and slaveholder) and Thomas Smyth were unconvinced by Agassiz’s polygenism. Each wrote a book formally answering him.27
So for Agassiz the daguerreotypes he commissioned had both a scientific and a political purpose. Scientifically, he wanted to use them to demonstrate the physical differences between Black Africans and white Europeans. Politically, he wanted to use them to demonstrate the latter’s superiority over the former.28
In The Emperor’s New Clothes I described one of the key factors that led Darwin to decide not to include a discussion of humans in On the Origin of Species.29 In his classic essay “Darwin’s Delay,” Gould discussed reasons for Darwin’s hesitancy in publishing On the Origin.30 Darwin’s concerns centered on the materialist foundation of natural selection and the way it might be seen as a challenge to religion. However, there was another aspect of Darwin’s theory that he and his mentor Charles Lyell knew would probably cause a bigger stir than materialism: the idea that all humans shared a common descent. The Victorian English were assured of their superiority to all other races, so the idea that they shared descent with Africans would not have been well received. Lyell felt that teaching such an idea would guarantee expulsion from the university. Darwin was convinced that Agassiz would “throw a boulder” at him.31 (The boulder reference, of course, resulted from Agassiz’s association with Ice Age geology. It is from this quotation that I decided to name and organize my lecture “Smashing Agassiz’s Boulder.”)
I began my talk with a discussion of the Kuhnian model of the structure of scientific revolutions. Nonscientists rarely think much about how science is done and how scientific ideas change. On the other hand, many practicing scientists are not too concerned with the “science of science,” which concerns itself with the social and cultural factors that influence people’s choice of problems to research. Those who do concern themselves with this process in science are generally mired in the great man model of history, which in science plays out as the idea that a genius person (usually a white male) came up with a great idea and that everyone recognized how great it was and accepted it right away. This model has some usefulness for understanding the progression of pre-twentieth-century science, but even in that era, it failed to capture the social dynamics moving scientific ideas. Thomas Kuhn’s model helps us better understand in greater detail the drama associated with scientific change (even in Darwin’s day.)
For Kuhn, all scientific research is associated with a core organized idea (a paradigm).32 Prior to Darwin and Wallace, biology’s core paradigm was still special creation (by supernatural means). The science associated with (and that doesn’t challenge) the core paradigm is called normal science. Normal science tends to be dominated by taking measurements and cataloging things. However, as normal science proceeds, observations and measurements accumulate that just don’t fit the core paradigm. At first, these things are shrugged off (like the fossils of dinosaurs, which were the remains of organisms that clearly no longer existed on the planet). However, more and more of these anomalies begin to accumulate, so modifications of the core paradigm are made. For biology, the modification was catastrophism, the idea that there had been successive special creation events, until God finally settled on the organisms that currently occupy our planet. However, when these modifications of theory continue to poorly explain the observations, the paradigm eventually enters a crisis. It is during periods of crisis that new hypotheses emerge; the proponents of these new ideas are the revolutionaries and the defenders of the old paradigm are the reactionaries. So as much as Darwin didn’t want boulders thrown at him, his ideas meant that a crisis was inevitable. The revolution was coming, and the reactionaries could not stop it because evolution by means of natural selection was a much better explanation of the facts than any variant of special creation.
However, having participated in a scientific revolution myself (of the evolution of aging), I can attest that the superiority of an idea doesn’t guarantee it must win in the contest of worldviews. As Darwin struggled to complete On the Origin of Species, Agassiz was ascending to the status of icon in both the European and American scientific communities. His allegiance to the slaveholders and aristocrats certainly helped in that regard. Darwin was the child of a wealthy family, but he and many members of his family held abolitionist views.33 Table 12.1 contrasts the views of Agassiz and Darwin. Yet despite Darwin’s many initial disadvantages in contesting Agassiz and the polygenists, his ideas would eventually demonstrate their superiority. As a result he was propelled into a leadership position in British science, eventually to replace Lyell as the leading person in British natural science. Once Darwin achieved this status, he confidently attacked polygeny in The Descent of Man, and Selection in Relation to Sex, published in 1871.
Table 12.1. Worldview of Louis Agassiz versus that of Charles Darwin
| Specific attribute | Agassiz | Darwin |
| Geology | Catastrophism | Uniformitarian |
| Anthropocentrism | Humans the center of creation | Humans one of many species molded by natural selection |
| Species | Created for distinct periods/zones in the history of life | Descent with modification |
| Political values | Conservative | Progressive |
| Personality | Gregarious/public | Introverted |
| Human races | Separate species | One species |
| Racial hierarchy | Europeans superior to all others | Emotion/intellect similari |
| Slavery | Abolitionii | Abolition |
Sources: Desmond A and Moore J, Darwin: The Life of a Tormented Evolutionist (New York: Norton, 1991); Irmscher C, Louis Agassiz: Creator of American Biology (Boston: Houghton Mifflin, 2013).
After comparing Agassiz and Darwin, I concluded my lecture with a discussion of what neither Agassiz or Darwin knew about population genetics, human genetic diversity, and the migratory history of the human species. When the lecture concluded I fielded questions concerning my talk, but none from David Reich (a Harvard geneticist), who was in the audience. I would only learn of his disagreement with my conclusions when he published his 2018 book Who We Are and How We Got Here.34 On March 23, 2018, prior to the publication of the book, an excerpt from Chapter 11 appeared in the New York Times under the provocative title “How Genetics Is Changing Our Understanding of ‘Race.’” I say “provocative” because over the course of my more-than-thirty-year career, I had not yet encountered any new developments in genetics that had really altered the scientific understanding of race. I was not surprised when he began with an assault on Richard Lewontin. He claimed that since the publication of Lewontin’s work in the early 1970s on the amount of genetic variation within and between human populations, an “orthodoxy” had formed among scientists denying the significance of race.35 The primary purpose of the “orthodoxy,” he claimed, was to deny the existence of meaningful biological variation between human populations:
But over the years this consensus has morphed, seemingly without questioning, into an orthodoxy. The orthodoxy maintains that the average genetic differences among people grouped according to today’s racial terms are so trivial when it comes to any meaningful biological traits that those differences can be ignored.36
Reich was reiterating claims made by Armand Leroi some thirteen years earlier in an opinion piece also published in the New York Times.37 I responded to Leroi’s claims in an essay published by the Social Science Research Council.38 In that essay, I addressed Leroi’s mischaracterization of Lewontin. Lewontin’s seminal finding that there was more genetic variation within a given human group than there was between them was not a claim that no significant genetic differences existed between human populations. What it did mean was that genetic variation could not be used to unambiguously define human groups. Clearly Norwegians have a higher frequency of the blue eye allele than Nigerians do, but that doesn’t mean Norwegians and Nigerians should be classified into different racial groups.
Reich’s error in 2018 was the same as Leroi’s in 2005. Shortly after Reich’s piece appeared in the New York Times and before his book was published, Jonathan Kahn (of the Mitchell Hamline School of Law), Alondra Nelson (of Columbia University and now deputy director for science and society in the Biden administration), and I sent a statement countering its claims to the New York Times. Sixty-seven scholars, ranging from geneticists to sociologists, signed on to the statement. I wrote the bulk of the genetics argument in the piece, which opposed Reich’s formulation of race and its relationship to human society. The New York Times refused to publish it, so the piece was eventually published in the popular news outlet BuzzFeed on March 30, 2018, entitled “How Not to Talk About Race and Genetics.”39
Because our response was published before Reich’s book was available, I did not know that in Chapter 11 he had called me out by name as a defender of the orthodoxy. He specifically referred to my lecture at Harvard. In his book he was particularly interested in my comments concerning the difference between complex traits (such as intelligence) and simple traits (such as skin color). I wrote a full response to Reich’s claims shortly after I read his chapter.40 My response was published as a chapter in the 2020 volume Critical Race Theory in the Academy. Although I do not subscribe to the philosophical discipline known as critical theory, I published in the book because it was designed by a respected colleague, Vernon Farmer (distinguished professor of education at Grambling State University). He passed away suddenly in 2019 before the volume was published. He had included my earlier essay “Science in the Belly of the Beast,” which was designed as a cautionary tale to young URM scientists, in his volume on African American pioneers in science.41
The article in Critical Race Theory outlined the major problems with Reich’s statement about my ideas. First and foremost among them is that his characterization of my ideas about the complexity of adaptation associated with a complex trait such as intelligence is based solely on a statement in a lecture given to an audience of mixed levels of scientific literacy. In a sense, he was saying that I didn’t understand the most elementary principles of natural selection on complex traits, even though my entire career in evolutionary biology has been organized around the experimental evolution of complex traits! If he had bothered to take the time to search for some of my publications, he would have realized that I fully understand how selection on complex traits works. The second problem with his statement was the utter and complete lack of evidence for natural selection differentiating human populations for genomic variants associated with cognitive function. In his book he suggested there were genetic variants, differentiated by population, that would account for differences in intelligence. One study supporting his claim examined seventy-four genetic markers (SNPs) in a European population contributing to individual differences in educational attainment.42 The authors of the study suggested that the phenotype “educational attainment” is at least 20 percent genetic, but that also means that nongenetic factors account for 80 percent of the phenotype. (Attempts to associate genomic variants moved from IQ tests to educational attainment because the SNPs associated with the former performed so dismally with regard to repeatability and ability to explain variation in the trait.) So even if we accept a 20 percent genetic contribution to educational attainment, if the effects are additive (equal), we would expect that each SNP is only contributing 0.27 percent to the variance in the trait (just as abysmally poor a result as shown for IQ tests).
Clearly, an individual’s cognitive function is influenced by his or her genetic constitution.43 It is also likely that cognitive function may have some correlation with educational attainment. However, accepting that fact is not the same as believing there should be significant genetic differentiation between racialized populations for cognitive function. This point is illustrated by a study conducted in my laboratory. We examined the effect of different superoxide dismutase (SOD) genotypes on longevity in the fruit fly.44 Longevity is a complex trait similar in genetic structure to cognitive function. SOD is an enzyme associated with preventing free radical damage to cells. The accumulation of free radical damage over the life span is one of the mechanistic causes of aging. This study was conducted prior to modern NGS methods, so the genetic variation was studied via electrophoresis (passing a current through an agar gel in such a way that it moved different protein forms at different rates). In Drosophila the SOD alleles were characterized by how far they moved in the gel: the fast (F) variant moves further than the slow (S) variant. There was a strong difference in the frequency of F and S SOD alleles between the selection treatments (in the short-lived flies, S variant = 0.00; in the long-lived flies, S variant = 0.25). In the second portion of the experiment, SOD genotypes were extracted from wild Drosophila populations and crossed into hybrid genetic backgrounds. Once that was done, there was no statistically significant difference in SOD activity by genotype (FF, FS, SS). However, we did find strong familial effects within each selection treatment, which means that while longevity is definitely genetically determined, the main impact of the SOD genotypes in this case was due to family (genetic background effect). In this case, family background was more important than the specific genotype.
The study that examined the seventy-four genetic markers listed some of the top-ranking candidate genes contributing to educational attainment. Of them, only one could be found in the Allele Frequency Database (ALFRED, maintained by the Kidd Lab at Yale School of Medicine). The population subdivision statistic (FST) for this SNP based on seventy-five populations was 0.128. FST has a range of 0 (no population subdivision) to 1.00 (total population subdivision). Any value below 0.250 is an indication that there is no racial or subspecies-level subdivision. This demonstrates that there is geographically based variation in the frequency of this gene but that in this case the variation between groups is not strong. Given how little this SNP contributes to cognitive variation and its weak differentiation between human groups, it is not strong evidence for a racialized difference in educational attainment (and certainly not of cognitive function).
A paper by Gregory Cochran, Jason Hardy, and Henry Harpending suggests that direct natural selection for greater intelligence explains the reputedly higher cognitive function of the Ashkenazim.45 The argument begins with the observation that Ashkenazi Jews have the highest mean IQ of all tested ethnic groups (about 0.75–1.0 standard deviations above the rest of Europe).46 It was hypothesized that this superior cognitive performance resulted from a heterozygote advantage (similar to that for sickle cell anemia) in which variants associated with lipid-storage diseases account for superior cognitive function in heterozygotes. Finally, for natural selection to favor the heterozygotes, the intelligence trait has to be associated with greater evolutionary fitness (differential survivorship and reproduction). The genetic variants reputedly associated with greater intelligence are those causing sphingolipid-storage diseases and glycogen-storage disorders.
There are four sphingolipid-storage disorders at high frequency in the Ashkenazim: Tay-Sachs disease, Gaucher disease, Niemann-Pick disease, and mucolipidosis type IV (MLIV). In Tay-Sachs, Niemann-Pick, and MLIV, the homozygous condition is usually lethal. Gaucher disease includes more than one type of mutation. One of them (homozygous 84GG) is lethal; another (N307S) is generally not. There is some histological evidence that in Gaucher disease, because it increases the amounts of glucosylceramide (a storage compound), sufferers have increased axon length and branching.47 Finally, there is evidence that in modern-day Israel, Gaucher individuals are disproportionately located in intellectually taxing professions (engineers, scientists, accountants, physicians, lawyers).48
Taken together, these observations could be seen as a strong example of evolution acting through heterozygote superiority in an isolated social group. However, it is not clear that the elevated frequencies of Gaucher disease variants in the Ashkenazim resulted from natural selection, as genetic drift is also a possible explanation.49 The Cochran, Hardy, and Harpending paper suggested that genetic drift was not the case. The most difficult piece of this argument is the question of whether there is real evidence to suggest that individuals with Gaucher (or other storage-disease heterozygotes) really did, during the feudal period in Europe, demonstrate greater reproductive success than those not carrying the variant. There is no evidence supporting that claim. Nor is there evidence that modern-day Gaucher individuals display greater reproductive success than others; in fact every indicator of reproductive success suggests the opposite. However, modern capitalist societies do not operate in the same way as feudal societies, so that is less of an objection to the theory. Finally, even if we accept that this single mutation of high penetrance really does dramatically increase an individual’s intelligence, it is clearly an exception. The 84GG mutation is nearly unique to the Ashkenazim, occurring only at extremely low frequency in other Europeans and in the rest of the world’s populations. The vast majority of genetic variants that affect cognitive function do not have such dramatic positive effects and certainly could not serve as the basis of an argument for racialized genetic differences in intelligence (unless one is willing to classify the Ashkenazim as a race).
WINTER IS COMING.
—GEORGE R. R. MARTIN, MANTRA OF THE STARK FAMILY, A GAME OF THRONES, 1996
Given the weak differentiation between human populations, those who continue to champion the notion that genomic foundations to racial differences in intelligence must exist are left with a huge elephant in the living room. Why and how should such differences exist? The key adaptation of our species is its greater intelligence compared to other animals. Our evolutionary lineage was characterized by increased brain size and complexity along with facial reduction.50 Many authors also suggest that the primary driving force of all primate intelligence (including our own) is social interactivity.51 Primates are highly social. Over the period from 2.5 million to five hundred thousand years ago the brain size of our hominid ancestors doubled. Over this same period the technical sophistication of these hominids was somewhat stagnant. Yet something was driving the brain growth (and possibly the intelligence growth). The seeming disparity between brain growth and lack of technical innovation has led some to propose that sexual selection rather than natural selection might have been at play. Sexual selection for a trait differs from natural selection in that it need not lead to greater survivorship of the individual so long as individuals with the trait in question come to have greater numbers of offspring. Evolutionary psychologist Geoffrey Miller went so far as to propose that the main driving force of this early physical change in brain size was the competition for mates.52 In this model, the benefit of greater cognitive capacity for technical innovation was a by-product of runaway sexual selection, not something directly selected for.
Whether Miller’s model is true or not, it is important to recognize that the majority of behavioral traits and intelligence of our species evolved long before anyone left Africa. We spent about two-thirds of our three-hundred-thousand-year existence there. This time depth also explains why the majority of the genetic variation found in our species is found in Africans. There is also a direct linear relationship between a population’s migratory distance from Africa and its genetic variation.53 Given the amount of genetic variation in Africa, it is notable that psychometricians never argue (as the melanist Afrocentrics do; see Chapter 7) that Africans are more genetically endowed for cognitive function than the rest of humanity. If genes were all there was to cognitive performance, we might expect that the most intelligent and the least intelligent populations would be found there. Maybe the Wakandans are still successfully hiding away from the world?
So we are still left with the question of whether and how there could have been direct selection for greater intelligence as humans migrated into Eurasia. One convenient answer to this mystery is winter. In 1864 Alfred Russel Wallace (codiscoverer of evolution by means of natural selection) proposed that winter selection would have favored greater intelligence. In 1925 Ellsworth Huntington argued that adaptation to the temperate zones caused strong selection for intelligence in Eurasians that was not experienced by sub-Saharan Africans.54 It is notable that Carleton Coon, known for both his racialism and his racism, did not reiterate Huntington’s idea in his work on the origin of the human races.55 This is especially significant in that he spent some time in that work discussing the importance of adaptation to climate as a factor in human evolution. Seventy years later, the winter selection idea was resurrected by J. Philippe Rushton and Arthur Jensen in the context of r- and K-selection theory applied to human races (see Chapter 7 on this fundamentally flawed idea).56
Despite its obvious problems, the idea that climate drove evolution still seems to have some traction. A recent paper argued for the importance of cold winters in spurring the evolution of intelligence in Eurasians.57 The paper suggested that during the last glacial maximum (about ten thousand years ago), eastern European populations displayed remarkable technical innovations that allowed them to survive the harsh winter conditions of their latitudes. These innovations included both new technology and changes in social organization. The technical innovations were focused on keeping the cold out and warmth in (to create clothing, shelter, and fuel), on managing time (to conserve limited winter food resources), and on changing the sexual division of labor (to open new roles to women in garment making, shelter building, fire making, pottery design and manufacture, and ornamentation). The relationship between evolved greater intelligence and these new technical innovations is supported by the greater cranial size (and hence brain size) associated with populations that migrated to these northern latitudes.
The paper relied on a study of the relationship between head size and latitude showing a clear trend toward larger heads in populations further away from the equator.58 The study examined 122 human groups and showed a strong correlation between brain size and the variables of solar radiation intensity, vapor pressure, and latitude in both the Eastern and the Western hemispheres. The authors of the study concluded that populations under severe cold stress had larger cranial volumes more from a change in head shape (rounder) than from differentiation due to total body size. The authors also made it very clear that there was no established relationship between IQ and cranial case volume in human populations.59 Finally, they also pointed out that head size differentiation is always a function of total body size. They mentioned that particularly large brain-size-to-body-mass ratios are seen in San hunter-gatherers (Africa), the Andamanese indigenous peoples (islands in the Indian Ocean, India), and the Bengalis (India), and that particularly low brain-size-to-body-mass ratios are seen in the French (Europe), the Mapuche (indigenous Chile), the Choctaw (indigenous North America), and the Maori (Pacific islanders). The groups in the high and low categories don’t correspond to the groups psychometricians typically claim for the genetically high IQ (East Asians) and low IQ (sub-Saharan Africans) categories.
Finally, there is no reason to believe that modern humans did not already possess all the cognitive ability needed to technologically innovate to meet new conditions before they arrived in Eurasia. The reasoning used by the winter selection theorists is circular. They claim that new innovations were required because of the movement of human populations into temperature zones. They argue that the appearance of innovations is evidence of genetically based greater intelligence in temperate populations and their absence is evidence of a lack of that intelligence in African tropical populations. However, if the innovations required to survive in winter are unnecessary in tropical zones, why would anyone build them? The minds of tropical populations would have been engaged in other aspects of their survival and reproduction. Their challenges were different (the rainy season, the barren soils of the tropics, the greater abundance of toxic plants and animals), but they were no less sophisticated than those associated with living in temperate zones.
IF ALL HUMAN POPULATIONS WERE ENGAGED IN STRUGGLES TO survive and reproduce in the environments where they found themselves, then to defend an argument for genetically based differences in cognitive function, one has to come up with some reason why natural selection would have operated differently on specific groups with regard to the underlying architecture of cognitive function. It is precisely here that psychometricians have failed. Indeed, the origin of the field itself is connected to the fear of dysgenesis. Francis Galton wanted England to enact eugenical policies because he believed that people of lower IQ were reproducing at a higher rate than those of high IQ—hardly an argument for the importance of the trait from an evolutionary perspective.
Evolution can produce a change in a complex trait as a result of selection for another trait. This phenomenon is called a correlated response to selection and is well-known from quantitative genetics.60 Correlation can occur if the genes determining the traits in question reside close to each other on the same chromosome (linkage) or if the traits result from the action of the same genes (pleiotropy). My understanding of these mechanisms and their impacts on complex traits was behind my lecture at the Peabody. Given that at all times, human populations are undergoing various forms of selection, the chance of sustained selection of any single kind resulting in an “accidental” evolution of superior intelligence in one group compared to others is so low it simply isn’t worth calculating. In my paper responding to Reich’s accusation that I was a defender of orthodoxy, I gave a detailed explanation of this problem for the accidental evolution of racialized intelligence.
There is no doubt that individuals differ in their cognitive abilities. There is also no doubt that some of those differences result from genetic sources and some from environmental sources. Determining the balance between the two has been at the center of some of history’s most contentious debates, beginning in the Western world with Greek philosophers (Aristotle). Support for the ideas that genes played the most significant role in determining cognitive abilities and that such abilities were racially differentiated reached gale force in the nineteenth and twentieth centuries with the ideas of people like Arthur de Gobineau, Francis Galton, Madison Grant, Ellsworth Huntington, Charles Davenport, Audrey Shuey, William Shockley, Arthur Jensen, Richard Herrnstein, Charles Murray, J. Philippe Rushton, Richard Lynn, Stephen Hsu, and many others. The vast majority of these individuals were males of European descent. Originally the genetic determinists based their conclusions on the design of a white supernatural entity (Agassiz’s zones of creation: Agassiz felt that the separate Adams and Eves had been created to occupy specific zones of the earth—tropical, temperate, arctic—along with their associated fauna and flora). When special creation was debunked, they shifted their arguments to evolutionary foundations of differential adaptation favoring greater intelligence among Eurasians. They utilized their scientific expertise to seize the scientific high ground against their opponents. They proposed plausible evolutionary scenarios to explain Eurasian superiority based on winter adaptation and r- and K-selection life history theory.
The problem with all of this, of course, is that their science was and is wrong. There is no a priori reason to believe that winter is any harder to deal with than tropical conditions. Simplistic statements like “planning was necessary to address the lack of food in winter” ignore the fact that the tropics have seasonal variation as well, such as the rainy season, whose problems also need to be solved. To support their claims, proponents of racially based differences in cognitive abilities adopted the tools of modern genetics as soon as they were available. In the late twentieth and early twenty-first centuries, they rapidly deployed NGS tools to search for the enigmatic foundations of greater human intelligence, and they failed utterly. When that failure became apparent, they shifted their search for the genes supporting educational attainment. Those studies, with larger cohorts, performed slightly better, but even the proponents of the approach had to admit that social, cultural, and environment influences play a substantial role in determining who gets educational opportunities and who does not.
I am not the first scholar (anthropologist or biologist) to take a stand against this pseudoscience. Franz Boas and his students (e.g., Ruth Benedict) took on this struggle in the early twentieth century; Ashley Montagu in the 1940s, Montague Cobb (an African American anthropologist) and his students in the 1950s, and Stephen Jay Gould and Richard Lewontin (and a host of others) in the 1970s all battled against this nonsense. I am the first African American evolutionary biologist to do so, and my training as an evolutionary biologist provided me with important skills that allow me to powerfully critique the logical fallacies so deeply interwoven in this program of disinformation.
So the question remains, why do biological determinist ideas of intelligence continue to resurface like the critters in a Whac-a-Mole game? The answer is simple: It is the most convenient explanation for social injustice. It makes an individual’s position in society the result of his or her own shortcomings rather than of the economic system that placed them there. In addition, racializing a genetic predisposition for intelligence creates in persons of Eurasian descent a pride in their genetic heritage. Thus, though they may not themselves be rocket scientists, engineers, doctors, lawyers, or corporate CEOs, the majority of people they see in those positions are members of their “race.” If they see a Black or Brown person there, they can fall back on the idea that some government affirmative action program allowed him or her to get there (after all, their genetic potential is inferior to that of the average white man). This in part explains why some of the people most damaged by the policies and actions of former president Donald Trump were willing to storm the US Capitol in a misguided attempt to maintain him in the White House.
Finally, there is a not fully appreciated danger looming in this ongoing belief in the simple genetic foundation of intelligence. Once the idea was simply to breed better (smarter) people—so-called positive eugenics. Positive eugenics failed, in large part because well-educated wealthy women of European descent did not want to have larger families. So if eugenical breeding can’t be implemented, why not develop biotechnological tools to improve human cognitive ability? For example, if NGS methods (such as GWAS) could be used to find those genetic variants, molecular biology tools could conceivably be developed to improve individual cognitive function. Not too long ago, there was a claim that the Beijing Genomics Institute (BGI) had initiated just such a project.61 The idea was that by studying the genomic foundation of greater mathematics ability in children shown to be mathematically precocious, a method could be developed to scan embryos, allowing a couple to decide which to implant to get the greatest potential for higher intelligence.62 Leaving aside the fact that this is, in all likelihood, a fool’s errand, the fact that a story about it was reported in the Wall Street Journal indicates that some people of wealth and power believe such techniques could work and are potentially willing to invest in them. The nightmare scenario is that the methods do work, which would result in people with the wealth and technological means of screening human embryos utilizing molecular methods to produce cognitively elite people. Such technology would undoubtedly result in the eugenic war scenario envisioned on Star Trek in “Space Seed” (season 1, episode 22). That is where such thinking goes, and it is a destination I emphatically suggest that the world avoid, lest extinction of our species follow.
Footnotes
i. Darwin biographers Adrian Desmond and James Moore tend to paint Darwin in the context of what a progressive scholar of his time would look like. Reading Darwin’s correspondence suggests that he was not entirely free of the European racism of his age.
ii. Agassiz’s abolitionism was of the Jeffersonian/Lincolnian variety—that is, abolition with the aim of removing persons of African descent from the United States.