THE AIM OF THIS CHAPTER is to deepen the enactive approach to experience by addressing the relation between time and experience. The experience of time is a long-standing and core theme of phenomenological philosophy. Recently, a number of theorists have combined phenomenological treatments of time-consciousness with dynamic-systems approaches to the mind and brain (Gallagher and Varela 2003; Lloyd 2002, 2003; van Gelder 1999b; Varela 1999). This synthesis plays a central role in the approach to consciousness known as neurophenomenology, proposed originally by Francisco Varela (1996). Neurophenomenology is an offshoot of the enactive approach. Its aim is to incorporate phenomenological investigations of experience into neuroscientific research on consciousness. Neurophenomenology focuses especially on the temporal dynamics of conscious experience and brain activity (Lutz 2002; Lutz and Thompson 2003; Thompson, Lutz, and Cosmelli 2005). In this chapter, I discuss neurophenomenology and its relation to phenomenological accounts of time and experience.
In everyday life we usually act purposively, effectively, and spontaneously, without deliberation, reflection, or explicitly entertaining a purpose. We open the door, greet a friend, make tea, answer the phone, clear the table, and so on. Such actions have a unity or coherence in the present moment, while flowing one into another in a seamless way. Yet there is discontinuity too. We shift rapidly from one activity to another, creating and responding to new situations. Disturbances and breakdowns can happen, disrupting the flow of activity and forcing us to engage anew in what we need to be doing. These continuities and discontinuities testify to the complex relation between embodied action and the temporal structure of experience.
For Merleau-Ponty, as we saw in Chapter 9, embodied action is based on a kind of bodily intentionality he calls motor intentionality. Motor intentionality is the sort of intentionality that characterizes habitual actions and bodily skills, or what Hubert Dreyfus (1991, 2002, 2005) calls absorbed skillful coping. Skillful coping does not have a subject-object structure: one’s experience is not that of standing over against an independent object and then acting on it or finding out about it. Rather, as Merleau-Ponty says, the intentionality in play “is at its object rather than positing it” (1962, p. 446). As Dreyfus explains:
According to Merleau-Ponty, in absorbed, skillful coping, I don’t need a mental representation of my goal. Rather, acting is experienced as a steady flow of skillful activity in response to one’s sense of the situation. Part of that experience is a sense that when one’s situation deviates from some optimal body-environment relationship, one’s activity takes one closer to that optimum and thereby relieves the “tension” of the deviation. One does not need to know, nor can one normally express, what that optimum is. (Dreyfus 2002, p. 378)
In describing the kinesthetic experience of bodily movement in intentional action, Husserl had already stated that its intentional structure is “I can” (move this way) rather than “I think” (a particular thought) (1989, pp. 266–277). Merleau-Ponty takes up this formulation and applies it to motor intentionality: “Consciousness is in the first place not a matter of ‘I think that’ but of ‘I can’” (1962, p. 137). Consider his example (cited in Chapter 4) of the football player in action:
For the player in action the football field is not an “object” … It is pervaded with lines offorce (the “yard lines”; those which demarcate the “penalty area”) and articulated in sectors (for example, the “openings” between the adversaries) which call for a certain mode of action and which initiate and guide the action as if the player were unaware of it. The field itself is not given to him, but present as the immanent term of his practical intentions; the player becomes one with it and feels the direction of the “goal,” for example, just as immediately as the vertical and the horizontal planes of his own body. It would not be sufficient to say that consciousness inhabits this milieu. At this moment consciousness is nothing other than the dialectic of milieu and action. Each maneuver undertaken by the player modifies the character of the field and establishes in it new lines of force in which the action in turn unfolds and is accomplished, again altering the phenomenal field. (1963, pp. 168–169)
This passage contains a number of important ideas. As already mentioned, experience does not have a subject-object structure in immersed skillful action. Instead, experience is the phenomenal flow of one’s body-environment coupling. Furthermore, consciousness here is not detached observation or reflective self-awareness, but rather a nonreflective attunement to the interplay of action and milieu. Finally, this interplay has a certain temporal form or structure. There is clearly a phenomenal flow, but there are also changes from moment to moment. Consciousness has the structure of a nonreflective “I can” in the flow of habitual action, but the leading “I can” can change from moment to moment and modify the character of the flow (Rietveld 2004). As Merleau-Ponty says, “Each maneuver undertaken by the player modifies the character of the field and establishes in it new lines of force in which the action in turn unfolds and is accomplished, again altering the phenomenal field.”
This last idea raises a number of important questions: How do such momentary changes occur? What is the relationship between them and the flow overall? What sort of awareness or consciousness is proper to this experience of punctuated flow? These questions will occupy us throughout this chapter and the next one.
To prepare the way, we need to look more closely at the relation between experience and absorbed skillful action. Dreyfus states (in the passage quoted above) that in absorbed skillful coping “acting is experienced as a steady flow of skillful activity” (emphasis added). This characterization seems exactly right. In skillful coping, we experience our activity (it is not unconscious), and we experience it as a steady flow (unless there is some disruption or breakdown). Yet Dreyfus also suggests in other writings that “absorbed action in the world does not involve an experience of acting” (1991, p. 58; emphasis added). Sometimes he suggests that skillful coping is largely unconscious (2000, pp. 322–323) and that any subsequent report we feel inclined to make about our experience during absorbed action is “a retroactive rationalization of our ongoing activity” (1991, p. 57). This view seems inconsistent with the previous one and phenomenologically questionable. What accounts for this discrepancy?
The basic problem is that Dreyfus seems to think that the only kind of self-awareness is reflective awareness and hence that there is no prereflective self-awareness in skillful coping (see Poellner 2003, pp. 48–51). In this view, in skillful coping we experience a steady flow of activity, but we are unaware of our acting because we are absorbed in acting and do not reflect on it. Indeed, were we to reflect, we would no longer be immersed in the flow of activity but would have arrested it so as to comment on it after the fact. Although we should grant this point about reflection, it does not follow that we are completely unaware of our acting in skillful coping. The reason is that reflective self-awareness is not the only kind of self-awareness. As argued in the previous chapters, experience also comprises a prereflective self-awareness—including in particular a prereflective bodily self-consciousness—that is passive (involuntary) and intransitive (not object-directed). There is every reason to think that this sort of prereflective self-awareness animates skillful coping. If skillful coping were not prereflectively self-aware, then it would not be different from unconscious automaticity and would have no experiential character whatsoever. And if it had no experiential character, then there could be no genuine phenomenology of skillful coping, but only a logical reconstruction of it.
The most phenomenologically plausible way of interpreting the claim that absorbed action does not involve experience of acting is that it does not involve transitive consciousness of acting. To say that skillful coping does not have a subject-object structure means that there need be no transitive or object-directed awareness of either the environment or one’s acting in skillful coping. As Dreyfus says, “in active coping I need not be thematically aware of my current activity,” and although some skillful activities require me to be “explicitly aware of the environment. . . for most skills I can simply respond to the overall gestalt or affordance and need not be aware of it at all” (2000, pp. 322–323). Notice, however, that the kind of awareness being considered here is thematic or explicit awareness. It does not follow from the absence of this kind of awareness that there is no experience of acting at all. On the contrary, it makes sense to suppose that skillful coping, by contrast with mindless automaticity, involves a nonthematic and implicit experience of acting. More precisely, it makes sense to suppose that skillful coping involves—indeed requires—prereflective bodily experience.
Dreyfus appears to deny this idea when he writes, “background coping is necessarily nonthematic and largely unconscious” (2000, p. 323). But we need to draw the necessary phenomenological distinctions. First, there is clearly a crucial difference between activities that we nonthematically experience and activities that are unconscious. The former have an experiential character, and the latter are not experiential in any sense. Second, nonthematic experience can be transitive (object-directed) or intransitive (non-object-directed). Perceptual experience of one’s surroundings, for example, involves a transitive and nonthematic or marginal awareness of the background, as well as a thematic or focal awareness of the foreground. Recall the case of hearing the hum of a refrigerator while focusing intently on a conversation one is having with a friend. One’s awareness of the hum is marginal and nonthematic; one’s awareness of the conversation is focal and thematic.
One’s prereflective and nonthematic experience of one’s body, however, is not a transitive awareness of a marginal or background object, but rather an intransitive and direct acquaintance with one’s bodily subjectivity, an acquaintance whose form is that of a nonintentional and nonobservational bodily self-awareness (Gallagher 2003; Legrand 2006; Zahavi 2002a). If skillful coping were devoid of any experience of acting, if our activities in skillful coping had no experiential character whatsoever, then skillful coping would lack such bodily self-awareness. But then skillful coping would be completely nonsentient. If that were the case, then there could be no phenomenology of skillful coping and Dreyfus’s project would have no foundation.
The implication of this line of thought is that absorbed action in the world does involve experience of acting—or perhaps we should say acting experience—but this experience is usually lived in a prereflective way without becoming the focus of attentive awareness. Our ability to report without observation or inference what we are doing and experiencing depends on such prereflective experience and cannot be explained away simply as retroactive rationalization. Rather, this ability is an ability to make explicit or manifest aspects of our experience that are implicit or latent. Making aspects of experience explicit in this way unavoidably involves interpretation and the creation of meaning (Gendlin 1997). It also presupposes a pregiven background of tacit and unreflective experience that can never be made fully explicit. In these respects, accounts of prereflective experience are interpretive and not merely descriptive. It hardly follows, however, that they are nothing more than retroactive rationalization. Indeed, to claim that they are is ultimately self-defeating because it logically requires some criterion by which to distinguish retroactive rationalization from accurate phenomenological description, and what could the basis for such a criterion be other than some kind of prereflective acquaintance with lived experience?
These considerations reinforce the importance of the idea, shared by phenomenologists from Husserl to Heidegger to Sartre to Merleau-Ponty, that our experience or existence or manner of being comprises an immediate and nonreflective self-awareness, one in which we are acquainted with ourselves prior to any reflection (Zahavi 2005a). In the next section, we will see that this type of self-awareness or self-acquaintance is intimately connected to time-consciousness and the temporal structure of experience.
The analysis of time-consciousness occupies a special place in phenomenology. Any comprehensive analysis of intentionality must take into account the temporal character of the intentional objects of experience. Objects persist and undergo change and transformation; processes unfold and develop in time; and events arise, endure, and cease. Certain things, such as melodies, have different aspects that cannot exist simultaneously, but only across time. Furthermore, at a deeper level, there are temporal “syntheses of identity” at work in experience. A tree along your path presents different profiles as you walk by, but you experience them as belonging to one and the same tree, not as disconnected and momentary fragments. As Merleau-Ponty says, “the perceptual synthesis is a temporal synthesis” (1962, p. 239). In addition, we remember past objects and events, and anticipate future ones. You can remember the bare tree of the winter as you pass by and admire its flowering in the spring, and you can anticipate its changing colors in the fall. The tree as remembered, the tree as perceived, and the tree as anticipated are all intended as one and the same tree. In general, for something to be experienced as objective, it must be experienced as exceeding or transcending the particular act that intends it, and such transcendence in turn requires that we experience it as having an identity across different intentional acts and appearances.
Furthermore, experience itself is temporal, and particular experiences are temporally related to each other. We are aware of our experiences and mental activities as arising, enduring, and ceasing, and as followed by other experiences and mental activities, all related to one another in complex ways. Time-consciousness thus comprises both awareness of external things and their temporal characters, and awareness of experience itself as temporal and as unified across time. Husserl’s account of the structure of time-consciousness is meant to explain both sorts of awareness—how we can be aware of temporal objects (outer time-consciousness) and how we can be aware of our own ebbing and flowing experiences (inner time-consciousness). This account and the myriad analyses it comprises remain the reference point for any phenomenological consideration of time-consciousness.1
Husserl argues that it would be impossible to experience temporal objects if our consciousness of the present moment were the experience of a punctual or instantaneous now. Consciousness of the present moment must rather be an experience of the present as having temporal width. Husserl thus agrees with William James that “the practically cognized present is no knife-edge,” but instead a “duration-block,” a temporal expanse containing past and future phases, or as James says, “a bow and a stern … a forward-and rearward-looking part” (1981, p. 574). But whereas James takes this duration-block, which he calls, following E. R. Clay, “the specious present,” as a basic datum of experience needing neurological explanation, Husserl takes it as requiring further phenomenological analysis (Gallagher 1998, pp. 32–52). More precisely, the duration-block of the present moment is an intentional object of time-consciousness. Hence it needs to be analyzed by tracing it back to the structure of the intentional processes that constitute it—that bring it to awareness, in the sense of enabling us to experience it in the way that we do.
According to Husserl, time-consciousness has a threefold structure. Three intentional processes, called primal impression, retention, and protention, work together in any intentional act and cannot operate on their own apart from one another. Husserl’s favorite example is listening to a melody. For each now-phase of the melody, each currently sounding note, there is a corresponding primal impression directed exclusively toward it. (There are also, of course, primal impressions of the now-phases prior to and after completion of the melody.) Primal impression involves no reference to either past or future, and so by itself is insufficient for the perception of the melody (and for the experience of any temporal object, no matter how brief its duration). Primal impression must be accompanied by retention and protention.
Retention is directed toward the just-elapsed phase of the melody, the just-heard note, which though no longer actual, is still being heard in the mode of just-past. The note still subjectively sounds not because it is really present to consciousness; on the contrary, it is no longer actual. It is present only intentionally; it is mentally intended as just-past.
Protention, by contrast, is directed in a more indefinite way toward the immediate future, in this case toward the notes of the melody sensed as just-about-to-occur. Unlike retention, which is intentionally filled or determinate in content, protention is unfilled or indeterminate. This openness, however, does involve a sense of anticipation, for were the melody to stop abruptly or were a wrong note to sound, we would be surprised. It is always possible in principle for us to be surprised in this way, no matter what the content of our present experience. Thus our consciousness always involves an open and forward-looking horizon.
The unified operation of protention, primal impression, and retention underlies our experience of the present moment as having temporal width. Primal impression is the continuous intending of the new, even if it be only the newly unchanging (as in the case of a continually sounding tone); retention is the continuous holding onto the present as slipping away and sinking into the past; and protention is the continuous going beyond the present as opening into the future. This threefold structure is invariant and present synchronically as a unified whole throughout the course of experience. Although the specific experiential contents of this structure change from moment to moment, the structure itself does not (unless consciousness suffers a radical breakdown).
It is important to distinguish retention and protention (intentional processes proper to the structure of any intentional act) from recollection and expectation (understood as specific types of intentional acts). There is a clear difference between retentional consciousness of the just-played notes of a melody and recollection of a past and finished hearing of the melody. Similarly, there is a clear difference between protentional consciousness of the about-to-be-played notes and expectation of hearing the melody at some time in the future. Retention and protention are structural features of any occurrent experience, and therefore recollection (the remembering of a past experience) and expectation (the anticipation of a future experience) themselves presuppose the work of retention and protention. For this reason, Husserl sometimes calls retention primary memory and recollection secondary memory, and protention primary anticipation and expectation secondary anticipation.
These secondary types of intentional act are re-presentational, whereas the primary forms of experience are presentational. Retention and protention function to make the immediate past and the immediate future intentionally available in our perceptual experience of the now. Recollection and expectation function to bring something absent into a kind of presence by mentally evoking a perceptual experience of it. We recollect a past playing of a melody by recollecting our previous experience of it, and we anticipate a future playing of the melody by anticipating our future experience of it. Finally, whereas recollection and expectation are intentional acts that we can initiate deliberately, retention and protention are passive and involuntary processes that operate continuously.
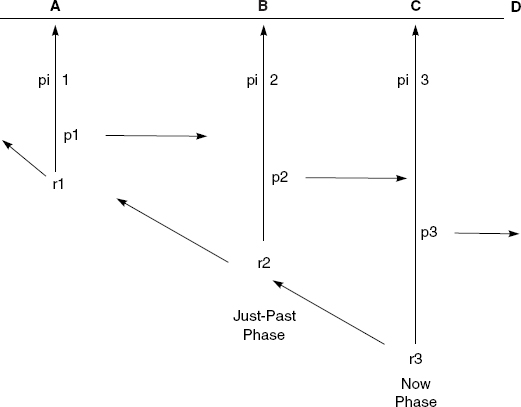
Figure 11.1 illustrates this continuous operation of primal impression-retention-protention. The horizontal line ABCD represents a temporal object, such as a stretch of a familiar melody. The vertical lines represent abstract momentary phases of an enduring act of consciousness. Each phase is structured by the three intentional processes of primal impression (pi), retention (r), and protention (p). If we let phase 3 be the actual now-phase, then there is a primal impression (pi3) of the now sounding note C, a protention (p3) of the next impending phase of consciousness (to be filled by note D), and a retention (r3) of the just-past phase 2 of consciousness, whose primal impression (pi2) was of note B.
Figure 11.1. Time-consciousness according to Husserl. Reprinted with permission from Shaun Gallagher, The Inordinance of Time (Evanston, IL: Northwestern University Press, 1998), 79, fig. 5.1.
As this figure illustrates, any given now-phase of consciousness retains the whole just-past phase of consciousness, specifically this phase’s primal impression, retention, and protention. This just-past phase in turn retains its whole immediately prior phase, this prior phase its immediately prior phase, and so on, so that at any given moment there is a retentional continuum stretching back over past experience. Of course, this stretching back is intentional, not real. Present consciousness does not literally reach back into the past, nor are past phases really inherent in the present phase. Rather, the past is only intentionally contained in the present. The present phase intends the past ones and thereby keeps them in view. Yet the past phases are also perpetually slipping away and disappearing from view. The retentional continuum is characterized by decreasing clarity and affective force as it sinks back into the past, but it is also renewed at each moment, simultaneously being filled with new content while old content perpetually slips away into a sedimented and ultimately unconscious substratum.
Two important types of intentionality belong to this retentional continuum. The first type is the intentional unification of consciousness over time. This unification happens automatically because retention retains the previous phases of consciousness, specifically their interlocking primal impressions, retentions, and protentions. Husserl calls this retention of consciousness the horizontal or lengthwise intentionality of time-consciousness. Thanks to this lengthwise intentionality, consciousness is internally related to itself and self-affecting.
The second type of intentionality ensures the continuity of the experienced temporal object. This intentional process also happens automatically, because the retention of consciousness necessarily includes the retention of the intentional object belonging to the previous phases of consciousness. Husserl calls this type of intentionality the transverse intentionality of time-consciousness. Notice that the transverse intentionality, the retention of the temporal object, is founded on the lengthwise intentionality, the retention of the previous phases of consciousness. In other words, any given now-phase of consciousness retains the just-past phase of its temporal object by virtue of retaining the just-past phase of consciousness. To be aware of intentional objects across time, consciousness must also be retentionally aware of itself.
We can now begin to appreciate the relation between time-consciousness and prereflective self-awareness. As we have seen, each phase of experience retains not only the intentional contents of the just-past experience, but also the just-past phase of the experience itself, including its retentional-impressional-protentional structure. The retention of the just-past notes happens through the retention of one’s just-past experience of them. In other words, retention explicitly grasps the just-past intentional content by implicitly holding onto the just-past experience. Thus the threefold structure of time-consciousness entails prereflective self-consciousness. At the same time one is aware of the melody, one is implicitly co-aware of one’s ongoing experience of that melody, thanks to the threefold temporal structure of one’s experience.
At this point, however, a problem arises. Our focus so far has been on the experience of external temporal objects, such as melodies. But we are also aware of our own transitory intentional acts or experiences and their temporal relations. Indeed, we just saw that in being aware of a melody, we are also implicitly aware of our enduring act of listening. Here then is the problem. In analyzing how there can be such things as melodies for our consciousness (how melodies are constituted), we have appealed to the threefold structure of time-consciousness. But what about our intentional acts or experiences themselves? Is their constitution for consciousness also to be analyzed in terms of the threefold structure of primal impression-retention-protention? But then what is to prevent such an analysis from leading to an infinite regress? If the phenomenal duration and unity of a melody are constituted by time-consciousness, and if our act of listening to the melody is itself also experienced with duration and unity, then are we not forced to posit yet another consciousness to account for this inner duration and unity? Is there any way to stop this regress?
Husserl grappled with this problem throughout his writings on time-consciousness. According to the standard interpretation of his views, Husserl distinguishes among three levels of temporality—external temporal objects (such as melodies), experiences of those objects or intentional acts directed at them, and the experiencing of those intentional acts themselves. Given that we are or can become aware of our experiences and intentional acts as unfolding in time, they also have a temporal character, but one belonging to subjective or immanent time, by contrast with the objective or transcendent time of external temporal objects. Whereas these external temporal objects are brought to awareness or constituted by intentional acts existing in immanent time, these intentional acts are brought to awareness or constituted by a deeper level of inner time-consciousness, which Husserl calls the absolute consciousness or the absolute flow. The absolute flow is supposed to be the deepest level of time-consciousness, a bedrock stratum that is presupposed by every other type of consciousness and intentional performance, and that makes all of them possible.
Given this framework, two questions immediately arise. The first concerns subjective or immanent time. Is it right to think of our intentional acts and experiences as objects of inner time-consciousness (i.e., as internal temporal objects) on analogy with objects of outer time-consciousness or external temporal objects? Does this way of thinking not distort self-awareness and how we experience ourselves in time? The second question concerns the absolute consciousness. Is this consciousness itself experienced? If it is not experienced, then it seems a theoretical construction, with no intuitable content. If it is, then it too must somehow be brought to awareness or constituted by consciousness, but then there looms an infinite regress.
To address the first question properly, it will help to know more about Husserl’s conception of the absolute consciousness. So let me set this first question aside for the moment and return to it after addressing the second one.
Husserl’s view is that consciousness as sheer flow is phenomenologically discernible. We can see, if we look closely and carefully enough (adhering to the epoché and phenomenological reduction), a flowing or streaming belonging to our consciousness that is distinct from the objects or contents of consciousness, that is, from whatever our experience is about or directed toward, whether in the objective and transcendent world, or the subjective and immanent sphere of our own intentional acts and experiences (Husserl 1991, p. 389). This flowing or streaming consciousness is absolute, ultimate, or original, in the sense that it is a condition of possibility for every other kind of consciousness, but is not itself made possible by some other, still deeper level of consciousness.
But given this idea, how can we avoid an infinite regress? If the absolute flow is itself experienced as flowing or unfolding in time, then its temporal phases must be brought to awareness or constituted by some still deeper level of consciousness. In this case the absolute flow is not really absolute, and the whole problem arises once more, with no end in sight.
Husserl tries to deal with this problem in two striking ways. First, he tries to make plausible the idea that the absolute flow is self-constituting, thanks to its horizontal or lengthwise intentionality (Husserl 1991, pp. 390–394). The flow of consciousness retains and protends itself—including retaining its protending of itself and protending its retaining of itself—and is in this way self-unifying. The absolute flow, it is tempting to say, is self-organizing. Second, Husserl denies that the absolute flow is a process unfolding in time or moving through time. The absolute time-consciousness is not one or another temporal process located in the flow of phenomenal time; it is the self-constituting flow of phenomenal time itself. Temporal predicates—“before,” “after,” “simultaneous,” and so on—apply to the temporal objects of our experience, to the enduring, changing, and temporally ordered intentional objects of time-consciousness. But they do not apply literally and univocally to the absolute time-consciousness whereby we experience those objects. Although the absolute flow is temporal or has a kind of temporality, it is not in time in any familiar or ordinary sense. Thus the infinite regress is blocked.
The line of argument here is conceptual, transcendental, and phenomenological. It is a familiar conceptual point that an experience of something as red or loud, for example, is not itself red or loud. Similarly, the timing of what is experienced is not the same as the timing of the experience. For example, an experience whose temporal content is “A before B” does not have to take the form of an experience of A followed by an experience of B (see Dennett 1991a, pp. 148–149). To borrow an example from William James, in experiencing the silence before the thunder, we do not need to have first an experience of silence and then an experience of thunder. As James writes: “Into the awareness of the thunder itself the awareness of the previous silence creeps and continues; for what we hear when the thunder crashes is not thunder pure, but thunder-breaking-upon-silence-and-contrasting-with-it” (1981, p. 234). In general, an experience that presents something as having a beginning, an end, and a duration cannot itself be assumed to have a beginning, an end, and a duration in the same way (see Dennett 1991a, pp. 101–170; Dennett and Kinsbourne 1992; Pylyshyn 2003, pp. 369–370). Husserl recognizes this point, but applies it in a radical way consistent with his transcendental phenomenology.
In general, one cannot assume that concepts and terms applicable to the constituted (that which is given to experience) apply univocally to the constituting (that which makes such experience possible). If phenomenal or lived time (including scientifically understood time) is constituted by the structure of time-consciousness, then the concepts and terms applicable to phenomenal time are not univocally transposable to time-consciousness. But—and this is the crucial point—all our familiar temporal concepts pertain to time as constituted, and hence they cannot apply literally to the time-constituting absolute flow. With this thought in mind, Husserl admits that even the term flow here is metaphorical: “For all of this, we have no names” (1991, p. 382).
This line of thought is also phenomenological. We can distinguish within experience between what changes or varies and what remains invariant. The contents of the present moment—the particular things of which we are aware—arise and perish. But the present moment as a structure of awareness does not change or vary. No matter what we experience, it is always there, or rather always here. It is not a changing content of experience, but an unchanging structure of experience, the threefold structure of primal impression-retention-protention. Husserl calls the present moment understood this way the “living present.” The living present does not flow through time; time “wells up” within it. Husserl describes the living present as “standing-streaming.” It is standing because it does not move in or through time; it is streaming because as the continuous operation of primal impression-retention-protention, it underlies any appearance of flow whatsoever, including the appearance of consciousness to itself as flow.
We can now return to the question about whether it is right to think of our intentional acts and experiences as objects of inner time-consciousness or internal temporal objects. The discussion so far has been premised on the idea that we need to distinguish within experience overall between the experienced external object and the experienced intentional act—between our experience of the melody proceeding in time and our simultaneous experience of our listening act unfolding in time. But in what sense do we experience our listening as distinct from the melody? In particular, does it make sense to say that we experience our listening (or any intentional act) as a temporal object, albeit one of a special, immanent sort?
To address this question, let us return to the point that we experience absorbed activity not as having a subject-object structure, but instead as an immediate coupling or dynamic attunement to our environment. From this perspective, Husserl’s classic descriptions of listening to a melody seem disembodied and abstract. He does not tell us whether the experience is an absorbed listening to a familiar and cherished piece of music or a hearing of a new and unfamiliar work, whether the performance is live or a recording, whether the setting is a celebratory event in the company of others or a solitary and contemplative listening. Of course, Husserl is interested in precisely the invariant structure of time-consciousness, which presumably constitutes the listening experience as such in any possible situation. It is important to grant and recognize the force of this point. Nevertheless, in abstracting away from the concrete setting of the experience, we run the risk of objectifying the experience and thereby misdescribing it. Especially if the listening is an absorbed or immersed one, there will not be any explicit or thematic awareness of the melody as a distinct object, or any explicit or thematic awareness of our ongoing listening.
This observation seriously challenges the idea that we ordinarily experience our listening as any kind of internal temporal object distinct from the heard melody. Recall that object-directed consciousness is transitive. To suppose that we experience our listening as an inner temporal object is accordingly to suppose that in being transitively conscious of the melody we are also transitively conscious of our listening. But this supposition seems mistaken, for the way we ordinarily experience our listening (and our intentional activities generally) is prereflectively and intransitively, not reflectively and transitively.
This point brings us back to the topic of prereflective self-awareness. Dan Zahavi (2003a, 2003b, 2005a) has proposed a different reconstruction of Husserl’s account of temporality, one that equates inner time-consciousness with prereflective self-awareness. According to this view, transitive consciousness of intentional acts and experiences happens only in reflection and certain sorts of recollection. Prior to reflection or recollection there is no transitive or object-directed awareness of experience, certainly no explicit or thematic awareness, but also not even a marginal one. Rather, there is only experience of temporal objects and events in the world, as well as the prereflective and intransitive self-awareness of those very experiences. When we listen to a melody we hear the melody (transitive consciousness), but we also subjectively live through our listening (intransitive consciousness). The listening has a subjective character that makes it immediately manifest, without observation or inference, as one’s own experience. In this way, we experience our listening implicitly, without its becoming an object of awareness—not even a marginal or background one, for the first-personal quality of the experience is precisely its subjectivity and not a type of object-consciousness. The subjectivity of the experience consists essentially in its being intransitively and nonreflectively self-aware. Or rather it consists in its being prereflectively self-aware, for it can come to be reflected upon but is necessarily prior to any such reflection. Only when reflected upon does it come to figure in awareness as a temporal unity of its own in immanent or subjective time.2 Otherwise, it is simply lived through without being individuated as an experience.
Earlier we saw that prereflective self-awareness is a necessary condition for the experience of temporal phenomena. To be aware of phenomena across time, consciousness must be retentionally and protentionally aware of itself across time. Therefore, time-consciousness entails prereflective self-awareness. In other words, our being conscious of external temporal phenomena entails that our temporally enduring experiences of those phenomena are self-aware. Inner time-consciousness is thus nothing other than prereflective self-awareness.
If we understand inner time-consciousness this way, then what becomes of the absolute consciousness or absolute flow? How does it fit into this account?
In answering this question, we arrive at the following fundamental idea: “The absolute flow of experiencing simply is the pre-reflective self-manifestation of our experiences” (Zahavi 2003b, p. 169). Put another way, the structure of inner time-consciousness—primal impression-retention-protention—is exactly the structure of prereflective self-awareness and also precisely the absolute flow. We can appreciate this equivalence in the following way. The absolute flow is the standing-streaming living present. On the one hand, the living present is streaming because it is the continuous transformation (intentional modification) of the about-to-happen into the happening into the just-happened. On the other hand, the living present is standing, because the threefold structure of protention-primal impression-retention that constitutes the streaming is always present and unchanging.
The crucial point now is that this description of the absolute flow or standing-streaming living present also applies precisely to prereflective self-awareness. Preflective self-awareness is streaming because it is constitutive of the streaming or flowing experiences themselves, not a pure and empty awareness that appears on its own. By the same token, it is standing because it is an ever-present and unchanging feature of consciousness. As Zahavi writes, “Whereas we live through a number of different experiences, our self-awareness remains as an unchanging dimension. It stands—to use the striking image of James—permanent, like the rainbow on the waterfall, with its own quality unchanged by the events that stream through it” (2003b, p. 170).
In recent years, scientists and philosophers interested in the temporal dynamics of consciousness have rediscovered Husserl’s analyses of time-consciousness (Lloyd 2002, 2003; van Gelder 1999b; Varela 1999). Varela in particular puts these analyses to use in his neurophenomenological approach to consciousness and offers a neurophenomenological account of time-consciousness as “an acid test of the entire neurophenomenological enterprise” (Varela 1999, p. 267).
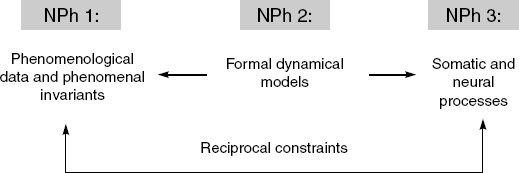
Varela formulates the “working hypothesis” of neurophenomenology in the following way: “Phenomenological accounts of the structure of experience and their counterparts in cognitive science relate to each other through reciprocal constraints” (1996, p. 343). By “reciprocal constraints” he means that phenomenological analyses can help guide and shape the scientific investigation of consciousness, and that scientific findings can in turn help guide and shape the phenomenological investigations. A crucial feature of this approach is that dynamic systems theory is supposed to mediate between phenomenology and neuroscience. Neurophenomenology thus comprises three main elements (see Figure 11.2): (1) phenomenological accounts of the structure of experience; (2) formal dynamical models of these structural invariants; and (3) realizations of these models in biological systems.
Given that time-consciousness is supposed to be an acid test of the neurophenomenological enterprise, we need to see whether phenomenological accounts of the structure of time-consciousness and neurodynamical accounts of the brain processes relevant to consciousness can be related to each other in a mutually illuminating way. This task is precisely the one Varela undertakes in his neurophenomenology of time-consciousness and in his experimental research on the neurodynamics of consciousness.
Varela’s strategy is to find a common structural level of description that captures the dynamics of both the impressional-retentional-protentional flow of time-consciousness and the large-scale neural processes thought to be associated with consciousness. We have already seen how the flow of time-consciousness is self-constituting. What we now need to examine is how this self-constituting flow is supposed to be structurally mirrored at the biological level by the self-organizing dynamics of large-scale neural activity.
There is now little doubt in cognitive neuroscience that specific cognitive acts, such as the visual recognition of a face, require the rapid and transient coordination of many functionally distinct and widely distributed brain regions. Neuroscientists also increasingly believe that moment-to-moment, transitive (object-directed) consciousness is associated with dynamic, large-scale neural activity rather than any single brain region or structure (Cosmelli, Lachaux, and Thompson, 2007). Hence, any model of the neural basis of mental activity, including consciousness, must account for how large-scale neural activities can operate in an integrated or coherent way from moment to moment.
This problem is known as the large-scale integration problem (Varela et al. 2001). According to dynamical neuroscience, the key variable for understanding large-scale integration is not so much the activity of the individual neural components, but rather the nature of the dynamic links among them. The neural counterparts of mental activity are thus investigated at the level of collective variables that describe emergent and changing patterns of large-scale integration. One recent approach to defining these collective variables is to measure transient patterns of synchronous oscillations between different populations of neurons (Engel, Fries, and Singer 2001; Varela et al. 2001). According to Varela (1995, 1999), these synchrony patterns define a temporal frame of momentary and transient neural integration that corresponds to the duration of the present moment of experience.
Varela presents the reasoning behind this view in the form of three connected, but logically independent, working hypotheses (1995, 1999, pp. 274–277):
Hypothesis 1: For every cognitive act, there is a singular, specific neural assembly that underlies its emergence and operation.
According to this hypothesis, the emergence of any cognitive act requires the rapid coordination of many different capacities (attention, perception, memory, motivation, and so on) and the widely distributed neural systems subserving them. The neurophysiological substrate for this large-scale coordination is assumed to be a neural assembly, which can be defined as a distributed subset of neurons with strong reciprocal connections.3
In the context of large-scale integration, a dynamic neural assembly engages vast and disparate regions of the brain. There are reciprocal connections within the same cortical area or between areas at the same level of the network; there are also reciprocal connections that link different levels in different brain regions. Because of these strong interconnections across widely distributed areas, a large-scale neural assembly can be activated or ignited from any of its smaller subsets, whether sensorimotor or internal. These assemblies have a transient, dynamic existence that spans the time required to accomplish an elementary cognitive act and for neural activity to propagate through the assembly.
Various empirical and theoretical considerations suggest that the time-scale of such neurocognitive activity—whether it be a perception/action state (such as an eye or head movement), passing thought or memory, or emotional appraisal—is in the range of a fraction of a second, roughly 250–500 milliseconds or more (see Dennett and Kins-bourne 1992; Pöppel 1988). Varela (1999) calls this scale of duration the “1 scale” of large-scale integration and he distinguishes it from the “1/10 scale” of elementary sensorimotor and neural events (10–100 milliseconds), and the “10 scale” of descriptive-narrative assessments involving memory. During successive time intervals at the 1/10 and 1 scale, there is competition between different neural assemblies: when a neural assembly is ignited from one or more of its smaller subsets, it either reaches coherence or is swamped by the competing activations of other overlapping assemblies. If the assembly holds together after its activation, then one can assume it has a transitory efficacy. This holding time is bound by two simultaneous constraints: (1) it must be larger than the time (at the 1/10 scale) for spike transmission either directly or through a small number of synapses; and (2) it must be smaller than the time (at the 1 scale) it takes for a cognitive act to be completed. Thus the relevant neuronal processes are distributed not only in space, but also over periods of time that cannot be compressed beyond a certain limit (a fraction of a second).
Given Hypothesis 1, the question arises of how large-scale dynamic integration actually happens in the brain so as to make possible a flow of coherent and adaptive cognitive acts. One answer currently being explored in neuroscience is that large-scale integration happens through some form of temporal coding, in which the precise time at which individual neurons fire determines whether they participate in a given assembly. The most well-studied candidate for this kind of temporal coding is phase synchrony. Populations of neurons exhibit oscillatory discharges over a wide range of frequencies and can enter into precise synchrony or phase-locking over a limited period of time (a fraction of a second).4 A growing body of evidence suggests that phase synchrony is an indicator (perhaps a mechanism) of large-scale integration (Engel, Fries, and Singer 2001; Varela et al. 2001). Varela states this idea in the form of a second hypothesis:
Hypothesis 2: A specific neural assembly is selected through the fast, transient phase-locking of activated neurons belonging to subthreshold, competing neural assemblies.
Neural phase synchrony (the phase-locking of oscillatory discharges) occurs at various scales in the brain. Short-range synchronies occur within local networks (columns in primary visual cortex, for example), long-range synchronies over widely separated brain regions. In keeping with Hypothesis 2, long-range synchronization has been proposed as a mechanism for (or at least indicator of) the transient formation of a coherent large-scale neural assembly that selects and integrates multimodal networks (for example, between occipital and frontal lobes, or across hemispheres, which are separated by dozens of milliseconds in transmission time). Animal and human studies demonstrate that specific changes in synchrony occur during arousal, sensorimotor integration, attentional selection, perception, and working memory (Engel, Fries, and Singer 2001; Varela et al. 2001). It has also been hypothesized that whether a local neural process participates directly in a particular (transitive) conscious state depends on whether it participates in a coherent, synchronous large-scale assembly (Dehaene and Naccache 2001; Engel and Singer 2001; Tononi and Edelman 1998; Varela 1995).
Phase synchronies occur in a broad range of frequencies. Fast rhythms (above 15 hertz) in beta and gamma frequencies meet the requirement for rapid integration, and have been shown to play an important role in perceptual and attentional processes on the time-scale of fractions of a second (the 1 scale) (Engel, Fries, and Singer 2001; Varela et al. 2001). Slow rhythms (4–12 hertz) in alpha and theta bands have been shown to play important roles in attention and working memory and in sensorimotor integration.5 These slow rhythms probably also play an important role in motivation and emotion (Lewis 2005), and may provide the context or carrier for fast-rhythm processes (VanRullen and Koch 2003). Taken together, these ideas and evidence support the general notion that phase synchrony subserves the overall integration of all aspects of a cognitive act, including associative memory, affective tone, emotional appraisal, and motor planning (Damasio 1990; Varela 1995; Varela et al. 2001).
Large-scale integration must involve not only the establishment of dynamic links, but also their active uncoupling or dismantling, in order to make way for the next cognitive act. Therefore, not only phase synchrony, but also desynchrony or “phase scattering” is thought to play an important role in large-scale integration (Varela 1995; Rodriguez et al. 1999; Varela et al. 2001, p. 236).6
Varela’s third hypothesis is that this dynamic of synchronization and desynchronization is the neural basis for what we experience as the present cognitive moment:
Hypothesis 3: The integration-relaxation processes at the 1 scale are strict correlates of present-time consciousness.
To explain this hypothesis we need to go back to the distinction between the 1/10 scale and the 1 scale. Elementary sensorimotor and neural events at the 1/10 scale (10–100 milliseconds) are integrated into the 1 scale (250 milliseconds to several seconds) thanks to long-range integration and the formation of large-scale neural assemblies. In the language of dynamic systems theory, a neural assembly must have a “relaxation time,” a time span during which it arises and stabilizes, followed by a bifurcation or phase-transition to a new assembly (a new pattern of long-range integration).
Varela’s hypothesis is that this relaxation time (which is variable and dynamically dependent on a host of factors, such as fatigue, interest, motivation, affect, age, and so on) defines a temporal frame or window of simultaneity, such that whatever falls within this window counts as happening “now” for the system (Varela, Thompson, and Rosch 1991, pp. 72–79; see also VanRullen and Koch 2003). This neurodynamical “now” has an incompressible width or thickness, corresponding to the (variable) frame or window of the 1 scale, within which it incorporates elementary events of the 1/10 scale. It is therefore precisely not a knife-edge present, but a duration-block, having forward and rearward ends, a bow and a stern. According to Varela, this neurodynamical “now” is the neural basis for the present cognitive moment. In other words, the temporal structure of experience, specifically the rearward and forward-looking “now,” depends on the way the brain dynamically parses its own activity. The term strict correlates in Hypothesis 3 is thus misleading because this hypothesis is meant to be causal, not merely correlative. The aim is to explain how the temporal structure of experience is caused by and realized in the dynamic structure of biological processes.
An experimental study by Varela and his colleagues can be used to illustrate these ideas (Rodriguez et al. 1999). This study used the visual recognition of high-contrast, black-and-white patterns as faces (so-called Mooney figures) as a paradigmatic example of a moment of consciousness. This study showed that conscious perception of meaningful complex forms is accompanied by synchronous activities in distant brain regions as recorded at the scalp level by EEG (electroencephalogram) (see Plate VI). Subjects were shown upright and upside-down Mooney figures, which are easily perceived as faces when presented upright, but usually perceived as meaningless black and white forms when presented upside-down. Subjects had to decide as rapidly as possible whether or not they perceived a face at first glance, and then they were asked to report accordingly by pressing on a two-choice button.
When the pattern of phase synchrony between pairs of electrodes in the gamma frequency band (30–45 hertz) was directly measured, it showed marked quantitative and qualitative differences between the “perception” and “no perception” conditions. Between 200 and 260 milliseconds, a first period of significant synchronization was observed in the perception condition but not in the no-perception condition. The synchrony involved left parieto-occipital and frontotemporal regions. Notice that this phase synchrony pattern is long-distance (between pairs of electrodes over widely separated regions of the scalp), unfolds at the 1 scale of duration (roughly 250 milliseconds after stimulus presentation), and corresponds to the moment of perceptual recognition (seeing the face “now”). This synchronization was followed by a period of massive loss of synchronization or phase scattering that peaked at around 500 milliseconds and was observed between parietal and occipitotemporal regions bilaterally. This finding suggests that an active uncoupling or desynchronization may be necessary for the transition from one synchronous neural assembly (and cognitive moment) to another.
Finally, a second period of synchrony increase appeared around 700 milliseconds. This final period coincided with the subject’s button pressing and appears to be related to the motor response in both conditions. This second period is the only period of time in which some similarity between the phase synchrony patterns in the perception and no-perception conditions was noted. Although this study is correlative, not causal, it provides suggestive evidence that long-distance phase synchronization and desynchronization may subserve the temporal parsing of cognition into coherent and momentary acts.7
A crucial feature of this approach is that cognitive time is not based on any external or internal uniformly ticking clock, but rather arises from an endogenous and self-organizing neurodynamics. According to Varela, this dynamics can be described as having a retentional-protentional structure. Each emergent assembly arises as a bifurcation from the immediately previous one and is attracted along a certain forward trajectory, while containing the trace of its predecessor. At one level, each assembly can be seen as an individual pattern that arises, stabilizes, and subsides, thereby giving way to the next assembly. At another level, there is one itinerant trajectory, which wanders among various intrinsically unstable, attractor regions (corresponding to transient neural assemblies), without ever settling down to any one of them, illustrating a type of behavior known as chaotic itinerancy (Tsuda 2001). Varela’s proposal is that the threefold structure of time-consciousness can be redescribed using these concepts of dynamic-systems theory, and that the resulting dynamical description can be grounded in biological features of the brain. In this way, he proposes to “naturalize” the phenomenological account of the structure of time-consciousness.
The key to this naturalized account is the principle of emergence through collective self-organization. As we have seen, emergence through self-organization has two sides: local-to-global determination (the formation of macrolevel patterns through microlevel interactions) and global-to-local determination (the constraining of microlevel interactions by macrolevel patterns). In the present case, the microlevel corresponds to arrays of neurons that behave as coupled nonlinear oscillators (at the 1/10 scale) thanks to their intrinsic cellular properties (see Le Van Quyen, Schuster, and Varela 1996). These oscillators enter into a synchrony or phase-locking that is described by the collective variable of relative phase (at the 1 scale). This collective or large-scale phase-synchrony pattern is a global indicator of the large-scale integration underlying a particular cognitive act.
This emergent act or behavior arises on the basis of the elementary components, but these components act as they do because of the global pattern in which they participate. As Varela observes (1999, p. 283), this sort of neurocognitive self-organization is not an abstract computation, but an embodied behavior subject to initial conditions (what the agent has just done and intends to do) and various exogenous and endogenous parameters (changes in environmental conditions, attentional modulation, motivation, and so on). Neural assemblies and large-scale integration are thus always embedded in and modulated by particular bodily and environmental contexts (Thompson and Varela 2001). Gallagher and Varela summarize this approach in the following way:
The fact that an assembly of coupled oscillators attains a transient synchrony and that it happens within a certain temporal window is the explicit substrate of the living present. The dynamical models and the data show that this synchronization is dynamically unstable and will thus constantly and successively give rise to new assemblies (these transformations define the trajectories of the system). Each emergence bifurcates from the previous ones determined by its initial and boundary conditions. Thus the preceding emergence is still present in the succeeding one as the trace of the dynamical trajectory (retention on the phenomenological level). The order parameters (initial conditions and boundary conditions) are important here. They are defined by the embodiment and experiential context of the action, behaviour, or cognitive act. The boundary conditions shape the action at the global level and include the contextual setting of the task performed, as well as the independent modulations arising from the contextual setting where the action occurs (i.e., new stimuli or endogenous changes in motivation). (Gallagher and Varela 2003, p. 123; see also Varela 1999, p. 283)
So far we have seen that neural coherence (phase synchrony) at the 1 scale is supposed to be the neurobiological correlate of the present cognitive moment and that the dynamical trajectory of neural coherence—the way each transient pattern of coherence contains a trace of its predecessor and is attracted along a certain trajectory—is supposed to be the neurobiological correlate of retention-protention. What about the self-constituting absolute flow or standing-streaming structure of time-consciousness? How is it supposed to be dynamically described and biologically realized?
Recall that the absolute flow is an invariant structure of experience, not a changing content of experience. This point suggests that, in dynamic-systems terms, the absolute flow cannot correspond to any particular attractor region (synchrony pattern) or trajectory in phase space. Rather, it must correspond to an invariant and generic feature of the dynamics, which needs to be described in terms of the geometry of the entire phase space.
Varela proposes that this generic feature is the self-organizing nature of the system’s dynamics, in particular the intrinsic instability or metastability of the system’s dynamics. There are two important aspects to this idea. First, the absolute flow has to be described in terms not of any particular trajectory, but of the collection of all trajectories (all possible behaviors)—in other words at the level of what is known as the system “flow” in dynamic systems theory. Second, the dynamics that shapes this flow, in the case of the brain, is self-generating and metastable. Metastability means that the surface or manifold over which a trajectory courses is unstable, so that the trajectory shifts constantly from one unstable region to another without ever settling down to any one of them. In a metastable system, noise and dynamic instability are intrinsic to the system, so that the system can switch autonomously from one behavior to another, without any external instruction.8 On the one hand, the system is constantly self-moving, transiting from one local instability to another, without ever settling down or staying put. On the other hand, the self-movement that manifests in any trajectory is generically the same. There is thus a kind of standing-streaming character to the dynamics. Self-movement is the nature of the system and is generically the same for any trajectory (standing), but this self-movement exists or manifests itself only in the individual, unstable trajectories (streaming).
So far I have presented neurophenomenology as a theoretical proposal to naturalize the phenomenological account of time-consciousness by redescribing the structure of time-consciousness in the language of dynamic systems theory and mapping this description onto biological processes going on in the brain. But neurophenomenology is not meant to be only a theoretical proposal; it is also meant to be an empirical research program for the experimental investigation of consciousness in cognitive and affective neuroscience (Lutz and Thompson 2003; Varela 1996). In the next section, I discuss experimental neurophenomenology. Then, in the remainder of this chapter, I address some conceptual issues about the neurophenomenological project overall.
There is growing awareness in cognitive and affective neuroscience that detailed and precise first-person descriptions of subjective experience are needed in order to relate moment-to-moment experience to the complexity of brain activity (Jack and Roepstorff 2002).9 Experimental neurophenomenology stresses the importance of collecting descriptive first-person reports of experience from phenomenologically trained subjects as a heuristic strategy for uncovering the physiological processes relevant to consciousness. Such first-person descriptive reports can be called “first-person data,” for they record observations made by individual subjects of their experiences from their first-person perspective.10 Biobehavioral data, by contrast, are “third-person data,” for they record observations of biobehavioral phenomena made by scientists from their third-person perspective. The neurophenomenological approach is to obtain detailed first-person data through careful phenomenological investigation of experience and to use these original first-person data to uncover new third-person data about the physiological processes crucial for consciousness. One central aim of experimental neurophenomenology is thus to generate new data by incorporating careful phenomenological forms of investigation into the experimental protocols of neuroscientific research on consciousness.11
To this end, experimental neurophenomenology employs first-person phenomenological methods in order to obtain original and refined first-person data. Individuals vary in their abilities as observers and reporters of their own experiences, and these abilities can be enhanced through various phenomenological methods. First-person methods are disciplined practices individuals can use to increase their sensitivity to their experience at various time-scales (Depraz, Varela, and Vermersch 2000, 2003; Varela and Shear 1999a, 1999b). These practices involve the systematic training of attention, mindfulness, and self-regulation of emotion. Such methods exist in phenomenology (Moustakas 1994), psychology (Petitmengin-Peugeot 1999; Price and Barrell 1980; Price, Barrell, and Rainville 2002), psychotherapy (Epstein 1995; Gendlin 1981; Stern 2004), and contemplative traditions of mental training (Wallace 1998, 1999). Some are routinely used in clinical and health programs (Kabat-Zinn 1990), and physiological correlates and effects of some of these practices have been investigated (Austin 1998; Lutz, Dunne, and Davidson 2007).
The relevance of these practices to neurophenomenology derives from the capacity for sustained attentiveness to experience they systematically cultivate. This capacity enables tacit and prereflective aspects of experience, which typically are either inaccessible or reconstructed after the fact according to various biases, to become subjectively accessible and describable in more accurate ways. Using first-person methods, individuals may be able to gain access to aspects of their experience (such as transient affective state and quality of attention) that otherwise would remain unnoticed and hence unavailable for verbal report and description. The experimentalist, however, using the phenomenological accounts produced with these methods, may be able to gain access to physiological processes that otherwise would remain opaque, such as the variability in brain response recorded in neuroimaging experiments, often treated as noise. Thus one rationale for using first-person methods in an experimental context is to generate new data (both first-person and third-person) for the science of consciousness.
The working hypothesis of experimental neurophenomenology is that phenomenologically precise first-person data, produced by employing first-person methods, strongly constrain the analysis and interpretation of the physiological processes relevant to consciousness. Moreover, third-person data produced in this manner might eventually constrain first-person data, so that the relationship between the two would become one of dynamic, “mutual,” or “reciprocal constraints” (Varela 1996; Lutz 2002). Reciprocal constraints means (1) that the subject is actively involved in generating and describing specific phenomenal features of his or her experience, (2) that the neuroscientist is guided by these first-person data in the analysis and interpretation of physiological data, and (3) that the (phenomenologically enriched) neuroscientific analyses can provoke revisions and refinements of the phenomenological accounts, as well as facilitate the subject’s becoming aware of previously inaccessible aspects of his or her mental life.12
To establish such reciprocal constraints, both an appropriate candidate for the neurophysiological basis of consciousness and an adequate theoretical framework to characterize it are needed. Experimental neurophenomenology is guided by the theoretical proposal (discussed in the previous section) that the most promising current candidate for the neurophysiological basis of consciousness is a flexible repertoire of dynamic large-scale neural assemblies that transiently link multiple brain regions and areas. This theoretical proposal is shared by a number of neuroscientists, though specific models vary in their details (for a review, see Cosmelli, Lachaux, and Thompson 2007). In this approach, the framework of dynamic systems theory is essential for characterizing the neurophysiological processes relevant to consciousness.
In summary, experimental neurophenomenology is based on the following threefold synergy (compare to Figure 11.2):
1. (NPh1) First-person data from the careful examination of experience with specific first-person methods.
2. (NPh2) Formal models and analytic tools from dynamic systems theory.
3. (NPh3) Neurophysiological data from measurements of large-scale, integrative processes in the brain.
I turn now to review two experimental studies of consciousness and large-scale cortical dynamics based on this approach. Each study concerned the temporal dynamics of visual perception. In the first study, individual subjects gave detailed, trial-by-trial accounts of their experience, and these descriptions were used to define original and stable experiential categories or phenomenal invariants for each individual subject. These phenomenal invariants were then used to detect and interpret neurodynamic activity (Lutz et al. 2002). In the second study, a strict structural invariant of experience for all subjects was defined and then used as a heuristic to reveal a corresponding structural invariant of cortical dynamics (Cosmelli et al. 2004). Both studies were explicitly motivated by the methods and hypotheses of neurophenomenology.
When an awake and alert human subject is stimulated during an experiment, his or her brain is not idling or in a state of suspension, but engaged in cognitive activity. The way the brain responds derives from the interaction between its ongoing activity and the afferent stimulation. Yet because this ongoing activity has not been carefully studied, most of the brain response is not understood. Successive exposure to the same stimulus elicits highly variable responses; this variability is usually treated as unintelligible noise (and may be discarded by techniques that average across trials and/or subjects). The source of this variability is thought to reside mainly in fluctuations of the subject’s attentional state, spontaneous thought processes, strategy to carry out the task, motivation, and so on. Although it is common to control, at least indirectly, for some of these subjective factors, the ongoing subjective mental activity is not usually analyzed systematically.
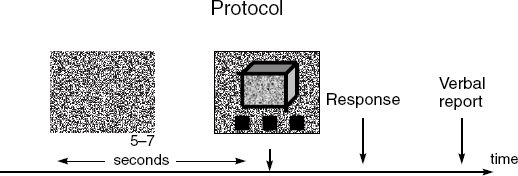
One strategy would be to describe in more detail this ongoing activity by obtaining first-person reports about its subjective character. These reports should reveal subtle changes in the subject’s experience, whether from trial to trial or across individuals. This type of qualitative first-person data is usually omitted from brain-imaging studies, yet if methodological precautions are taken in gathering such data, they can be used to shed light on cognition via a joint analysis with quantitative measures of neural activity. Following this approach, in a pilot neurophenomenological study, Lutz et al. (2002) investigated variations in subjective experience for one limited aspect of visual perception—the emergence of an illusory 3D figure during the perceptual fusion of 2D random-dot images with binocular disparities.
The task began with subjects fixating for seven seconds a dot pattern containing no depth cues (see Figure 11.3). At the end of this “preparation period,” the pattern was changed to a slightly different one with binocular disparities. Subjects then had to press a button as soon as the 3D shape had completely emerged. Throughout the trial EEG signals were recorded, and immediately after the button-press subjects gave a brief verbal report about their experience. In these reports, they labeled their experience using phenomenal categories or invariants that they themselves had found and stabilized during a prior training session. The recording session thus involved the simultaneous collection of first-person data (phenomenological reports) and third-person data (electrophysiological recordings of neural activity and behavioral measures of button-pressing reaction time).
In the training session, subjects intensively practiced performing the task in order to improve their perceptual discrimination and to enable them to explore carefully variations in their subjective experience during repeated exposure to the task. Subjects were instructed to attend to their own immediate mental processes during the task and to the felt-quality of the emergence of the 3D image. In phenomenological terms, the aim was to induce awareness not simply of the “what” or object-pole of experience (in this case the 3D image), but also of the correlated “how” or act-pole of experience (in this case the performance of perceptual fusion and its subjective character).
In this pilot study, this phenomenological attitude was either self-induced by subjects familiar with it or facilitated by the experimenter through the method of open questions (Petitmengin, in press; Petitmengin-Peugeot 1999). For example: Experimenter: “What did you feel before and after the image appeared?” Subject: “I had a growing sense of expectation but not for a specific object; however, when the figure appeared, I had a feeling of confirmation, no surprise at all.” Or “It was as if the image appeared in the periphery of my attention, but then my attention was suddenly swallowed up by the shape.”
Figure 11.3. Visual task for neurophenomenological pilot study of ongoing conscious perception. Reprinted with permission from Antoine Lutz, Jean-Philippe Lachaux, Jacques Martinerie, and Francisco J. Varela, “Guiding the Study of Brain Dynamics by Using First-Person Data,” Proceedings of the National Academy of Sciences USA 99 (copyright 2002, National Academy of Sciences, U.S.A.), 1587, fig. 1.
Subjects were repeatedly exposed to the stimuli, and trial by trial they described their experience through verbal accounts, which were recorded on tape. In dialogue with the experimenters, they defined their own stable experiential categories or phenomenal invariants to describe the main elements of the subjective context in which they perceived the 3D shapes. The descriptive verbal reports from a total of four subjects were classified according to the common factor of degree of preparation felt by the subject and quality of perception. This factor was used to cluster the trials into three main categories: Steady Readiness, Fragmented Readiness, and Unreadiness. Subcategories (describing the unfolding of the visual perception, for example) were also found in individual subjects, but were not investigated in the pilot study.
• Steady Readiness. In most trials, subjects reported that they were “ready,” “present,” “here,” or “well-prepared” when the image appeared on the screen, and that they responded “immediately” and “decidedly.” Perception was usually experienced with a feeling of “continuity,” “confirmation,” or “satisfaction.” These trials were grouped into a cluster SR, characterized by the subjects being in a state of “steady readiness.”
• Fragmented Readiness. In other trials, subjects reported that they had made a voluntary effort to be ready but were prepared either less “sharply” (due to a momentary “tiredness”) or less “focally” (due to small “distractions,” “inner speech,” or “discursive thoughts”). The emergence of the 3D image was experienced with a small feeling of surprise or “discontinuity.” These trials formed a second cluster corresponding to a state of “fragmented readiness.”
• Unreadiness (Spontaneous Unreadiness, Self-Induced Unreadiness). In the remaining trials, subjects reported that they were unprepared and saw the 3D image only because their eyes were correctly positioned. They were surprised by it and reported that they were “interrupted” by the image in the middle of a thought (memories, projects, fantasies). This state of distraction occurred spontaneously for subjects S1 and S4, whereas S2 and S3 triggered it either by fantasizing or by thinking about plans (S3), or by visualizing a mental image (S2). To separate passive and active distraction, these trials were divided into two different clusters: “spontaneous unreadiness” for S1 and S4, and “self-induced unreadiness” for S2 and S3.
These phenomenal invariants found in the training session were used to divide the individual trials of the recording session into corresponding “phenomenological clusters.” The EEG signals were analyzed to determine the transient patterns of local and long-distance phase synchronies between electrodes, and separate dynamical analyses of the signals were conducted for each cluster. The phenomenological clusters were thus used as a heuristic to detect and interpret neural activity. The hypothesis was that distinct phenomenological clusters would be characterized by distinct “dynamical neural signatures” before stimulation (reflecting state of preparation), and that these signatures would then differentially condition the neural and behavioral responses to the stimulus. To test this hypothesis, the behavioral data and the EEG data were analyzed separately for each cluster.
The overall result was that original dynamical categories of neural activity were detected, and hence the opacity in brain responses (due to their intrinsic variability) was reduced. For an example, we can consider the contrast between the two clusters of Steady Readiness and Spontaneous Unreadiness for one of the subjects (see Plate VII). In the first cluster (A), the subject reported being prepared for the stimulus presentation, with a feeling of continuity when the stimulation occurred and an impression of fusion between himself and the percept. In the second cluster (B), the subject reported being unprepared, distracted, and having a strong feeling of discontinuity in the flux of his mental states when the stimulus was presented. He described a clear impression of differentiation between himself and the percept. These distinct features of subjective experience were correlated with distinct dynamical neural signatures (transient patterns of local and longdistance phase synchrony and frequency amplitude).
During steady preparation, a frontal phase-synchronous ensemble emerged early between frontal electrodes and was maintained on average throughout the trial, correlating with the subject’s impression of continuity. The average reaction time for this group of trials was short (300 milliseconds on average). The energy in the gamma band (30–70 Hz) increased during the preparation period leading up to the time of stimulus presentation. This energy shift toward the gamma band occurred in all subjects and was specific to the “prepared” clusters. The energy in the gamma band was always higher in anterior regions during the prestimulus period for subjects in the “prepared” clusters than for subjects in the “unprepared” clusters, whereas the energy in the slower bands was lower. These results suggest that the deployment of attention during the preparation strategy was characterized by an enhancement of the fast rhythms in combination with an attenuation of the slow rhythms.
In the unprepared cluster, however, no stable phase-synchronous ensemble can be distinguished on average during the prestimulus period. When stimulation occurred, a complex pattern of weak synchronization and massive desynchronization or phase-scattering between frontal and posterior electrodes was revealed. A subsequent frontal synchronous ensemble slowly appeared while the phase-scattering remained present for some time. In this cluster the reaction time was longer (600 milliseconds on average). This complex pattern of synchronization and phase-scattering could correspond to a strong reorganization of the brain dynamics in an unprepared situation, delaying the constitution of a unified cognitive moment and an adapted response. This discontinuity in the brain dynamics was strongly correlated with a subjective impression of discontinuity.
Apart from these patterns common to all subjects, it was also found that the precise topography, frequency, and time course of the synchrony patterns during the preparation period varied widely across subjects. These variations should not be treated as noise, however, because they reflect distinct dynamical neural signatures that remained stable in individual subjects throughout several recording sessions over a number of days (see Plate VIII).
In summary, this study demonstrated that (1) first-person data about the subjective context of perception can be related to stable phase-synchrony patterns measured in EEG recordings before the stimulus; (2) the states of preparation and perception, as reported by the subjects, modulated both the behavioral responses and the dynamic neural responses after the stimulation; and (3) although the precise shape of these synchrony patterns varied among subjects, they were stable in individual subjects throughout several recording sessions and therefore seem to constitute a consistent signature of a subject’s cognitive strategy or aptitude to perform the perceptual task.
As we saw earlier in this chapter, time-consciousness has the phenomenal structure of a standing-streaming, an unceasing yet continually changing flow of moments of awareness. Is it possible to take account of this phenomenal temporality while studying the brain processes relevant to consciousness? This question motivated the second experimental study, in which binocular rivalry and magnetoencephalography (MEG) were used to investigate the temporal dynamics of brain activity during conscious perception (Cosmelli et al. 2004).
When two different visual patterns are presented simultaneously, one to each eye, they seem to alternate back and forth at irregular intervals, in a phenomenon known as binocular rivalry. In binocular rivalry, as in other forms of multistable perception such as ambiguous figures, the perception changes while the visual stimulation remains the same. For this reason, binocular rivalry has been used as a probe for finding the neural processes associated specifically with the content of a moment of conscious perception (see Blake 2001; Blake and Logothetis 2002). Yet binocular rivalry also offers an ideal experimental condition to assess the dynamics of cortical activity during ongoing conscious perception, because the perception of a given stimulus fluctuates spontaneously and unpredictably in time.
The rivalrous stimuli used in this experiment were an image of a face and an image of a series of expanding checkerboard rings. The subject was instructed to indicate when one or the other image became visually dominant by pressing one or the other of two buttons. The expanding rings spanned from 0 to 4 degrees of visual eccentricity at a rate of five times per second (5 hertz). Stimuli such as this one that have an intrinsic frequency will produce a concomitant, evoked cortical response (a posterior-anterior wave of activation) at the same frequency. Thus the 5 hertz spanning velocity of the checkerboard stimulus was used as a “frequency tag” to mark neural activity and thereby guide the localization of both the cortical network specifically evoked by this pattern and the modulation of this network throughout successive dominance and suppression periods.
Prior to the recording session, subjects familiarized themselves extensively with the rivalry experience for these stimuli. They gave detailed phenomenological accounts, guided by the method of open questions asked by the experimenter. From these descriptions, the following structural invariant of experience was found to be strictly valid for all subjects: during binocular rivalry, periods of dominance are recurrent through time, while the transitions between periods of dominance are highly variable in the way they arise. Though quite general, this invariant is nonetheless a significant constraint on the detection and interpretation of the underlying neural activity. In particular, one can hypothesize that in binocular rivalry, cortical activity during conscious perception may exhibit a corresponding structure of recurrent dynamic patterns with highly variable transitions.
Testing this hypothesis requires novel tools. One needs to be able to map recurrent brain sources with no restrictions on the temporal sequence of their activation. One also needs to be able to take explicitly into account the variable dynamics of each perceptual transition without averaging (which could destroy potentially relevant information). For these reasons, novel source localization techniques (for inferring underlying cortical source activity from MEG scalp data) and novel statistical analyses (for studying recurrence and variability without averaging) were developed and tested (David et al. 2002; David, Cosmelli, Hasboun, and Garnero 2003).
Using these tools, phase synchrony analysis was performed on the underlying cortical signals reconstructed from the MEG scalp data. The hypothesis was that during conscious perception of the expanding checkerboard rings, phase synchrony in this frequency band would rise and fall in concert with perceptual dominance. A consistent correlation was found between the time-course of the synchronization of the cortical network and the transitions in conscious perception. Thus the overall evoked cortical synchrony correlated with the conscious perception.
To analyze the spatiotemporal dynamics of the specific cortical networks engaged during conscious perception, the ongoing pattern of synchronous brain activity throughout a series of perceptual transitions was followed. A dynamic buildup of the synchronous network was apparent throughout the perceptual transitions. At the beginning of each transition, few synchronous pairs were evident. As perceptual dominance developed, the occipital pole showed an increase in local synchrony involving primary visual cortex and more dorsal occipital areas. Long-range coactivation was then established between occipital regions and more frontal areas, including mainly medial frontal regions. This pattern of an occipitofrontal distributed network was maintained for several seconds and coincided with full perceptual dominance of the frequency tagged, expanding checkerboard rings. Then, as suppression began, long-range synchronous activity fell apart, leaving coactive areas in the occipital pole and, in some cases, inferotemporal regions up to the temporal pole. During full suppression, few coactive regions were left and the brain patterns returned to the pretransition situation. This study thus presented ongoing patterns of cortical synchronous activation correlated to the spontaneous stream of conscious perception.
By using first-person methods to generate new first-person data, and then using these data to make sense of the highly variable brain responses, these two studies illustrate the potential of neurophenomenology for the neuroscience of consciousness. Nevertheless, these studies do not fully embody the neurophenomenological aim to create mutual constraints between phenomenology and neuroscience. Although novel phenomenological data are used to uncover new patterns of brain activity, there is no reciprocal use of biological data to uncover novel phenomenal features of experience. Furthermore, although these studies provide temporally extended, dynamic neural correlates of ongoing conscious states (unlike most studies to-date of the neural correlates of consciousness), they nevertheless remain correlational. Hence they do not cast light on the nature of the relation (beyond correlation) between experiential and biological properties.
In the remainder of this chapter, I address some conceptual issues about neurophenomenology. I begin with how we should think about neurophenomenology in relation to the current scientific aim to discover the neural correlates of consciousness (Metzinger 2000; Rees, Kreiman, and Koch 2002).
In his article “What Is a Neural Correlate of Consciousness?” David Chalmers (2000) distinguishes between two types of neural correlates of consciousness or NCCs—neural correlates of background states of consciousness (background NCCs), such as wakefulness, dreaming, and so on, and neural correlates of the specific contents of consciousness (content NCCs). He defines a content NCC as follows:
An NCC (for content) is a minimal neural representational system N such that representation of a content in N is sufficient, under conditions C, for representation of that content in consciousness. (Chalmers 2000, p. 31)
As this definition indicates, the content-NCC approach is based on the assumption that the contents of consciousness match the contents of neural representational systems. Alva Noë and I call this assumption (which we criticize) the “matching content doctrine” (Noë and Thompson 2004a, 2004b). According to the matching content doctrine, for every conscious experience E, there is a neural representational system N, such that (i) N is the minimal neural representational system whose activation is sufficient for E, and (ii) there is a match between the content of E and the content of N.
The matching content doctrine can be called into question. Although neural correlates of various types of conscious states have been found experimentally, these correlates do not provide any case of a match between the content of a neural system and the content of a conscious state (“match” means equivalence, not mere correlation or correspondence, as can be seen from the above definition of a content NCC) (Noë and Thompson 2004a). Moreover, the very idea that neural systems described neurophysiologically could match conscious states in their content seems inadequate. Experiential content and neural content are different kinds of content, and so it is a category mistake to confuse the two. Experience is intentional (world-presenting), holistic (constituted by interrelated perceptions, intentions, emotions, and actions), and intransitively self-aware (has a nonreflective subjective character). Neural content as standardly described has none of these features. Although there are various sorts of systematic relations between experiences and neural processes, we need not assume that these relations include any content match (Noë and Thompson 2004a, 2004b).
In this context, we need to remember that the neurophenomenology of time-consciousness is not supposed to be a proposal about the neural correlates of the contents of time-consciousness, but rather a proposal about the neural basis of the phenomenological structure of time-consciousness. The proposal is a formal or structural one, not one about representational content. Neurophenomenology derives from embodied dynamicism in its enactive version and thus does not assume the standard representationlist view of the brain (see Chapter 3).
Furthermore, unlike the content-NCC approach, neurophenomenology is not committed to there being a minimally sufficient neural substrate of consciousness.13 It is possible (as argued in Chapter 8) that the minimally sufficient condition for consciousness includes bodily processes. This possibility is not consistent with the NCC view, according to which the neural correlates of conscious states are minimally sufficient for those states. It is consistent, however, with the neurophenomenological proposal that the formal structure of time-consciousness depends on the dynamic structure of brain activity. Neurophenomenology maintains that large-scale integration is necessary, but not minimally sufficient, for time-consciousness. In Varela’s view, bodily processes of emotion and motor activity are crucial to the flow of experience and act as global order parameters for neural events at the 1 and 1/10 scales of duration (Varela 1999; Varela and Depraz 2005). Thus, according to Varela, bodily activity in skillful coping sculpts the entire dynamical landscape of time-consciousness. I discuss this idea in the next chapter.
The content-NCC approach goes hand in hand with a particular research strategy that Searle (2000a) calls the building block model of consciousness. This model is aggregative: it assumes that ordinary multimodal experience can be analyzed into individual sensory experiences (visual experiences, auditory experiences, and so on). The building block approach tries to isolate content NCCs for specific types of individual sensory experiences, such as the visual experience of a perceptually dominant stimulus in binocular rivalry. The ultimate aim is to determine not mere correlates of particular types of conscious states, but the causally necessary and sufficient conditions of these states, including a theory or model of the neurophysiological mechanisms involved. The conjecture is that if we could determine the content NCCs for a particular sort of sensory experience, such as the visual experience of color or visual experience in general, this finding might generalize to other sorts of conscious experiences. To put it another way, if we could determine for color vision or vision in general what makes a given content a phenomenally conscious content, then we might be able to determine what makes a content conscious for any modality.
The building block model also can be criticized. It is problematic to assume that consciousness is made up of various building blocks corresponding to constituent individual sensory experiences, which are then somehow bound together to constitute the unity (or apparent unity) of consciousness. As Searle points out, “Given that a subject is conscious, his consciousness will be modified by having a visual experience, but it does not follow that the consciousness is made up of various building blocks of which the visual experience is just one” (2000a, p. 572).
In criticizing the building block model, Searle singles out another approach to the brain basis of consciousness, which he calls the unified field model. According to this model, the neural substrates of individual conscious states should not be considered sufficient for the occurrence of those states, for those states themselves presuppose the background consciousness of the subject. Any given conscious state is a modulation of a preexisting conscious field. An individual experience or conscious state (such as visual recognition of a face) is not a constituent of some aggregate conscious state, but rather a modification within the field of a basal or background consciousness: “Conscious experiences come in unified fields. In order to have a visual experience, a subject has to be conscious already, and the experience is a modification of the field” (Searle 2000a, p. 572).14 The unified field approach accordingly investigates (1) the neural basis of the whole unified field at the level of large-scale activity, (2) the neurophysiological processes that characterize the dynamic modulation of the conscious field in time, and (3) the differences in brain activity across basal or background states (sleeping, dreaming, wakefulness, and so on).
Experimental neurophenomenology is better interpreted in light of the unified field model than the building block model. Consider the study using binocular rivalry. Binocular rivalry is one of the main experimental paradigms for the content-NCC approach. In the NCC literature, binocular rivalry is usually presented as revealing a competition between two separate and distinct states that alternately become conscious. The research task is to discover the neural activity that represents what the subject sees (the content NCC) for each distinct conscious state (Crick 1994). Each conscious state is treated, in effect, as a building block of a composite rivalry experience. Careful phenomenological examination of the rivalry experience, however, suggests that this experience is not composite in the building block sense. It is not a composite experience having the normal (nonrivalrous) experience of one image and the normal (nonrivalrous) experience of the other image as constituent repeating elements (Noë and Thompson 2004a, pp. 24–25). Rather, it is better described as one bistable experience. In other words, the bistable experience of seeing a face/expanding checkerboard ring is not equivalent to the normal experience of seeing a face, plus the normal experience of seeing an expanding checkerboard ring, plus the two alternating with the appropriate temporal dynamics. As a bistable perception, it is a unique sort of experience, which accordingly must be assessed on its own terms.
Kelso (1995) takes precisely this approach to the perception of multistable figures such as the Necker cube, describing this kind of perception as a metastable state. Similarly, the neurophenomenological study of binocular rivalry takes a dynamic structural approach rather than a building-block content approach. The temporal dynamics of rivalry is considered to be constitutive of the experience rather than an added element in building block fashion. By taking into account detailed phenomenological accounts of the temporal structure of rivalry, a dynamic pattern of recurrence with highly variable transitions is revealed to be a structural invariant of the rivalry experience, a pattern irreducible to an alternation of two stable and clearly differentiated perceptual experiences. By using this structural invariant as a guide to detect and interpret neural activity, the neurophenomenological approach is able to show that ongoing patterns of cortical synchronization and desynchronization correlate with fluctuations of conscious perception.
The neurophenomenological study of depth perception and ongoing mental activity also provides a nice case for the unified field model. This study can be interpreted as an investigation of the dynamic structure of the field of consciousness at phenomenological and neurophysiological levels. It examines “modifications of the already existing field of qualitative subjectivity” (Searle 2000a, p. 563), in the form of modifications of perceptual experience that depend on the subjective experience of readiness or unreadiness. To map the neurodynamics of ongoing conscious states, the study makes use of both proximal and distant baselines, corresponding respectively to the one-second period immediately before stimulation and the seven-second preparation period leading up to stimulation. Using the proximal baseline enhances the contrast between the synchronous process immediately preceding the arrival of the stimulation and the processes triggered by the stimulation. Using the distant baseline reveals a resemblance in synchrony patterns between prestimulus activity and the response induced by the stimulation.
From a unified field perspective, these two baselines correspond to different time slices of the conscious field. The distant baseline takes in more of the preexisting field of subjective experience and thus reveals patterns of dynamic activity that are not seen using the proximal baseline. It also enables one to show that the preexisting conscious field is not only modified by, but also differentially conditions, the emergence of the 3D image. In phenomenological terms, the subjective context of readiness/unreadiness leading up to perception is a noetic factor of the field of consciousness, which differentially conditions the noematic factor of the depth image.
In addition, this study suggests that we need to reconsider the temporal interval of interest for the neural correlate of a conscious cognitive act. The correlate of depth perception clearly occurs between the appearance of the stimulus and the motor response. Yet this momentary conscious state extends from a previous one and finds its place within a temporal horizon defined by retention and protention. This horizon is not neutral but has a certain subjective character (minimally characterized here as readiness/unreadiness). Hence, in order to describe the dynamics of a transitory conscious state, we need to characterize not only the activity following the stimulus, but also the ongoing activity preceding the stimulus and this activity’s protentional character.
According to the unified field approach, “what we have to look for is some massive activity of the brain capable of producing a unified holistic conscious experience” (Searle 2000a, p. 574). One of the working hypotheses of neurophenomenology is that this massive activity involves large-scale integration via phase synchrony (or other forms of “generalized synchrony” across different frequencies; see Friston 2000a, 2000b; Schiff et al. 1996). Large-scale integration is metastable (new patterns constantly arise without the system settling down to any one of them) and rapid (occurring at the 1 scale of 100 milliseconds or more), and it defines a global reference pole for the selection and modulation of local neural activity (Le Van Quyen 2003).
Searle’s proposal is that states of consciousness arise as modifications of the field of basal consciousness, but he does not always clearly distinguish between the two notions of state consciousness and background consciousness. The distinction is important for conceptual, phenomenological, and neurophysiological reasons.
On the one hand, there is good reason to believe that the coherent large-scale activity characterizing transient conscious states occurs in thalamocortical networks (Llinas et al. 1998; Tononi and Edelman 1998). Strong corticocortical and corticothalamic projections can generate a coherent neural process on a fast time scale (the 1 scale), through ongoing reentrant and highly parallel interactions among widely distributed brain areas.
On the other hand, background consciousness depends crucially on brainstem structures and processes (Parvizi and Damasio 2001). Background consciousness is inextricably tied to the homeodynamic regulation of the body and includes a primary affective awareness or core consciousness of one’s bodily selfhood (Damasio 1999; Panksepp 1998b). Background consciousness in this fundamental sense is none other than sentience, the feeling of being alive, the affective backdrop of every conscious state. Sentience—or primal consciousness (Panksepp 1998b) or core consciousness (Damasio 1999)—is evidently not organized according to sensory modality, but rather according to the regulatory, emotional, and affective processes that make up the organism’s basic feeling of self. For this reason, the search for content NCCs in a particular sensory modality such as vision runs the risk of missing the biologically and phenomenologically more fundamental phenomenon of sentience, whose affective character and ipseity (nonreflective self-awareness) underlie and pervade all sensory experience.
Given the dependence of sentience on midbrain and brainstem structures, as well as regulatory systems of the whole body, it is unsatisfactory to say that consciousness, in the sense of a single and unified conscious field, “is in large part localized in the thalamocortical system” (Searle 2000a, p. 574). Although the thalamocortical system is crucial for transient conscious states, midbrain and brainstem structures are crucial for affective core consciousness or sentience. Thus virtually the entire neuraxis seems essential for consciousness in the widest sense of the term (Watt 1998).
In addition to producing new data about the dynamic neural activity relevant to consciousness, neurophenomenology might be able to shed light on the causal structure of this activity. For example, it seems natural to interpret the depth perception study in terms of the dynamic-systems principle of circular causality, in which local events collectively self-organize into coherent global patterns that in turn constrain and modulate local events. Thus the antecedent and “rolling” experiential context of perception modulates the way the object appears or is experientially lived during the moment of perception, and the content of this transient conscious state reciprocally affects the flow of experience. At the neurodynamical level, the brain response presumably results from the intertwining of the endogenous brain activity and the peripheral afferent activity evoked by the stimulus. Therefore the way the stimulus is directly lived by the subject (as indicated by rigorous phenomenological reports using first-person methods) could serve as a heuristic to shed light on the causal structure by which local afferent activity both modulates and is modulated by the contextual influence of the current large-scale neural pattern, such that this local activity either participates in or is discarded from the emergence of the large-scale pattern.
In summary, according to this neurophenomenological perspective, a transitory conscious state is neurally embodied as a dynamic large-scale state that shapes local activities. This transitory state is embedded within a more encompassing state of background consciousness configured by dynamic integration along the entire neuraxis (Thompson and Varela 2001). These multiple level, large-scale states not only emerge from lower level activities, but also mobilize and channel these activities along particular trajectories. In Kelso’s words, “Mind itself is a spatiotemporal pattern that molds the metastable dynamic patterns of the brain” (1995, p. 288).
From the perspective of phenomenological philosophy, an important question to ask about neurophenomenology is what sort of naturalism its naturalized account of time-consciousness presupposes or requires. I will conclude this chapter by addressing this issue.
As a starting point, we can consider the following statement from Husserl’s 1925 lectures on phenomenological psychology:
[T]he synthesis of consciousness is completely different from the external combinations of natural elements . . . instead of spatial mutual externality, spatial intermingling and interpenetration, and spatial totality, it pertains to the essence of conscious life to contain an intentional intertwining, motivation, mutual implication by meaning, and this in a way which in its form and principle has no analogue at all in the physical. (1977, p. 26)
The thought expressed here is that the forms or structures that phenomenology discerns as constitutive of consciousness are completely different from the forms or structures that empirical science discerns in nature. Science treats nature as a realm of entities located in space and interacting causally; phenomenology treats consciousness as a temporal flow of intentional experiences related to each other motivationally. According to Husserl, no analogue of the synthetic unity proper to consciousness is to be found in physical nature.
The neurophenomenology of time-consciousness directly challenges this claim. As we have seen, according to neurophenomenology, the formal structure of time-consciousness or phenomenal temporality has an analogue in the dynamic structure of neural processes. This analogue is revealed by a nonlinear dynamical form of description, a kind of description not available to Husserl (see Roy et al. 1999). This kind of description sees in nature more than simply externally related spatial elements, but also forms or structures that are irreducibly relational (see Chapters 3 and 4) and immanently purposive (see Chapter 6). In this respect, we can say that the kind of phenomenological antinaturalism espoused in the above passage has been outstripped by science (in a way not unlike the way autopoietic biology has outstripped the limits of reason as Kant saw them in relation to the organism and self-organization).15
Yet, according to Varela (1997b), the naturalism neurophenomenology offers is not a reductionist one. Because dynamic systems theory is concerned with geometrical and topological forms of activity, it possesses an ideality that makes it neutral with respect to the distinction between the physical and the phenomenal, but also applicable to both. Dynamical descriptions can be mapped onto biological systems and shown to be realized in their properties (for example, the collective variable of phase synchrony can be grounded in the electrophysiological properties of neurons), and dynamical descriptions can be mapped onto what Husserl calls eidetic features, the invariable phenomenal forms or structures of experience. One reason the naturalism of neurophenomenology is supposed to be nonreductive is that these three types of analysis—phenomenological, biological, and dynamical—are equally needed, and no attempt is made to reduce one to the other or eliminate one in favor of another. For this reason, Varela (1997b) describes neurophenomenology as “triple-braided.”
The question we now face is whether neurophenomenology offers anything more than an isomorphism or one-to-one correspondence between the phenomenological and biological domains, revealed by applying a formal dynamical model to both (Bayne 2004). It is undeniable that Varela’s Hypotheses 1–3 are isomorphic in form, for they propose a one-to-one correspondence between the structure of dynamic neural activity and the structure of time-consciousness. In this respect, neurophenomenology is reminiscent of Wolfgang Köhler’s principle of isomorphism. In Gestalt Psychology Köhler writes: “The principle of isomorphism demands that in a given case the organization of experience and the underlying physiological facts have the same structure” (Köhler 1947, p. 301). Köhler seems to have meant this principle to be a working hypothesis subject to empirical test, not an a priori postulate that any explanation must be isomorphic in form (Scheerer 1994). For this reason, and also because it pertains to structural or organizational features, Köhler’s principle is not equivalent to analytical isomorphism, which assumes that the content of experience is isomorphic to the representational format of brain processes (see Chapter 10). Varela, like other dynamic-systems theorists (Kelso 1995), seems to rely on this structural version of the isomorphism idea as a heuristic to guide the search for dynamic patterns of neural activity pertinent to the structure of consciousness, specifically its temporal structure. Isomorphism serves as a working tool for tracking brain dynamics at the level of large-scale integration but not as an explanatory principle.
Yet neurophenomenology also aims to go beyond this sort of structural isomorphism. Isomorphism suggests an epistemological parallelism in which the biological and phenomenological accounts run on parallel tracks, with no mutual interaction or influence. Neurophenomenology, following Merleau-Ponty’s lead, proposes a more daring idea—that biology and phenomenology can stand in a mutually enlightening, explanatory relation.16 The idea that biology can stand in an explanatory relation to phenomenology is familiar and widely accepted; it is a basic premise of mind science. The idea that phenomenology could stand in an explanatory relation to biology, however, will sound odd to many readers. What could phenomenology possibly explain in this domain? The answer is nothing less than how certain biological processes are also realizations of selfhood and subjectivity, as argued throughout this book.
Clearly, “how” here does not have a causal meaning; the job of phenomenology is not to give a causal account of how selfhood and subjectivity arise in the natural world. Nor does phenomenology aim to give a logical derivation of phenomenal concepts from natural concepts. Rather, phenomenology provides a way of observing and describing natural phenomena that brings out or makes manifest their properly phenomenological features—selfhood, purposiveness, normativity, subjectivity, intentionality, temporality, and so on—which otherwise would remain invisible to science. Put another way, phenomenology offers a way of seeing the inner life of biological systems. That the autopoietic form of an organism realizes a kind of purposive selfhood normatively related to the world, that the dynamic form of neural activity realizes a special structure of temporality—these insights become available only if we bring the resources of phenomenology to bear on biological phenomena. In this way, phenomenology helps to make intelligible the relation between biological life, and mind and consciousness.
We thus arrive at another reason the naturalism of neurophenomenology is nonreductive: in bringing the resources of phenomenology to bear on our understanding of nature, the very idea of nature is transformed (Roy et al. 1999, p. 54; Zahavi 2004b). The physicalist conception of nature as an objective reduction base for the phenomenal no longer holds sway, and instead nature is reexamined from a phenomenological angle. In this way, we find ourselves needing to use certain concepts, most notably that of the lived body, that are essentially mixed or heterogeneous, in the sense that they cannot be factored into the dichotomous categories of the physical and the phenomenal, or the objective and the subjective. It is the possibility of such a mixed discourse—one already actualized in Merleau-Ponty’s writings—that Varela anticipates in describing neurophenomenology as triple-braided. The next chapter pursues this sort of mixed discourse in a discussion of emotion, valence, and consciousness.