What was the Transylvanian region like some 70 million years ago, during the Maastrichtian age of the Late Cretaceous? To Franz Baron Nopcsa, the prehistoric landscape had a distinctly different appearance than it does in modern times. Here is how he might have imagined it, with a bit of a current paleogeographic spin.
Seen from high in the air, the Transylvanian region is an island—a speck among other specks—that, together with its neighbors, forms a chain that extends across the northern flank of a broad seaway. This great body of water, called the Neotethyan Ocean, covers what is now most of Europe and Russia to the north and northern Africa to the south. Some islands are small oceanic volcanoes no more than 100 square km in area, but a few are the size of present-day Madagascar. One of the latter, which would form what is now part of southern France and the majority of Spain, is the westernmost island of the chain, some 2,500 km away from Haţeg Island. The other end of the chain, in what is now the Caucasus, lies 2,000 km away from us, and we are some 3,000 km from the nearest coast of northern Africa (Plate VI).
Thus our island of interest—Haţeg Island—is halfway along this chain, some 20°–30° North Paleolatitude, just below the Late Cretaceous position of the Tropic of Cancer. As we descend through the tropical clouds toward our island, the air becomes more humid and we see several highflying birds and pterosaurs. The green flanks of volcanoes stand out against the deep blue of the surrounding Neotethyan Ocean as we begin to perceive the horseshoe shape of the island and its extent, some 75,000 square km in area (about the size of present-day Sri Lanka).1
As we get ever closer, the neighboring islands disappear from view. An occasional ash cloud drifts up from the cauldron of one of the volcanoes on Haţeg Island, but otherwise the air is clear—clear and thick with humidity and the white noise of insect cacophony. The climate on Haţeg Island is largely controlled by the surrounding warm currents of the Neotethyan Ocean and the prevailing westerly winds. Parts of the island receive abundant but seasonal rains, such that the island undergoes alternating wet and dry periods.
The island’s details emerge and we see not a single geometric unit, but tiers of diverse habitats and their denizens. Beaches ring the periphery; the island is rugged in places, and lava hints at a violent volcanic past. Most of the lower elevations of the island are covered with diverse rain forests. Farther inland, a series of low hills and terraces leads away from the shore. This gently sloping alluvial plain is well drained by numerous, rapidly flowing, shallow rivers. Their channels are interspersed with sand bars and occasional, short-lived, shallow lakes. Abundant fetid swamps occupy the lower-lying regions. Floodplains, on which soils have formed and vegetation flourishes, are broadly distributed among water and wetlands. There is luxuriant plant growth everywhere: dense forests towering above a lush tropical understory, and river banks covered with a riot of colors from flowering angiosperms.
The uplands of the island are often cloudy and humid, although where there is fresh lava, the gray rock is barren and sunbaked. Yet even these highlands have stands of woodland, though not nearly as dense or as common as in the lowlands. Instead, shrubs, weedy plants, and other ground cover dominate the landscape.
Lecturing at the 27 November 1912 meeting of the Zoological and Botanical Society in Vienna, Nopcsa summarized what was then known about the dinosaurs from the Upper Cretaceous of Transylvania.2 He then described their evolutionary relationships within Dinosauria, the conditions under which their remains were found, and their paleoecology, which he regarded as swamp dwelling. In doing so, he emphasized that, compared with the dinosaurs of the same age elsewhere in the world, all of the dinosaurs of Transylvania were strikingly smaller:
During the time of the Upper Cretaceous [Haţeg] formed an island and the smallness of the Transylvanian Cretaceous dinosaurs can be examined as a consequence of insular life [italics in original], which in other groups, as for example in the fossil elephants of the Mediterranean islands, likewise led to a reduction in body size in comparison with the continental forms.3
Figure 4.1. Othenio Abel (1875–1946)
Audience members at this lecture were impressed with Nopcsa’s inference that the members of the Haţeg fauna were dwarfs, but they were somewhat less than sanguine about whether the dinosaurs were dwarfs because of their island habitation. Pointing out that this was the first time that the Transylvanian dinosaurs had been clearly identified as a dwarfed fauna, Othenio Abel (figure 4.1), the session’s convener, objected to Nopcsa’s insular hypothesis, predicating his critique on three arguments.4 First, Abel asserted that the basis for dwarfing could not be directly attributed to island life per se, as was found in the dwarf elephants of Malta and the dwarf hippos of the Mediterranean islands. Instead, he argued that their diminution in size was due to the inevitable inbreeding that comes with insular isolation, as well as through other geographical interventions, such as isolation caused by mountain uplift and river dynamics. As a second objection, Abel noted that insular habitation does not always lead to dwarfing, pointing to examples in which certain reptiles and birds have tended toward gigantism through isolation on islands (e.g., crocodiles in Madagascar, Komodo dragons in Indonesia, turtles from the Galápagos Islands, moas in New Zealand). Third, Abel worried about Nopcsa’s use of the word “island” in reconstructing the Haţeg paleoenvironment, questioning whether the region that supported the fauna was actually small enough to be considered an island, or instead was sufficiently large to be considered a mainland.
Abel’s comments stayed with Nopcsa as he assembled his case for dwarfed Transylvanian dinosaurs. In 1915, he provided further documentation of body-size differences between the animals he had recovered from the Haţeg Basin and those elsewhere, in order to answer Abel’s criticisms.5 While suggesting that dwarfing either was due to the Transylvanian dinosaurs retaining a primitive stage of evolution or was a symptom of evolutionary degeneration, Nopcsa still appears to have been uncertain about the importance of either. As a good neo-Lamarckian, he explicitly advocated degeneration produced by insular isolation, opining that growth in the bones from Transylvania may have been pathologically limited through illnesses (caused perhaps by malnutrition or starvation).6 He called these changes in body size—from dwarfing in the Transylvanian dinosaurs to gigantism among dinosaurs in general—arrostic (literally, “arresting”) alterations.7 For Nopcsa, arrostic changes were nothing more than the heritable acquisition of endocrine disease: hyper-function of the pituitary gland in the evolution of gigantism, hypofunction of the thyroid gland in the evolution of dwarfing. We will return to these biological explanations in chapter 5.
Nopcsa used local geology to explore the paleogeography of the region that would have spurred the evolution of increasingly smaller descendants from larger ancestors. To do so, he took to the rocks, returning to the research he conducted previously while a graduate student at the University of Vienna under the supervision of Eduard Suess.8 Nopcsa’s dissertation research had taken him back home to Transylvania, in the heart of the Southern Carpathians, to conduct a comprehensive and detailed study of the geologic history of the hills, mountains, and rolling plains that he knew so well as a child living in Săcel. In the valleys of the Sibişel, Cerna, and Jiu rivers, the slopes of the Retezat, Sebeş, and Poiana-Ruscă mountain ranges, and the villages of Zeicani, Fărcădin, and Pui, he occupied himself with the real grunt work of a field geologist—walking, sample collecting, note taking, and mapping—as well as the marveling, wondering, and questioning that comes from these physical labors. Nopcsa not only looked at the rocks from oldest to youngest—from the granites, schists, and gneisses dating back to the Precambrian through the overlying sediments dating from the Early Jurassic to the last Ice Age—he also interpreted the ancient environment of depositions and their significance with respect to Earth’s major upheavals.
From Nopcsa’s detailed dissertation research, published in 1905 as a nearly 200-page monograph, we learn that the Haţeg region 150 million years ago was generally a warm, marine-dominated environment.9 The widespread limestones and chalk marls that dominate the Lower Jurassic and Lower Cretaceous were precipitated in the warm Neotethyan Ocean. Nopcsa thought that the central part of the Retezat Mountains, whose uplift he regarded as beginning no later than the Early Jurassic, may already have risen above the sea. Had the Haţeg region been an island in the Late Jurassic and Early Cretaceous? Nopcsa, thus far, remained uncertain. Sedimentary evidence of beaches that would have ringed the island—key to his insular hypothesis—is absent on the flanks of these mountains (though Nopcsa attributed this lack of evidence to erosion occurring since the Jurassic). However, following its emergence during the mid-Cretaceous, the Haţeg region was again covered by a warm sea, its waters abounding with predatory ammonites and the strange, reef-building bivalves called rudists, as well as oysters, clams, snails, and single-celled foraminifera. Some evidence of this remains in the region. Sea level fluctuated greatly through this interval, peaking some 80 million years ago, during the Campanian.
Terrestrial conditions at the very end of the Cretaceous are well preserved in the Transylvanian region. Outcrops of these Maastrichtian rocks are known in the Haţeg Basin itself, as well as in the southern parts of the Poiana-Ruscă Mountains and in the Apuseni Mountains (Plates VII and VIII). Nopcsa interpreted these rocks (a thick sequence of conglomerates, sandstones, and mudstones) as a system of nearshore, fluvial, and lacustrine environments, finding support for his interpretation of terrestrial conditions in the unionid clams, turtles, crocodilians, and dinosaurs often found in abundance at various Transylvanian locations.10 This emergent habitat endured until the early Eocene, approximately 55 million years ago. Sea level rose again in the middle Eocene, as the shales interbedded with coarse limestones overlying the terrestrial deposits testify, only to recede again in the Oligocene.
Nopcsa’s last word on the paleogeography of the Haţeg Basin came in 1923.11 Using the paleogeographic maps available to him at that time, he described the area as a “mere archipelago” of five major islands, distinctly separated from each other and the mainland since the beginning of the Late Cretaceous. Extensive overthrusting, which began some 80 million years ago, produced the present-day Carpathian ranges and the main outlines of current European topography. As Earth’s crust buckled, the seas withdrew, uncovering what were to become the Southern Carpathian Mountains. Here, the Transylvanian dinosaurs and other members of the fauna were entombed in the region’s riverbeds, freshwater lakes, and formidable bogs.
Nopcsa’s schooling at the hands of Eduard Suess introduced him to the possibility that the global positions of the continents may not have been fixed over geologic time. In his nineteenth-century attempts to explain the formation of folded mountains ranges like the Alps, Suess suggested that such major geologic upheavals were due to the gradual contraction of the planet.12 With this shrinking, due to Earth’s cooling, its outer crust was forced to wrinkle, fold, and subside. In so doing, large regions of the crust created depressions into which the seas drained, thereby exposing regions of dry land. As Suess’s student, Nopcsa came to see Earth as a changeable, mobile matrix of continents and oceanic basins formed through shrinking, rather than as a permanently fixed geography.
Nevertheless, Suess’s contractionist theory didn’t survive long into the twentieth century before it collapsed—there were just too many improbable estimates of cooling and contraction rates, a recognition that continental and oceanic crusts were different in their composition and density, and the discovery that radioactivity in the crust created a stable heat balance across geologic time. Yet Suess had opened the door widely for the possibility of large-scale crustal mobility.
Although he was not the first to propose major lateral displacements of the continents, one person has been rightly credited as the principal developer of the ideas of continental drift, Alfred L. Wegner.13 In 1912, Wegener proposed that, in the distant past, all the continents had been united, had later broken apart, and thereafter drifted through the ocean floor to their current locations. Much of his evidence came from the jigsaw fit between the margins of the continents, biogeography, paleoclimatology, and evidence that the present-day continents are actually, though very slowly, moving. Nopcsa himself contributed to Wegener’s theory by incorporating aspects not only of paleontology and biogeography, but also of geochemistry and theoretical geophysics.14 For their time, Nopcsa’s arguments for continental mobility—involving the relationship of different kinds of volcanism that came from the movement of continental plates over the top of zones of subducting oceanic crust—were quite innovative, agreeing with the present-day view of magma origin.15 Nevertheless, many felt that the evidence in support of Wegener’s multi-faceted theory, called continental drift, was ambiguous, and it failed to attract a powerful following—that is, until the late 1950s and the 1960s.
In the 1950s, geophysicists initiated studies of Earth’s past magnetic field, as recorded in geologic deposits, and this work later established considerable variation in the position of Magnetic North through time.16 In particular, the pole position on one continental landmass at any given time in the past was not the same as the pole positions on the others; instead, there were many pole positions. One explanation for this apparent polar wandering was continental drift. By moving the continents across the face of the globe, the pole positions could all become superimposed. Drift received a spark of renewed interest.
In addition, thanks in part to research spawned by the events of World War II, the oceanic basins were investigated as never before. The discovery of oceanic ridges, trenches, volcanoes, and mountains came from mapping the ocean floor using radar, which imparted considerably more topography than had ever been expected before. Abundant shallow-earthquake activity and volcanism were associated with the ridges, whereas deep earthquakes were linked with the trenches. Soon it was discovered that the age of the oceanic crust was not uniform. By mapping, sampling, and radiometrically dating the oceanic crust, it was revealed that the youngest oceanic floor was at the midoceanic ridges, and the oldest was in proximity to the continents. New oceanic crust was formed at the ridges and spread away laterally, indicating that the continents must have been closer to each other in the past. In other words, they had to have moved to their present positions.
As these new data flooded into laboratories during the 1960s, Wegener’s theory of continental drift was to receive a vigorous dusting off. Wegener’s early evidence and all of the more recent discoveries could be causally related in what is now known as plate tectonics. Developed in further detail by geologists, geochemists, geophysicists, and paleontologists, this unified theory links all of the dynamic processes of the globe—how mountains are formed, why volcanoes are distributed in the Ring of Fire around the Pacific margin, how the sea floor is formed, and why certain zones are earthquake prone—with the relative motions of a mosaic of large, rigid, lithospheric plates that constitute Earth’s outermost shell. A variety of motions can occur at the boundaries between these plates. Lithospheric plates moving toward each other produce a zone of convergence, while in moving away from each other they form zones of divergence. Plates can also slide past each other to produce transformational movement, such as that along the San Andreas fault system in California. New plate material is produced at divergent boundaries (e.g., along the crests of midoceanic ridges), while old material is destroyed and recycled by subduction at convergent boundaries, as is the case along the deepsea trenches off the Pacific coast of Japan. Given the available evidence, geologists think that each lithospheric plate floats on Earth’s upper mantle, a 660 km thick, plastic layer of dense, semimolten, magnesium- and iron-rich silicates. Plate motion is driven by large-scale convection currents in this upper mantle. It is this motion, imparted to the continents, which produced the global patterns that originally caught Wegener’s eye.
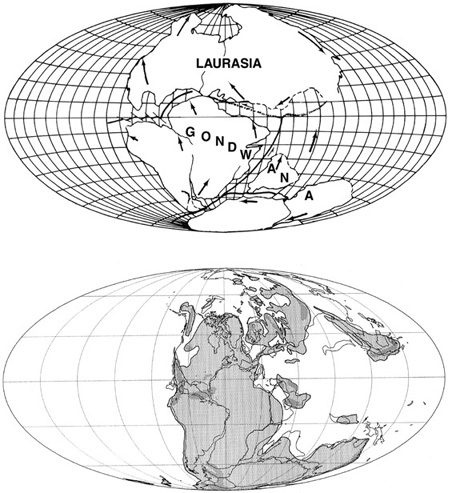
With the acceptance of plate tectonics in the late 1960s, scientists clamored to reconstruct Earth’s geography in bygone times, and in doing so they produced a flurry of paleogeographic maps. At first, these were done by hand, but with the coincident increase in computer access, computer modeling of plate motion and continental repositioning became the working tool of choice. The global models provided in 1970 by Robert S. Dietz and John C. Holden were based on the best fit of the continental positions of present-day coastlines, the margins of the continental slope, and patterns of fracture zones in the ocean floor.17 Later computer simulations, such as those by Alan G. Smith and his collaborators from England, followed suit, but they added further constraints (figure 4.2).18 First, based on their present coastlines and shelf margins, the continents were reassembled into their positions prior to the opening of the Atlantic, Indian, and other oceans, much like the jigsaw-puzzle approach used by Dietz and Holden. Second, a paleogeographic latitude–longitude grid was superimposed on the reassembled continents, and successive stages of breakup—with reference to the positions of the paleomagnetic poles for each interval of time (from those apparent polar-wandering curves)—were determined.19 It is through such paleogeographic reconstructions that we came to know of the existence of the late Paleozoic supercontinent called Pangea, with its huge eastward-opening Tethyan Ocean that provided Pangea with a northern Laurasian half and a southern Gondwanan half. This kind of research has also been used to graphically document the opening of the North Atlantic in the Late Triassic–Early Jurassic, and then the South Atlantic in the mid-Cretaceous.
Figure 4.2. Global paleogeographic reconstructions for the Triassic, approximately 220 million years ago. (Dietz and Holden 1970 reconstruction [above]; A. Smith et al. 1994 reconstruction [below])
As these computer models of plate tectonics became available, other scientists began investigating more fine-grained changes in global conditions. Most important have been new models of paleoclimatologic change, including variations in oceanic circulation patterns, fluctuations in the surface temperature of the oceans and the atmospheric temperature, changes in patterns of atmospheric pressure and circulation, and alterations in weather patterns.20 Data taken from depositional environments (based on surface outcrops and boreholes), as well as information on fossil plants thought to indicate particular environments, can also be plotted on paleogeographic maps to characterize important aspects of regional paleoenvironments.
With both these new global and more regional paleogeographic models, we have come to appreciate that the history of the Tethyan region was much more complicated than previously thought. Instead of a single oceanic basin, it was formed by two different, spreading ridge systems, and therefore had two successive basins—the Paleotethys and the Neotethys. By the Late Jurassic, the Paleotethyan Ocean, the great incisure into Pangea that had formed in the Devonian, was on its way out as the Neotethyan Ocean increased at its expense throughout the rest of the Mesozoic. The Paleotethys formed in the Devonian, when a stretch of European terranes (which were to become northern Europe) separated northward from the northern margin of what is now known as Africa. This opening ocean, created by the Paleotethys midoceanic spreading ridge, gave the world’s geography a huge incisure that opened to the east and partially separated Pangea into Laurasia and Gondwana for much of the remaining Paleozoic. The Neotethyan Ocean began to close toward the east as the African plate rotated counterclockwise and the northern margin of the Neotethyan Basin began undergoing subduction. This produced a number of island arcs in what is now the region of the Middle East and the Caucasus.21 In addition, subduction of the Neotethyan oceanic crust under the more southerly Arabian portion of the African continental plate formed an island chain across present-day Iran.22
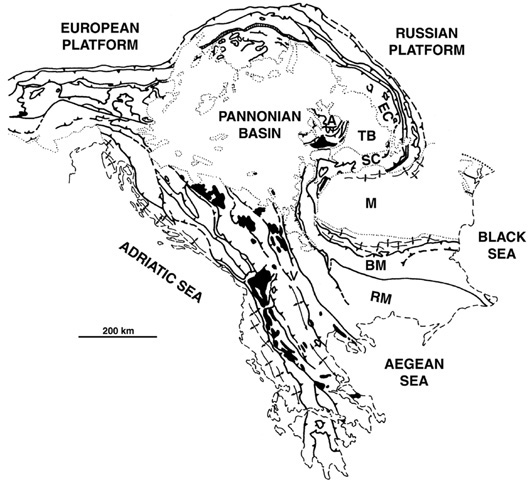
Figure 4.3. A generalized tectonic map of the Eastern European Alpine system. A: Apuseni Mountains; BM: Balkan Mountains; EC: Eastern Carpathian Mountains; M: Moesian Platform; RM: Rhodopian Mountains; SC: Southern Carpathian Mountains; TB: Transylvanian Basin. (After Burchfiel 1980)
In order to understand how these Neotethyan tectonics relate to the Transylvanian region, we now need to explore the history of what today are the Western, Eastern, and Southern Carpathian Mountains, that great mountainous loop crossing Slovakia, Ukraine, and western Romania (figure 4.3). The history of these mountain belts is very messy, to say the least. The region has seen times of incredible deformation, severe faulting, great compressional forces, crustal warping, zones of extension and collision, and the suturing of continental fragments against the irregular European-Russian platform. These events took place throughout the Cretaceous, and they were subsequently highly modified by further regional deformation during the Cenozoic.23 Research by Mircea Săndulescu (Universitatea din Bucureşti), by B. Clark Burchfiel (Yale University), and, most recently, by Ernst Willingshofer (Vrije Universiteit Amsterdam) has deciphered a good deal of the complex paleogeo-graphic history of this region, thus providing us with a view of the events that gave the Transylvanian dinosaurs a place to live.24
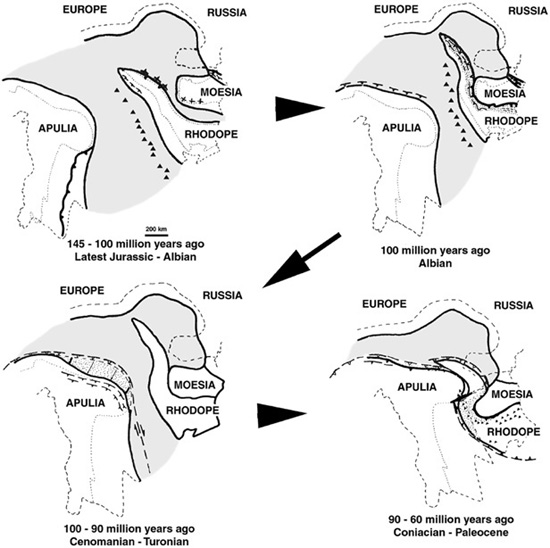
By the Late Jurassic, numerous fragments of continental crust lay immediately south of the European platform in the northern Tethyan realm (figure 4.4). Two of these fragments—Apulia and Rhodope25—are of particular importance to us. Here the margin of the European platform was very irregular, with a substantial promontory—the Moesian platform26—extending westward to form an important northern embayment between it and the European plate.27 This embayment has its origin in the opening of the Alp-Tethys oceanic basin in the Early Jurassic. At that time, Rhodope lay to the west and south of Moesia. The short-lived Severin Ocean separated Rhodope and Moesia, while the Vardar Ocean was to the south and west of Rhodope, separating it from Apulia. Both of these oceans were relatively sizeable deepwater domains, with Vardar larger than Severin. On Apulia, shallow-water carbonates dominated, but there also appear to have been regions of exposed land.28 Likewise, Rhodope consisted of a mixture of one or more exposed islands, shallow water carbonates, silts, and sands. In contrast, Moesia and adjacent parts of the European platform appear to have been covered by shallow seas.29
From the Late Jurassic onward, the kinematics of convergence of these microplates ultimately controlled by movements of the African platform relative to the European platform, has Rhodope beginning its north- and eastward migration toward Moesia, thereby closing the Severin Ocean sometime during the Aptian, about 120 million years ago. By the Early-to-Late Cretaceous transition (approximately 105–90 million years ago), the Severin Basin had closed and the Rhodope microplate began docking with Moesia, a process that continued for another 15 million years, into the Campanian. By that time, having wrapped itself around the western margin of the Moesian promontory, Rhodope was shifting to more continental conditions.
The northward and eastward movement of the Apulian fragment toward Rhodope began in earnest early in the Late Cretaceous and by the Campanian–Maastrichtian interval, some 80–70 million years ago, Apulia had begun its docking with Rhodope. As this migration of Apulia continued during the Paleocene–Miocene (65–15 million years ago), it induced the clockwise rotation of Rhodope, which had the effect of wrapping Rhodope completely around the Moesian promontory into the European embayment. Various parts of the Apulian plate thrust north-and eastward to occupy the wedge between Rhodope, Moesia, and the European platform and add to the continental conditions of the Transylvanian region. It is through all of these collisions, which began in the Cretaceous and continued through much of the Tertiary, that the Western, Eastern, and Southern Carpathian mountain belts were formed (figure 4.3). As it was shoved against the margins of Moesia, Rhodopian tectonics formed the Southern Carpathians—which includes the Haţeg Basin—and small portions of the adjacent Eastern Carpathians. The remainder of the Eastern and all of the Western Carpathians are due to the collision and suturing of Apulia with the European platform in the Tertiary. The Apuseni Mountains—the heart of Transylvania, lodged between the Southern and Eastern Carpathians—are also the tectonic result of the forward regions of Apulia pushing north- and eastward toward the European plate.
Figure 4.4. Relative positions of the Apulian, Rhodopean, and Moesian microplates from the latest Jurassic through the beginning of the Tertiary. The shaded regions represent the possible extent of the oceanic crust; triangles are suggested locations of volcanism; barbed lines indicate subduction zones; the dashed line at the top of each diagram indicates the present location of the Carpathian–Balkan front; the dashed line around the boundaries of the microplates are present-day boundaries along the Black, Aegean, and Adriatic seas; the dotted lines are the present sizes of microplate boundaries. (After Burchfiel 1980)
Not only were continents adrift and new oceanic basins forming in the Neotethyan realm during the Cretaceous, but these great crustal dynamics also had profound effects on changes in the sea level, patterns of oceanic circulation, and the climate, all of which influence our understanding of the conditions faced by the dinosaurs of Transylvania. Sea level in Europe (and for much of the rest of the globe) rose and fell throughout much of the Early and Late Cretaceous.30 The largest of the Late Cretaceous transgressions began about 100 million years ago, and it is thought to be related to an increase in rates of seafloor spreading and the addition of new oceanic ridge systems.31 It certainly reduced the entire European coastline dramatically, allowing the Neotethyan Ocean to communicate not only with the North Sea, but also, through the Polish Trough and the Paris Basin, with the North Atlantic (Plate VI). Flooding of what is now the region of Ireland and England, and much of France, Spain, and central and eastern Europe, culminated approximately 80 million years ago (during the Campanian) and produced the greatest number of European islands, both large and small. To the north and west was the largest, Fennosarmatia (present-day Fennoscandia and northern European Russia). In the extreme west and south lay several smaller islands, including the Iberian Meseta (Portugal and western Spain), the Ebro High (northeastern Spain), and the Massif Central (southcentral France). Several other exposed regions are located farther north and closer to Fennosarmatia (including the Cornwall region of southwestern England, the Irish Massif, and Caledonia). Finally, the Rhenish Massif (northeastern France and southern Germany) occupied a more central European realm.
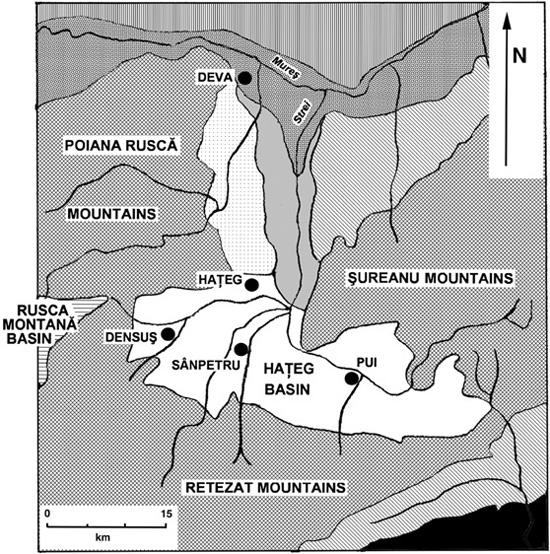
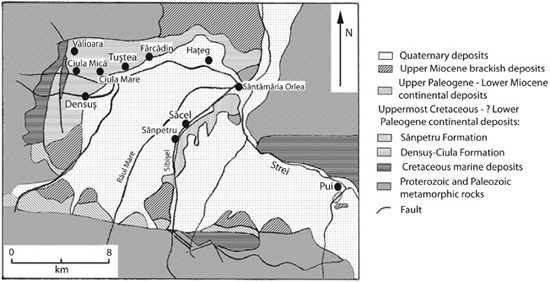
Since Nopcsa’s time, we have learned a great deal more about the paleo-geography of Transylvania during the Late Cretaceous. The Haţeg Basin is an intramontane basin measuring approximately 45 km from east to west, bounded by the Şureanu Mountains to the northeast, the Poiana-Ruscă Mountains to the northwest, and the Retezat Mountains to the south (figure 4.5). Narrow zones of crystalline basement rock and marine sediments separate the Haţeg Basin from the Petroşani and Rusca Montană intramontane basins. The Petroşani Basin to the east contains Oligocene and Lower Miocene freshwater lake and brackish water deposits, while the Rusca Montană Basin to the west contains Upper Cretaceous marine and fluviolacustrine deposits, preserving abundant coal beds and a reasonably well-known terrestrial flora. Finally, the Transylvanian Depression (also known as the Transylvanian Basin32)—a very large, subcircular basin to the north and northeast of the Haţeg Basin, within the bend zone connecting the Eastern and Southern Carpathian Mountains—hosts more than 10 km of sediment that date from the Late Cretaceous through the Pliocene.
During the latest Cretaceous, the Haţeg Basin was located on the eastern flank of the Rhodope microplate, whereas the localities of the adjacent Transylvanian Depression were scattered around the forward and northern edge of the Apulian microplate. At that time, all were located about 20°–30° North Paleolatitude.33 Surface currents in the nearby marine realm were probably westward, and surface water temperatures may have averaged no less than 24°C in the winter and 32°C in the summer.34 This part of the Neotethyan Ocean may also have been susceptible to violent storms.35 A diverse array of predatory ammonites and a host of single-celled foraminifera lived in these waters, but the marine vertebrate biota of Transylvanian is unknown. In nearby regions, however, teleost fish, sharks, marine crocodilians, and mosasaurs are present.36
The surrounding warm waters of the Neotethyan Ocean and the prevailing westerly winds most likely made the terrestrial climate of the Transylvanian region warm and humid, at least seasonally. In addition, what today are the Retezat and Poiana-Ruscă mountains were beginning to form, providing the principal topography of the Transylvanian landscape with a source of sediments during the latest Cretaceous.
Figure 4.5. Major geographic features around the Haţeg Basin. (After Weishampel et al. 1991)
The Transylvanian dinosaur fauna comes from strata exposed in numerous localities within the Haţeg Basin and the Transylvanian Depression (figure 4.6). Thus far, research on the Haţeg Basin is more extensive and detailed than that on the Transylvanian Depression, so the former will be covered first.
The Haţeg Basin includes two distinctly different sequences of rock, which, as a consequence, represent two formational units: the Sânpetru Formation outcrops in the central part of the southern end of the basin, and the Densuş-Ciula Formation in the west (Plate VIII). The Sânpetru Formation—a rock unit thought to be an outcropping at least 2,500 m thick on both sides of the Sibişel Valley, along the Râul Mare all the way to Sântămărie Orlea, and along the Râul Bărbat at Pui—has yielded the richest lode of fossil vertebrates from the Haţeg Basin.37 Sedimento-logical research indicates that the Sânpetru Formation consists almost exclusively of clastic sediments, from coarse conglomerates (including pebbles 15–20 cm in diameter—about the size of large grapefruits) to claystones.38 The Formation also includes irregular beds of calcium carbonate nodules. Toward the top of the Sânpetru Formation, strata not only include the finer-grained beds (now dark gray or green instead of brown or red39), as seen lower in the section, but also coarser and thicker conglomeratic beds. In contrast, the approximately 4,000 m thick Densuş-Ciula Formation covers a large area of the northwestern part of the Haţeg Basin.40 Its gray-colored lower portion consists of repetitive sequences of volcanic and terrestrial strata, revealing a turbulent history punctuated by volcanic eruptions, while the upper portion is dominated by red conglomerates, sandstones, and mudstones.
Figure 4.6. A geologic map of the western part of the Haţeg Basin. (After Weishampel et al. 1991)
The kinds of sediments in each of these Formations provide information about the environment in which they were deposited. Those of the Sânpetru Formation were laid down within rapidly flowing, braided river systems, probably deposited at the base of the newly exposed Retezat Mountains.41 The Densuş-Ciula Formation appears to consist of strata deposited near or along the flanks of volcanoes.42 The lower part of the Densuş-Ciula Formation is periodically interrupted by ashfall and mudflow deposits that contain volcanic debris, while the upper part lacks such beds, suggesting that this volcanic activity had decreased in the region.43 According to Dan Grigorescu, the river systems of the latest Cretaceous within the Haţeg Basin flowed predominately northward, away from their sources in the Retezat Mountains.44 The angularity of the grains and an abundance of feldspars and mica in the sandstones suggest that these sediments settled out in close proximity to their source. The conglomerates were deposited in rapidly flowing channels, while the sandstones and silty mudstones were laid down as river channels and bars, as well as overbank deposits during seasonal floods that burst the banks of the rivers and spread over the surrounding floodplain.
In the southern part of the Transylvanian Depression, the Upper Cretaceous Şard Formation is exposed along the southeastern margin of the Apuseni Mountains, between the village of Vurpăr and Pâclişa on the northern margin of the Mureş River, and near Sebeş.45 Here it consists of a 2,500 m stack of red sandstone and mudstone beds that represent continental fluviolacustrine deposits. Moderately to well-developed paleosols, often associated with abundant bioturbation, developed on top of the abundant overbank deposits. Unlike the situation in the Haţeg Basin, the rivers flowed southward, away from their sources in the southern Apuseni Mountains. The Jibou Formation along the more northerly margin of the Transylvanian Depression is as much as 1,200 m thick.46 Formed of coarse clastic rocks representing fluvial channels and red clay overbank deposits, it has only recently begun to yield much of a Late Cretaceous fossil record.
Based on studies of its Late Cretaceous vegetation, Transylvania probably had a tropical/subtropical climate, suggesting an annual mean temperature of approximately 22°C and annual precipitation of about 150 cm/year.47 Occasionally lakes and ponds occupied the landscape, while soils developed on the floodplains. Analyzed by François Therrien of the Royal Tyrrell Museum of Palaeontology (Drumheller, Alberta, Canada), these ancient soils—called paleosols—indicate that the Transylvanian climate, although generally subhumid and monsoonal, also had periods of aridity. Rainfall during these drier seasons was reduced to a range of 71 to 85 cm/year, and it lowered the mean annual temperature to 11.5°–11.7°C.48 Evidence for these dry times come from the carbonate nodules—known from the Sânpetru, Densuş-Ciula, and Şard formations—that were formed by the precipitation of calcium carbonate as the moisture in the soil evaporated. These paleosols indicate that evaporation must have exceeded rainfall during part of the year. Bioturbation, in the form of fingerlike burrows and root traces a few centimeters long and several millimeters wide, riddles the red mudstones of the overbank deposits and even some sandstones. The burrows, probably feeding traces of annelid worms and insects, indicate that the floodplain and the other sediments were undergoing active turnover.
During the wet season, Transylvania was a mosaic of greenery, revealed by the relatively rich palynofloras and mega- and mesofloras from the Haţeg and Rusca Montană basins and the Apuseni Mountains. These floras—found in the alluvial fan and the volcanoclastic, fluviolacustrine, and coal-bearing deposits—consist of a mixture of horsetails (sphenophytes), ferns (pteridophytes), and flowering plants (angiosperms).49 Dispersed conifer trees are present, while cycads, ginkgoes, and benettitaleans are rare or absent from these floras. Whether this paucity is biological (i.e., they did not live within the depositional basins) or taphonomic (i.e., they were present but their remains were not preserved) is unknown. Nevertheless, benettitaleans (cycadlike close relatives of angiosperms) and conifers, along with ferns and angiosperms, are abundantly known in the Bohemian Massif of Czechia from earlier times (Cenomanian; approximately 95 million years ago), and their presence in the latest Cretaceous of Romania, though speculative, might be anticipated.
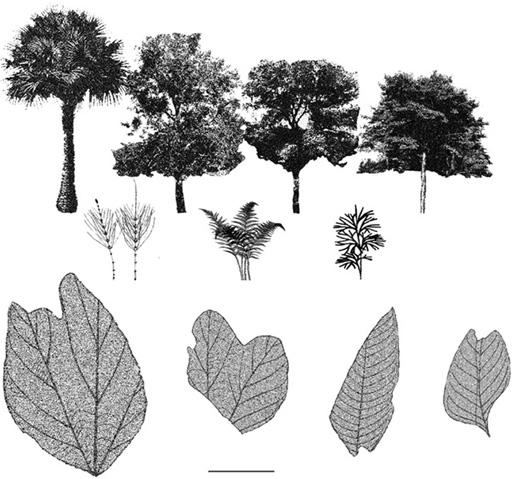
We can reasonably assume that the shady river margins and moist marshy habitats in Transylvania were colonized by stands of horsetails, some of which grew to approximately 1 m high and were towered over by tropical ferns that probably stood up to 2 m high. Away from the rivers, in patches of sunlight, horsetails and ferns flourished in the loose and gravely, seasonally wet soils. The better-drained habitats farther from the rivers were dotted with a heterogeneous mix of ground cover, shrubs, and stands of trees (figure 4.7). What the herbaceous ground cover might have resembled is unknown, but it probably consisted of small ferns, sphenophytes, and club mosses (lycophytes). Shrubbier understory vegetation was composed principally of angiosperms and low tree ferns. The canopy of the open woodlands probably extended to a height of 20–30 m and was formed of filicalean ferns (Gleicheniaceae, Polypodiaceae), broadleafed conifers (?Cheirolepidiaceae), and woody angiosperms, including palms (Arecaceae), screw pines (Pandanaceae), walnuts (Juglandaceae), and laurels (Lauraceae). However, it was the sycamores (Platanaceae) that appear to exhibit the greatest diversity and biomass of these Transylvanian forests.50
Figure 4.7. Probable elements of the Late Cretaceous flora of Transylvania. Arborescents: palm (Arecaceae), sycamore (Platanaceae), walnut (Juglandaceae), and laurel (Lauraceae) (above, from left to right). Ground cover: horsetail (Equisetaceae), fern (Polypodiaceae), and clubmoss (Lycopodiaceae) (middle, from left to right). Examples of leaves from dicot (bottom left two) and monocot (bottom right two) plants known from the Rusca Montană flora. Scale of leaves = 5 cm; the figures of the arborescents and the ground cover are not to scale. (After Petrescu and Duşa 1982)
The only invertebrates known from the Late Cretaceous of the Transylvanian region are small land snails, freshwater unionid clams, and ostracodes.51 Besides their biostratigraphic significance, these mollusks provide additional insight about the paleoecological relationships of the area. Like their present-day relatives, the Haţeg snails probably required constant moisture, food, and shelter, most likely living beneath rocks and the broadleaf litter of the forest floor. There, they crept about, each on its broad, flat, flexible foot, feeding upon detritus and each other or scavenging on the dead and dying. As for the unionids, these freshwater bivalves were burrowers, reworking the sediments on the bottoms of streams and rivers in order to feed on small food particles. As such, they formed an important part of the Haţeg freshwater ecosystem.
Other invertebrates almost certainly were present in the Late Cretaceous of Romania, but the fossil record for this part of the fauna is virtually silent. This applies especially to insects, for which the Transylvanian record is thus far absent (except for a dermestic beetle boring into a Magyarosaurus osteoderm52) and, indeed, is rather scanty throughout the remainder of Europe during this interval. This limited evidence of insect activity includes leaf damage, eggs, and body parts of butterflies, dragonflies, beetles, and caddisflies from the earliest Late Cretaceous of the Bohemian Massif (Czechia) and the Paris and Aquitanian basins of northwestern France.53 However, we do know from the records elsewhere, and from earlier times, that virtually all major groups of extant insects had evolved by the Maastrichtian. Based on this information, it’s reasonable to assume that the underbrush and leaf litter of Transylvania must have abounded with insect life. Termites, ants, and cockroaches tunneled through the detritus. Grasshoppers, crickets, and katydids hunted and chirped, while cicadas sang their whining all-day, all-night song. Beetles, bugs, mantises, and aphids held court in the trees and shrubs. In the air, bees, wasps, flies, and butterflies buzzed and flapped, while dragonflies silently hovered above it all.
Here we have presented a sketch of what the environmental setting of Transylvania might have been like some 70 million years ago. Some of the information is readily available from the rock and fossil record, but other aspects remain highly speculative. On this basis, do we now think that the paleogeography, paleoenvironment, and biota is well understood? Not yet we don’t.
As we have noted, Nopcsa and several recent paleogeographers of the Mesozoic of Europe depicted western Romania as a reasonably large island, often within an archipelago whose activity is due to movements of small tectonic plates associated with the conjoined motion of Africa and Apulia. However, things are not necessarily so straightforward; other studies indicate that this habitat reconstruction is far too simple.54 We cannot overemphasize how extremely difficult it is to reconstruct now-obliterated habitats, especially those terrestrial ephemera created and disposed of by tectonic dynamism during the evolution of the Cretaceous Neotethyan Ocean. Despite his strong belief that the Haţeg area was part of an island, Nopcsa also recognized that there was a paucity of geologic evidence to suggest that this was true—no trace could be found of beach or other tidally influenced deposits indicating the periphery of such an island. Abel also criticized Nopcsa’s reconstruction of the Haţeg region as an island, questioning whether the area that supported the fauna was actually small enough to be considered an island or, instead, was sufficiently large in this regard to be considered a mainland.
Just how do we identify islands in deep time? Unlike the present or recent past, where islands can be quite easily seen or are uncontroversial in their reconstruction, we need to be more careful with our prehistoric interpretations. Nopcsa’s strategy was to enlist both geology and biology, just about the only observations we still have available to us. From geology, we could look for sedimentological evidence of beaches or shorelines (which Nopcsa—and others since—have been unable to find). We could also look to the depositional context, in particular the thickness of the stack of terrestrial sediments, for an estimate of the size of the rapidly subsiding depositional basin. Given that the thicknesses of the terrestrial rocks observed in the Haţeg Basin and the Transylvanian Depression range from 2,500 to 10,000 m, this large accumulation of sediment means that the Haţeg region cannot be reconstructed as a small island, and it needed to have been a sufficiently large emergent area to be able to accommodate the tectonic activities that clearly occurred there. As we have seen in the earlier portions of this chapter, another source of data comes from paleogeographic reconstructions. Current paleogeo-graphic maps indicate a sizeable emergent area in this northern Peri-Tethyan realm, corresponding to the relatively small Haţeg Basin and much larger Transylvanian Depression. However, from what is now eastern Iran to Italy, the entire area was a huge swath of active mountain building (Plate VI).55 It is therefore difficult (if not impossible) to assess, amid all of this flux, many of the geographic details necessary to have made Transylvania a sufficiently sizeable and remote island.
Fortunately, the best indicators of islands seem to be biological.56 The nature of their isolation from more continental faunas will produce different effects on island faunas. First, it is highly likely that island faunas will be less diverse than those on the nearby mainlands from which they colonized. Second, these faunas are very likely to be unbalanced, with major clades known from the mainlands absent from the islands. Third, depending on how long ago the islands were colonized, their faunas may resemble the more primitive nature of earlier faunas elsewhere. Fourth, for those taxa present on islands, there is probably a high degree of endemism. Finally, for the individuals living on islands, changes in body size are common.
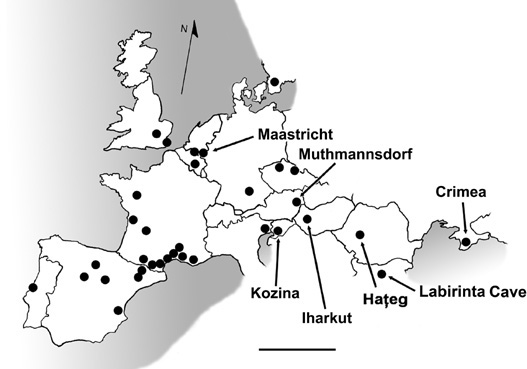
The issue of body size will be considered in some detail in chapter 5. Here, we will explore diversity, balance, primitiveness, and endemism in Transylvania, based on information about the membership of other faunas. We begin by comparing the Transylvanian fauna with other assemblages from Europe (figure 4.8). These include several sites in Sweden, England, Bulgaria, and Czechia; numerous localities in southern France, Spain, and Portugal; the assemblage from Muthmannsdorf in Austria; several nearshore marine sites around Maastricht in the Netherlands and adjacent parts of Belgium; a poorly known assemblage from Crimea in Ukraine; and new finds made in the late 1990s and early 2000s from Italy, Germany, Slovenia, and Hungary.57
Paleontologists working at sites in southern France have recovered some of the most important terrestrial and nearshore marine vertebrate faunas from the Late Cretaceous of Europe.58 From Provence in the east to the northern foothills of the Pyrenees in the west, a wealth of dinosaurs, turtles, crocodilians, pterosaurs, bony fish, frogs, lizards, and mammals have been collected since the earliest discoveries in the 1700s.59 There are theropods from southern France, quite similar to the dinosaurs of the Transylvanian fauna. Like their Transylvanian relatives, most of these French specimens are poorly understood as yet. They include small forms, as well as the 7–10 m long Tarascosaurus and some tantalizing remains of a large bird (neither of which is yet known from Transylvania). A titanosaurian sauropod is present—Ampelosaurus—as are the ornithopods Rhabdodon, some newly discovered hadrosaurid material referred to Pararhabdodon (see below), and the nodosaurid Struthiosaurus. There are also plenty of turtles, fish, and crocodilians, gigantic pterosaurs, and eggs displaying a megaloolithid architecture.60
Figure 4.8. A map of modern Europe, indicating known dinosaur-bearing localities from the Late Cretaceous. Scale = 600 km
Numerous terrestrial faunas have also been found across the Iberian Peninsula. Those from Portugal are less well known, but they include, among the dinosaurs, a variety of small theropods, titanosaurian sauropods, nodosaurids, and ornithopods, as well as crocodilians, turtles, mammals, amphibians, and fish.61 Spanish localities have yielded a variety of thus-far poorly understood small theropods, titanosaurian sauropods (including one named Lirainosaurus), the nodosaurid Struthiosaurus, at least two kinds of ornithopods (Rhabdodon and Pararhabdodon), dinosaur eggs, pterosaurs, and a rich array of crocodilians, lizards, snakes, turtles, placental mammals, amphibians, bony fish, and sharks.62
We have been introduced already to the Gosau fauna of Muthmannsdorf, Austria (chapter 2).63 The dinosaurs from this area consist of indeterminate theropods, an indeterminate rhabdodontid (formerly known as Mochlodon suessi),64 and the nodosaurid Struthiosaurus, while the rest of the fauna includes pterosaurs, crocodilians, a choristoderan (a small crocodilianlike fish-eater), lizards, and turtles.
Nearshore marine deposits from the Maastricht region of the southernmost Netherlands and adjacent Belgium have produced a rich marine fauna, but very few dinosaurs thus far.65 A hadrosaurid known as Orthomerus dolloi and a theropod called Betasuchus bredai are known only from isolated and often fragmentary bones and teeth. These remains may represent cast-off body parts from bloated carcasses that drifted out to sea, for otherwise the vertebrate fauna consists of marine crocodilians, plesiosaurs, mosasaurs, turtles, bony fish, and sharks.
Of the few remaining European faunas, the earliest known, described in 1945 by A. N. Riabinin from St. Petersburg, is from Crimea, the portion of Ukraine projecting into the Black Sea.66 Several postcranial skeletal elements belong to a hadrosaurid (originally called Orthomerus weberi, but this is probably not diagnostic). More recently, Peter Wellnhofer, a Bavarian paleontologist and one of the world’s leading specialists on Pterosauria, described a tantalizing, yet frustrating hadrosaurid femur from Maastrichtian marine limestones in Bavaria, Germany.67 At the opposite end of the scale of preservation, some excellent hadrosaurid material, including most of the skeleton and skull, occurs in Santonian deposits in northeastern Italy; it has been named Tethyshadros insularis.68 Finally, discoveries made over the past ten years in several areas in eastern and northern Europe—Kozina, Slovenia; Iharkút, Hungary; Labirinta Cave, Bulgaria; Mezholezy, Czechia; and the Belgorod and Volgorod regions of Russia—have disclosed additional dinosaur records from the Late Cretaceous. Thus far the dinosaur fauna from Slovenia, which is roughly contemporary with those of southern France, includes hadrosaurids and other ornithopods, small theropods, and two different crocodilians,69 while that from Hungary, dating from as early as late Santonian, indicates the presence of a neoceratopsian (Ajkaceratops kozmai), a nodosaurid ankylosaur (Hungarosaurus tormai), a rhabdodontid ornithopod, theropods (including an enantiornithine bird), an azhdarchid pterosaur (Bakonydraco galaczi), several kinds of crocodilians (including Iharkutosuchus makadi), turtles, amphibians, and fish.70 The Bulgarian occurrence is a disarticulated, incomplete hadrosauroid hind limb and a caudal vertebra that come from Labirinta Cave, southwest of the town of Cherven Bryag, in the northwestern part of the country.71 From Czechia comes a fragmentary yet tantalizing medium-sized femur that is thought to come from an ornithopod dinosaur. It was collected from shallow marine sediments (late Cenomanian in age) near the village of Mezholezy, between the towns of Kutná Hora and Čáslav. Currently, the Russian material from Belgorod consists of a tooth and a vertebra of what appears to be a close hadrosaurid relative, perhaps similar to Telmatosaurus, but of Albian–Cenomanian age.72 Finally, the Volgorod locality, Maastrichtian in age, has yielded two dinosaurs (an indeterminate hesperornithid and a basal theropod) and a few turtles.73
How do these European faunas compare with each other and with others globally, in terms of diversity, balance, primitiveness, and endemism? It can be argued that the Transylvanian fauna is not much different from those elsewhere in Europe at the end of the Late Cretaceous—it has approximately the same within-clade diversity of creatures as France, Spain, or Austria: one or two ornithopods, a titanosaur or two, small and rare theropods, maybe a nodosaurid, pterosaurs, eusuchian crocodylomorphs, lizards, and amphibians. From this quick census, we would be hard-pressed to see many differences in diversity among the European regions.
In this way, the European faunas appear to be reasonably ecologically balanced when compared with one another; that is, the makeup of the trophic organization of each region is roughly equivalent. For example, there are a variety of primary consumers in each fauna. Which group is at the top of the food chain (small theropods, pterosaurs, or eusuchian crocodilians) is somewhat questionable, but all are well represented from region to region. None of these faunas seems out of place with the others in terms of relative primitiveness. For all of these reasons, none of the European faunas appear to be loaded with endemics.
That said, we’ve been looking at the wrong level for our comparisons. Major clades are expected to be widely distributed, on mainlands and on islands. When looked at from a species-level perspective, however, the pattern of similarity across the region changes. Telmatosaurus transsylvanicus is known only from Romania, while Pararhabdodon isonensis comes just from Spain and France. Ampelosaurus atacis has been found only in France, while Magyarosaurus dacus hails solely from Romania. The three species of the nodosaurid Struthiosaurus (S. austriacus, S. transylvanicus, and S. languedocensis) are each found in different parts of Europe. At these lower taxonomic levels, each European fauna is completely endemic to its particular region.
Global comparisons of diversity, balance, and primitiveness are perhaps the most telling. At the global level, mainland faunas often have twice or three times (and more) the within-clade species diversity that is found in the different regions of Europe.74 Then there’s the issue of faunal balance, in particular regarding the top predator. If the European faunas of the Late Cretaceous differ in terms of the makeup at the top of the food chain, then they are not just microcosms of continental conditions elsewhere in the world. When compared globally, the most obvious difference between these European assemblages and those elsewhere in the world is who is represented in this top rank. The absence of a large theropod in Transylvania and in the other European faunas differs from the more continental situation during the Late Cretaceous. In western North America, we have Albertosaurus, Daspletosaurus, and Tyrannosaurus; central and eastern Asia has its Tarbosaurus and Alectrosaurus; and in South America there are Abelisaurus and Giganotosaurus.75 The lack of a large theropod in the European assemblages could be an artifact of collecting; none of the European localities can match the availability of fossils in the badlands and deserts of these other, larger regions. Nevertheless, if this lack is not an artifact, then the European faunas do depart from the more continental conditions of North and South America and Asia, making them unbalanced, at least in terms of who the top predator was. Further faunal absences from Europe consist of a host of major theropod groups, including tyrannosauroids, ornithomimosaurs, oviraptorosaurs, and therizinosauroideans. Another factor in this imbalance is the absence of diplodocoidean sauropods, ankylosaurids, pachycephalosaurs, and ceratopsians in Transylvania. Without many of these taxa, the European faunas tend to look rather primitive, more closely resembling those from the Early rather than the Late Cretaceous.
The Transylvanian region seems to have been an island or, at the least, it behaves like one biologically. Even so, its areal extent is nearly unconstrained. It assuredly was large enough to form basins with very thick terrestrial depositions. It was also big enough to support numerous interacting tetrapod species. Could it have been large enough or close enough to contemporary mainlands to operate as an outpost, part of a tectonically active region where sizeable terrestrial habitats were contiguous with continental landmasses, but on an off-and-on basis?76 At the moment, it’s difficult to say. And maybe it’s not really relevant to determine whether the Transylvanian region was an island or an outpost. Perhaps what is most important here is the degree of insularity or isolation of the Transylvanian region and its fauna from those elsewhere in the world.
Insularity need not be limited just to those landforms we call islands. Rather, it is used as a qualitative measure of isolation in present-day ecosystems. MacArthur and Wilson stated this in their opus on theoretical ecology and island biogeography:
Insularity is moreover a universal feature of biogeography. Many of the principles graphically displayed in the Galápagos Islands and other remote archipelagos apply in lesser or greater degree to all natural habitats. Consider, for example, the insular nature of streams, caves, gallery forest, tide pools, taiga as it breaks up into tundra, and tundra as it breaks up into taiga.77
In the case of our Transylvanian investigation, what matters is the insularity of the region, rather than the name of the landform we give to where it existed. Islands surrounded by water, remote lakes surrounded by land, clumps of woodland in a prairie, oases in the desert, mountain peaks, geologically ephemeral outposts in a tectonically active assemblage of microcontinental plates—all are characterized by being isolated from each other by a formidable barrier.78 However we reconstruct the paleogeography of Transylvania, it was certainly an insular place, by virtue of tectonics, geography, and ecology. The Transylvanian dinosaurs made their living on the edge, thanks to their insularity from the much larger continental faunas of Africa, Asia, and North America. Now it is a matter of understanding how and when this remarkable fauna may have arisen. To do so, we will take up the issues of phylogeny, heterochrony, and historical biogeography in chapters 6, 7, and 8.