The array of Late Cretaceous creatures from Transylvania goes well beyond its dinosaurs. Although they were the most obvious animals in the Haţeg landscape, Zalmoxes, Struthiosaurus, Magyarosaurus, Telmatosaurus, and others were among the rarest members of the fauna. Just as every ecosystem has its myriad players—primary producers, herbivores, predators, decomposers, and many more, so we must look for more than just the dinosaurs of Transylvania for a more complete picture of life in this region some 72 million years ago. Who were these other creatures also making a living there?
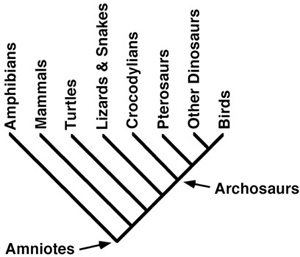
To answer this question, we turn to all of the non-dinosaurian members of the Transylvanian vertebrate assemblage known to date. To cover them in a reasonably logical sequence, we order our discussion around their kinship to dinosaurs (figure 3.1). As pterosaurs are the amniotes (land-dwelling vertebrates) most closely related to dinosaurs, we begin with these flying reptiles. Thereafter, we turn to crocodilians, squamates (lizards and snakes), turtles, and mammals. Finally, we will step outside of amniotes to cover the amphibians and fish from the Late Cretaceous of Transylvania.
Pterosaurs have a unique position in the Mesozoic world—they were the first fully flying tetrapods, establishing themselves as masters of the air as early as the Late Triassic and dominating the skies throughout the world for over 170 million years. Much of what we know about these flying reptiles comes from the Jurassic of Europe, but the Cretaceous pterosaur record indicates their presence on all continents, with the exception of Antarctica.1 By the Cretaceous, a variety of long-tailed forms had come and gone, to be replaced by short-tailed Pterodactyloidea, which were to become the largest flying animals—some species with a wingspan of nearly 12 m—ever to evolve.
Figure 3.1. Evolutionary relationships of Amniota
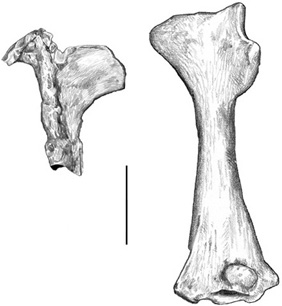
Wonderfully preserved bones of these flying behemoths have been recovered in abundance from Cretaceous beds in North and South America, although specimens from the Late Cretaceous of Europe are quite rare, known from only three localities: Choceň in northeastern Czechia (Czech Republic), Muthmannsdorf in eastern Austria, and Sânpetru and Vălioara in Romania. In 1881, Anton Fritsch, a Czech paleontologist, described some new fossils that he called Cretornis. Although he originally thought that they belonged to a bird, some small, well-preserved wing bones were collected that helped him identify this animal as a pterosaur. Fritsch then referred his Cretornis material to Ornithocheirus as a new species, O. hlavatschi, named after the pharmacist Hlaváč in the town of Choceň who had collected the fossils.2 Two years later, Seeley recognized the first pterosaur material (an articular bone from a lower jaw that Bunzel had interpreted as belonging to a lizard) from the Gosau fauna of Muthmannsdorf, Austria, calling it Ornithocheirus bunzeli (figure 3.2).3
Figure 3.2. Humeri of Ornithocheirus bunzeli (left) and Ornithocheirus hlavatschi (right). Scale = 2 cm
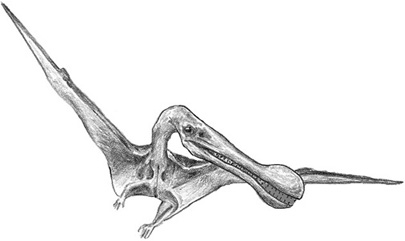
Even though all of these forms were called Ornithocheirus, the first known pterosaur bearing this name didn’t come from Austria, Czechia, or Romania. Ornithocheirus itself was originally described by Seeley on the basis of fossils found in mid-Cretaceous rocks in England.4 No complete skeletons of this pterosaur have ever been found, but we know from fragments of the skull, individual limb bones, and vertebrae that Ornithocheirus was a large pterosaur with a long, slender skull and probably a bony crest on the snout (figure 3.3). The jaws were lined from front to back with sharp teeth, suggesting that it was probably a fish eater.
Figure 3.3. A reconstruction of Ornithocheirus
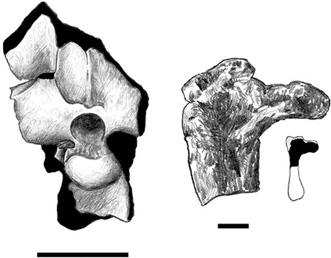
Nopcsa announced the first pterosaur from the Haţeg fauna in 1914.5 This specimen consists of several fused vertebrae in the shoulder region, which in some pterosaurs and birds is known as a notarium. In 1926, Nopcsa again mentioned this pterosaur in a discussion of other Late Cretaceous pterosaur material from Europe, but this time he provided further taxonomic information.6 He regarded the Haţeg pterosaur as a member of Ornithocheiridae, perhaps related to the poorly known Ornithodesmus from the Early Cretaceous of England (which is now thought to be a maniraptoran theropod dinosaur).7 Nopcsa never elaborated on these two comments, and he never described or illustrated the material. To make matters worse, the specimen was thought to be lost from the 1930s onward—that is, until 1995, when the two of us rediscovered the misplaced specimen in the collections of the Magyar Állami Földtani Intézet in Budapest.8 Our preliminary study of this specimen indicated that Nopcsa was correct and that this was a pterosaur notarium. However, more recent work suggests that this specimen may instead be a partial sacrum of a maniraptoran theropod.9 Whether a pterosaur notarium or a maniraptoran sacrum (it is difficult to say, and this certainly needs more study), we are fortunate in having additional pterosaur specimens—a right humerus, a left femur, and possibly another notarium—more recently discovered in the same area of Transylvania (figure 3.4). On the basis of all of the Sânpetru material, we suggested that this pterosaur may have been a member of Dsungaripteroidea, but one for which we could make no further resolution (box 3.1). Unlike its closest relatives, many of which had wingspans of up to 7 m, the Haţeg dsungaripteroid must have been one of the smallest, measuring no more than 75 cm from wingtip to wingtip, about the size of many present-day bats.
Figure 3.4. A pterosaur notarium (bottom left) and femur (bottom right) from Sânpetru. The figure of Pteranodon (above) indicates the position of these two elements and the humeri from figure 3.2. Scale = 1 cm
The most recently discovered pterosaur material from the Haţeg Basin, even more dramatic than that previously found in Romania, was recovered from the Densuş-Ciula Formation near the village of Vălioara.
BOX 3.1 Evolutionary Relationships in Pterosauria
Ranging in size from a sparrow to a small airplane, pterosaurs dominated the skies of the Jurassic and the Cretaceous, becoming extinct at or near the Cretaceous–Tertiary boundary. These flyers have been placed into two groups: long-tailed rhamphorhynchoids and short-tailed pterodactyloids. However, only the latter is monophyletic, while the paraphyletic former consists of basal taxa sequentially less closely related to Pterodactyloidea.

While this much is known about the phylogeny of Pterosauria, the more fine-grained relationships have been harder to come by. Recent cladistic studies by Alex Kellner (Museu Nacional in Rio de Janeiro, Brazil) have gone a long way to improve this situation. According to Kellner, pterodactyloids are divided into Archaeopterydactyloidea and Dsungaripteroidea. The former includes Pterodactylus, Germanodactylus, Ctenodactylidae, and Gallodactylidae, while the latter consists of Nyctosauridae and Ornithocheiroidea. This latter taxon includes Pteranodontoidea (Pteranodon, Ornithocheirus, and Anhanguera, among others) and Tapejaroidea (Dsungaripterus, Noripterus, Tapejara, Quetzalcoatlus, Azhdarcho, and others).
Note: See Kellner 2003.
Figure 3.5. A rear view of the braincase (left) and a ventral view of the proximal humerus (right) of Hatzegopteryx thambema. Scale = 10 cm
Consisting only of the back end of the skull and the left humerus, these bones were originally thought to belong to a moderately large theropod.10 However, a new study by Eric Buffetaut, Dan Grigorescu, and Zoltán Csiki has shown it to be a gigantic new flying reptile (figure 3.5), which they named Hatzegopteryx thambema (“the monstrous wing from Haţeg”).11 Hatzegopteryx, in contrast to the earlier-mentioned pterosaur, is currently thought to be a huge azhdarchid pterosaur, with an estimated wingspan greater than 12 m and a skull perhaps 3 m long, making it, if not the largest, then at least one of the largest flying animals ever to have evolved (figure 3.6).
Instead of having a wing like a bird or a bat, the wing membrane in all pterosaurs was supported solely by an elongate fourth finger. This membrane was reinforced by closely packed, parallel, flexible rods to maintain the airfoil dynamics of the wing. In addition, the bones of the skeleton are lightweight (as in birds), hollow, and thin-walled, yet strong. The small Transylvanian pteranodontid assuredly would have been capable of powered flapping flight that was both agile and maneuverable, but for Hatzegopteryx, the situation would have been different. For one thing, the huge head raises the question of how the animal was able to fly at all with such a heavy object stuck on the front of its body. The secret appears to be that the skull’s internal bone was constructed of a dense network of very thin trabeculae enclosing small spaces.12 This polystyrenelike structure would have been exceptionally well designed to combine strength with lightness, of obvious necessity for a flying creature. However, unlike the powered maneuverability of the small Haţeg pterosaur, we expect that Hatzegopteryx would have elegantly soared across the sky, silhouetted against the clouds.
Figure 3.6. A reconstruction of Hatzegopteryx thambema, after Quetzalcoatlus northropii, a closely related azdarchid pterosaur. Scale = 4m
Instead of making more specific claims about the form and lifestyle of the Haţeg pterosaurs, we can only form opinions by looking at their closest relatives. When ambling on the ground, pterosaurs appear to have had an awkward, quadrupedal gait,13 and this condition probably holds for the two Haţeg pterosaurs. In keeping with their presumed warm bloodedness, the body (but not the wings) of these fliers is thought to have been covered with a furlike pelage. The neck may have been long and the tail very short, as in other pterodactyloid pterosaurs. The Haţeg pterosaurs may have had a crest on the front of the upper jaw and perhaps on the undersurface of the lower jaw. Such a crest may have stabilized the head during flight—sort of like sticking the rudder in front instead of behind, as in most present-day airplanes—or it may have had some sort of significance in courtship and territorial displays, much like the cranial ornamentation in hadrosaurid dinosaurs we previously discussed.
What these Haţeg pterosaurs ate is anything but obvious. Based on evidence from other pteranodontid and azhdarchid pterosaurs,14 the snout was probably long, narrow, and pointed, and the jaws were almost certainly edentulous. The giant pterosaurs may have ventured far out to sea, skimming over the surface of the water to feed on fish. Alternatively, they may have been attracted to a more terrestrial food source, feeding on carrion, such as the carcasses of dinosaurs. In the case of Hatzegopteryx—by far the largest predator known from the Haţeg assemblage—it might be at the top of the food chain, the top predator among the area’s fauna.
Turning from the air back to land and water, we now consider the other major group of archosaurs represented in the Haţeg fauna: Mesoeucrocodylia. The remains of at least five crocodilian-related forms have been recovered from Transylvania. Two are ziphodonts (Doratodon and an as-yet unnamed form with molariform teeth), whereas another is a short-snouted taxon closely related to Acynodon (figure 3.7).15 The fourth, a newly recognized species named Theriosuchus sympiestodon, is another small, short-snouted terrestrial form that appears to be a relict taxon, given its older congeneric relatives from elsewhere in world (figure 3.8).16 The fifth, Allodaposuchus precedens (figure 3.9), is known from a portion of the skull roof, teeth, vertebrae, and limb elements, which were originally collected in 1914 at Vălioara, from the Densuş-Ciula Formation, by Ottokar Kadić, the chief geologist at the Magyar Állami Földtani Intézet, as well as a recently discovered, nearly complete skull collected by members of the joint expedition from Universitatea Babeş-Bolyai Cluj Napoca and the Institut Royal des Sciences Naturelle de Belgique.
Figure 3.7. The teeth of Doratodon (right) and a short-snouted taxon closely related to Acynodon (left). Scale = 1 cm. (After J. Martin et al. 2006)
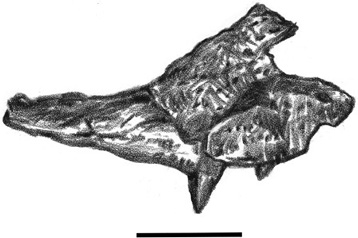
Figure 3.8. The maxilla of Theriosuchus sympiestodon in lateral view. Scale = 1 cm. (After J. Martin et al. 2010)
Nopcsa first described Kadić’s material in 1915.17 Originally not impressed by the details of his new material, Nopcsa considered this Haţeg crocodilian an example of Crocodilus affulvensis, otherwise know from the French Late Cretaceous and named by Philippe Matheron.18 However, by 1928, he had opportunity to examine Matheron’s original material and decided that the Haţeg form was not the same kind of crocodilian, so he provided it with a new name, Allodaposuchus precedens.19
Figure 3.9. A reconstruction of the head of Allodaposuchus precedens (above), a dorsal view of the skull roof of A. precedens (bottom right), and the newly discovered skull of A. precedens in dorsal view (bottom left). Scale = 5 cm. (Newly discovered skull after Delfino et al. 2008)
Based on Nopcsa’s original material and a newly discovered skull, we know that Allodaposuchus was a medium-sized crocodilian for its time, probably 2 to 2.5 m long.20 Like most present-day crocodilians, its skull is moderately long, deep, profusely ornamented with pits, and thick-boned. The largest teeth in the lower jaws insert into notches on the sides of the snout, but the number of teeth is fewer than those seen in other crocodilians the size of Allodaposuchus. The eye sockets are large and indicate that the eyes and their protective lids stood up from the top of the head. What is known of the rest of the skeleton is limited largely to limb elements that, according to Mason Meers at the University of Tampa, suggest that Allodaposuchus was probably a lightly built, mostly terrestrial crocodilian.21
In 2001, we joined Angela Buscalioni and Francisco Ortega from the Universidad Autónoma de Madrid to explore the evolutionary relationships of Allodaposuchus.22 According to our cladistic analyses (box 3.2), Allodaposuchus appears to occupy a position very close to the base of the group comprising all living crocodilians. A similar position—basal in Eusuchia—was also assigned in a 2008 analysis Delfino et al. made after their discovery, based on a new and nearly complete skull of Allodaposuchus, so we can be doubly sure that this basal position is likely to be correct.23
Much less is known (from isolated teeth only) about Acynodon, another of the Haţeg crocodilians, than about Allodaposuchus.24 These teeth had short crowns that were globular in occlusal view. A recent study of Acynodon by Delfino, Martin, and Buffetaut, based on much better material (cranial and postcranial remains from the Santonian of northeastern Italy), indicates that it was a short-faced alligatoroidean.25 Its jaws were lined with molariform teeth and, unlike many other crocodilians, there was no caniniform tooth. This dentition, as well as aspects of the skull, suggest that Acynodon fed on a mixture of invertebrates (mollusks and crustaceans) and possibly plants.
Finally, Doratodon from Transylvania is also known only from isolated teeth.26 Similar to those from elsewhere in Europe, the Transylvanian teeth are compressed transversely, with large serrations on the forward and rearward edges. A newly described, nearly complete mandible from Spain indicates that Doratodon was relatively small and had a short, narrow rostrum equipped with a reduced, heterodont dentition. On the basis of these and other features, Julio Company and his colleagues considered Doratodon to be a mesoeucrocodilian, positioning it as the sibling taxon to Sebecosuchia within Ziphosuchia.27
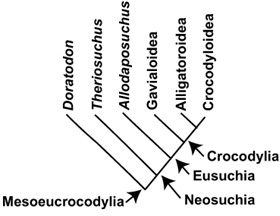
BOX 3.2 Evolutionary Relationships in Mesoeucrocodylia
Mesoeucrocodylia contains quite a number of fossil forms at its base, as well as Crocodylia, the great clade of all living crocodilians: Gavialoidea, Alligatoroidea, and Crocodyloidea, plus all their extinct relatives. Close to crown-group Crocodylia—indeed, often regarded as its sister taxon—is none other than our own Allodaposuchus precedens. Theriosuchus and then Doratodon have more distant relationships with crown-group Crocodylia and Allodaposuchus.
Note: See Brochu 1997, 1999; Buscalioni et al. 2001; Delfino et al. 2005; Pol et al. 2009.
These discoveries suggest that there is greater ecological diversity in Transylvanian taxa on the line toward living crocodilians than had been previously recognized.28 We suspect that Allodaposuchus, the best known among them, was probably an opportunistic, highly efficient predator, preferring to hunt and feed mainly at night, as is true in present-day crocodilian forms.29 These prehistoric creatures may have tracked their next meal, overpowering and dismembering the body before swallowing chunks of the carcass in single gulps. The ability of crocodilians to eat almost any animal they can overpower or drag underwater and drown has contributed to their success as predators.
The crocodilian habit of basking during daylight hours has led to the popular misconception that these animals are idle predators with limited social acumen. However, present-day crocodilians are known to engage in complex social behavior, including gregariousness, visual and vocal display, territoriality, and courtship behavior. Mothers build and guard nests, and they aid newborns in their struggle to leave their eggs. In addition, parental care extends well into the lives of these hatchlings. Whether a similar behavioral repertoire was present in any of the Transylvanian crocodilians is, unfortunately, still unknown.
As we have noted, the role of the top dinosaurian predator from the Transylvanian assemblages may have been occupied either by small, agile theropods or perhaps by the enormous pterosaur Hatzegopteryx. Could it be that Allodaposuchus also shared in this predatory biological role? Nopcsa, for one, regarded this ecological status as a distinct possibility. He even suggested that the fragmented, disarticulated dinosaurian remains in places like Sânpetru represented the ancient feeding grounds of Allodaposuchus.30 While we’re not so sure about this, it is true that Allodaposuchus and its crocodilian cohort would have been formidable predators on whatever they could get a hold of, not only the young and debilitated, but also any of the Transylvanian creatures caught unawares.
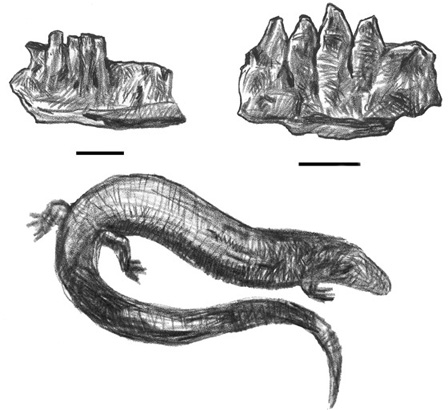
Although archosaurs dominate the Transylvanian assemblages, they do not represent the complete roster of taxa known from this region during the Late Cretaceous. The squamate (lizards and snakes) portion of the Haţeg fauna is known from screening sediment for microfaunal remains. Through these efforts, we know that scincomorphs predominate (box 3.3). Known principally from jaws, teeth, and isolated parts of the skull, Haţeg forms—such as Becklesius nopcsai, B. hoffstetteri, and Bicuspidon hatzegiensis—were probably swift, long-tailed forms resembling their present-day relatives (figure 3.10 on p. 74).31
In contrast, anguimorphs represent the slow-moving, secretive predators among present-day lizards. Known only from an upper jaw of an indeterminate anguimorph, this clade is very rare in the Haţeg fauna thus far.32
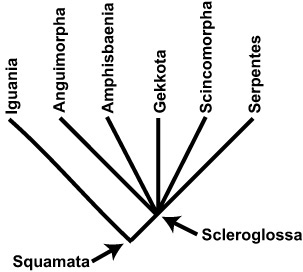
BOX 3.3 Evolutionary Relationships in Squamata
Living snakes, lizards, and amphisbaenians, plus all the descendants of their most recent common ancestor, form the clade known as Squamata (meaning “the scaled ones”). Systematically, squamates include iguanians (agamids, chameleons, and iguanids) and scleroglossans. The latter include anguimorphs (monitors, Gila monsters, and alligator lizards, among others), amphisbaenians (worm lizards), geckos, scincomorphs (skinks, whiptail lizards, and others), and serpents (snakes). The phylogenetic relationships of these taxa are provided in the figure.
Note See Vidal and Hedges 2005; Conrad 2008.
Finally, a single vertebra represents all that we know about snakes in the Late Cretaceous of Transylvania. This snake, an indeterminate madtsoiid, apparently killed its prey by constriction, much like present-day boids, to which, however, they are not particularly closely related.33
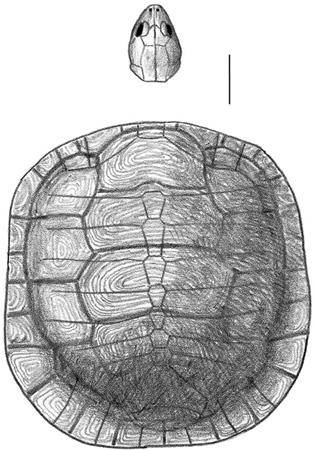
Turtles are ubiquitous—scutes, limb elements, and rare skull materials have been collected nearly everywhere, ranging from localities in the Haţeg Basin to Vurpăr, Jibou, Bărăbanţ, Lancrăm, Sebeş, and Oarda de Jos in the Transylvanian Depression.34 One of these has been known for some time. Named Kallokibotion bajazidi by Nopcsa in 1923,35 it may have been quite abundant: fragments of its shell occur everywhere in rocks from the region, although other parts of the skeleton are relatively rare. Kallokibotion was a moderately large, broad-headed turtle, with an oblong shell measuring much as 50 cm in length and 40 cm in width (figure 3.11), about the size of a present-day snapping turtle. Like present-day turtles, it was toothless, and the margins of its upper and lower jaws were covered by a horny beak. A slow ambler on land, Kallokibotion most likely fed on low-lying foliage and fleshy fruits. Living turtles lay clutches of leathery eggs, which are then abandoned; there is no parental care. We think this may also have been the case with Kallokibotion.
Figure 3.10. The scincomorphs Becklesius nopcsai (top left), Bicuspidon hatzegiensis (top right), and a modern scincomorph, Chalicides chalicides (the three-toed skink) (bottom). Scale = 1 mm (top left), 500 mm (top right)
Nopcsa regarded Kallokibotion as closely related to several primitive cryptodiran turtles, those forms that pull their heads into their shells by folding their necks vertically (the other major group of turtles are pleurodires—side-necked turtles—that bend their necks sideways). Recent studies now position it as the sister taxon to Testudines, the crown group consisting of the most recent common ancestor of all cryptodires and pleurodires and all the descendants of this ancestor (box 3.4).36
Figure 3.11. A restoration of the skull and carapace of Kallokibotion bajazidi. Scale = 5 cm
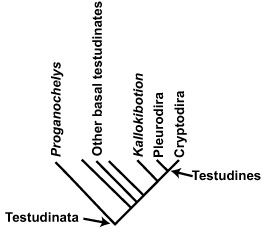
BOX 3.4 Evolutionary Relationships in Testudinates
Turtles—all living forms and a myriad of fossil taxa—are scientifically known as Testudinata. Characterized by the presence of a shell (composed of a dorsal carapace and a ventral plastron) that completely encloses both of the limb girdles, the absence of teeth lining the jaws, and other features, turtles are known from as far back as the Late Triassic and Early Jurassic, not only from their most primitive taxon, Proganochelys, but also from the earliest members of many of today’s major groups.
Testudines basally consist of Proganochelys and Rhaptochelydia, the latter divided into Austrochelyidae and Casichelydia. Casichelyds make up all modern turtles and their extinct relatives, including Pleurodira (side-necked turtles) and Cryptodira (hidden-necked turtles). The monophyly of these two groups is well supported by both morphological and molecular evidence. Cryptodires make up the greatest diversity among living and extinct turtles. Basally, this group includes the Early Jurassic Kayentachelys from Arizona, an unnamed clade consisting of Meiolaniidae and others, and Selmacryptodira. Kallokibotion is thought to be the sister taxon of Diacryptodira, consisting of the remainder of all cryptodirans.
Note: See Hirayama et al. 2000; Joyce 2007.
Another turtle—a yet-to-be-determined dortokid—has recently been announced by Vlad Codrea and his coworkers.37 Previously known from the Cretaceous of southern France and northern Spain, and from the early Tertiary of Romania,38 they have now been collected from Oarda de Jos and Vurpăr. Dortokids are thought to be primitive pleurodires.
By this time in Earth’s prehistory, there was one great mammalian dynasty—Multituberculata—and, emerging from its shadow, two fledgling groups, marsupials and our own group, placentals (box 3.5). The most diverse and abundant of Mesozoic mammals, multituberculates are first known with certainty from Upper Jurassic deposits,39 but especially so in the Cretaceous fossil record (they survived the Cretaceous–Tertiary mass extinction, flourishing for another 35 million years before dying out in the early Oligocene 30 million years ago). These mammals ranged in size from that of a small mouse (25 g) to that of a woodchuck (4 kg).
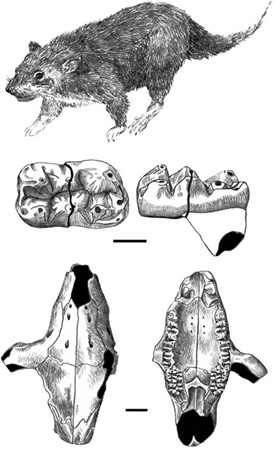
Only multituberculates have been found in the Haţeg strata thus far, but it is not known whether the absence of marsupials and placentals is biologically real or whether they will be discovered eventually in Transylvania. The first multituberculate to be described, in 1985, was based on an isolated upper incisor from the Pui locality, and two lower molars from the same site were described in 1986.40 Known as Barbatodon transsylvanicus,41 the lower molar from this creature was the best material available from the Haţeg Basin until 1996, when the first multituberculate skull from Sânpetru—named Kogaionon ungureanui—was described by two paleontologists from Institutul speleologic “Emil Racoviţă” Bucureşti, Costin Rădulescu and Petre-Mihai Samson (figure 3.12).42 A possible new species of Kogaionon was reported by Vlad Codrea and his coworkers in 2002.43 Finally, a multituberculate called Hainina, otherwise known from Paleocene rocks in western Europe, was also discovered in 2002 from the Late Cretaceous of the Haţeg Basin.44
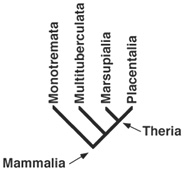
BOX 3.5 Evolutionary Relationships in Mammalia
Mammalia, the large group of vertebrates in which we claim membership, presently dominates the world’s biota of large animals. The northern high latitudes have their musk ox, elk, and polar bears; the tropics, their tapir, giraffe, elephants, rhinos, hippos, tigers, and lions. Placentals all, they are united by having a special construction of the embryonic membranes between mother and embryo called the placenta. By comparison, marsupials have only an incomplete placenta, but most females carry their young in an abdominal pouch called a marsupium. Today, marsupials are best known from Australasia, but they are also found in South America, and—thanks to the pesky Virginia opossum (Didelphis virginiana)—in North America as well. Together, placentals and marsupials are grouped as Theria. The remaining living mammal group, the egg-laying Monotremata, includes platypuses and echidnas from Australia and New Guinea.
Because all these mammalian groups are still living, we consider them crown-group Mammalia, but they alone aren’t all of the major groups represented in this clade. Another group, Multituberculata, is more closely related to Theria than Monotremata is. First appearing in the Late Jurassic and going extinct in the Oligocene, these small herbivores are well represented all over the Northern Hemisphere.
Note: See Kielan-Jaworowska and Hurum 2001; Kielan-Jaworowska et al. 2004; Lillegraven et al. 1979; McKenna and Bell 2000.
Figure 3.12. A reconstruction of Kogaionon ungureanui (top), and the lower left first molar of the Haţeg multituberculate Barbatodon transsylvanicus, in occlusal (middle left) and lingual (middle right) views. Scale = 1 mm. Dorsal (bottom left) and ventral (bottom right) views of the skull of the Haţeg multituberculate Kogaionon ungureanui. Scale = 5 mm. (Top and middle after Rădulescu and Samson 1986; bottom after Rădulescu and Samson 1996)
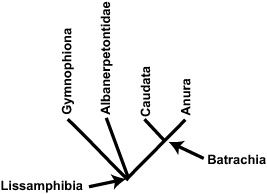
BOX 3.6 Evolutionary Relationships in Amphibia
Modern amphibians (Lissamphibia) include frogs and toads (Anura), salamanders and newts (Caudata, or Urodela), and caecilians (Gymnophiona, or Apoda). In addition, a number of extinct taxa are known within Lissamphibia. As concerns us here, Albanerpetontidae is known from the Jurassic to the Miocene. Members of this wholly extinct clade most closely resemble modern salamanders.
Note: For Gymnophiona, see Frost et al. 2006. For Albanerpetontidae, see Gardner 2000, 2002; Gardner et al. 2003; McGowan 2002; Venczel and Gardner 2005.
Multituberculates have a long, broad skull that is superficially rodent-like in appearance, with long lower incisors, followed by a gap (called a diastema) between them and the cheek teeth. The lower premolars are specialized into shearing blades, whereas the molars are designed for crushing or grinding food, suggesting that multituberculates, including those from Haţeg, had a varied diet that included seeds as well as other larger, hard food items.45
Based on features of the most complete skeletons, paleontologists believe that at least some multituberculates were arboreal.46 With a prehensile tail, a hind limb held in a crouched position, a reversible ankle similar to that seen in squirrels, and a wide range of forelimb motion, these forms could have descended tree trunks headfirst to forage on the ground. However, other multituberculates may have been burrowers, and a third interpretation is that some forms were leapers.47 Whatever the case (and these interpretations are not mutually exclusive), it would be premature to characterize the Transylvanian forms as leapers, burrowers, or tree climbers without any postcranial skeletal material. Whatever their mode of locomotion, Barbatodon transsylvanicus and Kogaionon ungureanui would probably have been found scurrying by moonlight in the mosaic of shadowed alcoves, branches, or undergrowth, as did many nocturnal Mesozoic mammals, since the best escape from predatory dinosaurs was the cover of darkness.
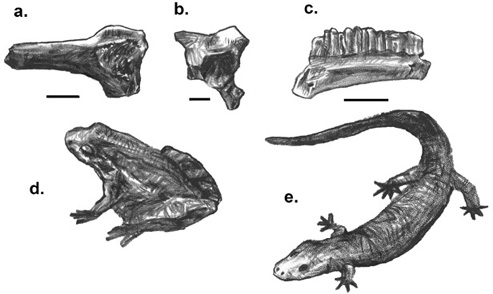
Figure 3.13. The dentaries (with teeth) of (a) Hatzegobatrachus grigorescui (an anuran), (b) Paralatonia transylvanica (an anuran), and (c) Albanerpeton sp. Scale = 0.5 mm. A living anuran, (d) Discoglossus galganoi, and (e) a reconstruction of the extinct Albanerpeton sp.
Active screening for microvertebrate remains in Transylvania in the past two decades has produced a number of new frogs and an albanerpetontid (box 3.6).48
Hatzegobatrachus grigorescui and Paralatonia transylvanicus are small frogs (with an estimated 30–50 mm snout–vent length) known from jaw elements and various postcranial elements. Other indeterminate frogs have been reported from the Haţeg Basin: one that appears to be closely related to Eodiscoglossus, and another having an affinity with Paradiscoglossus (figure 3.13).49
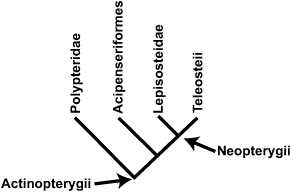
BOX 3.7 Evolutionary Relationships in Actinopterygii
Actinopterygii, also known as ray-finned fish, possess fins supported by lepidotrichia (bony or horny spines). Currently consisting of nearly 30,000 species, actinopterygians are found in both freshwater and marine environments, from the highest mountain streams to the deepest sea. Three groups of actinopterygians (acipenseriforms, lepisteostids, and teleosts) are recognized from the Late Cretaceous of Transylvania.
Note: See Stiassny et al. 1996.
Albanerpeton, an enigmatic, extinct, salamanderlike taxon, is the last of the amphibians recovered from the Late Cretaceous of western Romania.50 Otherwise known from the Middle Jurassic to the Miocene of Europe, North America, and central Asia, albanerpetontids were approximately 10 cm long, predominantly terrestrial (although some may have been fossorial), and predatory.51 Albanerpetontids are placed within Lissamphibia (the crown group that includes frogs, salamanders, and caecilians).52
The Haţeg fauna, despite coming from sediments laid down in streams and rivers (chapter 4), is counterintuitively very poor in fish of any kind (box 3.7). Thus far, the only known fossil remains from members of this fully aquatic community are acipenseriforms (chondrosteans), lepisteostids (holosteans), and teleosts. Living acipenseriforms consist of sturgeons and paddlefish, both with cartilaginous skeletons, elongate bodies, and triangular snouts. Acipenseriforms are bottom feeders. Rhomboidal, thick scales and a conical, slightly recurved tooth attest to the Haţeg presence of the gar Lepisosteus.53 Present-day gars are long-snouted carnivores, with a long, slender body and a heterocercal caudal fin. Finally, characiform teleosts have been reported from the Haţeg Basin.54 Today we know some of these characiforms as the popular aquarium pets called tetras (figure 3.14).
Figure 3.14. Living representatives of the Transylvanian actinopterygians: gar (Lepisosteus oculatus; top); blue neon tetra (Paracheirodon simulans; middle), a characiform teleost; and sturgeon (Acipenser transmontanus; bottom)