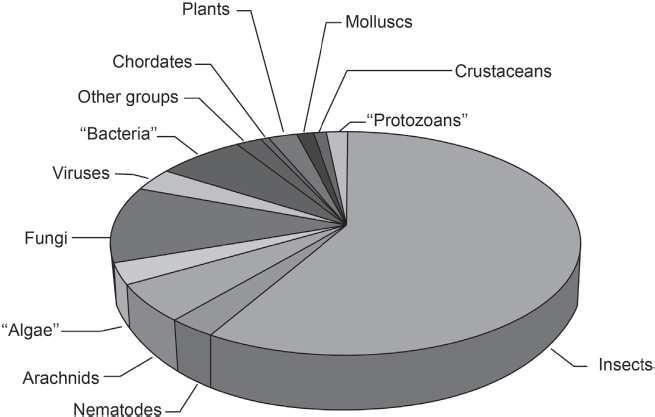
The “jointed legged” animals of the phylum Arthropoda are by far the most numerous, diverse, and successful creatures on this planet. The arthropods include not only insects but also spiders, scorpions, millipedes, centipedes, crustaceans, the extinct trilobites, and dozens of other groups (fig. 15.1). There are over a million different species of arthropods, and some biologists think there may be many millions more, most having not yet been described. Insects alone make up 870,000 species of which there are 340,000 species of beetles. When the great biologist J. B. S. Haldane was asked what nature told him about the Divine, he reportedly said that “God has an inordinate fondness for beetles.” In contrast, there are only 42,000 species of backboned animals and only about 4,000 species of mammals. By nearly any reckoning, arthropods make up 95 to 99 percent of the animal species on Earth (fig. 15.2).
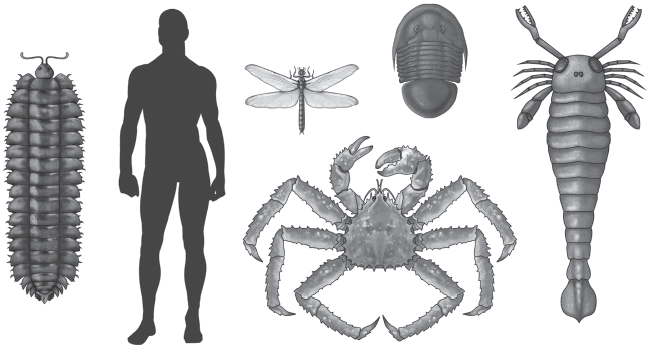
Figure 15.1 ▲
Arthropods come in a wide variety of body forms, including such extinct groups as (top row) trilobites and eurypterids, (second row) chelicerates like scorpions as well as crustaceans, and (bottom row) myriapods like centipedes, plus the largest group of all, the insects. (Courtesy of Wikimedia Commons)
Figure 15.2 ▲
This pie chart shows the diversity of species of the various organisms. Arthropods include arachnids and crustaceans as well as insects, but insects alone outnumber all other groups on the planet combined. In contrast, we chordates (fish, amphibians, reptiles, birds, and mammals) are only a tiny part of the diversity of life. (Courtesy of Wikimedia Commons)
In addition to their diversity, arthropods are also the most numerous animals on the planet. A typical anthill or termite nest might have thousands of individuals. A single pair of cockroaches, if they have unlimited resources and space, could have 164 billion offspring in seven months. In the tropical rainforests, a single hectare may only contain a handful of birds and mammals, but it will have over a billion arthropods, especially mites, bees, ants, termites, wasps, moths, and flies. In the ocean, the plankton in a single cubic meter of seawater may include almost a million tiny crustaceans, especially copepods, krill, ostracodes, and other shrimp relatives.
Arthropods can live in almost any habitat on the planet, from the oceans to fresh water to the land, and many can fly through the skies. They can survive in the hottest deserts, on the coldest ice, and in the saltiest lagoon. Many are parasites that live inside or outside other organisms.
What makes arthropods so successful? Certainly, the fact that they can reproduce rapidly in the right conditions helps. They can lay hundreds of eggs that hatch out and develop, and then they can breed again in a matter of days or weeks. Their small body size means they can live in dense numbers on a limited resource, and they can subdivide into many small subniches that are very closely packed. Most important of all, however, is their modular body plan, which consists of a series of segments that can be added or subtracted or modified. On each segment is at least one pair of jointed appendages, which give the phylum its name: arthros means “joint” and podos means “foot” in Greek. These appendages can be modified into legs, mouthparts, antennae, claws, pincers, swimming paddles, copulatory structures, and a great variety of other possible functions. This modular construction gives them great evolutionary flexibility. Just by changing their embryonic development, they can end up with more or fewer segments, and limbs can be modified as they grow. Some groups (such as the metamorphosis of a caterpillar into a butterfly) can completely change their fundamental body plan in just a few weeks by rearranging and modifying the modules that control each segment.
Another factor that helps them is their external hard shell or skeleton, known as an exoskeleton. In most groups, it is made entirely of the carbohydrate chitin, but in others (like trilobites) it is reinforced with the mineral calcite. The exoskeleton provides support for the body, making it a hollow tube or shell, and the muscles run within it to make it move. It also provides some protection against predators because it is harder than the soft bodies of worms or many other invertebrates, and the exoskeleton also protects against drying out. This allowed arthropods (first millipedes, then centipedes, scorpions, and finally insects like silverfish and springtails) to emerge onto land in the Late Ordovician, and they continued to flourish for more than 100 million years before the fish-like amphibians managed to crawl onto land.
About the only ecological niche that arthropods don’t dominate is the niche for large body size. As animals get larger, their volume (and thus their mass) increases by a cube of their linear dimensions, so organisms get heavier much quicker than they get longer. Their length may increase by only a few centimeters, but their volume increases so fast that they soon test the limits of supporting the weight of their bodies under the rapid increase in gravity. Arthropods have another key limitation: their exoskeleton cannot grow with them, so they must molt by breaking out of it and hardening a new exoskeleton. This is not a problem when they are tiny, but as they get larger and larger, the effects of gravity increase dramatically. Once they pass a certain size, their unsupported soft body after a molt would collapse without support. So the house-sized ants and praying mantises from old-fashioned science fiction movies are a complete impossibility.
The largest land arthropods (fig. 15.3) today are goliath beetles, which are at most about 4.3 inches (110 millimeters) long. However, during the Carboniferous, gigantic millipedes like Arthropleura were over 10 feet (3 meters) long, dragonflies had wingspans of almost 2.5 feet (70 centimeters) across, and cockroaches were almost 1 foot (30 centimeters) long. The Carboniferous was a time when the atmosphere was extremely rich in oxygen, which allowed arthropods (which have relatively inefficient respiratory systems) to get bigger than they ever had been at any other time. The higher density of water supports their bodies better than air, so larger-bodied arthropods exist in the water. The largest today is the king crab, whose legs can span 10 feet (3 meters), but in the past giant eurypterids (sea scorpions) were over 8 feet (2 meters) long.
Figure 15.3 ▲
A few examples of giant arthropods. On the left is the huge millipede relative Arthropleura from the Carboniferous coal swamps. On the extreme right is the eurypterid (sea scorpion) Pterygotus. In the middle at the bottom is the king crab, the largest living arthropod. Above it on the left is the eagle-sized Carboniferous dragonfly Meganeura, and on the right the largest of the trilobites, Isotelus rex. (Illustration by Mary Persis Williams)
Even though arthropods are the most successful, diverse, and abundant animals on the planet, they have a relatively poor fossil record. Only about 30,000 species have been described from fossils, despite the fact that millions are alive now and that many more millions must have lived in the geologic past. Most arthropods have a chitinous exoskeleton, which only fossilizes in exceptional conditions. The one group that fossilizes well is the trilobites, which reinforced their chitinous sheath with the mineral calcite, so there are about 2,000 genera and thousands of species of trilobites. Most collectors or paleontologists will not have the opportunity to collect fossil insects or crustaceans or other types of arthropods. So I will focus instead on the trilobites and a few other extraordinary extinct groups from the geological past.
SUBPHYLUM TRILOBITOMORPHA: TRILOBITES
Trilobites are the most popular fossil among collectors, as well as among many professional paleontologists. Not only are they abundant and easy to collect in many places, but they have an extraordinary variety of shapes, including some with weird appendages and spines, and even some with amazing eyes (fig. 15.4). Trilobites have fascinated people for a long time. A Silurian trilobite carved into an amulet was found in a 15,000-year-old Paleolithic rock shelter at Arcy-sur-Cure, France. Australian aborigines chipped a Cambrian trilobite preserved in chert to form an implement. Both of these specimens were clearly imported from a long distance because they were not present in the local area. The Ute people used to make amulets out of the common Middle Cambrian trilobite Elrathia kingi, from the House Range of western Utah. They called it timpe khanitza pachavee, or “little water bug in stone house.” This fossil is so abundant that it can be commercially mined with backhoes; it is found in virtually every rock shop and commercial fossil seller’s catalog.
Figure 15.4 ▲
Diagram showing trilobite diversification. (Redrawn from several sources)
Trilobites first appeared in the third stage of the Cambrian (Atdabanian Stage), and they are by far the most common fossils of the Cambrian, both because they diversified rapidly and because most other Cambrian animals were soft bodied and did not have hard, fossilizable body parts. By the Late Cambrian, there were 65 families with over 300 genera. In Cambrian shales or limestones, virtually every outcrop yields some trilobites, and they evolved so rapidly that we tell time in the Cambrian with them.
By the Ordovician, trilobites began to decline in importance as other invertebrate groups diversified and came to dominate the seafloor. In addition, trilobites faced their first large predators in the enormous shelled nautiloids (see chapter 16). The groups that survived became much more specialized and distinctive, with adaptations for rolling up for protection, burrowing, swimming, or hiding from their predators. Trilobites were hit hard during the Late Ordovician extinction event. During the Silurian and Devonian, there were only about 60 genera in 17 families. The Late Devonian extinction nearly wiped trilobites out, leaving only a handful of genera in a single family that survived until the end of the Paleozoic. They finally vanished during the great Permian extinction event that wiped out tabulate and rugose corals, most groups of brachiopods and bryozoans, and many other Paleozoic groups.
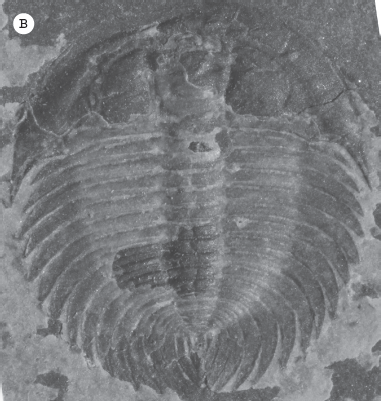
The body of the trilobites can be divided into three parts (fig. 15.5A): the head shield (cephalon), the middle part or thorax made of many segments, and the tail shield, or pygidium. In addition, the trilobite shell has three lobes (hence the name “trilobite”) from side to side: a central axial lobe, and the two sides or pleural lobes. The axial lobe on the cephalon forms a bulbous ridge called the glabella. Each side of the cephalon adjacent to the glabella is called the “cheeks,” and they are split by a line called the facial suture. The pattern of the suture is valuable in identifying major group of trilobites. In primitive forms, the back end of the suture splits the back of the cephalon (known as opisthoparian suture), but in one specialized group with a proparian suture, the back end of the suture splits the side of the cephalon in front of the genal angle (the outer back corner of the cephalon). When the trilobite molted, it split the cephalon along this suture. The part that remained attached to the glabella is called the fixed cheek, and the two parts that split away are called the free cheeks. Many trilobite fossils consist only of a partial cephalon: the glabella plus fixed cheeks (known as the cranidium). However, for many trilobites, these are often enough to identify the species.
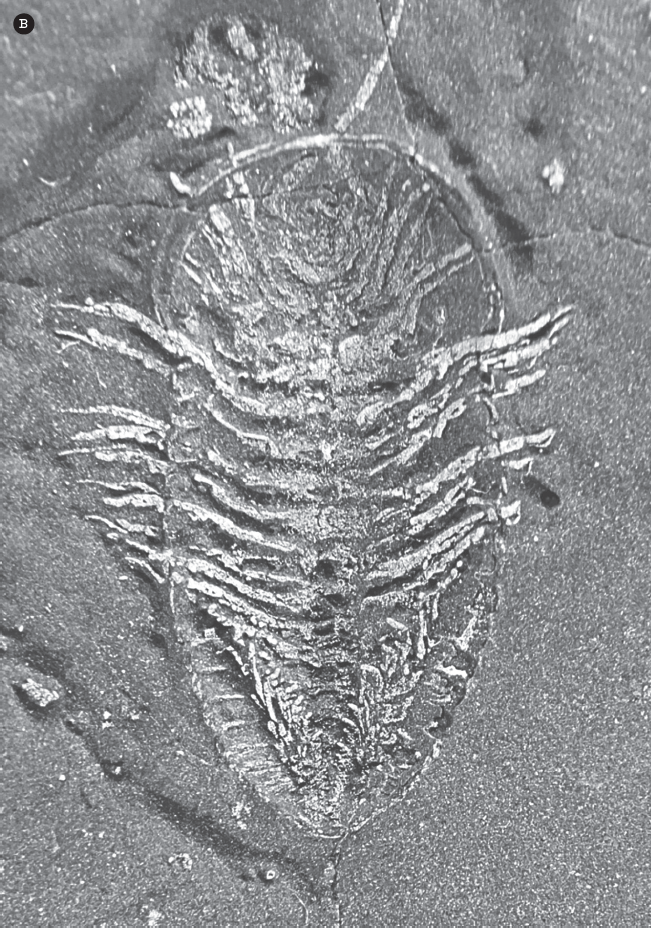
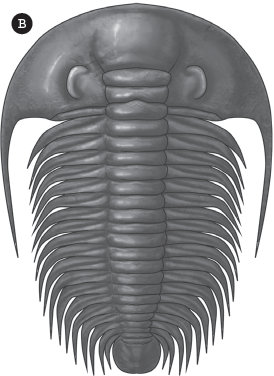
Figure 15.5 ▲
(A) The anatomy of the trilobite shell: The three parts of the shell from front to back are (1) the cephalon or “head shield,” (2) the thorax, and (3) the pygidium or “tail shield.” Number (1) also points to the “nose” or glabella. On each side of the glabella are the eyes. The two bold lines running through the eyes are the facial sutures, lines along which the cephalon splits during molting. The two areas outside the facial sutures are the “free cheeks”; the segment of the cephalon between them including the glabella is called the “fixed cheeks.” The three “lobes” of the trilobite from side to side are (5) the axial lobe and (4, 6) the two pleural lobes. (B; color figure 5) Occasionally, black shales preserve the soft parts of trilobites, here replaced by pyrite. This specimen shows the legs, gills, and antennae. (Courtesy of Wikimedia Commons)
The eyes are usually on each side of the glabella, right along the facial suture. They may be simple semicircular eyes with hundreds of tiny closely packed calcite lenses (holochroal eye) or large elevated bulging eyes with a handful of lenses, each built of two nested parts that correct for spherical aberration (schizochroal eyes). A few trilobites had no eyes at all, and they must have lived in dark muddy bottoms where there was no need for vision.
The cephalon has the most diagnostic features, but in complete trilobites, the number and shape of the thoracic segments, and the presence of spines in certain segments, is often diagnostic. The earliest trilobites had no fusion of the tail segments into a pygidium at all, but once trilobites evolved the pygidium, it took lots of shapes and often diagnoses certain trilobite groups.
Trilobites have an exoskeleton made out of calcified chitin, so the external shell preserves quite readily, even though they often break apart during molts. In fact, most fossils of trilobites consist of only one or two segments because they are simply shed molts; the animal probably swam away to live another day. Trilobite specialists usually need only a few diagnostic parts, such as the cranidium, to identify most species.
Although only the hard calcified shells of trilobites are usually found today, like all arthropods trilobites had legs, antennae, gills, and other appendages surrounded only by their exoskeleton of chitin but uncalcified. Under the right circumstances these softer parts may be preserved, such as in deepwater black shales where pyrite replaces the organic tissues (fig. 15.5B). This provides a rare glimpse at what the legs, gills, and antennae of other trilobites must have looked like.
With about 2,000 genera of trilobites, it is not possible to talk about every group in this book. The sections below summarize some of the most distinctive and commonly collected genera and orders.
ORDER REDLICHIIDA
The redlichiids are the most primitive group of trilobites, starting in the late Early Cambrian and vanishing at the end of the Middle Cambrian. The most familiar family is the extremely primitive Olenellidae (fig. 15.6A), which are collected in many places in the western United States.
Figure 15.6 ▲
Examples of very primitive trilobites from the Early-Middle Cambrian, often put in the “wastebasket” group Redlichiida. (A) The primitive earliest Cambrian trilobite Olenellus, with its simple eyes, many thoracic segments with spines, and a tail spike (telson) instead of a pygidium. (B) Paradoxides, an important Middle Cambrian trilobite. (C) A fossil of Paradoxides davidis, the largest trilobite of the Cambrian, which reached 15 inches (37 centimeters) in length. ([A–B] Illustrations by Mary Persis Williams; [C] courtesy of Wikimedia Commons)
Olenellids were so primitive that they did not fuse their final segments into a pygidium but had a large spike on their tail instead. They had simple half-moon shaped eyes on each side of a glabella with a round knob at the front end, and large genal spines on the outer corners of the cephalon. They had lots of thoracic segments, often with large spines on the “shoulders” of the first few thoracic segments. Early forms like Olenellidae had no visible suture, but later redlichiids such as Paradoxides had opisthoparian sutures (fig. 15.6B–C).
ORDER PTYCHOPARIIDA
The ptychopariids is a large wastebasket group for many different primitive trilobites that dominated the Middle and Late Cambrian. They had a simple glabella that tapers toward the front, a large area in front of the glabella, and many thoracic segments.
The classically well-known Cambrian trilobites include Ptychoparia, Olenus, Triarthrus, and the most common trilobite on the commercial market, Elrathia kingi (fig. 15.7A). This last species is mined in huge quantities from the Middle Cambrian Wheeler Shale in the House Range of Utah and is sold in every commercial market, website, and rock shop in the world.
Figure 15.7 ▲
(A) The common Middle Cambrian ptychopariid trilobite, Elrathia kingi. (B) Lancastria elongata, a corynexochid trilobite. (C) Peronopsis interstrictus, a tiny agnostid trilobite, with no eyes, a large cephalon and pygidium, but only a few thoracic segments. ([A, C] Illustrations by Mary Persis Williams; [B] courtesy of Wikimedia Commons)
The corynexochids are another important Early to Middle Cambrian group. They have a box-like glabella and subparallel facial sutures that are opisthoparian (fig. 15.7B). The thorax has only seven or eight segments, and some have a large pygidium that is roughly the same shape and size as the cephalon (isopygous).
ORDER AGNOSTIDA
These tiny peculiar fossils are so unusual that not all scientists agree that they are trilobites; they may just be a related group of arthropods (fig. 15.7C). They have a button-shaped pygidium the same size and shape as the cephalon (isopygous), so it is the eyes that tell you which is front and which is back. Only two or three short thoracic segments separate their cephalon and pygidium. They are tiny (only a few millimeters long) and may have lived in the plankton rather than grubbing in the bottom muds as most trilobites did. They are most common in the Early to Middle Cambrian but straggled on into the Ordovician. Huge numbers of the tiny agnostid Peronopsis interstrictus are collected from the Wheeler Shale in the House Range of Utah, and they can be found in collections and rock shops everywhere.
ORDERS ASAPHINA AND ILLAENIDA
During the Ordovician, most of the unspecialized Cambrian groups declined or vanished, replaced by many highly distinctive and specialized groups. This was probably due to the pressure of new predators, especially the huge new nautiloids, which could see and grab any trilobite within reach of their tentacles (chapter 16). Two groups, the asaphids and illaenids, coped with this by becoming “snowplow” burrowers and living just beneath the seafloor surface sediments to hide from predators. In both orders, they have a large smooth cephalon shaped like a plow and an isopygous pygidium that mirrored the shape of the cephalon except for the eyes (fig. 15.8A–C). They had only six to nine thoracic segments. Typical asaphids include Homotelus (fig. 15.8C) and Isotelus rex (fig. 15.8A–B), the largest trilobite known, which reached almost a foot in length. Illaenids like Bumastus and Illaenus are also common in collections and on the commercial market.
Figure 15.8 ▲
(A) Asaphids are represented by the “snowplow trilobite,” Isotelus. (B) A specimen of the giant trilobite Isotelus rex from the Ordovician of the Cincinnati area. (C) Another common asaphid is Homotelus, and death assemblages like this one from the Ordovician can be found in the Criner Hills of Oklahoma. (D) Trinucleid trilobites are typified by Cryptolithus, the “lace collar trilobite,” with its distinctive “lacy” brim on the cephalon, prominent glabella, enormous cheek spines, and tiny thorax and pygidium. ([A, D] Illustrations by Mary Persis Williams; [B–C] courtesy of Wikimedia Commons)
Another distinctive group of mainly Ordovician trilobites were the trinucleids. Amateur collectors often call them “lace collar trilobites” because they have a huge cephalon and tiny thorax-pygidium, with a perforated “lacy” edge around the brim of the cephalon. They also have a large knob-like glabella that looks like a big nose. Trinucleids like Cryptolithus (fig. 15.8D) and Trinucleus are common Ordovician fossils and probably survived due to their tiny body size and by burrowing just beneath the surface of the seafloor.
ORDER HARPIDA
Harpids are highly distinctive trilobites, recognized by their broad horseshoe-shaped cephalic brim that wraps around the small thorax and pygidium (fig. 15.9A). The broad cephalic brim was probably suited for plowing through the seafloor sediment.
Figure 15.9 ▲
(A) Harpes, a typical harpid trilobite, with the broad horseshoe-shaped cephalon and tiny thorax and pygidium. (B) Lichas, a typical lichid trilobite, with the “prow” on the front and the large fused pygidium, and tiny bumps all over its surface. (C) Arctinurus boltoni, a typical spiny odontopleurid. (Courtesy of Wikimedia Commons)
Unlike trinucleids, harpids had small eyes on tubercles, a glabella that tapers to the front, and lots of thoracic segments.
ORDER LICHIDA
The lichids are another peculiar and distinctive group of trilobites. Their glabella stretches to the front edge of the cephalon and is subdivided by long glabellar furrows, and they had relatively small free cheeks. Many of them, like Arctinurus (fig. 15.9B) and Radiaspis, had glabella that taper sharply forward with a small “prow” on the front. Their most distinctive feature is their relatively large pygidium, which was often larger than the cephalon and was made of three pairs of expanded spiny pleural segments.
ORDER ODONTOPLEURIDA
Odontopleurids are easy to recognize because most of them are extremely spiny, with spines sticking up from their cephalic brim, genal angle, top of the glabella, all along the thorax, and even from the back of the pygidium (fig. 15.9C). They were relatively rare but were most diverse in the Devonian. They are collected from the Devonian rocks of the Tindouf Basin of Morocco and can be found in many commercial markets.
One of the most interesting and distinctive of the trilobite orders was the phacopids. They are the only group with proparian facial sutures, and most of them were capable of rolling up into a tight ball like sowbugs or pillbugs or “roly-poly bugs” (which are not true bugs but isopod crustaceans that have moved from the tide pools to the land) found in leaf litter today. In the Ordovician and Silurian, they were represented by the distinctive calymenids such as Calymene and Flexicalymene (fig. 15.10A–B). Calymenids had lots of thoracic segments, and they even had segmented furrows on the glabella, with their eyes out on stalks. Another branch includes the Silurian index fossil Dalmanites (fig. 15.10C), and the common Devonian phacopids, including Phacops, Greenops, Eldredgeops (fig. 15.10D–E), and others. These phacopids have a broad curved cephalon and a broad glabella that expands forward covered with tiny bumps. They had large schizochroal eyes that stick up above the cephalon, giving them almost a spherical view of the world above them, with stereovision between the lenses. Some, like Walliserops (fig. 15.10F) have spines not only along the top and above their eyes, but also a strange trident-like spine in front.
Figure 15.10 ▲
(A) The Silurian index fossil Dalmanites, with the pointed spiky pygidium and large schizochroal eyes. (B) Flexicalymene, a typical Ordovician calymenid, with lots of segments in the thorax, eyes on stalks, and a furrowed glabella. (C) Flexicalymene rolled up in a defensive ball. (D–E) The distinctive Devonian phacopid trilobite Phacops, with huge schizochroal eyes, a broad cephalon covered in bumps, and many thoracic segments. (F) Walliserops, a spiny phacopid with a remarkable spiny fork on the front and hooks on its eyes. ([A–B, E] Illustrations by Mary Persis Williams; [C–D, F] courtesy of Wikimedia Commons)
ORDER PROETIDA
The Late Devonian extinction event wiped out the phacopids and odontopleurids and the other dominant Devonian trilobite groups; only one order managed to survive through the rest of the Paleozoic. Proetids were small trilobites with relatively primitive body forms, a large vaulted glabella, opisthoparian sutures, and most of them are isopygous. They had primitive holochroal eyes and a furrowed pygidium without spines. When these last stragglers of the trilobite radiation finally vanished in the great Permian mass extinction, trilobites were completely extinct.
SUBPHYLUM CHELICERATA
The chelicerates include the arachnids (spiders, scorpions, whip scorpions, pseudoscorpions, harvestmen or “daddy long legs,” mites, ticks, and chiggers) plus the merostomes. With over 70,000 living and extinct species, the chelicerates are the second largest group of arthropods after insects. They all have a distinctive anatomy of head region (prosoma or cephalothorax) and abdomen (or opisthoma). They do not have antennae; instead, their first pair of appendages, known as chelicerae, are found in their mouthparts. Their second pair of appendages, the pedipalps, sometimes serve as mouthparts, but in scorpions, some spiders, and eurypterids, they are modified into pincers for capturing prey.
The most familiar and only living merostomes are the horseshoe crabs (fig. 15.11A), which are not closely related to true crabs (Subphylum Crustacea) at all. They are often called living fossils because some of the Mesozoic forms (fig. 15.11B) look somewhat like modern species, but in the Paleozoic they experimented with many other body shapes as well.
Figure 15.11 ▲
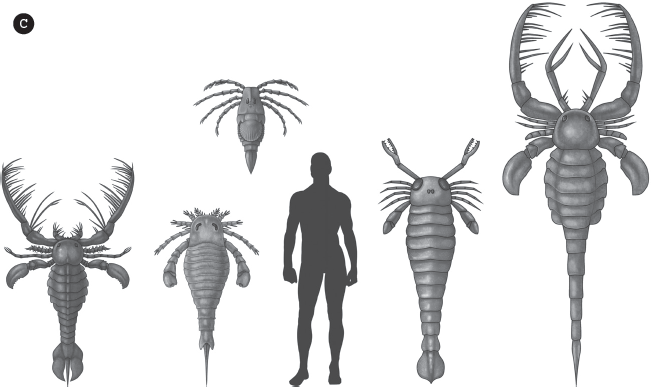
Merostomes: (A) The living horseshoe crab, Limulus polyphemus, still common on the Atlantic Coast during very high tides; (B) a fossil horseshoe crab, Mesolimulus walchi, from the Upper Jurassic Solnhofen Limestone of Germany. (C) Eurypterids came in many different sizes and shapes. (D) Specimens of smaller Eurypterus remipes, a common fossil from the Bertie Limestone of New York. (E) Life-sized model of a huge eurypterid. ([A–D] Courtesy of Wikimedia Commons; [E] photograph by the author)
For paleontologists, however, the most impressive chelicerates in the fossil record are the “sea scorpions,” or eurypterids. Their nickname is a misnomer because they are not closely related to scorpions (which are arachnids, not merostomes) and apparently lived in fresh water and not just in the sea. However, their body form is reminiscent of scorpions with large pincers out front, and spidery legs flaring out from their prosoma (fig. 15.11C). More impressive is the fact that they were one of the world’s largest predators during the Silurian. The largest nearly complete eurypteid fossil is Pterygotus (fig. 15.11D), which as about 7 feet (2.1 meters) long, and the giant Jaekelopterus, which may have been about 8.2 feet (2.5 meters) long. Pterygotus had a long body with a flat swimming tail and a pair of appendages modified into paddles, with huge pincers up front. Stylonurus was more spider-like with a spike on its tail and no swimming paddles. Mixopterus had long spiny chelicherae and pedipalps, spiny walking legs, and a tail like a scorpion.
Altogether, there were about 70 genera and 250 species of eurypterids, ranging from the Ordovician to the Permian. They were the largest marine predators of the Silurian, and they continued that role in the Devonian. In the United States, the famous Silurian Bertie Limestone of upstate New York produces a majority of the specimens of Eurypterus remipes (fig. 15.11E) found in collections and in the commercial market.