CHAPTER 6
NATURAL SELECTION
It was not until the 1930s that evolutionists fully appreciated (as was shown in Chapters 2–4) that none of the explanations of evolution based on essentialism was valid. Curiously, the correct explanation had been found by Darwin 100 years earlier in 1838, although it was not published until 1858/1859. This was the concept of natural selection. The dramatic novelty of the Darwin–Wallace theory was that it was based on population thinking instead of essentialism. Alas, essentialism was the dominant way of thinking of the period and it took several generations before natural selection was universally adopted. However, population thinking had a compelling logic as soon as one adopted it.
Natural selection as proposed by Darwin and Wallace was a most novel and daring theory. It was based on five observations (facts) and three inferences (see Box 6.1). When one refers to populations in discussing natural selection, one ordinarily has sexually reproducing species in mind, yet it also takes place among the clones of asexual organisms.
The theory of natural selection proposed by Darwin and Wallace became the cornerstone of the modern interpretation of evolution. It was a truly revolutionary concept, having never before been suggested by any philosopher, and only rather casually referred to by two of Darwin’s contemporaries (William Charles Wells and P. Matthews). Even today many people have difficulty understanding how this principle works. Yet when population thinking is employed, it would seem to be simplicity itself. However, because the concept met strong resistance from long-established traditions and ideologies as the exclusive direction-giving factor to evolution, it remained a minority view from 1859 to the 1930s.
Box 6.1 Darwin’s Explanatory Model of Natural Selection
Fact 1. Every population has such high fertility that its size would increase exponentially if not constrained. (Source: Paley and Malthus)
Fact 2. The size of populations, except for temporary annual fluctuations, remains stable over time (observed steady-state stability). (Source: universal observation)
Fact 3. The resources available to every species are limited. (Source: observation, reinforced by Malthus)
Inference 1. There is intense competition (struggle for existence) among the members of a species. (Source: Malthus)
Fact 4. No two individuals of a population are exactly the same (population thinking). (Source: animal breeders and taxonomists)
Inference 2. Individuals of a population differ from each other in the probability of survival (i.e., natural selection). (Source: Darwin)
Fact 5. Many of the differences among the individuals of a population are, at least in part, heritable. (Source: animal breeders)
Inference 3. Natural selection, continued over many generations, results in evolution. (Source: Darwin)
To better appreciate the difficulty of understanding natural selection, one must take a closer look at this process. We must ask Darwinian questions. For instance, what happens in a given population through time? How does a population change from generation to generation? What is responsible for these changes and how do they affect the populations of a species?
POPULATION
Wherever a species occurs it is represented by a local population. Owing to unequal survival and reproductive success of its individuals, there is a continuing genetic turnover in each population as a result of chance and natural selection. Neighboring populations grade into each other if the habitat is continuous. However, favorable habitats are often discontinuous, resulting in a “patchy” distribution of the populations. Even greater breaks in the continuity of the populations occur where geographical barriers (mountains, water, unsuitable vegetation) inhibit dispersal. Along the border of a species’ range, populations are often rather isolated.
An understanding of the nature of populations is of the utmost importance for an understanding of evolution, because all evolution, and particularly selection, takes place in biopopulations. All aspects of populations are, therefore, of interest to the evolutionist. A local population is sometimes called a deme. It may be defined as the community of potentially interbreeding individuals at a given locality.
As we have seen, the concept of natural selection is based on observations of the natural world. Every species produces vastly more offspring than can survive from generation to generation. All the individuals of a population differ genetically from each other. They are exposed to the adversity of the environment, and almost all of them perish or fail to reproduce. Only a few of them, on the average two per set of parents, survive and reproduce. However, these survivors are not a random sample of the population; their survival was aided by the possession of certain attributes that favor survival.
NATURAL SELECTION IS REALLY A PROCESS OF ELIMINATION
The conclusion that these favored individuals had been selected to survive requires an answer to the question, Who does the selecting? In the case of artificial selection, it is indeed the animal or plant breeder who selects certain superior individuals to serve as the breeding stock of the next generation. But, strictly speaking, there is no such agent involved in natural selection. What Darwin called natural selection is actually a process of elimination. The progenitors of the next generation are those individuals among their parents’ offspring who survived owing to luck or the possession of characteristics that made them particularly well adapted for the prevailing environmental conditions. All their siblings were eliminated by the process of natural selection.
Herbert Spencer, when saying that natural selection is nothing but “the survival of the fittest,” was indeed quite right. Natural selection is a process of elimination, and Darwin adopted Spencer’s metaphor in his later work. However, his opponents claimed that it was a tautology, a circular statement, by defining “the fittest” as those who survive, but this is a misleading claim. Actually, survival is not a property of an organism but only an indication of the existence of certain survival-favoring attributes. To be fit means to possess certain properties that increase the probability of survival. This interpretation is equally applicable to the “nonrandom survival” definition of natural selection. Not all individuals have an equal probability for survival because the individuals that have properties making survival more probable are a restricted nonrandom component of the population.
Do selection and elimination differ in their evolutionary consequences? This question never seems to have been raised in the evolutionary literature. A process of selection would have a concrete objective, the determination of the “best” or “fittest” phenotype. Only relatively few individuals in a given generation would qualify and survive the selection procedure. That small sample would be able to preserve only a small amount of the whole variance of the parental population. Such survival selection would be highly restrained.
By contrast, a mere elimination of the less fit might permit the survival of a rather larger number of individuals because they have no obvious deficiencies in fitness. Such an enlarged sample would provide, for instance, the needed material for the exercise of sexual selection. This also explains why survival is so uneven from season to season. The percentage of the less fit in a population would depend on the severity of each year’s environmental conditions.
The larger the sample of the population that successfully passes the nonrandom process of elimination of the unfit, the more the success of the survivors will depend on chance factors and on selection for reproductive success.
The metaphor of selection pressure is frequently used by evolutionists to indicate the severity of selection. Even though it is a picturesque expression, this term, borrowed from the physical sciences, could be misunderstood, for there is no force or pressure connected with natural selection that corresponds to the use of the term in the physical sciences.
SELECTION IS A TWO-STEP PROCESS
Almost all of those who opposed natural selection failed to realize that it is a two-step process. Not realizing this, some opponents have called selection a process of chance and accident, while others have called it deterministic. The truth is that natural selection is both. This becomes obvious as soon as one considers the two steps of the selection process separately.
At the first step, consisting of all the processes leading to the production of a new zygote (including meiosis, gamete formation, and fertilization), new variation is produced. Chance rules supreme at this step, except that the nature of the changes at a given gene locus is strongly constrained (see Box 6.2).
At the second step, that of selection (elimination), the “goodness” of the new individual is constantly tested, from the larval (or embryonic) stage until adulthood and its period of reproduction. Those individuals who are most efficient in coping with the challenges of the environment and in competing with other members of their population and with those of other species will have the best chance to survive until the age of reproduction and to reproduce successfully. Numerous experiments and observations have revealed that certain individuals with particular attributes are clearly superior to others during this process of elimination. They are the ones that are “fittest to survive.” On the average, only two individuals of the abundant offspring of a set of parents will survive and become the progenitors of the next generation. This second step is a mixture of chance and determination. Clearly, those individuals with characteristics providing the greatest adaptedness to the current circumstances have the greatest probability of survival. However, there are also many chance elimination factors, so that there is no pure determination even at this step. Everything is somewhat probabilistic. Natural catastrophes, like floods, hurricanes, volcanic eruptions, lightning, and blizzards, may kill otherwise highly fit individuals. Furthermore, in small populations superior genes may be lost owing to sampling errors.
Box 6.2 The Two Steps of Natural Selection
Step One: The Production of Variation
Mutation of the zygote from its origin (fertilization) to death; meiosis, with recombination through crossing-over at the first division, and random movement of homologous chromosomes during the second (reduction) division; any random aspects of mate choice and fertilization.
Step Two: Nonrandom Aspects of Survival and Reproduction
Superior success of certain phenotypes throughout their life cycle (survival selection); nonrandom mate choice, and all other factors that enhance the reproductive success of certain phenotypes (sexual selection). At the second step much random elimination occurs simultaneously.
The fundamental difference between the first and the second steps of natural selection should now be clear. At the first step, that of the production of genetic variation, everything is a matter of chance. However, chance plays a much smaller role at the second step, that of differential survival and reproduction, where the “survival of the fittest” is to a large extent determined by genetically based characteristics. To claim that natural selection is entirely a chance process reveals total misunderstanding.
IS SELECTION A MATTER OF CHANCE?
Natural selection, unexpectedly, provided the solution to an old philosophical problem. An argument had been raging since the days of the Greek philosophers as to whether the events of this world are due to chance or to necessity. As far as evolution is concerned, Darwin put an end to this controversy. In short, owing to the two-step nature of natural selection, evolution is the result of both chance and necessity. There is indeed a great deal of randomness (“chance”) in evolution, particularly in the production of genetic variation, but the second step of natural selection, whether selection or elimination, is an antichance process. The eye, for instance, is not a chance product, as so often claimed by anti-Darwinians, but the result of the favored survival of those individuals, generation after generation, who had the most efficient structures for vision. (For an expanded analysis, see Chapter 10.)
Another widespread erroneous view of natural selection must also be refuted: Selection is not teleological (goal-directed). Indeed, how could an elimination process be teleological? Selection does not have a long-term goal. It is a process repeated anew in every generation. The frequency of extinction of evolutionary lineages, as well as their frequent changes in direction, is inconsistent with the mistaken claim that selection is a teleological process. Also there is no known genetic mechanism that could produce goal-directed evolutionary processes. Orthogenesis and other proposed teleological processes have been thoroughly refuted (see Chapter 4).
To say it in other words, evolution is not deterministic. The evolutionary process consists of a large number of interactions. Different genotypes within a single population may respond differently to the same change of the environment. These changes, in turn, are unpredictable, particularly when caused by the arrival at a locality of a new predator or competitor. Survival during a mass extinction may be strongly affected by chance.
CAN NATURAL SELECTION BE PROVEN?
After one has fully understood natural selection as a population process, it seems so obvious that one is at once convinced of its correctness. This is indeed what happened to Charles Darwin. However, in 1859 when he published the Origin, he actually did not have a single clear-cut piece of evidence for the existence of selection. The situation has completely changed since then. In the nearly century and a half since 1859, great amounts of concrete evidence have been acquired (Endler 1986).
The response of the genotype to a selection pressure is sometimes extraordinarily precise, as in some cases of mimicry, but far less so in other situations. As Cain and Sheppard have shown, bandedness in the snail Cepaea nemoralis is in certain habitats advantageous over an unbanded shell, but it would be difficult to prove that five bands are selectively superior to three bands.
The first proof of selection was the discovery of mimicry. The tropical explorer Henry Walter Bates (1862) observed in Amazonia that some palatable species of butterflies had the same pattern and coloration as sympatric toxic or at least unpalatable species, and that wherever the noxious models varied geographically, the palatable mimics followed the same geographic variation (
Fig. 6.1). This became known as
Batesian mimicry. A few years later, Fritz Müller (1864) discovered that poisonous species also mimicked each other so that the insect-eating birds had to remember only one model to avoid, thus protecting three or four or even a dozen different toxic species. This greatly reduced predation loss in the toxic species that mimicked each other, since the young birds had to learn only a single pattern for a whole group of Müllerian mimics
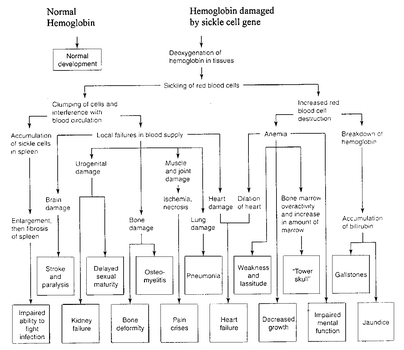
(Müllerian mimicry).Drug resistance of pathogens, as well as pesticide resistance in agricultural pests, eventually forced everybody to accept the importance of selection. In recent years, numerous occurrences of selection have been discovered by medical and public health workers. The relationship between the sickle cell gene and malaria resistance in Africa is a good example (
Fig. 6.2 and Box 6.3). Industrial melanism, in which moths and other organisms adapt to polluted habitats by changing body coloration, is a phenomenon in which the occurrence of selection has been particularly well tested experimentally.
Box 6.3 Sickle Cell Gene and Human Hemoglobin
The human sickle cell gene demonstrates the drastic effects that a mutation may have, even if it results only in the replacement of a single amino acid. The sickle cell gene is common in most malarial regions, particularly in Africa, because it protects the heterozygous carrier against malaria. In the sickle cell mutation the amino acid glutamic acid in the (beta) globin chain is replaced by valine. The blood disease caused by this mutation is sooner or later fatal for the homozygous carrier, but the heterozygotes are protected against malarial infections. This advantage is lost when a carrier of the sickle cell gene moves into a malaria-free region, like the United States. The frequency of the sickle cell gene among the descendants of slaves is gradually being reduced owing to the mortality of the homozygous carriers without any recompensating advantage of being heterozygous.
Geographic races of the Batesian mimic Papilio memnon (left) vary in parallel with the variation of their model (right).
Source: Reprinted from the Biology of Butterflies, R. I. Vane and E. B. Ford, page 266, copyright © 1984, by permission of Academic Press, London.
Pleiotropic effects of the sickle cell mutation.
Source: Strickberger, Monroe M. (1985). Genetics 3rd ed. Prentice-Hall: Upper Saddle River, N.J.
STRUGGLE FOR EXISTENCE
Darwin chose the metaphor “struggle for existence” as the title of the third chapter of the Origin. Every individual, whether animal, plant, or other kind of organism, “fights” every minute of its life for survival. If it is potential prey, it struggles with predators; if it is a predator, it fights for prey against other predators. In order to survive an individual has to meet successfully all conditions of life. As Darwin said: “A plant on the edge of a desert is said to struggle for life against the drought, though more properly it should be said to be dependent on the moisture” (1859: 62). The plant that is superior in drought resistance to the other members of the same population will survive best. The competition is usually most severe among members of the same population; this competition is not only for food, but also shelter and all the needs for successful reproduction, such as territory and mates. And, as Darwin continues, “as more individuals are produced than can possibly survive, there must in every case be a struggle for existence” (1859: 63).
But such struggle takes place not only among members of the same species, but often among individuals of different species. For example, seed-collecting ants in the American West compete for plant seeds with rodents. Red squirrels compete with red crossbills for pine seeds. On pastures and salt marshes I have seen passing flocks of starlings compete with beautiful yellow-chested local meadowlarks. In the tidal zone there is a pitched struggle for space among barnacles, mussels, kelp, and other marine organisms. In many instances, two species with similar requirements may manage to coexist. Yet when one of the species is experimentally removed, the other may strikingly increase in number. Many other pairs of species cannot coexist because their requirements are too similar and one is a little superior. This is referred to as the competitive exclusion principle. Sometimes it is quite puzzling how two seemingly quite similar species can successfully coexist. In the Galapagos Islands, species of Darwin’s finches coexisting on the same island have bills with different mean sizes and ranges of variation. If one of these species inhabits an island all alone, free of competition with the other species, its bill may have a much greater range of variation, including part of the range of variation of the species that competes with it elsewhere.
The importance of competition is demonstrated most graphically when a species becomes extinct as the result of an alien species successfully colonizing its range. Darwin called attention to the extinction of many native New Zealand species of animals and plants when introduced European species successfully established themselves there and outcompeted the natives.
Competition and other aspects of the struggle for existence exert an enormous selection pressure. Understanding the interactions among species has often been of great value for agriculture. Various pests of citrus orchards (aphids and scale insects) have been successfully controlled by ladybird beetles or other predatory insects. When introduced Opuntia cactuses were spreading like wildfire on Queensland sheep and cattle pastures, an Argentine moth (Cactoblastis) in no time virtually eliminated the cactus and restored tens of thousands of square miles of pastures to productivity. What these cases, and scores more in the ecological literature, demonstrate is that normally coexisting species live in steady-state harmony with each other, which is continuously adjusted by natural selection.
THE OBJECTS OF SELECTION
Who or what is being selected? Curiously this simple-sounding question has been the source of a long and continuing controversy. For Darwin, of course, as for virtually all naturalists since then, it was the individual organism that survived and reproduced. The genetics of the whole individual, however, cannot be dealt with mathematically, and most mathematical population geneticists, therefore, adopted the gene as the real “unit of selection.” Other authors proposed still other putative targets of selection, such as groups of individuals or whole species.
Some students of animal behavior and some ecologists thought that selection acted to “improve” the species. Up to 1970 some geneticists still thought that not only genes but also populations were the units of selection. It was not until about 1980 that reasonable unanimity was reached that the individual is the principal target of selection.
Much confusion about this problem can be avoided by considering two separate aspects of the question: “selection of” and “selection for.” Let us illustrate this with the sickle cell gene. For the question “selection of” the answer is an individual who either does or does not carry the sickle cell gene. In a malarial region the answer to “selection for” is the sickle cell gene, owing to the protection it gives to its heterozygous carriers. When one makes the distinction between the two questions, it becomes quite clear that a gene as such can never be the object of selection. It is only part of a genotype, whereas the phenotype of the individual as a whole (based on the genotype) is the actual object of selection (Mayr 1997). This does not reduce the importance of the gene in evolution, for the high fitness of a given phenotype may be due to one particular gene.
The reductionist thesis that the gene is the object of selection is also invalid for another reason. It is based on the assumption that each gene acts independently of all other genes when making its contribution to the properties of the phenotype. If this were true, the total contribution of genes to the making of the phenotype would be accounted for by the addition of the action of all individual genes. This assumption is referred to as the “additive gene action” assumption. Indeed some genes, perhaps even many genes, seem to act in such a direct and independent manner. If you are a male with the hemophiliac gene, you will be a bleeder. Many other genes, however, interact with each other. Gene B may enhance or reduce the effects of gene A. Or else the effects of gene A will not occur unless gene B is also present. Such interactions among genes are called epistatic interactions.
Obviously, epistatic interactions are not as easily determined as additive gene actions and their study has therefore been generally avoided by geneticists. One such interaction is designated as “incomplete penetrance.” In such a case an individual may have a particular gene but does not show its effect, while it is fully reflected in the phenotype of another member of the population who has a somewhat different genotype. For instance, in a widely adopted model of the inheritance of schizophrenia, it is postulated that the major gene contributing to this illness has only a 25 percent penetrance, that is, is manifested in only 25 percent of the carriers of this gene. Some of the combinations of interacting genes are apparently tuned so finely that any deviation from the optimal balance is being selected against. Pleiotropy and polygeny are well-known instances of such interactions of genes (see Chapter 5).
The importance of this interaction of genes was not fully realized until the discovery of the regulatory genes, like hox and pax genes. With these genes we can observe very drastic interactions, but minor interactions among genes are very common. The question of what all of these interactions add up to is controversial. But there is a good deal of indirect evidence for the existence of an “internal balance” of the genotype or, as it has also been called, a “cohesion of the genotype.” It has been postulated that this is a conservative element in evolution and accounts for the stasis in so many evolutionary lineages. It has also been postulated that this is the reason why founder populations may change drastically so often and so rapidly. Founder populations have a greatly reduced variance and may have a rather unbalanced set of genes. Such gene pools may respond to new selection pressures rather differently than the parent species and may be able to produce greatly diverging phenotypes.
It is important for the clarification of various evolutionary controversies to clearly understand how variable a contribution to fitness a gene may make. Many genes do not have a standard selective value. A gene may be beneficial when placed in one particular genotype, but it may be deleterious when placed in a genotype with different genes. Hence, the interaction of genes is of considerable importance for the selective value (fitness) of an individual. So-called neutral evolution (see below) is a meaningless concept considering the fact that the gene as such is not the target of selection.
A gene may have a very different effect on the fitness of an individual whether it is present in a single dose (heterozygote) or a double dose (homozygote). A single dose of the sickle cell gene greatly adds to the fitness of its heterozygous carrier in a malaria region, while in double doses (as in homozygotes) it is sooner or later lethal. This shows particularly graphically that a gene does not necessarily have a fixed selective value, but that this value may depend on the other genes with which it is associated in the genotype.
PHENOTYPE
What do we mean when we say that the individual is the object of selection? What is it that is encountered by natural selection and induces it either to favor or to disfavor an individual? Not its genes or genotype, because these are not visible to selection, but rather its phenotype. The word phenotype refers to the totality of morphological, physiological, biochemical, and behavioral characteristics of an individual by which it may differ from other individuals. The phenotype originates during development of the zygote from the fertilized egg to adulthood owing to the interaction of the genotype with its environment. The same genotype may produce quite different phenotypes in different environments. A semiaquatic plant, for instance, may produce entirely different leaves on land than in the water (
Fig. 6.3)
Phenotypic variation in leaf form in the semiaquatic plant Ranunculus aqu atilis. Compare the filamentlike leaves on submersed branches (a) with normally structured leaves on branches above the water (b).
Source: Herbert Mason, Flora of the Marshes of California. Copyright © 1957 Regents of the University of California, copyright renewed 1985 Herbert Mason.
The phenotype consists not only of the structure of an organism and of its physiology, but also of all the products of the behavioral genes. This includes the nest a bird builds, or the web of a spider, or the path of migration of a migratory bird. Dawkins (1982) has referred to these aspects of the characteristics of an organism as the extended phenotype. It is as much (and often more so) the target of selection as the structural characteristics of an organism.
The range of phenotypic variation that a particular genotype is able to produce is referred to as its norm of reaction. Thus the phenotype is the result of the interaction between genotype and environment. Some species have a very wide reaction norm; they can adjust their phenotype to wide variations of the environment and have a high phenotypic plasticity. The fact that it is the phenotype, rather than the genotype, that is the target of selection allows the existence of considerable genetic variation in a gene pool. Such variation is compatible with selection as long as the produced phenotypes have an acceptable selective value.
Because the phenotype is the product of the genotype, it has both evolutionary stability and evolvability. Many core processes at the cellular level are conserved throughout the metazoa, such as many signaling pathways and genetic regulatory circuits, others are conserved throughout the eukaryotes (e.g., the cytoskeleton), and yet others throughout all life-forms such as metabolism and replication. Sequence conservation is so strong that more than half of the coding sequences of yeast are recognizable in mice and humans. The actins of yeast and humans, for example, are 91 percent identical.
These core processes, however, must not be so structured that they prevent further evolution. Indeed there is ongoing selection for evolvability of the phenotype. Only this flexibility permits the occupation of new adaptive zones and the successful coping with new environmental challenges. The study of the properties of a genotype that allow it to cope with the constraints of the conserved portions and to maintain optimal evolvability is one of the frontiers of current evolutionary biology.
OTHER POTENTIAL OBJECTS OF SELECTION
The individual is not the only entity that has been suggested by one or another evolutionist to be the object of natural selection. We have already refuted this claim for the gene, and will now discuss gametes, groups, species, higher taxa, and clades.
Gamete Selection
All gametes are subject to selection between the completion of the meiotic cycle and either fertilization or death. Elimination is extremely severe since only a very small fraction of the gametes is successful. Unfortunately, we know very little about the eliminating factors. Experiments have revealed that the proteins of the egg wall in certain marine invertebrates have the capacity to prevent the entry of some spermatozoa while admitting others. What fitness criteria are involved here is still unknown. The properties responsible for gametic selection are important isolating mechanisms, referred to as gametic incompatibility.
Gamete interaction has been studied much more intensively in plants, particularly the compatibility reactions between pollen tubes and the stigma or style. In many taxa, special mechanisms prevent self-pollination. Less is known about incompatibilities among out-crossing species and how it is controlled. As early as the 1760s the botanist J. G. Kölreuter showed that when conspecific and alien pollen were placed on the stigma simultaneously, it was always the conspecific pollen that fertilized the seed. If the alien pollen alone was placed, it successfully fertilized in some species pairs.
Group Selection
There has been much argument whether a group of individuals can or cannot be the object of selection. The situation is clarified if one makes a distinction between “soft” and “hard” group selection (Mayr 1986). Soft group selection refers to selection of casual groups, and hard group selection to cohesive social groups. In the case of soft group selection, the fitness of the group is the arithmetic mean of the fitness values of the members of the group. This mean value has no effect whatsoever on the fitness of the composing individuals. The evolutionary success or failure of such a group (“group selection”) is simply the automatic consequence of the fitness of the composing individuals. The fact that they are associated in a group makes no contribution to their fitness. Such soft group selection makes no independent contribution to evolution. This is the kind of “group selection” one finds in casual groups. Soft group selection really should not be referred to as group selection, because the group as such is not selected. A population as a whole is subject to such soft “group selection.”
However, in certain species a special kind of group occurs, social groups, that can indeed be a target of selection. Such a group, owing to social cooperation among its members, has a greater fitness value than the arithmetic mean of the fitness values of its individual members. This may be called hard group selection. Members of such groups cooperate by warning of enemies, sharing newly discovered sources of food, and joint defense against enemies. This cooperative behavior enhances the survival propensity of such a group. The human species, at least during the hunter-gatherer stage, benefited from such social cooperation and this led to increased survival of certain groups. As a result, any genetic contribution toward cooperative behavior would be favored by natural selection. It is believed that this social cooperation has been an important factor in the development of human ethics (see Chapter 11). Hard group selection does not replace individual natural selection, but is superimposed on it.
Kin Selection
A form of selection called kin selection is recognized by many evolutionists, particularly in connection with the evolution of altruism. It is defined as selection for characteristics that favor the survival of close relatives of a given individual who shares part of the same genotype (known as inclusive fitness altruism). Except for parental care and in social insects, kin selection is probably not as important a factor in evolution as sometimes believed, particularly when there is a considerable exchange of individuals among neighboring groups. The altruism that members of a social group show to other related members of the group (excluding offspring) is apparently never anywhere near as great as the altruism displayed by parents (particularly mothers) to their own offspring. It is perhaps misleading to combine the two kinds of relationship under the single term kin selection. However, since members of a social group are often closely related to each other, much hard group selection is simultaneously kin selection. (See also Chapter 11.)
Species Selection
The history of evolution is a steady extinction of species and the origin of new species. This turnover is often apparently due to the superiority of a new species over an established species. Also, when species of different biota come into competition, as did those of North and South America after the isthmus of Panama had been established in the Pliocene, it can result in considerable extinction, part of it caused by competition between the invaders and the indigenous species. This phenomenon has been called species selection. As mentioned earlier, Darwin called attention to the frequent extinction of native species of plants and animals on New Zealand after European species had been introduced. The mistake was made by some authors to consider this an alternative to individual selection. In reality, this so-called species selection is superimposed on individual selection. The individuals of both species coexist after the entry in the same niche, and extinction takes place when the individuals of the invading species are, on average, superior to those of the indigenous species. Clearly, it is a selection of individuals. Misunderstandings are avoided if this process is called “species turnover” rather than “species selection” (see Chapter 10). A species as a unit is never the object of selection, only its individuals.
An even higher taxonomic level is involved in so-called clade selection, a clade being a holophyletic group of taxa forming a branch of a phylogenetic tree. Owing to the Alvarez extinction event at the end of the Cretaceous, the clade of the dinosaurs became extinct, but not the clades of birds and mammals. During every mass extinction certain higher taxa have fared better than others. Again, the actual objects of selection were individuals, but the individuals of some clades shared characteristics that favored survival through the extinction event, while these characteristics were absent in the individuals of the losing clades. What is remarkable for mass extinctions is that a whole higher taxon may be eliminated almost instantaneously or at least during a relatively short period. Clade extinctions also sometimes occur that are not clearly the result of a mass extinction. The extinction of the trilobites may be an example.
Competition Among Higher Taxa
Mass extinctions have called attention to the possible competition between higher taxa. Mammals had existed for some 100 million years prior to the mass extinction at the end of the Cretaceous, but they were small, insignificant, and quite likely nocturnal. Why did they enjoy such an explosive radiation in the ensuing period of the early Tertiary? The most widely accepted answer to this question is that they were able to enter all the ecological niches vacated by the demise of the previously dominant dinosaurs. Evidently the two classes of animals had been competing all along, but the dinosaurs had been superior competitors. Obviously, the mammals did not cause the extinction of the dinosaurs, but they replaced them when the dinosaurs became extinct owing to a nonbiological cause.
This case of the mammalian flourishing also illustrates the phenomenon of explosive speciation in previously vacant niches. Other examples are the species flocks of fish, molluscs, and crustaceans in ancient lakes and the rapid radiation of colonists of oceanic archipelagos. There are more than 700 species of drosophilid flies and over 200 species of crickets on the Hawaiian Islands. The honey creepers (Drepanididae) in the Hawaiian Islands and the geospizid finches in the Galapagos are other well-known cases of such radiation.
In all of these cases it was the absence or the removal of competition that made the radiation possible. One speaks of displacement when an incumbent taxon is exterminated by competitive exclusion owing to the arrival of a superior competitor. To actually prove the causal connection in such a sequence is difficult. For instance, the multituberculates were a flourishing group of nonplacental mammals in North America in the late Cretaceous and Paleocene. But when, in the Eocene, the first rodents appeared (probably from Asia) and became extremely successful, common, and widespread, the multituberculates became rare and finally died out. The extinction of the trilobites when the bivalves became so successful is another possible case, but an environmental catastrophe has also been proposed for their demise. Throughout the history of paleontology there are numerous similar cases of the sudden decline and ultimate extinction of a previously prosperous taxon after a new taxon with seemingly similar ecological requirements had appeared. It is, of course, impossible to prove in any of these cases that it was the arrival of the new competitor that caused the extinction, but this scenario often fits the known facts better than any other explanation.
WHY IS EVOLUTION USUALLY SO SLOW?
When the pharaohs’ tombs were opened in Egypt early in the nineteenth century, not only human mummies were found but also those of sacred animals such as cats and ibises. When the anatomy of these animal mummies, estimated to be about 4000 years old, was carefully compared by zoologists with living representatives of these species, no visible differences could be found. This finding was in striking contrast to the rapidity with which animal breeders had produced pronounced changes in domestic animals in a much shorter time. Consequently, the absence of any visible change in these mummies was used as an argument against Lamarck’s theory of evolution. Now we know that it usually takes many thousands if not millions of years for visible changes in evolving species to occur, except in a few special situations. Hence, the constancy of the Egyptian mummies is no evidence against evolution.
With drastic selection taking place in every generation, it is legitimate to ask why evolution is normally so slow. The major reason is that owing to the hundreds or thousands of generations that have undergone preceding selection, a natural population will be close to the optimal genotype. The selection to which such a population has been exposed is normalizing or stabilizing selection. This selection eliminates all of those individuals of a population who deviate from the optimal phenotype. Such culling drastically reduces the variance in every generation. And unless there has been a major change in the environment, the optimal phenotype is most likely that of the immediately preceding generations. All the mutations of which this genotype is capable and that could lead to an improvement of this standard phenotype have already been incorporated in previous generations. Other mutations are apt to lead to a deterioration and these will be eliminated by normalizing selection. There are also some special genetic mechanisms, such as genetic homeostasis (including heterozygote superiority), that favor maintenance of the steady state.
FOUNDER POPULATIONS
The genotype is a carefully balanced system owing to the epistatic interaction of the composing genes. Selection for the replacement of a gene by a new one may therefore require adjustments at other gene loci. The larger a population, the slower will be the incorporation and spread of new genes. By contrast, a small founder population established by the offspring of a single fertilized female or by a few founders may be able to shift more quickly to a new adaptive phenotype since it is unconstrained by the cohesive forces of a large gene pool.
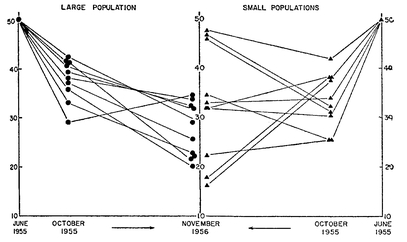
There is much observational evidence that evolutionary change to the level of completed speciation proceeds more rapidly in peripheral populations than in large widespread species (Mayr and Diamond 2001). The explanation for this is still controversial. Dobzhansky and Pavlovsky (1957) (
Fig. 6.4) showed a long time ago that a group of small, originally identical populations diverged from each other much more rapidly than a group of identical large populations. Other laboratory studies of founder populations did not find a drastic change in such populations. However, most of the studies were made with
Drosophila melanogaster, a species with a phenotype that is apparently particularly stable as indicated by its various sibling species. This constancy of
D. melanogaster raises the possibility that different founder populations may react differently to their isolation. Traditionally, the greater evolutionary inertia of large populations was ascribed to the greater amount of pleiotropy and polygeny. However, another cause is presumably the distribution of different regulatory genes. Conservative gene flow does not reach isolated populations and interfere with their increasing divergence. There is every reason to believe that new discoveries in developmental genetics will contribute a better understanding of the causes for different rates of evolutionary change in general and in speciation in particular.
WHAT IS THE EVOLUTIONARY ROLE OF BEHAVIOR?
For Lamarck, behavior was an important cause for evolutionary change. He thought that changes in organisms caused by any kind of activity would be transmitted to future generations by the inheritance of acquired characters. For example, when giraffes stretched their neck to reach higher leaves, the resulting elongation of the neck would be inherited by the next generation. Even though this theory of inheritance is now refuted, evolutionists still believe, but for very different reasons, that behavior is important in evolution. A change in behavior, for instance, adoption of a new food item or increased dispersal, is apt to set up new selection pressures, and these may then lead to evolutionary changes (Mayr 1974). There are reasons to believe that behavioral shifts have been involved in most evolutionary innovations, hence the saying “behavior is the pacemaker of evolution.” Any behavior that turns out to be of evolutionary significance is likely to be reinforced by the selection of genetic determinants for such behavior (known as the Baldwin effect).
Variation, epistatic effects, and population size. The frequency (percent, vertical scales) of PP chromosomes in twenty replicate experimental populations of mixed geographic origin (Texas to California). The populations that have gone through a bottleneck of small population size show far greater variance after 17 months than the continuously large populations.
Source: Mayr, E. Animal Species and Evolution. Belknap (HU Press), 1966.
SELECTION FOR REPRODUCTIVE SUCCESS (SEXUAL SELECTION)
When we speak of natural selection, unconsciously we always think of the struggle for existence. We think of the factors that favor survival, such as a capacity to overcome adverse weather conditions, to escape enemies, to better cope with parasites and pathogens, and to be successful in competition for food and habitation, in short to have any property that would enhance the chances for survival. This “survival selection” is what most people have in mind when they speak of natural selection.
Darwin, however, saw quite clearly that there was a second set of factors enhancing the probability of leaving offspring: all factors contributing to an increase of reproduction success. He called these factors sexual selection. Darwin listed here all cases of pronounced sexual dimorphism, such as the large antlers of deer, the magnificent tails of male peacocks, and the resplendent plumage of male birds of paradise and hummingbirds. Since females usually have the opportunity to choose their mates, those males will be favored by sexual selection who are most successful in attracting the favor of mate-seeking females. Other male characters that in some species are also favored by sexual selection are those that help males to be victorious in fights with rivals and that permit such males to acquire a larger harem of females, as occurs among seals, deer, sheep, and other species of mammals. Those males who benefit from such characteristics have increased reproductive success. However, enhanced reproductive success may also be effected by other means, such as the ability to obtain superior territories, sibling rivalry, aspects of parental investment, and other aspects of the interaction among individuals in a family or in a population. Darwin defined sexual selection as “the advantage which certain individuals have over others of the same sex and species solely in respect to reproduction.” The term “selection for reproductive success” actually fits this broad definition better than the term sexual selection.
The cases in which competition or fighting among members of the same sex, such as among seal bulls or stags, leads to reproductive success have been designated “intrasexual selection.” Other cases, such as female choice, in which the selection takes place between the two sexes, have been called “intersexual selection.” There has been much discussion in recent years concerning the criteria employed by females when making their choice. Zahavi (1997) postulated that females may choose particularly conspicuous males, because their survival in spite of the handicap of conspicuousness indicates that they have superior qualities (the so-called handicap principle).
Simultaneous Mate and Niche Selection
One would expect particularly strong normalizing selection to achieve a complete constancy of the behavioral components of a species’ isolating mechanisms to prevent loss of fitness through hybridization. This is indeed ordinarily true. However, it appears that in some cases mate selection is correlated with niche selection and that a heterogeneity of available niches may lead to a diversification of mate selection. Different kinds of males in a population may have different kinds of reproductive success in different subniches or habitats. In a species of freshwater fish, cichlids for example, some males may have a preference for benthic feeding and others for open-water feeding. Different kinds of females may originate in such a species, some with a preference for benthic and others for pelagic males. Eventually, through the process of sympatric speciation, two species may evolve. In such cases, sexual selection leads to speciation.
Sympatric speciation in plant-feeding insects may take place by simultaneous mate and host selections, as Guy Bush has maintained for many years. If insects that are on the whole host specific on plant A succeed in colonizing plant species B, and if the new colonists on B develop a mating preference for other individuals who have become adapted to species B, a host-specific new species will evolve on plant B, and reverse colonization from B back to A will be rare.
Sexual Dimorphism
Male and female appearances differ from each other in most animals, and there is an extraordinary range of variation in the degree of this sexual dimorphism. Certain deep-sea fish have dwarf males that are attached to the females, because free-swimming males might have difficulty finding females in these vast and rather lifeless spaces. At the other extreme, in certain species of seals, like the elephant seal, males may be several times larger than females because larger males can better defeat their rivals in territorial fights and so acquire larger harems. The magnificent plumages of male birds of paradise, hummingbirds, and other birds with polygamy were already mentioned under sexual selection. None of these cases poses any real difficulty for the theory of natural selection because all of these special male characteristics have the selective advantage of enhanced reproductive success. Invariably there is some counterselection against a runaway evolution of too extreme development of male characteristics. When they reduce survival, they will be selected against.
WHY DOES NATURAL SELECTION SO OFTEN FAIL TO ACHIEVE OR MAINTAIN ADAPTEDNESS?
Some enthusiasts have claimed that natural selection can do anything. This is not true. Even though “natural selection is daily and hourly scrutinizing, throughout the world, every variation even the slightest,” as Darwin (1859: 84) has stated, it is nevertheless evident that there are definite limits to the effectiveness of selection. Nothing demonstrates this more convincingly than the fact that 99.99 or more percent of all evolutionary lines have become extinct. We must ask ourselves, therefore, why is natural selection so often unable to produce perfection? Recent studies have revealed numerous reasons for such limitation. A discussion of these constraints can contribute a great deal to the understanding of evolution. I recognize seven classes of such constraints.
1. The limited potential of the genotype. The existing genetic organization of an animal or plant sets severe limits to its further evolution. As Weismann expressed it, no bird can ever evolve into a mammal, nor a beetle into a butterfly. Amphibians have been unable to develop a lineage that is successful in salt water. We marvel at the fact that mammals have been able to develop flight (bats) and aquatic adaptation (whales and seals), but there are many other ecological niches that mammals have been unable to occupy. There are, for instance, severe limits on size, and no amount of selection has allowed mammals to become smaller than the pygmy shrew and the bumblebee bat, or allow flying birds to grow beyond a limiting weight.
2. Absence of appropriate genetic variation. A given species population can tolerate only a limited amount of variation. In the case of any drastic change of the environment, whether a climatic deterioration or the appearance of a new predator or competitor, the kind of genes needed for an appropriate immediate response to this new selection pressure may not be present in the gene pool of the population. The high frequency of extinction documents the importance of this factor.
3. Stochastic processes. Much of the differential survival and reproduction in a population are not the result of selection, but rather of chance. Chance operates at every level in the process of reproduction, beginning with the crossing-over of parental chromosomes during meiosis to the survival of the newly formed zygotes. Furthermore, potentially favorable gene combinations are undoubtedly often eliminated by indiscriminate environmental forces such as floods, earthquakes, or volcanic eruptions before natural selection has had the opportunity to favor specific genotypes.
4. Constraints of phyletic history. Several alternate responses are usually possible to any environmental challenge, and it is the existing structure of the organism that often prescribes what response prevails. When the selective advantage of a skeleton developed among the ancestors of the vertebrates and of the arthropods, the arthropod ancestors had the prerequisites for developing an external skeleton, and the vertebrate ancestors for acquiring an internal skeleton. The entire evolution of these two large groups of organisms has since been affected by this choice among their remote ancestors. It permitted vertebrates to develop such huge creatures as dinosaurs, elephants, and whales, whereas a large crab is the largest type that the arthropods were able to achieve. The need for a regular molt of the external skeleton sets up in arthropods a formidable selection pressure against size increase.
Once a particular body structure has been acquired, it may not be possible to change it again. For instance, in terrestrial vertebrates the respiratory tract from the oral cavity to the trachea crosses the digestive tract, which also runs from the oral cavity to the esophagus. This arrangement was adopted in rhipidistian fishes, our aquatic ancestors. Although it poses forever the danger of food entering the trachea, no reconstruction of this inferior pathway occurred in several hundred million years.
Pelagic floating has been achieved by descendants of sessile, benthic, and actively swimming ancestors, belonging to many different animal phyla, who became adapted to the pelagic form of life through such very different adaptations as the inclusion of oil droplets, an increase of the surface area, and various other mechanisms. Each solution is a different compromise between the constraints or opportunities of this new adaptive zone and a species’ previously existing physical structure. The adoption of a particular response to a new environmental opportunity may greatly restrict the possibilities of future evolution.
5. A capacity for nongenetic modification. The more pliable the phenotype is, that is, the larger its norm of reaction (owing to developmental flexibility), the more this reduces the force of an adverse selection pressure. Plants, and particularly microorganisms, have a far greater capacity for phenotypic modification than do higher animals. However, a capacity for nongenetic modification is present even in humans. This is exemplified by the physiological changes in a person when he or she ascends from the lowlands to high altitudes, where over the course of days and weeks the individual can become reasonably well adapted to the lowered atmospheric pressure and consequent decrease in oxygen. Natural selection is, of course, involved even in this phenomenon, since the capacity for nongenetic modification is under strict genetic control. Also, when a population shifts to a new specialized environment, genes will be selected during the ensuing generations that reinforce and eventually largely replace the capacity for nongenetic adaptation (the Baldwin effect).
6. Unresponsiveness of the postreproductive age. Selection cannot eliminate genetic propensities for diseases of old age. In the human species, for instance, most genotypes responsible for Parkinson’s, Alzheimer’s, and other afflictions that manifest themselves primarily in postreproductive life are relatively immune to selection. To some extent this is even true for diseases of middle age like prostate cancer and breast cancer, which usually strike toward the end of the active reproductive age.
7. Developmental interaction. It was realized by students of morphology as far back as Étienne Geoffroy St. Hilaire that there is competition among an individual’s organs and structures. Geoffroy expressed this in his La Loi de Balancement (Law of Balancing, 1822). The different components of the morphotype are not independent of each other, and none of them responds to selection without interaction with the other components of the morphotype. The whole developmental machinery is a single interacting system. An organism’s structures and functions are compromises among competing demands. How far a particular structure or organ can respond to the forces of selection depends to a considerable extent on the resistance of other structures and other components of the genotype. Wilhelm Roux, more than 100 years ago, referred to the competitive developmental interactions as “the struggle of parts” in organisms.
The morphology of every organism reveals to what degree it is the result of a compromise. Every shift into a new adaptive zone leaves a residue of no longer needed morphological features that then become an impediment. One only needs to think of the many weaknesses in humans that are remnants of our quadrupedal and more vegetarian past, for instance, the facial sinuses, the structure of the lower vertebral column, and the caecal appendix. Such vestiges of former adaptedness are referred to as vestigial characters (see Chapter 2).
8. The structure of the genotype. The classic metaphor of the genotype was that of genes lined up like beads on a string. According to this view, each gene is more or less independent of the others and all of them are more or less similar in their nature. Not much is left of this view, which was generally accepted 50 years ago. To be sure, all genes are composed of DNA and the information they contain is coded in the linear sequence of base pairs. However, modern research in molecular genetics has revealed that there are different functional classes of genes, some charged to produce material, others to regulate it, and still others that are seemingly without function (see Chapter 5).
Furthermore, there is a good deal of rather indirect evidence that groups of genes may be organized into functional teams, which, in many respects, act as wholes (known as modular variation). However, this is a rather controversial area of molecular biology, and perhaps the best one can do at the present time is to call attention to the fact that the old “beads on a string” image of the genotype is no longer valid, and that there is still great uncertainty as to the actions of the genotype. The fact that there are transposons, introns, middle repetitive DNA, highly repetitive DNA, and many other kinds of noncoding DNA suggests different functions, but most of what these elements are and how they work together is still to be determined. More will be learned about the process of evolution as our understanding of the structure and functioning of the genotype improves than by anything else.
THE ROLE OF DEVELOPMENT IN EVOLUTION
The fertilized egg, the zygote, is a formless mass. It is converted into the phenotype of the adult stage during the embryonic or larval stages of development. Changes occurring during development are responsible for the divergence of different evolutionary lineages. Hence the study of development, of the ontogeny of the developing zygote, is of major concern to every evolutionist. However, the methods of classic embryology and in particular those of experimental embryology (Entwicklungsmechanik ) were not suitable to produce the needed synthesis between embryology and genetics; this is finally being achieved by molecular biology. What was needed was a study of gene action, that is, a determination of the contribution to the development of the embryo made by each gene. This led to the discovery of the great diversity of genes, and in particular to the discovery of the regulatory genes (see Chapter 5).
Development is rarely direct. In a high proportion of animals the adult stage is reached through one or several larval stages, some of them requiring highly specific adaptations. One needs only to think of the caterpillar and butterfly or of the planktonic larva of the barnacle and its mollusclike adult. In these cases, new adaptations are acquired by some ontogenetic stages, but in other cases, particularly among parasites, certain phenotypic adaptations of the adult stage are lost, as in the Sacculina parasite of certain crabs.
DEVELOPMENT
Evolutionists, stretching all the way back to Darwin, realized that the “type” does not evolve as a unit and at the same rate in all of its parts, but that some components of the phenotype evolve faster, and some more slowly. This can be observed when a phyletic lineage shifts from one adaptive zone to a different one. Archaeopteryx, the earliest well-known fossil bird, had already acquired various avian characters—feathers, wings, the capacity for flight, enlarged eyes, and a birdlike brain—but had retained a reptilian stage of other parts of its structure (teeth, tail vertebrae). Such an unequal rate was referred to in earlier chapters as mosaic evolution. It would seem in such cases as if the phenotype was produced by more or less independent sets of genes. Consequently it has been postulated that the genotype is composed of an assemblage of gene modules, each controlling one of the mosaics of the phenotype. This thought was quite unpalatable to highly reductionistic geneticists, but the evidence for a somewhat modular structure of the genotype is increasing. If this is correct, a single regulatory gene may control such a module of genes. In other words, a mutation of the regulatory gene may result in a rather drastic change (discontinuity) of the phenotype. In other cases such a module may consist simply of a set of genes temporally brought together by selection for a particular state of adaptedness, but that might again be disassembled when the selective conditions change. There is a lot of structure in a genotype that cannot be discovered and explained by a purely reductionistic approach.
A Balance of Selection Pressures
No individual is ever perfectly adapted, as was stressed early on by Darwin. The main reason for this is perhaps that every genotype represents a compromise of genetic variability and stability. Most environments are perpetually changing, and at the end of a drought period a population will be better adapted for drought conditions than for an oncoming wet period. In the long run, the genotype strikes a balance between conflicting demands. And the same is true for the behavior of an organism toward predators and competitors. Mathematically inclined evolutionists have expressed this in terms of the game theory and superior strategies. Actually, of course, animals hardly test the various strategies in their minds. Rather their genotype predisposes some individuals in a variable population to be more timid and others to be more bold. Those individuals with the most successful balance of the two tendencies in a given situation will have the best chance to survive. There is no selection for the favorite type, rather the mean value of the population will reflect the balance of success of the various, sometimes rather conflicting genetic tendencies.
The response to a change in environmental conditions is often not predictable. When the climate of North America became more arid in the Pliocene, the vegetation responded and grasses, indeed eventually rather harsh, unpalatable grasses, took over. The browsing species of horses became extinct and were replaced by hypsodont species (see Chapter 10). When later a mesic period returned, several species of horses shifted back to browsing but retained their high teeth. In other cases, the return to an earlier environmental condition will be answered by a reversal of the selection. When industrial pollution was drastically reduced in recent years, the frequency of the black phenotype of the peppered moth (Biston betularia) also drastically declined in parallel with the reduction of soot and sulfur dioxide.