Ecology in the Vineyard and the Winery
One recent early summer day, we had the pleasure of visiting several wineries in northern California’s Alexander Valley, one of the state’s most outstanding growing regions. The sun was shining brightly; the sky was clear; and a warm wind swept over the valley. We sat close to the edge of the vine rows, sampling a glorious local Cabernet Sauvignon and watching as the light breeze ruffled the glinting leaves on the receding rows of vines. As lazy observers we appreciated the pastoral beauty of the scene. But what the biologists in us also saw was a landscape of sex and death.
The fields of ecology and evolution could be characterized as studies of sex and death in nature. And although both fields are fairly new as scientific disciplines, the spirit underlying them is age old. Both Aristotle and Hippocrates wrote descriptive accounts of the natural world around them, and it escaped neither that the objects through which they described that world were knit together, and given meaning, by the interactions among them. Aristotle’s fourth-century B.C.E. protégé Theophrastus wrote extensively about plants, and was specific about how they should be understood. In the prescient statement that opens his classic Enquiry into Plants he made his approach clear: “We must consider the distinctive characters and the general nature of plants from the point of view of their morphology, their behavior under external conditions, their mode of generation, and the whole course of their life.” Theophrastus was, in fact, describing an ecological evolutionary approach to understanding plants, and ultimately he extended this focus on interactions to grapes and other fruits in a short work titled On Wine and Olive Oil. There he discussed the ripening of fruit (especially wine grapes) in the specific context of environmental conditions, particularly sunshine and heat.
So why have biologists been so obsessed with sex and death in the natural world? No mystery there—that’s where much of the action is in the ecological and evolutionary spheres. Evolutionary biologists often use the term “life history” to denote the ways organisms have evolved to be reproductively successful. And each life history strategy they recognize carries with it a reference to the potential contribution each individual will make to the next generation of its population and species. This is a major part of the evolutionary dynamic, and although it is not the whole story of change in the living world, it is omnipresent. A cultivated grapevine goes through several stages on its way to making grapes: bud break, flowering, fruit set, véraison, harvesting, leaf fall, dormancy. This life cycle is a somewhat artificial one, because it differs significantly from the natural cycle (in which there is no harvesting: the grapes interact instead with a variety of frugivores). But the vine remains in rhythm with the seasons.
A life history, however, is not just the story of an organism’s development, or even of its life cycle. Instead, a life history encompasses all the traits that are important for the organism’s reproduction and individual survival. Age at first reproduction, fecundity, and age at last reproduction are all important to biologists studying the evolutionary success of species. So how do grapevines manage, and how have their life history strategies contributed to their success?

One of the most important evolutionary problems any organism needs to solve, whether it’s a bacterium or an elephant, is how to create the next generation. Even viruses, not usually considered living organisms, are prolific and rapid reproducers. Prions—proteins that do not even have a genome—also contrive to replicate themselves copiously. And in the same spirit, the need to reproduce has shaped various parts of the grapevine. These plants spend most of their energy budget making leaves, seeds, rootstocks, and fruit. Why? Well, leaves and rootstocks are easy to understand. They are essential for the vine’s maintenance, converting the sun’s rays to useful energy and transporting nutrients throughout the plant. But how about the key component, the grapes that hold the seeds? They, too, represent a huge energetic investment, but they are on the vine for a different reason.
Reproduction is pretty straightforward for organisms that can walk, crawl, or slither in the interests of spreading their gametes around. Such creatures move hither and yon, hoping to find another with which to reproduce. But a plant can’t do that. Male plants have solved half of the problem posed by their immobility by packaging their gametes into tiny, light particles called pollen, and have come up with many ways to ensure that the pollen is spread around. The most spectacular of these involve coopting organisms that can move, analogous to yeast’s exploitation of wasps. Some plants are thus experts at attracting unsuspecting insects to do the heavy lifting. Others have taken a different tack, dispersing their seeds over long distances by means of tricks that allow their gametes to float on the air. Yet others have evolved a carnival sideshow–like half-woman/half-man strategy. Grapevines are among this last group.
The importance of such mechanisms is reflected in their sheer ingenuity. Charles Darwin put it this way in On the Various Contrivances by Which British and Foreign Orchids Are Fertilised by Insects (1862): “An examination of their many and beautiful contrivances will exalt the whole vegetable kingdom, in most persons’ estimation.” He went on to explain the contrivances of what he clearly saw as the most ingenious orchid of all, the genus Catasetum. Also known as “Darwin’s bee trap,” this orchid has a hair-trigger organ near the entry of the male flower. When tripped by a bee, the trigger shoots out a dart with pollen attached to its end, at a speed of more than 300 centimeters per second. The dart, a “pollinium,” sticks to the back of the bee, which will then deliver the pollen to the ova of a female Catasetum on its next visit. Domesticated grapevines keep things simpler, since most of them are hermaphroditic and are thus able to pollinate themselves. Insects and wind only rarely intervene.
Yet even when they can accomplish fertilization unaided, grapevines still need help in dispersing their seeds. Plant seeds can be scattered in several ways. One is by simply adhering to an animal that passes by. Anyone who has tramped through a field of high grass knows that you’re likely to come away with burrs sticking to your socks or pants. Those seed-containing burrs will either fall off or be picked off, and they’ll probably wind up on the ground some distance away from the parent, where they can carry out the business of reproduction. Another plant trick is dispersing the seeds through the air. Dandelions do this by attaching their fertilized seeds to a fluffy apparatus that can float in the breeze; maple trees have evolved a helicopter-like mechanism to disperse their seeds; and tumbleweeds have invented the wheel.
Still, perhaps the most popular long-distance dispersal strategy plants use is having their seed-containing parts eaten by animals. This strategy has dictated three of the more important aspects of grape anatomy—their heavily coated seeds, which can withstand the rigors of the stomach acids and intestinal enzymes to which they will be subjected before they are excreted; their color, which will catch the eye of potential dispersers; and their sugary innards, which given them an appealing taste. Thus, the biochemistry that makes grapes sweet came about because the vine evolved to bear fruit that would be attractive to potential dispersers. And it’s a competitive world out there—lots of plants are designed to disperse their seeds this way, usually at around the same time. So the fruit of the vine needs to be both as eye-catching and as tasty as possible.
This may explain why grapes are often red. Recent experiments suggest that birds, at least, greatly prefer red things to blue, yellow, green, or black ones. Researchers have demonstrated this preference by removing newly hatched birds from their nests, raising them in isolation, and then offering them items of different colors. The birds showed a clear preference for red objects. Vines generate the red color found in their skins and pulp by producing anthocyanins, rather bulky molecules that belong in the flavonoid group. So effective are these pigments that they are used in the food industry to produce food colorings, and they have the possible additional advantage of being antioxidants, often touted by health advocates as helping to counteract damage to the tissues caused by electron-robbing free radicals. (There is now a medium-sized question mark hovering above the potential health benefits claimed for antioxidants as a group, but there are compounds in red wine—notably the phenol known as resveratrol—that may be associated with some degree of cardiovascular benefit.)
Scientists have understood the molecular pathway involved in producing anthocyanins for a long time. The chain of reactions involves many proteins, each of which has a specific job in molding the structure of the anthocyanin. In the past decade, researchers have also begun studying the genes controlling all this activity. Whenever a gene is responsible for a physical trait (such as red color in grapes), one of two processes can occur. The gene concerned can make a protein that has a direct, physical impact on the trait, in which case it is called a “structural gene.” Or it can act like a valve, regulating the production of the protein involved. Genes of the second type are called “regulators,” or “transcription factors,” because they regulate the amount of protein produced, as well as when and where it is made. By studying entire grape genomes, researchers in Portugal have determined that there are ten structural genes and five regulators involved in grape coloration, a complexity that explains the extraordinary range of tints and color densities found in wine.
One of the more surprising discoveries of recent studies is that structural genes tend not to vary much among related organisms, and are sometimes not even active in producing the traits with which they are usually associated. Rather, it is the regulatory genes that do a lot of the dirty work, and implement much of the variety we see in nature. Grape color is no exception. The major genes controlling the production of anthocyanins in grapes are called Myb, Myc, and WD40. Japanese investigators have shown that the light color of Koshu “pink grapes” is caused by lowered anthocyanin production due to a defective Myb gene. The Koshu Myb gene has two small extra domains (the 44 and 111 nucleotides of DNA) in one of the Myb genes, an alteration that apparently reduces the amount of anthocyanin in the grape skin.
As we have already mentioned, nobody has yet been able to produce a seedless grape that makes good wine, so the seeds buried within the grape must provide an essential element of the chemical complexity of concentrated red wines. But grape seeds nonetheless pose a bit of a dilemma for the winemaker. On the one hand, they are critical to the reproductive success of the wild vine, and their presence is associated with good wine. But they are also the waste products of the winery. Grape berries evolved to attract animals, who would eat them and disperse the seeds; but if the seeds are digested, they can’t be dispersed. It is thus necessary for the animal to excrete them intact, after carrying them away from the vine. Hence the seeds of grapes, and indeed those of any other plant using this mode of dispersal, have acquired hard coats to help them survive the journey through the masticatory and digestive tracts of large animals. These coats make grape seeds tough, but as a second line of defense they also contain some nasty chemicals that an animal consuming the fruit would prefer not to taste. And this, of course, poses a problem for winemakers.
Anthocyanin content analysis of Cabernet Sauvignon and Merlot. The amount and types of anthocyanins are shown in the bars in the diagram. Also shown on the graph are Koshu, Riesling, and Chardonnay; the two latter have insignificant anthocyanins, while Koshu has very few. “Mv,” “Pt,” “Dp,” “Pn,” and “Cy” are abbreviations for the various kinds of anthocyanins found in these wines. Redrawn and modified from Shimazaki et al., “Pink-colored Grape Berry Is the Result of Short Insertion in Intron of Color Regulatory Gene.”
The grape seed, as we saw in Chapter 4, is composed of an outer seed coat, an endosperm, and the embryo nestled in the center. The seed coat acts as armor during the hazardous journey through the digestive tract, while the embryo is the seed’s precious cargo. The intermediate endosperm is fleshy, and provides nourishment for the embryo until the seed decides to germinate. These three layers all contain unpleasant-tasting polyphenols, which can make up almost 10 percent of the seed’s volume. But many other compounds are present, too, some of them noxious, though not all. Indeed, grape seed extract is controversially promoted for a range of health advantages, and grape seed oil is an excellent frying medium because it will not burn until it reaches very high temperatures. But from the wine drinker’s point of view the key thing is that, for whatever reason, clarets, Chateauneufs, and Cabs would just not be the same without the seeds.
The interactions of grapes with the dispersers that eat them are easy to observe. This is why most of the initial thinking done about interactions among organisms was about animals and plants that are visible to the naked eye. But it is now some time since scientists began to discover the role of microbial life in infectious disease. Diseases such as childbed fever and other killers have decimated human populations since time immemorial, and studying the microbes that caused disease was an important step in advancing human health. But this is only part of the story. Around the turn of the twentieth century two microbiologists, Martinus Beijerinck and Sergei Winogradsky, realized that microbes were everywhere and affected many natural processes; they did not simply cause diseases. Winogradsky was the first to realize that microbes were responsible for the enrichment of soil with nitrogen, while Beijerinck was one of the first scientists to obtain cultures of agriculturally important bacteria and bacteria involved in plant ecosystems. For years, researchers interested in this fledgling science kept their day jobs, so to speak, and studied microbial ecology as a side interest. But the more that has been learned, the more important the microbiome has become to our understanding of its role in enabling the conduct of our daily lives. Each of us is crammed with microbes that are essential for innumerable bodily processes.
The major difficulty in studying microbes is that they can be seen only with powerful microscopes. And until the past decade, the amount of DNA and protein in a single bacterial cell was too small for microbial ecologists to study using the techniques at hand. These scientists had to resort to culturing microbes from environmental samples, and what they studied was whatever they could grow in a laboratory. If, for example, researchers wanted to study the microbes living on a grape, they would wash the grape in a solution of water or dilute salt, take the wash, and try to culture microbes from it. The problem was that many microbial species could not be cultured. In fact, even now scientists haven’t figured out how to culture between 95 and 98 percent of all bacterial species. As a result, other methods had to be invented to look at the microbes involved in the ecology of the grape.
One new approach capitalizes on the fact that DNA is not only a double-stranded molecule, but also a complementary one: a researcher with one strand of a double helix can figure out how the other is structured. Whenever there is a G (guanine) on one strand of the double helix, directly across from it on the other strand will be a C (cytosine). Likewise, if there is a T (thymine) on one strand, directly across from it will be an A (adenine). These stick together by means of chemical bonds (A with T, G with C). As we saw, DNA with the sequence GATCGATC on one strand will have CTAGCTAG on the other, and the Cs and Gs and the As and Ts will stick together, acting act like a zipper. If the double helical molecule is heated, it will start to unzip. And it will zip back up when it cools.
Now, imagine that our old friend the yeast Saccharomyces cerevisiae has a unique sequence in its genome such as GCATCATCGATCGAGCATGATCGCAGC. Somewhere in the yeast’s genome the complement to this sequence exists on one strand. If this sequence is mixed with DNA from a yeast cell, heated, and then cooled, that sequence will find its complementary sequence and stick to it. Next, imagine putting a little fluorescent molecule on the end of this sequence, and repeating the exercise. What will happen this time is that, as expected, the sequence will stick to the complementary sequence in the yeast cell. And where it sticks, a tiny bit of fluorescence will be visible, indicating that the cell has the marker sequence, and hence is a yeast cell. If we have a number of sequences that we know are unique to a particular organism, we can make as many DNA probes as we wish and connect them to different-colored fluorescent beacons.
This approach is called fluorescent in situ hybridization, or FISH, and it allows us to know what species are present in a particular microscopic field and how many there are. FISH is used both for clinical purposes and to identify bacterial and other microbial species in samples taken from nature. This colorful way of identifying microbial species can tell scientists what kinds and quantities of microbes live on a grape—or anything else—and give them an idea of the players in the ecological game of wine.
If we were to use FISH on a grape sample, we might get an idea of the kinds of microbes living on the skin of the grape or in the dirt around it. But we would see only the species we have probes for. How then do we “see” all of the microbial species on and around a grape? Researchers learned in the 1990s that from a spoonful of dirt or a swab from the outside of a grape, they could make DNA much as a human genetics lab makes DNA from blood. The only difference is that in the DNA from blood there would be a single genome (the subject’s), whereas in the spoonful of dirt or grape wash there would be the genomes of millions of microbes.
The mixture of DNA from the dirt or wash contains the genomes of all of the microbes in the spoonful. Since each piece of DNA in the sample comes from a particular species, the most logical procedure would be to sequence all the fragments of DNA in the sample. But prior to what scientists call next-generation sequencing (NGS: a total misnomer because we are seven years into “next” generation already), the process of obtaining DNA sequences was laborious and yielded little data for the effort expended. Only a small number of DNA sequences could be obtained from the sample—maybe between 500 and 10,000, a tiny proportion of the whole. These sequences could then be compared to a huge compendium of bacterial DNA sequence information, the ribosomal database (RDB). By matching the sequences from the sample with those from the RDB, a researcher would know which species had been sequenced in the dirt or grape wash.
Next-generation sequencing has upped the ante. Typically, it will yield between 400,000 and 10 million different microbe sequences. It also allows scientists to “see” the millions of microbes that can’t be cultured in the lab. What this means is that researchers are getting more complete pictures of an increasingly large number of different microbial communities—and there seem to be an incalculable number of such communities in the world. It also means that new kinds of microbes are being discovered every day in media such as dirt, pond scum, air, seawater, sewage, and even on and in the human body. And NGS has given vineyard scientists an unprecedented perspective on life processes on the surface of grapes, on the soil in which grapevines grow, and in the grape must itself.
The first big task of the grape microbial community biologist is to identify the species involved—basically, to do a broad census. Three major kinds of microbes are found living on the surface of grapes: filamentous fungi, yeasts, and bacteria. It is clear from microbial community studies that the particular species of all three kinds vary from strain to strain of grape; they also differ in presence or frequency from region to region. A study that counted microbes on Cabernet Sauvignon vines and berries showed that nearly half of the microbes on these plants consist of just ten major species, whereas the number of species found in a spoonful of dirt is generally more than a thousand. Equally intriguing is the finding that the community of microbes living on vine leaves is quite different from the community living on the grape skins.
Once the initial census is made, scientists can look at these communities in two major ways. First, they can ask what the differences are in the microbial communities living on different strains of grapes. This is important for an understanding of the extent to which a particular microbial community might be responsible for the characteristic taste of, say, a Cabernet Sauvignon. Second, researchers can ask how the microbial community changes on grapes of the same strain. The microbes on grapes change as the berries ripen, and there is a huge turnover of the major kinds of bacteria living on grapes at the beginning of the ripening season, as well as a strong component based on environmental and varietal factors. In a 2014 study using NGS, Nicholas Bokulich and colleagues at University of California, Davis, examined the dynamics of the microbial makeup on grape skins in northern California. They were interested in what factors were involved in the makeup of the bacterial communities on grapes and in grape must. This is an important question for viticulturists because it is a first step toward circumventing the damage done to grapes by bacterial species. If the interactions and communities had proved to be random, it would have complicated attempts to remedy bacterial infection. But by identifying the microbial community structure on grapes from the Napa and Sonoma Valleys and in the central coast region, the scientists showed that bacterial communities are nonandomly associated with grape must and vary by region, variety of grape, and other environmental factors.
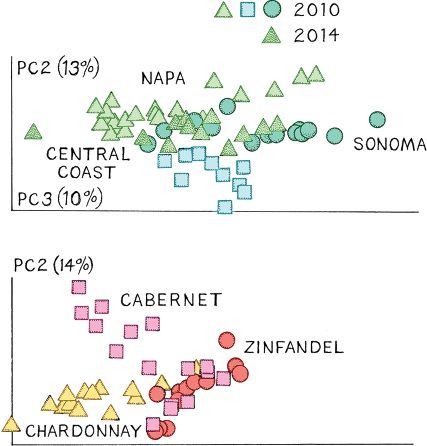
Graphs showing wine-growing region and grape variety correlated with bacterial community. The graphs were generated as multivariate statistical plots, where the axes become what are called principal components (PC) that explain the better part of the variation in the study. Note that the points cluster for the various shadings in both graphs, indicating that both geographic locality (above) and grape variety (below) are involved in determining the kind of microbes on the specific grapes. Redrawn and modified from Bokulich et al., “Microbial Biogeography of Wine Grapes Is Conditioned by Cultivar, Vintage, and Climate.”
Another study revealed that fungal species living on grapes often change depending on where the vine is in the vineyard, indicating a very fine spatial arrangement of microbes even within the same planting. Finally, it seems that the microbial communities in the same vineyard also vary from harvest to harvest, so clearly the interactions involved are incredibly complex. It remains for future research to explore the implications of these interactions for the production of wines having a particular desired character. But rest assured that this research will be done, with direct implications for the wine drinker.
Another unexpected discovery from this kind of mass census is that the winemakers’ yeasty friend Saccharomyces cerevisiae does not always flourish in vineyard environments. In fact, it rarely occurs naturally on the surfaces of harvested grapes, and is inoculated into them only shortly before crushing, after the wasps have intervened. The strain of yeast desired has to be added to the must, where it can dominate the fermentation process if it is present in adequate quantities. But it is not the only yeast that makes it into the must; wild yeast strains are also present. These uninvited guests may include undesired variants of S. cerevisiae that come from the vineyard or that are present in the winery as holdovers from earlier years when different strains were used for fermentation. They may come into contact with the must in a variety of ways, for instance as refugees on winemaking vessels or tools, or by transfer via insects or other animals.
Another category of “wild” yeasts includes species such as those from the genera Kloeckera, Brettanomyces, Candida, and Pichia. These yeast strains are often important components of wine fermentation, adding to the environmental individuality (terroir) of certain wines. Although such strains and species may do well in the vineyard, they are often less effective during fermentation because they lack high tolerance for alcohol. They also have low tolerance for sulfur dioxide, which is why many winemakers introduce this compound to the early stages of fermentation; they can get rid of the unwelcome yeasts before introducing Saccharomyces cerevisiae. Some winemakers, however, prefer to have the wild strains start the fermentation process. We say “start” because once the fermentation batch attains a 3–5 percent alcohol content the “wild” yeasts will often die, and the more alcohol-hardy S. cerevisiae will take over. Balancing the role of S. cerevisiae with the activity of wild strains is an important aspect of winemaking. Too little wild influence might fail to impart sufficient terroir to a wine, while too much might introduce contaminant compounds or undesired flavors. This delicate balance is an aspect of wine ecology that winemakers need to control carefully. Finally, we might note that small-scale winemakers sometimes make a point of using native yeasts, which should not be confused with wild yeasts, to produce their wines. Native yeasts might include the yeasts that are hanging around the winery and vineyards, but they might also be domestic strains that are traditional to a particular area.
The enormous microbial heterogeneity on grapes and in vineyards that recent research reveals is causing winemakers to think hard about their farming practices. One study showed that microbial community differences exist between organically farmed grapes and grapes grown the traditional way. Researchers have as yet merely scratched the surface of a huge and wide-ranging field of inquiry; as vine growers and winemakers increasingly use the new microbial community data, they will have an invaluable census of the microorganisms present and active in different kinds of grapes, in different vineyards, and at various stages of the winemaking process. This information promises to revolutionize vine growing and winemaking.
We hope we’ve convinced you that wine is the product of myriad interactions. Those interactions occur at many different levels. First and foremost, wine is the result of an interaction between chemicals and enzymes that leads to its color, nose, taste, and alcohol content. On another level, wine results from the interaction of different microbial species on and inside the grape and in the fermentation vessel. Wine also comes about as the result of the interaction of the parent vine with its environment, particularly with the other organisms living on and near it.
The life processes busily unfolding on the surface of a grape, and later in the grape must, are similar to the interactions going on in a busy industrial region like Elizabeth, New Jersey. Clusters of bacterial cells work together to take in raw materials and make products. The yeast in the must sit on the bottom of the tank and let carbohydrates come to them for processing. They open their bay doors and gather in tons of carbohydrates, which they then disassemble into their constituent parts. Carbon dioxide and alcohol arrive in the mix via the chemical pathways described in Chapter 3. The enzymes the yeast uses to produce the sugars and longer carbohydrates are like tiny machines on a factory floor, and they are continuously fed the raw materials from the grape must. All these interactions produce large amounts of exhaust and waste material, and require an immense amount of energy.
But that’s only part of the greater process. There are more than just carbohydrates in the mash. Because the grape skins, seeds, and some stems also get into the must, other little factories spring up in the filmy mats that form from yeast and bacteria. And many larger molecules, such as pigments and tannins, are delivered to those little factories for processing. If a molecule arrives at the wrong enzyme it is refused entrance and will move on to the next potential processing factory. These processes move on apace until the alcohol content of the mix reaches a certain concentration (usually about 15 percent), at which point the yeasts start to shut down. If the alcohol concentration goes too far above this level they will sicken and die. Accordingly, it is at around this point in fermentation that the winemaker moves on to the next stage of production, which involves racking the wine off the solids (pouring the liquid into a barrel and leaving the sediment behind) and, in the case of some reds, subjecting the wine to a secondary fermentation in which bacteria are used to convert astringent malic acid into softer lactic acid (as will be described in Chapter 11). When the wine arrives to rest in the barrel, the process still does not stop. The molecules in the wine interact with molecules emanating from the oak, and even when it is in the bottle in which it will be sold, the wine will continue to change.
Every one of these multifarious interactions, from enticing a bird to eat a grape to aging in the bottle, has combined with the others to make wine both possible to create and such a complex and rewarding product. The unique sum total of these interactions makes each wine you taste individual, different from every other in your experience. And the ultimate interactions are the many that the wine has with us, on its long and complicated journey from our noses to our brains.

