DAYDREAMS OF THE LOWER PALEOLITHIC
I dream things that never were and say why not?
THE EARLIEST KNOWN EXAMPLES of external thought appear roughly 1.5 million years ago in the form of chipped stone objects. They are pieces of rock chipped on both sides into an oval shape that archaeologists refer to as bifaces, or bifacially flaked tools. They have achieved the exalted status of an externalized mental representation because they bear little resemblance to the natural object—a variously shaped cobble or fragment of rock—from which they were made. They have been transformed by a pair of human hands in accordance with a mental template, or an internal mental representation.1
Archaeologists have been unable to determine whether the bifaces were meant to “stand for something” or were an arbitrary form imposed on pieces of rock. They may have represented a large leaf of exaggerated thickness or a giant coarse seed.2 And it is possible that the biface makers imposed representations on other natural forms—carving pieces of wood or drawing patterns in sand, for example—that have not been preserved (some rare examples of bifaces chipped in bone have turned up).3 In any case, humans continued to make bifaces for more than a million years before they began to create more complex representations outside their heads. Many of the later bifaces exhibit a more refined appearance, but their essential form remains unchanged.
The appearance of the bifaces was a major event in the prehistory of the mind, signifying the emergence of what I have termed the proto-mind. From that moment forward, humans began to match at least some of the thoughts generated in their heads with Those constructed outside the brain. Mental representations were no longer entirely confined to the brain. Equally important, it seems, was the fact that mental representations—not simply emotional states or simple bits of information—could be communicated from one individual brain to another. Humans now found themselves in a landscape populated not only by physical features and organic beings, but also by thoughts in an incredibly literal sense.
The Evolutionary Biology of the Mind
[M]an bears in his bodily structure clear traces of descent from some lower form; but it may be urged that, as man differs so greatly in his mental power from all other animals, there must be some error in this conclusion.
One of the more intriguing implications of the archaeological record is that the mind or something very much like it could have emerged in another form of life. There is nothing magical about humans; it is simply that the evolutionary biology of humankind created certain conditions from which the emergence of the mind was possible and perhaps even inevitable. What conditions would be necessary to produce a true mind, a mass of structured information that exists both inside and outside an integrated group of organically evolved individual brains that continually feed it with newly generated structures of information?
An organism must possess a large brain capable of constructing complex representations from incoming sensory data. Many vertebrates meet this criterion, and some of them have brains that exceed the volume of the human brain either in absolute terms or in relation to their overall body size.4 And while a brain might construct internal representations of the outside world from a sense of smell or sound, representations based on electromagnetic radiation within the spectrum of visible light would seem to offer the greatest potential for complex and intricate maps of the external world.5 Vision provides a potentially immense range—for all practical purposes unlimited, when humans look at distant stars and galaxies. Finally, there must be a means to project mental structures outside the brain in order to establish a feedback relationship between internal and external representations. In humans, the hands and later the vocal tract became the means, but any number of features might have evolved an analogous function in other organisms, such as a tail or antennae.
Humans evolved within the primates, and both comparative anatomy and the fossil record indicate that the aforementioned conditions have deep roots in this mammalian order. As early as 50 to 40 million years ago, the primates exhibited three important trends that provided a basis for the subsequent evolution of monkeys, apes, and humans: (1) increased size of the brain relative to the body, (2) increased emphasis on vision and corresponding reduction in sense of smell, and (3) increased use of extremities for manipulation.6 All were essential for the emergence of the human mind.
The basic function of a brain is to enable an organism to respond to spatial and/or temporal changes in its environment.7 A brain allows the organism to detect a drop in temperature, a potential food item, the approach of a predator, and other changes important to its survival and reproduction, and to generate an appropriate response. Among all but the most primitive metazoans (that is, sponges), the function of the brain is performed by specialized cells called neurons; among most multicellular animals, neurons are organized into a central nervous system. But even the simplest protozoa that appeared more than 3 billion years ago—the prokaryotes, or unicellular organisms that lack a nucleus—contain structures that evolved to perform the same basic function. The notorious and well-studied bacterium E. coli has protein molecules embedded in its cell wall that act as receptors to detect specific chemicals outside itself. These receptors trigger responses that vary according to whether the chemical detected is a food or a toxin. Propelled by six flagella, E. coli can maintain or change direction in response to the information received and transmitted by its receptors.8
When the human mind is placed into this universal functional perspective on the nervous system and brain, its unique capabilities become less curious. Although the ability of the mind to create alternative realities seems rather exotic in the context of metazoan brain evolution, it still serves the same basic purpose. Modern humans not only sense changes in the environment, but imagine and predict future changes. This feat is achieved by generating novel and often complex representations of the environment, either inside or outside the brain and often collectively among individuals. Cognitive psychologists often refer to it as “mental time travel.”9 In fact, the effort to predict animal movements, rains and floods, solar and lunar eclipses, and other natural events has been a major theme in human history.
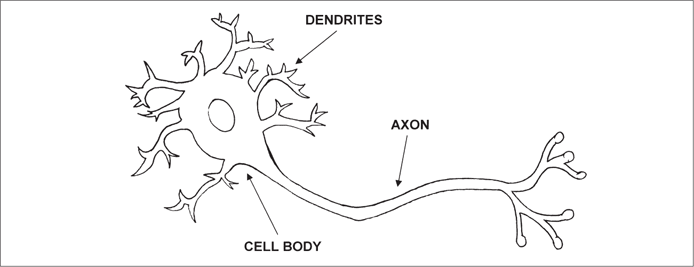
Neurons and the nervous system evolved more than 500 million years ago among the ancient phylum Cnidaria, which includes jellyfish, hydra, sea anemones, and corals.10 The design of nerve cells has not changed fundamentally since the appearance of the Cnidarians, only the ways in which they are organized. Neurons are true cells (eukaryotes) with a nucleus, but they also possess a long string-like structure called an axon and fine branching structures called dendrites (figure 2.1). Because they are dynamically polarized, neurons can transmit electrical signals to each other across synapses, the point at which they connect. They can transmit signals in digital (on or off) or analogical (continuous scale) mode. Networked together by synaptic connections, neurons can store, as well as transmit, information about the environment.
Just as there is continuity between the “brain” functions of the simplest organisms and the often wild imaginings of the human mind, responding to changes in the external world, there is continuity between the transmission and storage of information in the neurons of the early metazoans and human language. As the linguist Derek Bickerton observed, both are representational systems.11 Even the simple nervous systems of the Cnidarians contain information about the environment outside the organisms. As new metazoan groups evolved, they developed more complex neuronal networks to process and store more detailed information—more detailed and accurate maps—of the external world. They also evolved new organs that expanded the amount and type of information received—the eye is especially important to the evolution of humans—and those that expanded the range of responses to the environment from the organism.
Figure 2.1 The fundamental design of the specialized nerve cell, or neuron, has not changed for 500 million years.
The evolution of the neuron facilitated the great explosion of complex animal life that took place in the early Cambrian period roughly 540 to 520 million years ago. New forms included the worms, which exhibit a bilateral body plan (unlike the radial Cnidarians), with a defined head versus tail as well as dorsal and ventral sides. The first central nervous systems, comprising a brain in the head region and a nerve cord extending to the tail, developed in these phyla (Platyhelminthes and Annelida). The pattern of dorsal brain and ventral nerve cord was repeated in other phyla that emerged during the Cambrian explosion, including mollusks, crustaceans, and insects.12
Among these new phyla were the chordates (Chordata), which are distinguished by the embryonic development of a notochord. The first chordates were invertebrate marine organisms, but they subsequently gave rise to marine and terrestrial vertebrates: reptiles, dinosaurs, birds, and mammals with large complex brains. The living Amphioxus is thought to be representative of the earliest chordates. It possesses structures that correspond to several major components of the vertebrate forebrain, including the frontal eyes and hypothalamus, as well as the hindbrain; both the nerve cord and the brain are dorsal. The frontal “eye spot” in Amphioxus contains pigmented cells that seem to anticipate the vertebrate eye.13
Like the mind, the evolutionary process is creative. Although its products are the result of natural selection acting on randomly generated variation, novel forms are often characterized as “innovations.”14 The evolution of the early vertebrates, which began about 470 million years ago, following the end of the Cambrian, entailed a number of such innovations critical to the development of large brains. One of them was the neural crest, which eventually gave rise to the cranium, jaws, and teeth. Another was the protein hemoglobin, which plays an important role in meeting the high oxygen needs of the brain. Yet another was myelin, sheets of protein and fat molecules that insulate the axon and improve its conductivity. Insulated neurons can be packed into a small area to form a dense network.15
The early vertebrates evolved frontal eyes and nostrils, as well as several related brain components: the telencephalon and midbrain (mesencephalon) and the cerebellum (figure 2.2). The telencephalon contains expanded olfactory receptors, reflecting heavy reliance on the sense of smell. Of great significance for later brain evolution was the emergence of the midbrain, with optic tectum for receiving visual information and integrating data from other senses. The cerebellum also plays a role in vision by stabilizing visual representation as the head moves.16 Successful early vertebrates included the sharks, whose comparatively large brains probably are related to the substantial information needs of a large predator. The first terrestrial vertebrates (amphibians) emerged from the sea about 350 million years ago, and reptiles evolved roughly 300 million years ago.17
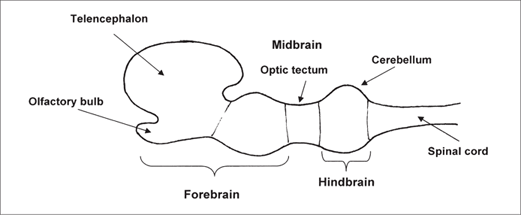
Figure 2.2 The basic vertebrate brain evolved more than 400 million years ago. A significant development was the appearance of the midbrain, with the optic tectum for receiving visual sensory input and integrating information from other senses.
Disaster struck 50 million years later, at the end of the Paleozoic era, with the most massive wave of extinctions in Earth history, probably a result of cooler climates. Ninety percent of the known fossil taxa vanished. However, the ecological vacuum opened opportunities for the evolution and proliferation of novel life-forms. On land, much of this vacuum was filled by warm-blooded vertebrates, including dinosaurs, birds, and—by 220 million years ago—mammals. The enormous energy demands of maintaining a constant body temperature seem to have been a principal factor in the evolution of bigger brains. Warm-blooded vertebrates need a large amount of accurate and detailed information about their environment.18
Among the mammals, this need was addressed in part by an evolutionary innovation in brain structure that was critical to the subsequent emergence of the mind. The neocortex—a uniquely mammalian feature—is a six-layer, sheet-like structure that covers the roof of the forebrain. Each layer is connected to other parts of the brain. The neocortex receives, integrates, and stores information about the environment from the various senses. Another innovation was a chain of ossicles that conducts sound from the eardrum to the inner ear, allowing mammals to hear high frequencies.19
The brain had acquired a new function especially evident in mammals. It was no longer simply identifying changes in the environment and coordinating appropriate responses to them. The information acquired through the senses and stored in the neuronal networks had become an essential part of how organisms survived and reproduced; learning and memory were now playing a role analogous to those of the genetically controlled (and naturally selected) structures of the body. Unlike fish and reptiles, mammals (as well as birds) require extensive parental care during the early phases of life, which reflects the increased importance of information.
The Visual Animal
Most primate-typical features involve the visual system.
The primates are a group of placental mammals that evolved from insectivores toward the close of the Mesozoic era, about 75 million years ago, and apparently occupied arboreal niches in what is now North America and Europe. They diversified after another massive wave of extinctions wiped out the dinosaurs in the late Cretaceous period. The early primates had small brains, long snouts, and laterally placed eyes, but forms resembling the living prosimians—tarsiers, lemurs, and lorises—were present by roughly 50 million years ago (Eocene epoch). They possessed larger brains with smaller olfactory bulbs (indicating reduced reliance on smell), more-forward-facing eyes, and long digits with nails instead of claws. These trends continued among the early catarrhines—a group that includes Old World monkeys and apes—during the Oligocene epoch, which began about 35 million years ago.20
Evolutionary innovations in brain structure, especially related to vision, in the Eocene primates took place among the catarrhines and were critical to the emergence of the mind. The neocortex expanded in size and in proportion to the brain as a whole. In the small prosimian Galago demidovii, the neocortex comprises 46 percent of brain volume; among living chimpanzees, the neocortex occupies no less than 76 percent of the brain.21 This surely reflected integration and storage of more information about the environment, but perhaps specifically more information about the social environment. Among primates, the ratio of the neocortex to total brain volume correlates significantly with the size of the social group.22
The primates evolved a complex of interrelated changes pertaining to vision, visual representation, and the visual coordination of muscle activity. The density of photoreceptors in the retina increased. The visual cortex of the brain expanded significantly. The optic tectum of the midbrain was restructured in relation to the overlapping fields of vision created by front-facing eyes. It functioned in a more specialized manner to allow primate eyes to fixate on objects of interest. In the forebrain, new components of the visual system improved the perception of motion and of the form of objects. Perhaps most important of all with reference to the subsequent evolution of the human hand was the development of a cortical area, unknown in other mammals, for the visual coordination of motor tasks.23
The catarrhines “reinvented” color vision. Most mammals possess limited color vision based on two types of cone photoreceptors. One of these cone types is sensitive to shorter wavelengths of light (blue portion of spectrum), while the other is sensitive to longer wavelengths (red portion of spectrum). As a result of a gene duplication, the ancestor of living catarrhines evolved a third cone type, which had been present among earlier mammals. Human vision is catarrhine vision.24
More than any other feature or complex of features, it seems, the visual system distinguishes the primates from other mammals. Why did it evolve so significantly in this group? The forward-facing eyes and overlapping fields of vision have been interpreted as part of an arboreal adaptation.25 They also have been ascribed to predation on small, fast-moving vertebrates and insects.26 Perhaps both explanations are valid. The (re)appearance of color vision among the catarrhines has been tied to their reliance on fruit and the need to identify it against a backdrop of green foliage.27 The rather generalized adaptations of the higher primates may have placed an added premium on a visual system that provides so much information about the environment.28 But another contributing factor seems likely to have been the growing importance of visual cues (and corresponding decline of chemical signals) in primate society, especially facial expressions.29
A theory of human vision was articulated by David Marr in 1982 and remains an important contribution to neuroscience.30 Marr placed primary emphasis on vision as information processing and on the creation of visual representations in the brain.31 He characterized the process as hierarchical and recursive, and the psychologist Michael Corballis later noted the parallels between Marr’s theory of vision and language—the generation of linguistic structures.32 According to Marr, visual information is processed through a sequence of stages that begins with the focusing of incoming light on the retina, the light-sensitive cells at the back of the eyeball (figure 2.3). The retinal image is received in the form of a raw primal sketch, comprising a map of variations in light intensity; these variations are hierarchically grouped into larger units to form a full primal sketch. The next stage entails processing information about the shape, orientation, and distance of objects; Marr referred to it as the 2½-D sketch.33 In the final stage, or 3-D model representation, the shapes and spatial arrangement of objects are organized hierarchically in an “object-centered coordinate frame.”34
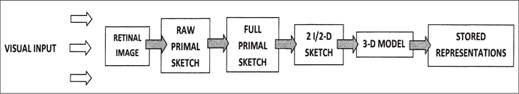
Figure 2.3 A theory of human vision developed by David Marr in 1982 stresses information processing and the recursive construction of mental visual representations, which seems to have been the foundation of the mind. (Modified from Michael C. Corballis, The Lopsided Ape: Evolution of the Generative Mind [New York: Oxford University Press, 1991], 220, fig. 9.1, based on David Marr, Vision: A Computational Investigation into the Human Representation and Processing of Visual Information [San Francisco: Freeman, 1982])
Molecular differences between living apes and Old World monkeys suggest divergence around 30 million years ago, although the earliest known ape fossils are dated to the beginning of the Miocene epoch (22 million years ago).35 Bones and teeth assigned to the genus Proconsul from Africa dominate the fossil record of early apes. Their analysis reveals that Proconsul was a mixture of ape and monkey characters—an arboreal quadruped as large as a modern female gorilla and lacking a tail. The skull is relatively ape-like in appearance, with fully forward-facing orbits and a shortened snout. Brain volume is calculated at just below 170 cubic centimeters, which is high for apes in relation to the small body size.36
During the middle Miocene (between roughly 18 and 12 million years ago), the apes underwent a dramatic expansion of their range, spreading out of Africa and across western and southern Eurasia. This was facilitated by tectonics as the African plate collided with the Eurasian plate, creating a new land connection between the continents. The apes diversified into many new genera. Among the European taxa was Dryopithecus—first discovered in France in the 1850s—which exhibits more features in common with the living African apes than does Proconsul.37 It is a plausible ancestor for the living African apes and humans and presumably possessed a brain and visual system fundamentally similar to that of both.
The Somesthetic Animal: Bipedalism and the Human Hand
[I]n the human race, a perfect tactual apparatus subserves the highest processes of the intellect … the most far-reaching cognitions, and inferences the most remote from perception have their roots in the definitely-combined impression which the human hands can receive.
When Carolus Linnaeus published his systematic classification of plants and animals in 1753, he placed humans and the great apes in the same genus. Humans were classified as Homo sapiens, and chimpanzees and orangutans (gorillas were unknown to him) as Homo troglodytes. The decision to assign apes and humans to the same genus was based entirely on comparative anatomy and the already well-known morphological similarities between apes and humans. It did not reflect evolutionary relationships and was not a shocking idea.38
In the latter half of the nineteenth century, Darwin, Huxley, and others made the case for an evolutionary relationship—a common lineage—between apes and humans. This was a shocking idea for many people. In any case, the apes had been moved to another genus or genera and even into a separate family. In the twentieth century, as more of the human fossil record was recovered, the evolutionary divergence between apes and humans was eventually placed in the middle of the Miocene epoch, roughly 15 million years ago.
By the mid-1960s, however, studies of the molecular differences between living apes and humans suggested a much later split between the two. The fossils were subsequently reinterpreted and fell into line with the results of the molecular research. In more recent years, new molecular studies have yielded further evidence of the close relationship between humans and the African apes, placing both in a group apart from the Asian apes, or orangutans. (Humans have lost their classificatory status as a separate family.) And within the evolutionary clade that contains African apes and humans, chimpanzees and humans compose an even tighter group. This classification is based on both anatomical and genetic comparisons.39 We have almost returned to where Linnaeus was in 1753.
The close relationship between chimpanzees and humans also is manifest in some aspects of behavior and ecology. In contrast to gorillas, chimpanzees forage over relatively open landscapes, engage in some hunting, and make a few simple tools. Their foraging adaptations and toolmaking, when combined with their anatomical and genetic overlap with humans, suggest that they probably resemble in many ways the last common ancestor of apes and humans. Despite this, both chimpanzees and gorillas evolved some features that probably were not present in this (still unknown) common ancestor. One of them is the unique mode of quadrupedal locomotion, knuckle walking, practiced by the African apes.40
There are inevitable gaps in the fossil record. Darwin attributed them to sampling and preservation bias—that is, “the imperfection of the geological record.”41 The problem may be further exacerbated by intervals of very rapid evolutionary change, or “punctuated equilibria.”42 There is nevertheless at least a broad evolutionary continuum from insectivores through prosimians and monkeys to apes and humans with respect to anatomy, genetics, and ecology. There are no massive gaps, for example, between Eocene prosimians and Miocene apes. But an immense gulf seems to lie between the cognitive world of the chimpanzee and the human mind—a difference that appears comparable to, if not greater than, that between prosimians and apes or, for that matter, between early chordates and placental mammals.43
How and why did the human mind emerge from the Miocene ape brain? How was the immense gulf crossed? Intermediate forms in the human fossil record fill much of the gap with respect to anatomy (and the genetic differences, as already noted, are not substantial). They include various fossil forms of Homo that reflect a continuum with respect to brain volume, which is undoubtedly a critical factor in the emergence of the mind. Equally important, there appear to be intermediate forms with respect to the mind, although they are more difficult to identify and interpret in the fossil record.
The short answer to the question is that early humans evolved a uniquely specialized extremity in the form of the hand, most likely an indirect consequence of bipedalism. The large neocortex and visual-motor system that had evolved over millions of years of primate evolution provided an essential context for this development. Humans eventually began to produce artifacts more sophisticated than those made by chimpanzees from which emerged the first externalized mental representations. They were relatively simple and exhibited little creativity, but they seem to have initiated a process—driven, I would suggest, by feedback between internal and external thought—that resulted in larger brains and more complex and creative representations. The social setting, the exchange of representations among members of the group, also must have been a factor in the process. The modern mind emerged when the ability to generate complex representations was expanded from hand to vocal tract in the form of syntactic language, creating the super-brain and unlimited creative potential.
Bipedal locomotion, or walking upright on two limbs, was the starting point, although for most of the time span that humans have walked the Earth, it had no significant consequences for brain volume. Bipedalism is equated with the evolutionary split between humans and African apes; it made the former out of the latter, and early humans have sometimes been described as bipedal apes. The early fossil record of humankind is very sparse, however, and the possibility cannot be excluded that the human lineage diverged from a chimpanzee-like ancestor for another reason and that bipedal locomotion came later. And it is also possible that bipedalism evolved among one or more extinct African apes that were not ancestral to humans. It is nevertheless the key feature that paleoanthropologists seek to identify among the few fossil remains of suspected human affiliation that are more than 3 to 4 million years old.
Bipedalism is an extremely rare form of locomotion among mammals, although like other features peculiar to humans it has roots in primate evolution—the upright posture found among apes, monkeys, and some prosimians. Most mammals are quadrupeds, and there are disadvantages attached to walking on the hind limbs. Around the world, humans struggle with the consequences in the form of knee-joint problems, back injuries, and related health issues. The advantages of bipedalism appear to be limited. A frequently cited study by Peter Rodman and Henry McHenry found that while it is less energy efficient than quadrupedalism at high speeds, bipedal locomotion is more efficient at slower (that is, walking) speeds.44 Other possible advantages include reduced body-surface area exposed to the sun, improved ability to see predators in tall-grass habitats, and better capacity to carry objects.45
How and why did human bipedalism evolve? Although most paleoanthropologists believe that it had profound consequences for humankind—freeing the hands for improved toolmaking and all that followed—they suspect that it evolved for more prosaic reasons related to the immediate foraging challenges faced by Miocene apes. A widely held view, buttressed by Rodman and McHenry’s study, is that bipedalism provided an energy-efficient means of foraging across landscapes with a low or medium density of trees. Climates became cooler and drier toward the end of the Miocene (roughly 8 million years ago), and many of the ape genera went extinct. In sub-Saharan Africa, forests shrank while grassland and open woodland expanded. The earliest humans may have evolved from Dryopithecus or a closely related African ape that spent an increasing amount of time foraging on the ground.46
The shift to bipedal locomotion affected many parts of the skeleton, not simply the limbs. In comparison with that in the African apes, the foramen magnum (through which the brainstem extends at the base of the skull to connect with the spinal cord) rotated forward, as the skull came to sit on top of the vertebral column, which, in turn, acquired an S-shaped curve. The pelvis became broader and shorter. The angle of the femur (thigh bone) was significantly altered. The lower limbs were elongated and the joint-surface areas expanded. The foot acquired a platform shape to support the upright body (figure 2.4). The changes in the bones were accompanied by changes in the musculature.47
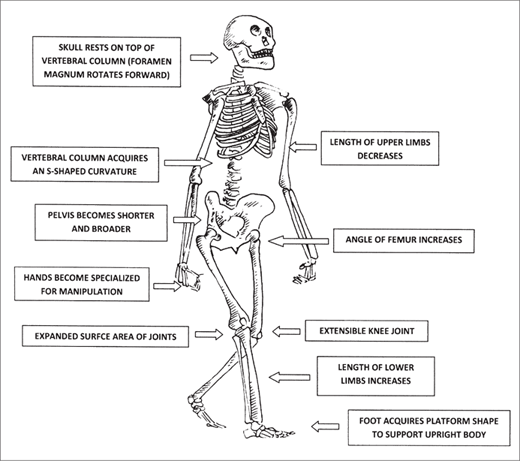
Figure 2.4 Bipedalism altered many elements of human anatomy and consequently has high visibility in the fossil record. The skeleton represents a reconstructed Australopithecus afarensis. (Redrawn from Richard G. Klein, The Human Career: Human Biological and Cultural Origins, 3rd ed. [Chicago: University of Chicago Press, 2009], 69, fig. 3.3)
The many changes in the skeleton mandated by the shift from quadrupedal to bipedal locomotion underscore the complexity of the transition, which probably took place over an extended period and, initially, perhaps was related to feeding posture.48 The myriad anatomical changes generated by the shift to bipedalism also yields a dividend for paleoanthropology—high visibility in a sparse and fragmentary fossil record. There are many potential signs of bipedalism in bones of suspected human affiliation dating to more than 3 to 4 million years ago, and paleoanthropologists search among them for these indicators.
The oldest potential signs of bipedalism presently lie in fossils dating to roughly 6 to 7 million years ago from west-central Africa assigned to the newly created taxon Sahelanthropus tchadensis (figure 2.5). The fossils include a nearly complete cranium, lower jaw fragments, and teeth. The shape of the cranium had been distorted by geologic processes, but was restored visually to its original form by means of software technology. The short face, small canine teeth, and thick enamel on the teeth are human (as opposed to modern ape) characteristics. Most important, the position of the foramen magnum may reflect bipedal locomotion.49
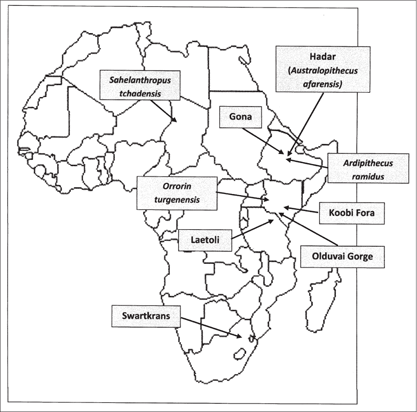
Figure 2.5 Map of fossil localities and archaeological sites in Africa, mentioned in the text, that date to between 7 and 2 million years ago.
The next oldest possibility dates to about 6 million years ago from northern Kenya. Fossils assigned to another new taxon, Orrorin tugenensis, comprise several limb bone fragments and teeth. In this case, the focus has been on the morphology of the femur, which exhibits a human-like angle an and elongated neck.50 Computerized tomography scans may indicate a human-like distribution of cortical bone thickness, but this is controversial.51 More recent fossils from Ethiopia, dating to between 5.7 and 4.4 million years ago, are classified as Ardipithecus. The youngest of these, assigned to Ardipithecus ramidus, include postcranial remains that reflect a primitive form of bipedalism.52
More human-like locomotion is found in the earliest fossils assigned to Australopithecus. Among the postcranial bones from two sites in Kenya dating to 4.2 to 3.9 million years ago and assigned to Australopithecus anamensis are a humerus (upper arm bone) and a tibia (lower leg bone) of a bipedal human.53 Especially powerful evidence of upright walking comes from Laetoli (Tanzania), where footprints—presumably made by australopithecines—were preserved in volcanic ash deposits dating to about 3.6 million years ago.54
Between 4 and 2 million years ago, australopithecines dominate the human fossil record. These early humans diversified into several species that occupied various ecological niches in sub-Saharan Africa, and one of them apparently was the direct ancestor of the genus Homo and modern humans. The early australopithecines are best known from the “Lucy” skeleton, discovered at Hadar (Ethiopia) by Donald Johanson in 1974 and eventually classified as Australopithecus afarensis. Lucy is roughly 3.2 million years old and represents a small ape-like human that walked upright but retained some striking arboreal adaptations: long chimpanzee-like fore-arms, a grasping foot, and curved fingers. Lucy evidently spent much of her time in trees. Her brain remained small (400 to 500 cubic centimeters) and only slightly larger, relative to her body size, than that of a modern chimpanzee.55
The most important developments in human evolution during this period concern changes in the anatomy and function of the hand. Until the appearance of the earliest Homo roughly 2.5 million years ago, there were no significant increases in brain size above that of the African apes, and there are no known stone artifacts. But changes in the morphology of the hand reflect a protracted transformation from a Miocene ape to a modern human hand, which was apparently completed by about 1.5 million years ago. This development—as critical as any to the emergence of the mind—was a logical consequence of bipedalism, which freed up the forelimbs for novel functions.
The evolved human hand is a remarkably sensitive, versatile, and precise instrument for translating internal thought to the external world.56 As Vernon Mountcastle observed in The Sensory Hand: “Movements and positions of the hand … are produced by the actions of 35 muscles (or 44 if one takes into account the three muscles that each have four insertion tendons) that control the movements of the 27 bones of the hand at their joints.”57 The movements of the hand are coordinated in the neocortex with the equally remarkable system of color vision that evolved among the catarrhines. And the hand functions not only as an instrument for effecting unusually sophisticated and subtle motor responses directed by the brain, but also as a sensory organ that gathers a special category of information from the external world. In this capacity, it is supported by a dense complex of nerve fibers in the glabrous (hairless) skin of the palm and fingers.58 Among humans, the hand acquired sensory functions that seem to parallel those of catarrhine vision, providing three-dimensional hierarchically structured representations of the external world through the tactile sense. Humans have become a somesthetic, as well as a visual, animal.59
The human hand is anatomically unique. As a consequence of bipedalism, the human hand is fully specialized for manipulation. The anatomist John Russell Napier (1917–1987) noted that the hand plays a locomotor role only in early childhood, when it is used like a monkey “foot-hand” for quadrupedal movement. Napier emphasized the primary importance of finger–thumb opposition, which he considered “probably the single most crucial adaptation in our evolutionary history”: “Opposition is a movement by which the pulp surface of the thumb is placed squarely in contact—or diametrically opposite to—the terminal pads of one or all of the remaining digits.”60 It is made possible by rotation of the thumb and facilitated by a thumb that is long in relation to the other digits. Among primates, humans have evolved the highest ratio of thumb length to index finger length.61
Napier subdivided human hand movements into nonprehensile and prehensile. Nonprehensile movements include “pushing, lifting, tapping, and punching movements of the fingers, such as typewriting or working the stops of a musical instrument.”62 Prehensile movements, such as gripping or pinching action between the fingers and thumb, include two primary and two subsidiary grips. The primary grips are the precision grip between the terminal pad of the thumb and the tips of one or more fingers, and the power grip between the fingers and the palm with the thumb providing reinforcement. The subsidiary grips are the hook grip, with the terminal joints bent acutely, and the scissor grip for grasping objects between the fingers (figure 2.6). The specialized skin of the hand, with its papillary ridges and sweat glands, also plays an important role in the grasping and manipulation of objects.63
While the range of hand movements is expanded by movements of the wrist, forearm, elbow joint, and shoulder joint, and even other parts of the skeleton, the basic movements, as Napier noted, are few.64 But the variety of combinations of movements and the variety of material structures that may be generated by hand movements coordinated by the modern human brain are potentially unlimited. The range of basic movements is analogous to the range of sounds that humans can produce with the vocal tract; it is a set of building blocks for a potentially infinite array of hierarchically organized technological creations. And through gesture, drawing, and writing, the role of the hands has been expanded to one similar to that of the vocal tract—the externalization of symbolically coded representations.
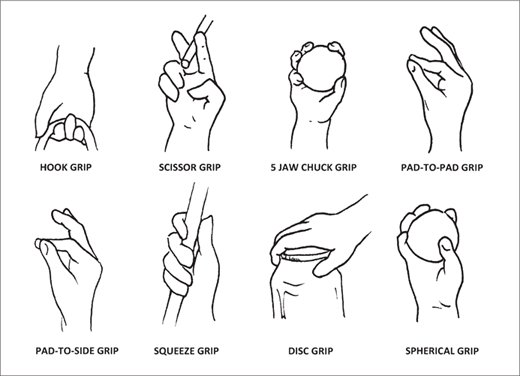
Figure 2.6 The basic grips of the human hand as defined by the anatomist John Russell Napier. (Redrawn by Ian T. Hoffecker, from Leslie Aiello and Christopher Dean, An Introduction to Human Evolutionary Anatomy [London: Academic Press, 1990], 372, fig. 18.1)
Like the other unique elements of human anatomy, the hand has deep evolutionary roots in the primates. The living primates are among the few groups of mammals that carry food to their mouths (as opposed to bringing their mouths to food),65 and as early as the Eocene, the fossil ancestors of modern prosimians had evolved grasping hands with nails instead of claws. Finger–thumb opposition appeared in the catarrhines, which eventually evolved a variety of specialized hand adaptations—all but one of which combines locomotor and manipulative functions.66
Fossil bones of the early Miocene African ape Proconsul reveal a hand that, like other aspects of its anatomy, combines typical ape and monkey features (figure 2.7).67 By the later Miocene (roughly 9.5 million years ago), a generic ape pattern is evident in hand bones of Dryopithecus from western Eurasia.68 The African apes subsequently evolved their own specialized hand, with shortened thumb and elongated fingers (for suspending from branches) and other features related to their peculiar knuckle-walking locomotion.69 The later fossil record of African apes is extremely sparse, however, and the sequence of events that yielded the modern ape hand is unknown.
The early human fossil record also is sparse and fragmentary, and there are only isolated and rather ambiguous specimens (no hand bones) dating to the interval between 8 and 4.5 million years ago. The 4.4-million-year-old hand and wrist bones of Ardipithecus ramidus, only recently unveiled in print, exhibit some novel features related to arboreal locomotion, but little indication of later trends.70 The earliest indication of subsequent changes appears 3.2 million years ago in East Africa with Australopithecus afarensis, represented by Lucy. Among the remains recovered from Hadar (Ethiopia) are more than fifty hand bones, including carpals, metacarpals, and phalanges (fingers).71 Analysis of these bones revealed many similarities to those of the living apes, including slender apical tufts (fingertips) on the ends of the phalanges. It also revealed some features lacking in apes but found among later humans, most notably a relatively long thumb (50 percent of the length of the middle digit). Lucy probably was capable of some, but not all, of the grips and other hand movements performed by modern humans.72
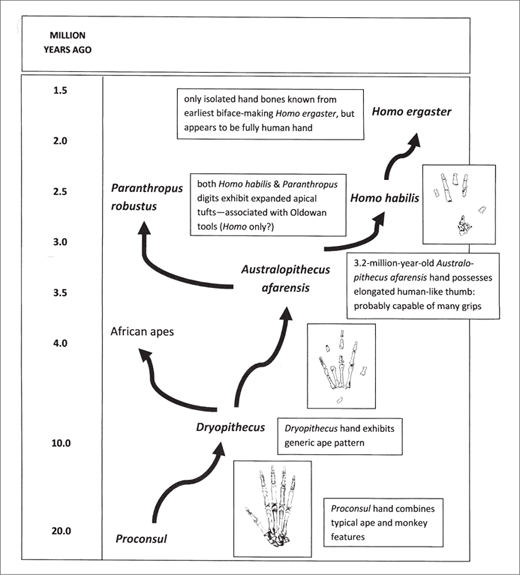
Figure 2.7 The evolution of the human hand.
The changes in hand morphology and function that took place 3.2 million years ago were not accompanied by an increase in brain volume or evidence of stone-tool production. But after 2.6 million years ago, there were increases in brain size (in both earliest Homo and the robust australopithecine Paranthropus), and stone tools are found in East Africa. Hand bones of early Homo associated with stone tools and dating to at least 1.75 million years ago were recovered in 1960 by Louis and Mary Leakey at Olduvai Gorge (Tanzania). The bones were described by Napier himself, who noted the expanded apical tufts (broad fingertips), similar to those of modern human fingers. But the Olduvai hand retains some ape-like features (for example, the morphology of the proximal and middle phalanges), and Napier wondered if it had been capable of a full precision grip.73
Of comparable age to the Olduvai finds are hand bones from Swartkrans (South Africa), also associated with stone tools and tentatively assigned to Paranthropus robustus. These hand bones differ from the earlier australopithecine pattern observed at Hadar and exhibit a number of human-like features, including expanded apical tufts like those of the Olduvai specimen.74 It is not certain, however, that these hand bones belong to Paranthropus, since some remains of Homo also are present in the deposits.75
The fully modern human hand seems to have evolved by roughly 1.5 million years ago in Homo (both Homo ergaster and Homo erectus), although the supporting fossil evidence remains somewhat elusive. A few hand bones were recovered with a partial skeleton of Homo ergaster from Nariokotome (Kenya), which is dated to about 1.6 million years ago.76 While these isolated specimens provide a less complete portrait of the hand than do the bones from Hadar and Olduvai, they are essentially modern in size and shape (like the arm bones also found at Nariokotome) and probably reflect the arrival of the fully modern human hand.
From Pebble Tools to Bifaces:
The Emergence of External Thought
Acheulean hand axes and other bifaces are a logical development from preceding Oldowan bifacial choppers.
The turning point for humans took place between 2.6 and 1.6 million years ago in sub-Saharan Africa. It began with the earliest known stone tools (pebble tools and flakes) and ended with the emergence of externalized mental representations in the form of ovate bifacial artifacts, or hand axes. The first bifaces were simple and crude, barely distinguishable from some of the more heavily flaked cobbles and fragments of the preceding era. But within a few hundred thousand years, they became more refined; over the next million years, some hand axes were shaped with admirable precision (especially when the quality of the raw material permitted fine flaking control). The initial appearance of the bifaces is an event of fundamental importance for the origin of the human mind because it marks the beginning of a relationship between internal and external representations.77
The hand axes emerged, I would suggest, as a consequence of the inter-play between the brain and the stone tools that humans began to make 2.6 million years ago, mediated by the sensory hand and internal visual representation. Most archaeologists do not recognize a gradual transition from the pebble-tool industry (termed Oldowan) to a stone-tool industry containing hand axes.78 The bifacial tools simply appear in the archaeological record 1.6 million years ago in Africa, marking the beginning of the Acheulean industry. But even a cursory glance at Oldowan artifact assemblages reveals that the technique of bifacial flaking—shaping a piece of stone by chipping it on both sides—can be found throughout the history of this industry, including the earliest known assemblages.79 Many of the partially bifacial artifacts of the Oldowan (some of which are classified as “protobifaces”) resemble crude hand axes to some degree and simply lack a discernible ovate form.80
It is impossible to verify the thesis that hand axes were a consequence of interaction between the early human brain and Oldowan toolmaking, but it seems to be the most logical and parsimonious explanation of their origin. An alternative explanation would be that the production of bifaces was triggered by an event outside the realm of the Oldowan brain and its tools—for example, a previously untapped food source that required an ovate bifacial tool for successful exploitation. Even in this case, however, the earlier history of Oldowan toolmaking probably would constitute an essential prerequisite to hand-ax manufacture. Moreover, I am not convinced that the ovate design of hand axes was critical to their function. The analysis of microscopic wear patterns on their edges indicates that many bifaces were used as tools, but it has yet to be demonstrated that the same tasks could not be performed with Oldowan tools.81
The appearance of the bifaces 1.6 million years ago also marks the emergence of what I have called the proto-mind. The term is applied to the cognitive apparatus of the humans who made bifacial artifacts before the appearance of anatomically modern humans and evidence of modernity or the modern mind (which occurred after 250,000 years ago). Most, if not all, of these humans can be assigned to Homo ergaster and a younger, larger-brained species often classified as Homo heidelbergensis. They evolved the unique ability to externalize mental representations in the form of stone artifacts—as well as probably other artifacts made from materials that have not been preserved in the archaeological record—but lacked the creativity of modern humans. They continued to produce the same external representation, the ovate biface, with only minor variations for more than a million years. In another respect, however, the proto-mind was similar to the modern mind: it was a collective phenomenon that entailed the sharing of one or more mental representations among individuals. The proto-mind was almost certainly a requisite precursor and stepping-stone to the modern mind.
If the identity of the makers of Acheulean bifaces is well established, the identity of those who made the Oldowan artifacts remains controversial. There is consensus that people assigned to the genus Homo—which first appeared at this time—made stone tools, but several other human taxa in sub-Saharan Africa are associated with Oldowan artifacts or evidence for artifact use. The remains of a gracile australopithecine (Australopithecus garhi) were found with mammal bones that exhibit traces of stone-tool damage, and it has been suggested that this species made the 2.6-million-year-old tools at nearby Gona (Ethiopia).82 The remains of a robust australopithecine (Paranthropus) are associated with artifacts in younger deposits at Olduvai Gorge (Tanzania) and Swartkrans (South Africa), and the bones from Swartkrans included some from the hand.83
The importance of the human hand is underscored by some experimental research suggesting that much of Oldowan tool production lay beyond the abilities of the African apes. Beginning in 1990, several pygmy chimpanzees (bonobos) were trained to flake stone tools at the Language Research Center at Georgia State University. The apes struck cobbles with hammerstones and produced sharp flakes. Although many of their artifacts resemble Oldowan forms, comparison with the 2.6-million-year-old Gona tools revealed significant differences. The chimpanzees were unable to produce flakes of the same quality or quantity from a cobble.84 Their brains are of comparable size to those of the australopithecines and not much smaller than those of earliest Homo, so the more constrained movement of their hands probably accounts for some of the contrast. Another factor related to bipedalism is that, from the outset, the quality of the Oldowan tools reflects the selection and transport (that is, carrying in the hands) of high-quality stone from its sources.85
The early Oldowan sites are found in eastern Africa, and, after 2 million years ago, they also appear in southern Africa. By 1.8 million years ago, Oldowan sites are present in southwestern Eurasia; sites containing similar artifacts are known from northern China by about 1.7 million years ago.86 In eastern Africa, Oldowan sites are typically found along the banks of streams and shores of lakes. In southern Africa, they have been discovered in limestone cave fillings that may contain a mixture of human occupation remains, carnivore accumulations, and debris simply washed in from the surrounding area. At the time, African landscapes were becoming drier, with more extensive grassland and fewer trees, while animal life was undergoing significant change (that is, extinctions and new species).87
The interpretation of the Oldowan artifacts has been debated for years. The most widely used approach to classify the tools was proposed by Mary Leakey. Reflecting a traditional functional typological interpretation (at least with respect to many of the artifacts), she assigned the artifacts to categories such as chopper and scraper.88 In 1985, Nicholas Toth published a classic paper on Oldowan artifacts that emphasized the processes of manufacture over the definition of formal types. Toth suggested that formal “types” may not have existed in the minds of the toolmakers and that the Oldowan typological classification had been imposed on a continuum of fractured cobbles and flakes (figure 2.8).89 These seemingly arcane discussions of artifact classification among archaeologists carry heavy implications for human cognition. If the Oldowan tools were not shaped in accordance with mental representations (or “mental templates”), the bifaces that emerged after 1.6 million years ago—which almost certainly do reflect such a template—constitute a fundamentally new and different phenomenon.
The Oldowan industry comprises (1) percussors, or heavy stones used to strike other pieces of stone; (2) variously shaped cores, from which flakes were struck; and (3) the flakes themselves, which also vary in size and shape. Many artifact assemblages also contain (4) retouched flakes, or flakes that were chipped along the edges. According to the traditional classification, retouched flakes are often assigned to functional type categories such as awl and side-scraper, but also may simply represent variations on a continuum of chipping. Nevertheless, retouched flakes become more common in Oldowan sites after 2 million years ago. Much of the spatial and temporal variation in the assemblages appears to reflect differences in the stone used for artifact manufacture or, more specifically, differences in how various stones fracture. Commonly used stone included volcanic lavas and ignimbrites, as well as quartz and quartzite; cherts and flints are less common.90
The making and using of Oldowan tools was driven by human ecology. The tools seem to have played a critical role in allowing early humans to occupy an ecological niche that entailed a significant consumption of meat, especially meat obtained from large mammals. Some sites contain the bones of various large mammals, many of which exhibit traces of damage from cobbles or heavy tools (used to smash them open for marrow) and/or sharp flakes (used to strip off meat). (although others yielded traces of woodworking and cutting soft plant materials).91 Analysis of microscopic wear on the edges of tools from Koobi Fora (Kenya) revealed that some were used for cutting meat92 And at least some of the large-mammal bones in Oldowan sites were derived from fresh carcasses, not leftovers abandoned by carnivores, and may reflect hunting or aggressive scavenging.93 In sum, the Oldowan tools were made and used for very practical purposes, and they probably paved the way for early human expansion into new habitats and climate zones in Eurasia.
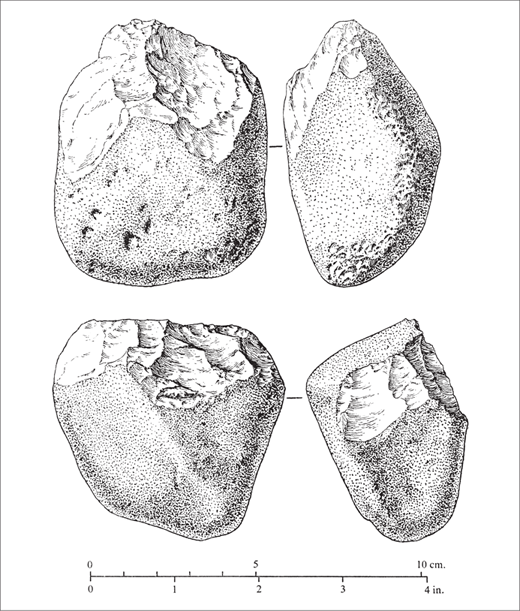
Figure 2.8 Stone tools of the Oldowan industry, classified as unifacial choppers, from Upper Bed I at Olduvai Gorge that do not appear to reflect the imposition of a mental template on a piece of rock. (From Mary D. Leakey, Olduvai Gorge, vol. 3, Excavations in Beds I and II, 1960–1963 [Cambridge: Cambridge University Press, 1971], 77, fig. 40. Reprinted with the permission of Cambridge University Press)
Many Oldowan sites have been discovered and excavated since 1971, when Mary Leakey published her volume on the artifacts from Olduvai Gorge. Nevertheless, the sequence of occupation levels at this most famous of early human sites still offers the best window on the emergence of external thought in the form of Lower Paleolithic bifacial stone artifacts. The occupation levels contain thousands of stone artifacts manufactured during the later Oldowan industry (roughly 2 to 1.5 million years ago) and the early Acheulean (roughly 1.5 to 1.2 million years ago). They represent the debris of human activities—utilized rocks, stone tools, and the waste products of toolmaking—scattered along the shore of an ancient lake that eventually retreated under drying conditions.94 As at many other Oldowan sites, the bones and teeth of mammals as well as other vertebrates are found with the stone artifacts. At least some of the large-mammal bones exhibit traces of tool damage and represent human food debris.95 The remains of both early Homo (H. habilis) and a robust australopithecine (Paranthropus boisei) are found in the occupations.
The major units in the lower part of the sequence at Olduvai Gorge (2 to 1.2 million years ago) are, from the top down:

The Oldowan assemblages represent the younger phases of the industry and yield a healthy quantity of retouched flakes. The upper levels of Bed II contain assemblages—in addition to those assigned to the Acheulean—classified by Mary Leakey as “Developed Oldowan B” that include a small number of bifaces.96 Although the oldest bifaces in Africa are currently dated to about 1.7 to 1.6 million years ago,97 the earliest Acheulean levels in Bed II at Olduvai Gorge do not seem to be significantly younger and represent an early phase of the industry. The most common raw materials used in both industries are quartz and quartzite.
One reason that archaeologists do not perceive a gradual transition from Oldowan to Acheulean is that the traditional typological classification forces individual artifacts into discrete categories. But as Nicholas Toth observed, many Oldowan tools actually occupy places along a continuum of variation.98 And while no formal bifaces have been identified among the assemblages below the upper part of Middle Bed II, there are many examples of bifacially flaked artifacts. They do not seem to reflect a mental template (although we can never be certain that this is the case), but they were partially and sometimes completely flaked; that is, there are no traces of cortex or the original weathered surface of the cobbles. In Lower Bed I, all the “choppers” were bifacially flaked to create a working edge, and some were completely flaked on all sides. Tools classified as discoids also exhibit bifacial working and sometimes complete flaking, creating a symmetrical appearance. And in what may represent a significant trend, proto-bifaces (“intermediate between a biface and a chopper”) become more common in the upper levels (figure 2.9).99
The bifacial tools in the early Acheulean levels at Olduvai Gorge and elsewhere are relatively crude in appearance—some are incompletely flaked and retain cortex—and the differences between them and many Oldowan tools are not dramatic. Nevertheless, when viewed as a group, the ovate artifacts in these and later Acheulean levels reveal a consistent pattern imposed on large flakes or cobbles that departs from the generally random shape of Oldowan tools (figure 2.10). They are subdivided into three types found throughout the Acheulean: hand axes, cleavers, and picks. All three exhibit an ovate form in plan view and cross section.100
The sequence of assemblages at Olduvai Gorge illustrates that humans were producing a variety of flaked-stone artifact shapes for hundreds of thousands of years before the emergence of formal bifaces. The experiments with the pygmy chimpanzees suggest that humans had evolved an unprecedented degree of control over the flaking process with their highly specialized sensory hands. Even more important, the analysis of stone artifacts from a 2.3-million-year-old site in Kenya indicates dynamic interaction between the brain of the toolmaker and the raw material. By refitting flakes and cores at Lokalalei 2C, Hélène Roche and her colleagues discovered that the Oldowan toolmakers were diagnosing—and correcting—imperfections in the surface of the area of the stone from which the flakes were struck.101
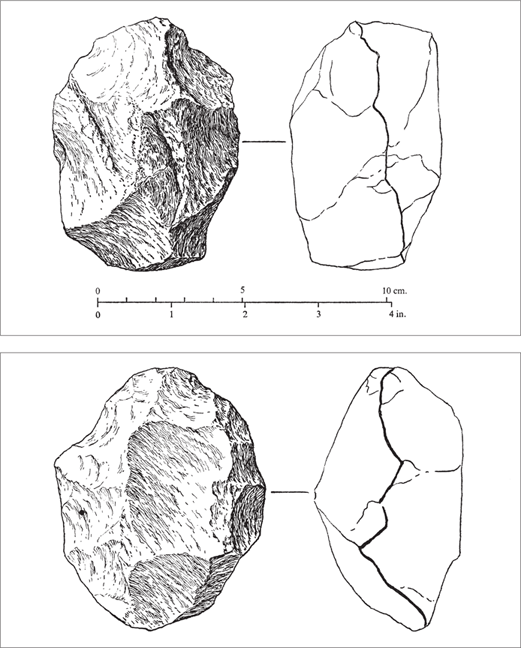
Figure 2.9 Stone tools of the Oldowan industry from the lower part of Middle Bed II at Olduvai Gorge that were flaked on more than one side. Classified as proto-bifaces, they seem to anticipate the flaking techniques used to produce the bifacial tools of the Acheulean industry. (From Mary D. Leakey, Olduvai Gorge, vol. 3, Excavations in Beds I and II, 1960–1963 [Cambridge: Cambridge University Press, 1971], 101–102, figs. 51, 52. Reprinted with the permission of Cambridge University Press)
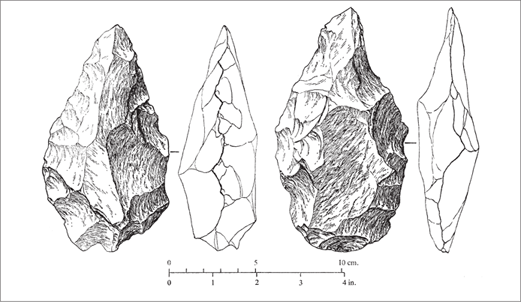
Figure 2.10 Bifacial tools, or hand axes, of the early Acheulean industry from Upper Bed II at Olduvai Gorge reflect an artificial or a semantic mental representation imposed on the rock. (From Mary D. Leakey, Olduvai Gorge, vol. 3, Excavations in Beds I and II, 1960–1963 [Cambridge: Cambridge University Press, 1971], 206, fig. 97. Reprinted with the permission of Cambridge University Press)
The ovate form, I suggest, was a result of the feedback between representations of the artifacts in the brain and the artifacts themselves. Each time an individual flaked a piece of stone, he or she created a series of new visual and tactile representations of the artifact—a nonverbal form of talking to oneself.102 Some early humans eventually reversed the process and worked the stone in accordance with an internal representation. It was a simple three-dimensional geometric form, initially imposed rather clumsily on pieces of rock, shared among individuals in space and time that became a social mental phenomenon.103
Why did early Homo start to make bifacial artifacts or, more specifically, bifacial artifacts shaped in accordance with a preconceived form? The question is a long-standing one among archaeologists, and a definitive answer has yet to be given. Most assume that there is a functional explanation for bifaces: they were made to perform one or more functions that could not be performed with Oldowan tools, or at least not performed as efficiently with Oldowan tools. This assumption is supported by microscopic traces of use-wear on the edges of some bifaces. It nests comfortably within an evolutionary biology framework for human evolution: bifaces were adaptive and enhanced the long-term reproductive success of the humans who made and used them.
A variety of functions have been proposed. A widely held view is that bifaces were used primarily to butcher large mammals. The heavy tools seem to be particularly suitable for chopping through limb joints, and there is supporting evidence from both microwear studies and the repeated association of bifaces with the remains of butchered carcasses. Another suggestion is that they were used to dig through hard soil to obtain plant roots or other buried resources. Some archaeologists argue that bifaces were multipurpose tools—that is, Lower Paleolithic Swiss Army knives—or that they also served as cores, a portable source of fresh flakes.104
One proposed function that emphasizes the shape of bifaces is that they were used as projectiles. According to this view, their design reflects aerodynamic considerations and is tied to the evolution of throwing. It has been advocated with particular vigor by the neuroanatomist William Calvin, who argued that the neural pathways involved in throwing objects provided a basis for the subsequent evolution of syntactic language abilities.105
The initial appearance of bifaces in Africa was broadly coincident with some changes in human anatomy and ecology. The remarkable skeleton from Nariokotome (Kenya) dates to about 1.6 million years ago, and a lower jaw was found with early Acheulean artifacts at Konso (Ethiopia). These fossils belong to Homo ergaster (sometimes they are assigned to the closely related Homo erectus). They reflect an increase in brain size—almost 30 percent larger than the mean for Homo habilis—although the significance of this is mitigated by an almost equal increase in over-all body size. More important, Homo ergaster was essentially modern from the neck down. The tree-climbing adaptations of the australopithecines and earliest Homo are absent, and the hand may be fully modern as well.106
The changes in human anatomy are almost certainly tied to changes in human ecology. Homo had expanded into new regions and climate zones in North Africa and mid-latitude Eurasia—as far as 40° North. Although climates were warmer than those of today, the movement north and out of Africa exposed humans to environments that were cooler, drier, more seasonal, and less biologically rich than those of tropical and subtropical Africa. Food was more dispersed in space and time. One response to this is an increase in foraging range; recent human foragers living in desert, tundra, or other resource-poor environments typically forage over very large areas during the year.107 The changes in postcranial anatomy provided Homo ergaster with a more efficient walking adaptation that probably reflects the demands of wider foraging. Another response is greater reliance on meat, a more efficient food source than a vegetarian diet (recent human foragers in higher latitude environments rely heavily on meat). The analysis of large-mammal remains from upper Bed II at Olduvai Gorge suggests a more intensified focus on meat about 1.5 million years ago.108
The question is therefore two fold: Why did humans start to make formal bifaces, and how were these bifaces related to the changes in anatomy and ecology that took place at roughly the same time? On balance, the strongest case for biface function—supported by several lines of evidence—is that hand axes were used for large-mammal butchery. I think that it is important to bear in mind, however, that Oldowan tools were used effectively for the same purpose. And during the extended period when some humans were making bifaces in Africa and parts of Eurasia, other humans—especially in East Asia—continued to butcher large-mammal carcasses with pebble tools and flakes. Although clearly suitable for butchery, bifaces do not seem to have been essential for this task, and it is difficult to see how they conferred an advantage on those who made and used them. Moreover, the unworn edges of many Acheulean bifaces reveal that a high percentage of them were either not used or used only lightly.109
Bifacial artifacts seem to have played no role in the initial expansion of Homo into Eurasia. The earliest sites in this part of the world were occupied by people who made Oldowan tools. Formal bifaces did not appear in Eurasia until several hundred thousand years after humans had invaded new habitats outside the tropical zone. The oldest site outside Africa is Dmanisi (Republic of Georgia), where humans made Oldowan artifacts 1.8 million years ago. Their skeletal remains reveal that they had already evolved the modern bipedal anatomy of Homo ergaster while retaining the small brain and body of Homo habilis.110
In sum, I would conclude—until a stronger case can be made for their technical and ecological significance—that the function of bifaces as tools was a secondary one. And when they were used as tools, I suggest, their symmetrical design contributed little to their function. The bifaces are not what they seem to be, and evolutionary biology does not provide an adequate framework to explain them. They are based on artificial representations generated in the brain, apparently by the mental manipulation of visual and tactile representations received from the external world through the eye and sensory hand. They are the product of daydreams. As such, they are a manifestation of the “emergent properties” of the mind alluded to in chapter 1 and probably cannot be fully accounted for in terms of the principles of evolutionary biology. Only a philosophy of mind perspective allows us to appreciate their significance.
The appearance of formal bifaces in the archaeological record 1.6 million years ago strikes me as a major event in Earth history. It is analogous to the origin of life itself, which has no function in the context of the natural world; life is dynamic structure generated according to a set of properties that emerged with the formation of complex organic molecules in Precambrian times. The bifaces also reflect the emergence of new properties—in this case, arising out of the recently enlarged human brain, or what I have termed the proto-mind. If they have any analogy to the products of modern human culture, it might be to music, which also lacks any obvious function (although it has acquired various social and economic roles among modern human groups).
Making bifaces nevertheless requires both time and energy, and they must have conferred some advantage on their makers—even if subtle and indirect—in order to “pay for themselves” in the context of human evolutionary biology. Otherwise, it seems that the cost of making them, especially when so many were either unused or under-used, would have conferred a disadvantage on their makers. Not only was biface making a habit that lasted for more than a million years, but the makers and/or the habit spread geographically—from Africa to the Near East by 1.4 million years ago. It is necessary to explain why humans who found the visual–tactile structure of a biface so pleasing prospered and multiplied.
Looking ahead in time, it is clear that the externalization of mental representations, especially after humans evolved the capacity to create increasingly complex representations in the form of technology, eventually conferred immense advantages. By externalizing complex mental representations in the form of language and other symbolic media, members of a group would eventually share complex thoughts among themselves, creating a super-brain with unlimited generative powers, which had extraordinary consequences for them as living organisms. All subsequent developments in the emergence of the mind took place among the direct descendents of the biface makers. These developments provide some insight into why the making of bifaces more than a million years ago may not have been a waste of biological time.
While the bifaces themselves may have lacked practical value, day-dreaming about bifaces may have been only one manifestation of the formation of artificial mental representations, some of which may have had very practical value. Biface making may have been intimately linked to an array of other daydreams externalized in the form of artifacts not preserved in the Acheulean archaeological record. They could have included tools and devices fashioned from plant and animal material that had some concrete benefits to their makers—for example, containers for carrying objects. Some of the microscopic traces of edge-wear from soft plant parts, wood, and hide found on Acheulean stone artifacts may reflect the making of such items.
More generally, thinking about and making bifaces may have been connected to the ability to think about something outside the immediate present in space and time—to create a visual representation, for example, of an object or a place that is not the product of processing incoming signals from the retina. The practical consequences of this ability, even in a limited proto-mind form, could have been profound, especially for humans foraging over wide areas for foods that were becoming increasingly variable in both space and time. It would have helped to imagine situations, predict future events, and plan ahead.
Making bifaces also may have had some impact on social relations within and perhaps even between groups because humans were not only externalizing a representation with their hands, but also communicating it to all other humans who could receive that thought by processing signals from the retina. And one of the most striking characteristics of Acheulean bifaces is their sameness; everyone expressed the same thought. This may have had some impact on social relations—acting as a binding agent within groups—which, in turn, may have had an effect on foraging success. Other mental representations with a potential cohesive influence on social relations may have been expressed not only in materials lacking visibility in the archaeological record, but in nonmaterial form altogether—through vocalization, gesture, or dance.
Landscape of the Proto-Mind
[I]t would be difficult to overemphasize just how strange the handaxe is when compared to the products of modern culture.
Evolutionary biology is not the only interpretive framework applied to Acheulean bifaces. Many archaeologists also have viewed them from a culture-historical perspective, assuming that bifaces were made according to a stylistic convention shared among people and communicated through generations as a cultural tradition. The archaeological literature contains many references to “Acheulean culture.”111 Unlike evolutionary biology, the culture-historical perspective is predicated (at least implicitly) on a philosophy of mind. But it, too, suffers from shortcomings as an interpretive frame-work for the Lower Paleolithic because it is based on the modern mind.
The hand axes or bifaces are almost certainly the product of a mind very different from our own. While unique among the animal kingdom, the mind of the biface makers lacked some fundamental properties of the modern mind. Much of the difference can be accounted for by the significant contrast in brain size. The cranial volume of the early biface makers was between 800 and 1,100 cubic centimeters, which is roughly two-thirds the size of the modern human brain. Their brains would have contained billions of fewer neurons and an exponentially lower potential for synaptic connections. The fossil crania of Homo ergaster nevertheless tell us little about the pattern of thinking that lies behind the bifaces. It is the stuff of the archaeological record that provides most of the insights into this earlier form of mind that I term the proto-mind.
The archaeology required to explain the bifaces and the mind that created them is also very different from the discipline established in the nineteenth century to classify and interpret the artifacts of the modern mind. It would be a variant of Colin Renfrew’s cognitive archaeology, designed to reconstruct or model a different mind on the basis of its external representations and their patterning in space and time. It is the sort of archaeology that would be needed to understand traces of an extinct form of intelligent life found on another planet.
Several major observations about the proto-mind emerge from the archaeological record of the Acheulean. The first is obvious and already discussed at length: the ability to create and externalize an artificial or a semantic representation. As noted at the beginning of this chapter, it is unclear if the shape of bifaces was inspired by a natural object, such as a leaf (figure 2.11). It would seem that their design was a geometric abstraction, like a square, that is unknown in the external world—that is, outside the brain. Perhaps we need look no further than their immediate source: the bifacially flaked Oldowan tools. In any case, the ovate biface was created in the proto-mind and articulated by hand in analogical, or continuous, form; it was not constructed from discrete elements like phonemes or numerals (although the process of shaping it by removing individual flakes has a digital character).
The bifaces of the Acheulean reveal that the proto-mind was capable of thinking about something beyond its internal state. They demonstrate the property of what philosophers of the mind have (rather confusingly) termed intentionality, or “that capacity of the mind by which mental states refer to, or are about, or are objects and states of affairs in the world other than themselves.”112 Despite the possibility or probability that the structure of the biface was generated in the proto-mind with limited reference to the external world, the fact that this representation was projected outside the head is a simple but unambiguous illustration of intentionality. In this way, the bifaces are analogous to a spoken or written sentence.
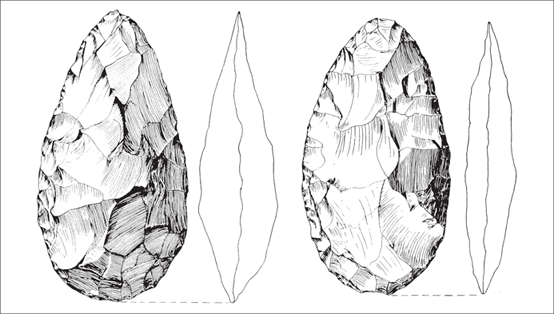
Figure 2.11 Bifacial tools of the Acheulean industry, dating to less than 500,000 years ago, from Kalambo Falls (Zambia) exhibit finer symmetry and shaping than their predecessors. (From J. Desmond Clark, Kalambo Falls Prehistoric Site III: The Earlier Cultures: Middle and Earlier Stone Age [Cambridge: Cambridge University Press, 2001], 337, fig. 6.6. Reprinted with the permission of Cambridge University Press)
The second major observation is that the proto-mind was severely lacking in what Noam Chomsky identified as the “core property” of language—the ability to creatively recombine the elements of externalized mental representations to yield new structures in the same way that modern humans recombine words and sentences to create a potentially infinite array of meaning. The biface makers continued to produce essentially the same structure for more than a million years with relatively minor variation. It is as though they were speaking the same sentence over and over again with minor alterations in grammar and pronunciation.113 There was no history because there was no historical process to create it, and there was no culture, at least in the sense that this term is applied to modern humans. The proto-mind was analogous to an asexually reproducing organism.
There was, nevertheless, some variation in the size and shape of Acheu-lean bifaces. In addition to the three most commonly recognized types (hand axes, cleavers, and picks), they include variants described as pointed, cordate, flat-butted, and ficron, among others.114 The analysis of the differences may provide insight into not only the proto-mind, but also the emergence of the modern mind. Just as Nicholas Toth addressed the question of whether the Oldowan tools reflect the imposition of a mental template, or an internal representation, on flakes and cobbles of stone, some archaeologists have attempted to determine if the people who made the Acheulean bifaces had more than one “type” in their minds. Beginning in the 1960s, François Bordes, Derek Roe, Glynn Isaac, and other Paleolithic specialists applied various measurements and quantitative analyses to samples of bifaces.115
The results of these studies reveal a consistent pattern. Despite the common perception that Acheulean bifaces fall into several types, analyses of large samples have shown that the various forms grade into one another on continuous scales of size and shape. It seems that there was only one rather general representation—subject to what Isaac described as a “random walk” variation—in the proto-mind of the biface makers. Moreover, the variation seems to have been heavily influenced by the size and type of stone used and/or the degree to which flakes or cobbles were reduced by chipping. And it should be noted that the bifaces measured for these studies were derived primarily from later occurrences—archaeological sites that are less than a million years old—and that most probably were made by the larger-brained Homo heidelbergensis. Thus they do not reflect the early phases of the Acheulean.
In his analysis of bifaces at Olorgesailie (Kenya), Isaac noticed an interesting pattern. Although as a whole, the bifaces exhibit the same pattern of continuous variation, specific variants or modes tended to cluster together at particular locations (figure 2.12). At one locality, for example, most bifaces were small, narrow, and relatively thick; at another, they were of intermediate length, contained many chisel-ended forms, and exhibited minimal retouching. Isaac wondered if a single individual had produced most of the bifaces at any one locality or if members of one family or social group had coalesced around a particular design mode within the broader spectrum of variation.116
The same pattern has been noticed elsewhere117 and probably tells us something about the workings of the proto-mind. It suggests a constant or recurring feedback relationship between the internal and external representations of bifaces. In addition to matching mental representations with the received visual and tactile representations of the bifaces they were making, individuals must have received the visual representations of bifaces made by others, especially in their immediate social setting. The feedback process reinforced individual or local trends that perhaps had begun as random variations, a sort of mental “founder’s effect.”
While the production of bifaces as a whole may have followed a random walk pattern, this apparently was not the case among individuals and/ or local groups. Just as Oldowan people spent a million years tinkering with various forms of chipped stone and eventually matched internal and external representations of the ovate biface, Acheulean individuals and/or small groups toyed with the biface form (or “talked to themselves” in this way) for roughly the same length of time. There may have been other externalized representations not preserved in the archaeological record with which they developed a feedback relationship. I have suggested that the matching of internal and external representations by the Oldowan tool-makers underlies the emergence of the proto-mind; similarly, the feedback between internal and external biface representations may have contributed at least in part to the emergence of the modern mind.
The third major observation about the proto-mind is that, like the modern mind, it was a collective enterprise. The biface makers shared mental representations among individual brains, a phenomenon that had evolved in the honeybees but probably not among earlier vertebrates. In comparison with the iconic dancing of the honeybee, laden with vital economic information, the making of bifaces seems rather silly. But perhaps the chief importance of the bifaces was that they constituted shared structure—in this case, “talking to one another”—regardless of content or meaning. This may have had a cohesive effect on social groups. In any case, the archaeological record shows that the biface representations were communicated through time as well as space, from one generation to the next, and that the collective Acheulean brain transcended individual organisms in a way that the shared representations of the honeybee do not.
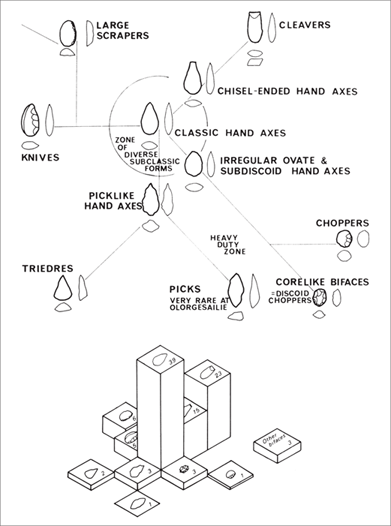
Figure 2.12 Variation in bifaces appears to be continuous and without clear division into subtypes, as observed by Glynn Isaac on the basis of his analysis of stone artifacts from Olorgesaile (Kenya). (From Glynn Ll. Isaac, Olorgesailie: Archeological Studies of a Middle Pleistocene Lake Basin in Kenya [Chicago: University of Chicago Press, 1977], 123, fig. 40. Reprinted with the permission of Barbara Isaac)
A collective brain is a logical, if not inevitable, result of the evolution of the ability to externalize mental representations in a social setting. In addition to their effect on group cohesion, the bifaces also probably reflect a more dynamic cognitive environment. Even if the representations communicated back and forth within and between social groups were devoid of meaningful content, the simple fact of their existence—of the flow of mental representations among a group of individuals—seems significant. I would guess that it was critical to the subsequent emergence of the super-brain and modern mind.
Either the biface makers or the habit of biface making spread from Africa into the Near East roughly 1.4 million years ago. Acheulean bifaces occur at sites of this age in the Jordan Valley at ‘Ubeidiya (Israel).118 They did not spread beyond the Near East into other parts of Eurasia at this time, however. Homo populations in East Asia and the earliest known occupants of Europe did not make bifaces. As far as we know, there were no significant changes in human anatomy or ecology at this time.
After a million years ago, some changes are apparent in Africa that seem to have set the stage for the evolution of modern humans and the modern mind. The volume of the brain significantly increased, and a larger-brained form of Homo is recognized in the African fossil record by about 800,000 years ago. Often referred to as Homo heidelbergensis, reflecting the discovery in 1907 of a fossil jaw in Germany that appears to belong to the same species,119 its estimated brain volume is between about 1,100 and 1,400 cubic centimeters and overlaps substantially with that of modern humans. Homo heidelbergensis retained primitive features of the skull, including a forwardly projecting face, thick supraorbital torus (brow ridge), and receding frontal bone (sloping forehead) (figure 2.13). Some paleoanthropologists nevertheless prefer to classify it as an archaic form of Homo sapiens.120
Coincident with and probably related to the changes in anatomy are changes in artifacts. Acheulean assemblages containing a high proportion of cleavers made on large flakes appear in East Africa after a million years ago (for example, at Olorgesailie [Kenya]), and similar assemblages in the Near East are dated to after 800,000 years ago (for example, at Gesher Benot Ya’aqov [Israel]).121 Both human fossils assigned to Homo heidelbergensis and biface assemblages (often containing many cleavers) are found in Europe by 640,000 years ago and document the spread of the larger-brained Homo into higher latitudes (as far as 52° North in Britain).122 The new biface makers also moved eastward into the Indian subcontinent, where they seem to be represented by both Acheulean artifacts and a partial cranium (Narmada) (figure 2.14).123
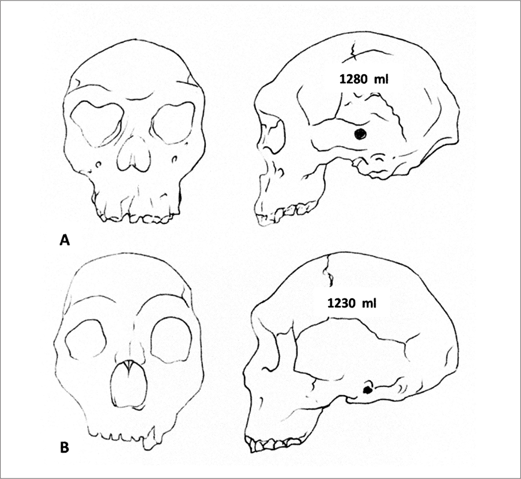
Figure 2.13 Crania often assigned to the later biface maker Homo heidelbergensis reflect a significant increase in brain volume (within the lower range of that of modern humans [mean = ∼1,400 ml]): (A) Kabwe (Zambia) and (B) Petralona (Greece). (Redrawn from G. Philip Rightmire, “Homo in the Middle Pleistocene: Hypodigms, Variation, and Species Recognition,” Evolutionary Anthropology 17 [2008]: 10, fig. 1)
The workings of the proto-mind are very much in evidence during this great dispersal event and provide a sharp contrast to the subsequent migration, also out of Africa, of modern humans. There is evidence for regional variation in the late Acheulean, and many bifaces are shaped with greater refinement and precision than those of the early Acheulean.124 But the lack of creativity is manifest in an essentially uniform pattern of stone-artifact production, especially the bifaces. There is nothing comparable to the spatial and temporal differentiation of material culture that took place after 60,000 years ago when modern humans spread throughout the Old World.
Figure 2.14 Although early biface makers occupied the Near East 1.4 million years ago, the major dispersal of biface-making humans (Homo heidelbergensis) out of Africa took place roughly 750,000 years ago, when they moved into the Near East, western and southern Europe, and southern Asia.