CHAPTER 8
THE UNITS OF DIVERSITY: SPECIES
The early naturalists of Europe had no idea of the overwhelming richness of the world’s organic diversity. The more conspicuous animals and plants of their neighborhood were all they knew. But this changed rapidly after the Middle Ages. The exploring voyages of the sixteenth to nineteenth centuries revealed the fact that each continent had an indigenous biota and also that there were great latitudinal differences, with the tropics having a very different life from the temperate and arctic regions. Oceanic research revealed a rich marine life, from the surface down to the greatest ocean depths, and the microscope disclosed the enormous world of planktonic and soil eukaryotes, small arthropods, algae, fungi, and bacteria. And this was not the end of the discoveries. Paleontology added an entire new dimension, the life of past geological periods.
This is not the place for a review of the enormous achievement of taxonomy to have described and classified nearly four million species of organisms (with somewhere between five and twenty million species still remaining undescribed). Instead I will focus on an explanation of the evolutionary aspects of this amazing diversity.
HOW MANY SPECIES OF LIVING ORGANISMS?
Few nonspecialists realize how difficult it is to answer this question. First of all, the agamospecies of asexual organisms, particularly of Prokaryotes, are something entirely different from the biological species of the sexually reproducing taxa. More importantly, the majority of taxa are still poorly known. It occurs commonly that in a revision of a tropical genus of insects or spiders, 80 percent of the recognized species are new to science. The same is true for nematodes, mites, and numerous obscure groups. In 1758 Linnaeus knew some 9,000 species of plants and animals. By now about 1.8 million species of animals have been described (excluding agamospecies) and the grand total of species is estimated to be at least 5 to 10 million. Most of these live in the canopy of the tropical rain forest and, with 1–2 percent of this forest being destroyed every year, this number will soon be reduced appreciably.
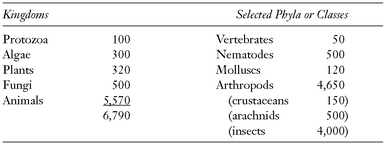
TABLE 8.1 Number of Described Living Species (in thousands)
SOURCE: From May (1990).
The figures suggested by Robert May in
Table 8.1 are very conservative. They are based on the biological species concept. If one uses instead a typological (including phylogenetic) species concept (see below), one can more than double these figures. May’s figures are also low because they do not allow for sibling species. A figure of 5.57 million species for living animals is surely too low, but other estimates that range as high as 30 million are surely much too high. The greatest value of these figures is for comparative purposes. For example, the terrestrial warm-blooded mammals have less than half as many species (4,800) as the warm-blooded aerial birds (9,800 species) (
Table 8.2).
Mammals and birds are best known, yet even in birds about three new species are discovered each year, and in mammals, in addition to bats and rodents, spectacular new large mammals were recently discovered in Vietnam. The figure of 9,800 species for birds is based on a liberal interpretation of polytypic species, in which peripherally isolated populations are mostly listed as subspecies (see
Fig. 8.1 for an example). If many of these were ranked as allospecies, the number of bird species could rise to 12,000. By far the largest group of animals are the beetles. For many families of animals, even for some orders and classes, there is at the present time not a single specialist in the world. It is feared that the description of the hitherto unknown species of organisms will proceed in the future at a slower rate than in the past. For a survey of this problem see May (1990).
TABLE 8.2 Number of species in Major Classes of Vertebrates
| Teleost fishes | 27,000 |
| Amphibians | 4,000 |
| Reptilians | 7,150 |
| Birds | 9,800 |
| Mammals | 4,800 |
Naturalists have long been faced by a puzzling conflict. On one hand, there is a pervasive continuity in the gradual change of the populations of a species through time and space and, on the other hand, there are gaps between all species and all higher taxa. Nothing has more impressed the paleontologists than the discontinuous nature of the fossil record. This is the reason why so many of them were such strong supporters of saltational theories of evolution. However, because we now know that saltations do not occur, we must ask the question: How do the gaps between species originate?
SPECIES CONCEPTS AND SPECIES TAXA
Obviously one cannot study the origin of gaps between species unless one understands what species are. But naturalists have had a terrible time trying to reach a consensus on this point. In their writings this is referred to as “the species problem.” Even at present there is not yet unanimity on the definition of the species. There are various reasons for these disagreements, but two are most important. The first is that the term species is applied to two very different things, to the species as concept and to the species as taxon. A species concept refers to the meaning of species in nature and to their role in the household of nature. A species taxon refers to a zoological object, to an aggregate of populations that, together, satisfy the definition of a species concept. The taxon Homo sapiens is an aggregate of geographically distributed populations that, as a whole, qualify under a particular species concept (see below). The second reason for “the species problem” is that within the last 100 years most naturalists have changed from an adherence to the typological species concept to acceptance of the biological species concept.
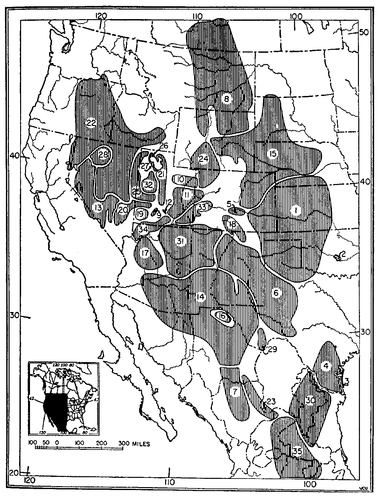
Polytypic species. The distribution of 35 subspecies of the kangaroo rat Dipodomys ordii Woodhouse. Numbers designate the ranges of various subspecies.
Source: Mayr 1969.
If the differences among the populations throughout the geographic range of a species are minor, not justifying taxonomic recognition, a species is called monotypic. Quite often, however, certain geographic races of a species are sufficiently different to be recognized as subspecies. A species taxon consisting of several subspecies is called a polytypic species (
Fig. 8.1).
Species Concepts
Traditionally any class of objects in nature, living or inanimate, was called a species if it was considered to be sufficiently different from any other similar class. Such a species has a number of species-specific characteristics by which it can be distinguished from other species. Philosophers referred to such species as “natural kinds.” This species concept, in which the species is considered to be a well-circumscribed class, is called the typological species concept. According to this concept, a species is a constant type, separated from any other species by an unbridgeable gap. In sexually reproducing species at a given time, it is usually not difficult to sort the organisms one finds at a given place into different species. One refers to the stated conditions as the “nondimensional situation.” Such species coexist at the same time and at the same place and are usually separated from each other by a well-defined discontinuity.
Toward the end of the nineteenth and at the beginning of the twentieth century, more and more naturalists realized that species of organisms are not types or classes, but rather are populations or groups of populations (see Chapter 5). Also, it was found that the basic operational principle of the typological species concept—“species status is determined by the degree of phenotypic difference”—ran into practical difficulties. For instance, sympatric natural populations were found ever more frequently that did not interbreed even though they showed no evident taxonomic differences. This did not fit the typological species definition at all. Such species are now called cryptic or sibling species. These species show the same genetic, behavioral, and ecological differences from traditional species as do phenotypically different species but do not possess the traditional taxonomic differences. Sibling species also occur in plants (Grant 1981) and protists.
SIBLING SPECIES
Coexisting species that do not differ by noticeable taxonomic characters are remarkably common. Malaria in Europe had a very puzzling pattern of distribution until it was discovered that the malaria mosquito, Anopheles maculipennis, was actually an aggregate of six different sibling species, some of them not vectors of the malaria parasite. The famous protozoologist T. M. Sonneborn worked for over 40 years on the ciliate Paramecium aurelia and its varieties, until he realized that it consisted of 14 sibling species. Almost 50 percent of the North American species of crickets were discovered only by their different songs, they are that similar to each other. So far, very little is known about the frequency of sibling species in most phyla and classes of animals (see Box 8.1).
The discovery of what one might consider the opposite situation was equally disturbing for the typological taxonomist. One found in many species individuals that were strikingly different from other members of their population, yet they reproduced successfully with them. The Blue Goose and Snow Goose complex is one example; scores of others are cited in Mayr (1963: 150–158). Both of these situations did not at all fit the typological species definition.
Taxonomists finally came to the conclusion that they had to develop a new species concept, not based on degree of difference but on some other criterion. Their new concept was based on two observations: (1) species are composed of populations, and (2) populations are conspecific if they successfully interbreed with each other. This reasoning resulted in the so-called biological species concept (BSC): “Species are groups of interbreeding natural populations that are reproductively isolated from other such groups.” In other words, a species is a reproductive community. Its reproductive isolation is effected by so-called isolating mechanisms, that is, by properties of individuals that prevent (or make unsuccessful) the interbreeding with individuals of other species.
Box 8.1 Sibling species
Sibling species are natural populations that are reproductively isolated from each other even though they often coexist sympatrically without interbreeding. Yet they are totally or virtually indistinguishable by traditional taxonomic characters. They are remarkably common in many higher taxa.
ARE THERE OTHER SPECIES CONCEPTS AND DEFINITIONS?
In the last 50 years, some six or seven additional so-called species concepts were proposed (Wheeler and Meier 2000). Are these new species concepts legitimate? To summarize my conclusion, they are not. None of the authors of these new concepts has understood the difference between a species concept and a species taxon. Instead of new concepts, they have proposed new operational criteria of how to delimit species taxa (see Box 8.2).
A species concept describes the role that the species plays in the living world. Up to now, only two qualifying concepts have been proposed: a species is either a kind, a different thing, and the species definition specifies the criteria according to which species are delimited (typological concept), or a species is considered a reproductive community (biological concept). There is some leeway in the choice of the criteria by which species can be delimited under a given species concept. In Willi Hennig’s species definition, the biological species concept was adjusted to the needs of cladification to permit the delimiting of appropriate clades. The recognition concept of Hugh Paterson is nothing but a different wording of the BSC. G. G. Simpson’s so-called evolutionary species concept contains undefinable criteria and is useless in praxis. And the various so-called phylogenetic species concepts are simply typological prescriptions of how to delimit species taxa. None of the putative new species concepts is actually a new concept. They are either rewordings of the two standard concepts or instructions on how to delimit species taxa.
Box 8.2 The Three Meanings of Species
The word “species,” unfortunately, has different meaning for different people. It leads to great confusion when these differences are not clearly recognized. Most importantly, one must distinguish three different uses of the word species (Bock 1995).
The species concept. I have described how the typological species concept, the concept prevalent among all classic taxonomists, was supplemented (and largely replaced) toward the end of the nineteenth and beginning of the twentieth century by the biological species concept (BSC). Philosophers have referred to typological species as natural kinds. This typological concept is in conflict with the populational nature of species and with their evolutionary potential. Whenever one is in doubt whether to recognize a particular population as a species or not, one can apply the yardstick of the biological species concept—reproductive compatibility. When one is dealing with sympatric populations, the decision is usually clear-cut. However, when allopatric populations are involved, it must be inferred whether they do or do not have the degree of incompatibility one would find in sympatric species. Inevitably such an inference will be somewhat arbitrary. Only two species concepts are or have been in general use, the typological and the biological.
The species taxon. When species are studied over geographical space, it is found that most of them consist of numerous local populations that differ either slightly or more drastically from each other. Such an assemblage of populations distributed in geographic space is a species taxon, as defined by the biological species concept. A species taxon is always multidimensional, whereas the species concept is based on the nondimensional situation. Species taxa that have well-defined subdivisions (subspecies) are called polytypic species.
The species category. This is the rank in the Linnaean hierarchy given to a taxon considered to be a species. The agamospecies recognized by the students of asexual organisms are also ranked as species in the Linnaean hierarchy, even though they do not form populations in the sense of the populations of biological species.
The biological species concept is applicable only to sexually reproducing organisms. Asexual organisms are assigned to agamospecies (see below). In recent years, various other species concepts were proposed, but none of them has been able to take the place of the biological species concept.
The paleontologist G. G. Simpson thought that in paleontology one needed a separate species concept and proposed the evolutionary species concept. However, his definition contains several criteria that cannot be defined. Furthermore, his species definition does not help in the delimitation of species in a phyletic lineage. The phylogenetic species concept is not a concept at all but simply a typological instruction on how to delimit species taxa in a phylogenetic tree. Likewise, the recognition species concept is simply a different formulation of the biological species concept.
THE MEANING OF SPECIES
A Darwinian always wants to know why each property of a living organism has evolved. So, he or she asks, “Why are there species? Why are living individuals of sexually reproducing organisms combined into species? Why does the living world not simply consist of independent individuals, each reproducing with some other, somewhat similar individual that he or she encounters?” The reason is obvious, and the study of hybrids between species gives the answer to these questions. Hybrids (particularly in genetic backcrosses) are almost invariably inferior, and often inviable or more or less sterile. This is particularly true for animal hybrids. This demonstrates that genotypes, being well-balanced and harmonious systems, have to be very similar for successful interbreeding. If they are not, as is usually true for the product of species crosses, the hybrid zygotes are apt to be an unbalanced, disharmonious combination of parental genes, resulting in more or less inviable or sterile individuals.
The meaning of species is now quite obvious. The isolating mechanisms of species are devices to protect the integrity of well-balanced, harmonious genotypes. The organization of individuals and populations into species prevents the breakup of well-balanced, successful genotypes as would occur if they crossed with alien, incompatible genotypes, and so it prevents the production of inferior or sterile hybrids. Therefore the integrity of species is maintained by natural selection.
Isolating Mechanisms
But what are these isolating mechanisms? Their definition is: Isolating mechanisms are biological properties of individual organisms that prevent the interbreeding of populations of different species where they are sympatric.
This definition makes it quite clear that geographic barriers or any other kinds of purely extrinsic isolation are not isolating mechanisms. For instance, a mountain range that separates two populations that would be able to interbreed if sympatric is not an isolating mechanism. Also, isolating mechanisms, particularly in plants, are often “leaky,” that is, they do not prevent the occasional “mistake” that results in the production of a hybrid. However, such occasional hybridism is not sufficiently successful to lead to a general interbreeding and fusion of the two species populations.
Various ways to classify isolating mechanisms have been suggested. The one I have adopted arranges them in the sequence in which these barriers have to be overcome in potential mates (
Table 8.3).
Different groups of organisms may have different isolating mechanisms. Mammal and bird species, for instance, are usually kept apart primarily by behavioral incompatibilities. Such species may be fully fertile, as are many species of ducks, yet fail to mate. It is not correct to assume that sterility is the prevailing isolating mechanism. Sterility is apparently more important in plants than in animals, because fertilization in plants is “passive,” that is, it is effected by wind, insects, birds, or other extrinsic agents. For this reason, hybrids usually occur more frequently in plants than in higher animals. Yet the production of occasional hybrids leads only rarely to a complete fusion of the two parental species. In plants, however, hybridization may lead through allopolyploidy to the production of new species (see Chapter 9). The study of the genetic basis of the various isolating mechanisms is still in its infancy. The number of genes involved to establish reproductive isolation ranges from one, as in the ratio of pheromones in two butterfly species, to the 14 or more that account for the sterility of hybrid males between two closely related species of Drosophila.
TABLE 8.3 Classification of Isolating Mechanisms
| 1. Premating or prezygotic mechanisms: Mechanisms that prevent interspecific matings. |
| (a) Potential mates are prevented from meeting (seasonal and habitat isolation) |
| (b) Behavioral incompatibilities prevent mating (ethological isolation) |
| (c) Copulation attempted but no transfer of sperm takes place (mechanical isolation) |
| 2. Postmating or postzygotic mechanisms: Mechanisms that reduce full success of interspecific crosses. |
| (a) Sperm transfer takes place but egg is not fertilized (gametic incompatibility) |
| (b) Egg is fertilized but zygote dies (zygotic mortality) |
| (c) Zygote develops into an F1 hybrid of reduced viability (hybrid inviability) |
| (d) F1 hybrid is fully viable but partially or completely sterile, or produces deficient F2 (hybrid sterility) |
Hybridization
Hybridization is traditionally defined as the intercrossing of established species. A hybrid is the product of such a cross. Gene exchange among different populations of the same species is frequent (referred to as gene flow), but should not be called hybridization. Rather, hybridization occurs whenever the isolating mechanisms are inefficient (“leaky”). Successful hybridization leads to the transfer (“introgression”) of genes of one species into the genome of another species. In some populations, particularly highly inbred ones, this may lead to an enhancement of fitness.
The frequency of hybridization is highly variable. It is rare in most higher animals but frequent in an occasional genus. For instance, there is extensive hybridization among the six species of ground finches (
Geospiza) on the Galapagos Islands, without apparent loss of fitness. It is also frequent in some families of plants. In spite of the frequency of introgression in such families, hybridization apparently only rarely leads to a fusion of two species and even more rarely to the production of a new species. In plants the doubling of the chromosome number of a sterile species hybrid may lead to the production of a near-fertile allotetraploid species (see
Fig. 5.2). In certain groups of vertebrates (reptiles, amphibians, and fishes), species hybrids may shift to parthenogenesis and function as separate species. The F1 hybrid generation may show increased viability (“hybrid vigor”) in some species crosses, but this is reversed in the F2 and later generations and in backcrosses. In general, hybrid zones occur when two populations (“species”) that have not yet acquired fully effective isolating mechanisms come into secondary contact.
Species Specificity
Even though every individual in a population is uniquely distinct, and every local population is genetically somewhat different from all others, this variability within a species does not mean that members of a species do not share “species-specific” characters. However, these characters are not constant, like an essence, but are always somewhat variable and, more importantly, they have the capacity to evolve in subsequent generations. By far the most important species-specific characters are the isolating mechanisms; others may be ecological properties, such as niche preference.
In spite of numerous diversifying local factors, the continued maintenance of every species is assured by a number of integrating processes. Most important among these is gene flow (see Chapter 5). Equally important is the basically conservative nature of the genotype. The average genotype of a local population is the result of hundreds or thousands of preceding generations of natural selection. Any deviation from this optimum is apt to be selected against by normalizing selection.
However, the selection factors are not the same everywhere in the range of a given species. There is, for instance, the latitudinal change of temperature, and local populations of many species are selected to be best adapted for the local temperature. This results in gradients of characteristics in such a species that parallel the climatic gradients. Such a character gradient is called a cline. A cline always refers to a particular character. The geographic variation of a species may involve as many clines as it has geographically variable characters.
Species in Asexual Organisms (Agamospecies)
The equivalent of biological species of sexually reproducing organisms does not exist in asexual organisms. Reproductive communities, such as biopopulations, do not exist in the prokaryotes. Hence, there is considerable uncertainty of how many “species” of bacteria to recognize. Furthermore, bacteria as different as eubacteria and archaebacteria, sometimes classified in two different kingdoms, are known to exchange genes quite frequently by lateral transfer. In such cases, one is forced to fall back on the typological species definition and recognize these species, so-called agamospecies, by the degree of difference.
However, asexual reproduction is also widely found in the eukaryotes. Each asexually reproducing individual belongs to a clone of genetically identical individuals. Whenever a new mutation occurs, it signifies the origin of a new clone. Each clone is a target of selection. Owing to natural selection, many clones are eliminated, producing gaps between bundles of successful clones. If these bundles are separated from each other by sufficiently large gaps, they are considered different species. Speciation in prokaryotes, induced by mutation and the extinction of intermediate clones, is something entirely different from speciation among biological species. Agamospecies (asexual lineages), considered to be as different from other groups of such lineages as are biological species taxa, are ranked in the Linnaean hierarchy as species.
In the next chapter I will show how new species can be produced in spite of the various isolating mechanisms to preserve the cohesion of the existing species.