We must, however, acknowledge, as it seems to me, that man with all his noble qualities, still bears in his bodily frame the indelible stamp of his lowly origin.
—Charles Darwin, The Descent of Man
Throughout the second half of this book, we have documented example after example of transitional forms in everything from microfossils to mollusks to mammals. We could continue to do this for hundreds of pages more, but it wouldn’t really make a difference to the creationists or to those who are confused and misled by them. The only transition that really matters is, of course, the evolution of humans. Many creationists readily concede many of the examples we have just discussed as evolution as just variation within “created kinds,” although we have documented many examples of macroevolutionary changes that exceed anyone’s definitions of “kinds.” But to many people, those are just stories about critters. They only care about humans and whether or not we are specially created in God’s image or “just another ape.”
The idea that we might be related to apes was shocking when it was first proposed after On the Origin of the Species was published in 1859. Darwin deliberately avoided the subject in his already controversial book and finally dealt with it near the end of his life with The Descent of Man and Selection in Relation to Sex (1871). Thomas Henry Huxley, however, was not afraid of offending the sensibilities of the Victorians and boldly published Zoological Evidences of Man’s Place in Nature in 1863, with explicit diagrams showing the detailed skeletal similarities between humans and the great apes. Nevertheless, because of the strong religious beliefs of the nineteenth century, most people still refused to accept the idea. They were horrified to look into the mirror and see an image of themselves as an ape (fig. 15.1).

FIGURE 15.1. Satirical cartoon from Darwin’s time, posing the question that most British were struggling with: Are we an ape’s reflection? (By permission of the Trustees of the British Museum)
As the years have passed, however, the gulf between humans and the rest of the apes has narrowed considerably. Instead of the old “screaming hooting monkey” stereotype, we have discovered just how similar the apes are to humans. Decades of field research by pioneering anthropologists like Jane Goodall with the chimpanzees and the late Dian Fossey with the mountain gorillas have demystified these majestic creatures and surprised us with their amazing behavioral similarities to humans. Both chimpanzees and gorillas can learn sign language, communicate in simple sentences, and make and use simple tools. Their societies are very sophisticated compared with those of any other animal and show us many insights into the complexities of human societies as well. Over a century of research by hundreds of anthropologists has documented more and more connections between apes and humans. In nearly every other westernized country, polls show that a majority of educated people no longer object to the idea that humans and apes are related or at least have come to terms with the fact that humans are part of the animal kingdom and a part of nature as well, not above it.
Yet the idea is still offensive to many in the United States, where many polls show that a majority of Americans still do not accept that we are related to the apes. Nearly all of this is driven by strong religious beliefs, of course, plus common misconceptions about apes (not yet defused by all the documentary footage of amazing chimpanzees on television), and especially by a determined campaign of creationist misinformation. Still, it is surprising that more than 150 years after Darwin’s book was published, we still cannot come to terms with overwhelming evidence from both biology and the fossil record. My colleagues in the anthropology departments all over the country face this all the time. More than any other group of scientists, they are under attack. They have had to waste a lot of time undoing creationist mischief and clarifying ape and human evolution for the general public when they could be doing real research, discovering something new and useful instead.
Whatever one’s personal religious beliefs about humans and their “specialness,” as scientists we must stick to objective evidence and testable hypotheses. Many different religious beliefs have held many different concepts of humanity and its relation to God or the gods, but that cannot concern us here in a work of science. As discussed in chapter 1, science cannot and should not deal with the supernatural or with untestable hypotheses. Science cannot and should not deal with issues of the soul or other concepts that are important in the religious perspective but cannot be dealt with in a scientific fashion. This is not to say that the soul does not exist (science cannot decide one way or the other) or that humans don’t have some special element of God in them (again, not a scientific question). As long as we are talking about scientific evidence for any hypothesis (whether it is the evolution of rhinos or of humans), we have to stick by the rules of science and exclude supernatural hypotheses because they cannot be tested in a scientific manner. Of course, everyone is entitled to their own personal beliefs, but they are not entitled to impose them on others or to call their ideas scientific when they are clearly not.
And that is where we run into the most outrageous lies and distortions broadcast by the creationists. They are pretty bad at misleading people about the evolution of other animals, but when it comes to discussing the human fossil record, they hit rock bottom. To a creationist, every human fossil has to be discredited somehow because it goes against their very innermost beliefs to acknowledge the existence of these fossils. The hominin fossil record has improved enormously over the past several decades. We can spread out the impressive array of hominin fossils that connect us to the apes and to all of the rest of the animal kingdom (fig. 15.2) just to get a sense of the quality of the human evolutionary record. If these were any other fossils other than hominins, most people would be duly impressed and agree that the case was well established. Simply because they are our relatives, the stakes are much higher, and ideas and specimens are attacked that much more vigorously by creationists.

FIGURE 15.2. The hominid fossil record is now becoming very complete, contrary to creationist falsehoods. This table displays the huge collection of fossils of Australopithecus afarensis discovered in Ethiopia and described by Don Johanson and Tim White. In the foreground is the partial skeleton of “Lucy,” the oldest nearly complete hominid skeleton known. In the background is a selection of modern chimpanzee skulls for comparison, and a skeleton of Homo sapiens. If we were to lay out all the thousands of fossils of hominids in the cabinets of the Kenya National Museum, it would be even more impressive. (Photo courtesy D. Johanson, Institute of Human Origins)
Unfortunately, some anthropologists are unduly naïve and create all sorts of opportunities for misunderstanding and for creationist distortions. The study of human fossils is one of the most crowded and contentious of any scientific field I’ve ever seen. Although the human fossil record is now quite impressive and includes thousands of specimens (fig. 15.2), there are also thousands of physical anthropologists who must “publish or perish,” and who need to make a career somehow. Most of the best fossils are typically studied by those who have the funding and the access to the key sites in Africa and elsewhere, so the rest of the profession has to make their careers whatever way they can. Consequently, every idea and every specimen in hominin paleontology is challenged and restudied and reinterpreted many times, and any creationist who wants to mine for quotes can find some person (no matter how unqualified he or she might be) to say something that out of context seems to deny the validity of a given fossil.
Let us not forget that anthropologists are humans and make mistakes, too. Like all people who are trying to find out new information, they sometimes get ahead of their data or their interpretations are colored by their expectations. Roger Lewin’s (1987) excellent book Bones of Contention: Controversies in the Search for Human Origins shows this side of anthropology very nicely, and I highly recommend it for anyone who wants to see what the profession is really like. Sometimes those biases can lead to serious mistakes. A good example is how a clever forger duped the British anthropological establishment with the Piltdown hoax, which fit their prejudices that humans evolved large brains first, and seemed to put Raymond Dart’s Australopithecus africanus out of the picture. In another case, Henry Fairfield Osborn got excited about a curious tooth that looked very like a hominin and published a premature report on “Nebraska Man” before he was corrected by other scientists.
The key point to all of this is that science is self-correcting. Individual scientists can make mistakes or be misled by their biases, but there are so many other hard-boiled skeptical scientific critics out there that mistakes are soon caught and corrected. The Piltdown forgery was eventually exposed when new scientific methods came along that could show its fraudulent nature, and it was already suspected as a fake before then because it was not consistent with the emerging fossil record from Africa.
“Nebraska Man” was not accepted by any other paleontologist except Osborn and was quickly corrected when better specimens appeared that showed it was the misleadingly worn tooth of a fossil peccary or javelina known as Prosthennops. (I’m currently working on a book on the fossil peccaries of North America, so I’ve gotten to know these fossils well). It is not a “pig” as creationists claim. Typical of their ignorance of most biological topics, creationists don’t know enough zoology to know that pigs and peccaries are separate families that evolved on different continents; they are only distantly related. In addition, Osborn’s mistake was not as ludicrous as the creationists try to make it appear. The tooth itself is very badly worn and apparently rotated in the socket, giving it a peculiar appearance. In addition, peccaries and pigs both have omnivorous diets and their teeth have squared crowns with low rounded cusps, very similar to another group of omnivores, the primates. I’ve surprised creationists on more than one occasion by putting the teeth of apes and peccaries side by side, and they can’t tell the difference.
A quick look at some of the creationist pamphlets and books shows just how misleading and dishonest their presentations are. Typical of the genre is the little pamphlet Big Daddy, published by creationist Jack Chick. It is an insidious cartoon for easily swayed young minds, with a professor proclaiming the facts about human evolution, then being “corrected” by a polite young Christian who shatters the professor’s ideas about science. The garish yellow centerfold features a “march of hominins” that we discussed in chapter 5, perpetuating the misconception that human evolution is a single linear sequence, not a branching bush of many species. And each example in the cartoon picks just one hominin fossil and attempts to discredit it. One by one, they distort the hominin fossil record or just plain make it up.
For example, they say of “Peking Man” that “all evidence has disappeared.” Not true! The original specimens of “Peking Man” (the Homo erectus specimens from Zhoukoudian cave near Beijing, not a distinct species) were lost when the Chinese and Americans were fleeing with the fossils during the Japanese invasion in 1939. But many good casts were made of the original material, and many more new and better specimens have been found in subsequent excavations. Creationists point to Piltdown man, because this forgery was accepted for a while. Of course, it was scientists (not creationists) who eventually discovered the forgery. “Nebraska Man,” as we outlined already, was the mistake of one scientist and was corrected within a year—and it is not the tooth of a pig! Creationists claim that Neanderthal man is just based on an arthritic skeleton. Not true! One famous specimen of Neanderthal was indeed arthritic, and its malformities influenced some early reconstructions, but there are now dozens of normal, undiseased specimens, and they clearly show that Neanderthals represent a distinct species that is not modern Homo sapiens. They were much more robust and heavy limbed than we are, with a brain larger than ours but with a distinctive flatter skull with large brow ridges, protruding face, no chin, and a bulge in the back of the skull (see fig. 15.6). The mysterious “New Guinea Man” is something that only appears in creationist publications. No legitimate anthropologist has ever made the claim that they are anything but modern Homo sapiens. Finally, Cro-Magnon was always considered to be modern Homo sapiens, so the creationist attempt to suggest otherwise is deceptive and misleading.
More importantly, these eight examples are used to discredit the entire human fossil record, yet every other human species that they cannot discredit (of which there are now dozens) is conveniently not mentioned—an obvious attempt at trickery and distortion. This brief summary of how they managed to get every single example in their attack on hominin fossils completely wrong is representative of their tactics and of the abysmal level of their understanding of human fossils. Any minor mistake, or any account that seems to discredit a fossil, is good enough to be perpetuated over and over again. These same examples pop up in nearly every other creationist publication and website as well, often cribbed word for word, complete with all the same mistakes and misspellings.
There is no point in wasting more space in this chapter to correct every single creationist lie about the hominin fossil record. The example given above is very typical, and I have not seen anything more sophisticated in any of their other publications. Just as is the case with all the other creationist misstatements we have already discussed, their tactics usually include quoting out of context, quoting old outdated sources that don’t reflect the modern knowledge of the fossils, or quoting scientists who were considered cranks even when they were active (such as Solly Zuckerman, Gish’s personal favorite). Creationists don’t do any legitimate peer-reviewed anthropological research themselves nor do they bother to actually work with the fossils or even learn the basics of anthropology and human anatomy to see what the fossils really look like. They just do book reports and pull quotes out of context; that’s as far as their scientific curiosity goes. Once they’ve found what they think is a damaging quote, either they don’t bother to find out the context or they deliberately mislead the reader about the entire context of the quotation. This may be a sneaky way to confuse people and win them to their cause, but it is dishonest, unethical, and unscientific.
So let’s stop wasting our time with the distorted creationist view and briefly review what the hominin fossil record does show.
It is…probable that Africa was formerly inhabited by extinct apes closely allied to the gorilla and chimpanzee, and as these two species are now man’s nearest allies, it is somewhat more probable that our early progenitors lived on the African continent than elsewhere.
—Charles Darwin, The Descent of Man
When Darwin published On the Origin of Species in 1859, there were still no good hominin fossils for him to point to, and this was still true when he wrote The Descent of Man in 1871. Although the first Neanderthal specimen was known, it was usually misinterpreted as the skeleton of a diseased Cossack who had died in a cave and didn’t figure in the early ideas about human evolution. The first genuine hominin fossil that was truly different from us was Eugène Dubois’s famous “Java Man” specimens of Homo erectus, originally described as Pithecanthropus erectus in 1896. As outlined by Swisher et al. (2000), the specimens were controversial and misinterpreted for many years because they were so incomplete (just a skull cap, a thigh bone, and a few other fragments) and did not fit the biases of anthropology at the time. On top of that, Dubois’s own paranoid behavior made his ideas harder to accept. But Raymond Dart’s 1924 description of the South African skull known as the “Taung Child,” Australopithecus africanus, was the first good fossil hominin species that was not a member of the genus Homo. With its discovery almost 100 years ago, it should have clinched the case that humans had evolved and did so in Africa. Nevertheless, the hominin fossil record (especially of our earlier ancestors, who all lived in Africa) was still quite poor for several decades after 1924 because most anthropologists focused on very young European material and could not accept Darwin’s insight that our closest ape relatives lived in Africa, and therefore we probably originated there, too.
Contrary to what creationists say, the fossil record of humans is no longer as poor as it was even 40 years ago. Decades of hard work in the field by hundreds of scientists has turned up thousands of hominin fossils (fig. 15.2), including a few good skeletons and many good skulls that show clearly how humans have evolved over 7 million years. This avalanche of new discoveries every year has occurred despite the handicap that hominin fossils are delicate and rare, and only one or two are found for every hundred fossil pig or fossil horse specimen found in the same beds in eastern Africa. A tour through the bomb-proof hominin vault in the Kenya National Museum in Nairobi is a revelation: a whole room full of fossils that document our evolution, and whose existence the creationists must deny. Many other museums in Africa, Europe, and Asia have similar large collections of our early ancestors, so there are lots of fossils to work with now (and even more ideas about how to interpret them, which is a good thing in science). Fortunately, there are now books such as Eric Delson’s (1985) Ancestors: The Hard Evidence, which was once part of a traveling museum exhibit. These books and exhibits allowed the public to see these precious fossils up close for the first time and realize that the creationist view of the human fossils record is a lie. Since then, anthropologists have been much better at displaying and publicizing the quality of the hominin record and demonstrating that they do not have any skeletons hiding in their closets. Now there are a number of beautifully illustrated books with excellent full-color photographs of the actual specimens published (listed at the end of the chapter), and there is no longer any excuse for someone who wants to see what the fossils really look like.
The entire story of human evolution is too long and detailed to be discussed in a single chapter, so we will just touch upon the highlights. The short version is this: dozens of human species and genera are now known, forming a very bushy family tree that spans almost 7 million years of human evolution, mostly in Africa (fig. 15.3). The exact details of how all these fossils should be named or how they are interrelated is always controversial because many of the specimens are incomplete, and anthropologists are famous for being argumentative and contentious. But no matter how the arguments swing from year to year, the amazing quality of the hominin fossil record is an objective fact, not someone’s interpretation or guesswork.
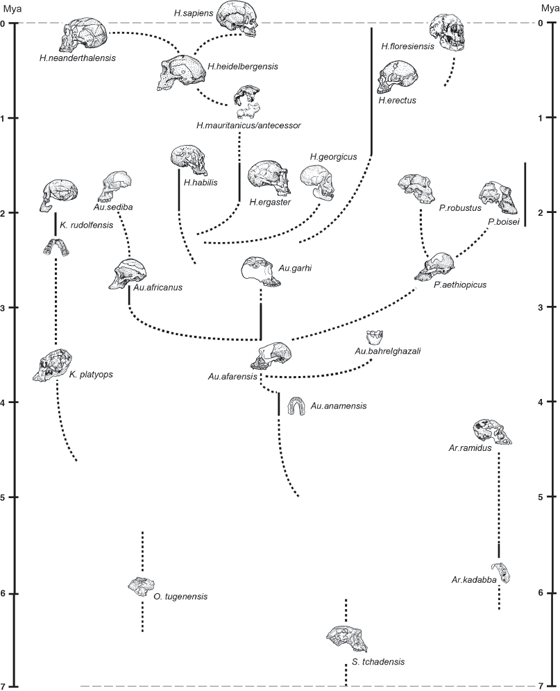
FIGURE 15.3. The current family tree of hominids, showing the extremely bushy branching pattern of human evolution, with many different fossil species at the same time at some time intervals. Sketches of the best specimens of each species are shown next to their time ranges. (Courtesy I. Tattersall)
First, let us place humans in a broader context. We are members of the order Primates, the group that includes not only ourselves and the great apes but also the Old World monkeys (Cercopithecidae), New World monkeys (Cebidae), and lemurs, lorises, bush babies, pottos, and many other archaic primates still alive today (fig. 15.4). We can trace the fossil record of most of these lineages back to the Cretaceous and Paleocene primate Purgatorius, whose name has interesting and ironic religious implications. (It was so named from Purgatory Hill in the Hell Creek Formation of Montana, where it was found). In the early Cenozoic, the globe was much warmer and more densely vegetated, and there was a huge diversity of different archaic primates distantly related to lemurs and other “prosimians,” including the plesiadapids, adapids, omomyids, and many other groups (see Beard 2004, for an outstanding account). If you collect fossils in Paleocene or early Eocene beds in places such as the Bighorn Basin of Wyoming, primates are one of the most common fossils, and enormous collections of their jaws and teeth are now stored in museums around the world. But as world climate became cooler and drier in the Oligocene and the forests vanished, primates became scarce, too. They vanished from North America and Europe, where they had once flourished, and became restricted to Southeast Asia and Africa. During the Oligocene, one group of primates, the Platyrrhini or New World monkeys, managed to make the crossing of the South Atlantic, where they radiated into the great diversity of prehensile-tailed monkeys including spider monkeys, colobuses, and howler monkeys, marmosets, and the like. Most primate evolution was confined to Africa, where the Old World monkeys (baboons, macaques, rhesus monkeys, and their relatives) flourished, alongside the earliest members of the ape lineage (Aegyptopithecus, Propliopithecus, Apidium, among others), which are documented from the Oligocene Fayûm beds of Egypt.
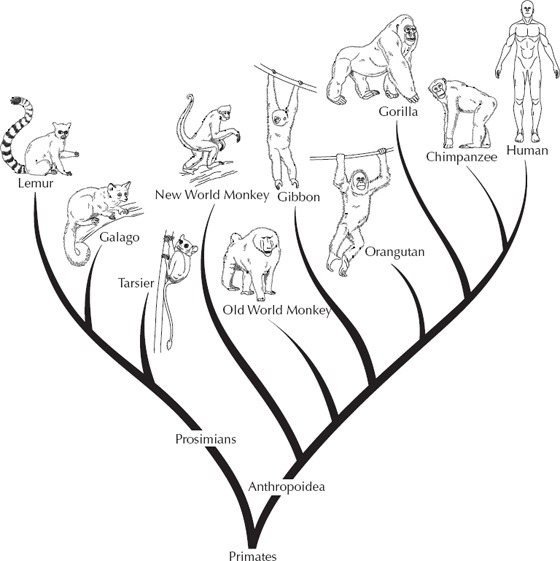
FIGURE 15.4. Family tree of the major groups of living primates, showing the close relationships between the great apes and humans and the more distant relationships of the Old World monkeys, New World monkeys, and lemurs, lorises, and bush babies.
In the Miocene, apes were more diverse and successful than the monkeys and occasionally were found in Europe as well as their African homeland. By the middle Miocene, primitive members of the orangutan lineage (Sivapithecus) are known from deposits dated at 12 million years old in Pakistan, giving us some of the first ape fossils that belong to modern lineages. Unfortunately, we have lots of fossil apes but no clear-cut fossils of the chimpanzee or gorilla clan just yet, probably because both of these apes have always lived in forests where there is scant chance of fossilization. Most of the fossil apes declined and died out by the end of the Miocene, and Old World monkeys came to dominate the primate adaptive zones ever since then.
The oldest specimen that can be truly described as a member of our own family was discovered and described only a few years ago. Nicknamed “Toumai” by its discoverers, its formal scientific name is Sahelanthropus tchadensis. The best specimen is a nearly complete skull (fig. 15.5A) from rocks about 6–7 million years in age from the sub-Saharan Sahel region of Chad (Brunet et al. 2002). Although the skull is very chimp-like with its small size, small brain, and large brow ridges, it had remarkably human-like features, with a flattened face, reduced canine teeth, enlarged cheek teeth with heavy crown wear, and an upright posture at the very beginning of human evolution. Just slightly younger is Ororrin tugenensis, from the upper Miocene Lukeino Formation in the Tugen Hills in Kenya, dated between 5.72 and 5.88 million years ago. Ororrin is known mainly from fragmentary remains, but the teeth have the thick enamel typical of early hominins, and the thigh bones and shin bones clearly show that it walked upright. Slightly younger still are the remains of Ardipithecus ramidus kadabba, found in Ethiopian rocks dated between 5.2 and 5.8 million years ago. These consist of a number of fragmentary fossils, but the foot bones show that hominins used the “toe off” manner of upright walking as early as 5.2 million years ago. Thus our human lineage was well established by the latest Miocene and fully upright in posture, even though our brains were still small and primitive and our body size not much different than that of contemporary apes.

FIGURE 15.5. Some of the better known fossil hominid skulls from Africa. (A) The oldest known hominid, Sahelanthropus tchadensis, nicknamed “Toumai,” from beds 6–7 million years old in Chad. (From Brunet et al. 2002; by permission of the Nature Publishing Group) (B) The famous “Black Skull,” KNM-WT 17000, a specimen of the very primitive robust australopithecine, Paranthropus aethiopicus, discovered in beds 2.5 million years in age on the west shore of Lake Turkana. (Photo courtesy A. Walker) (C) Mary Leakey’s “Nutcracker Man,” the super-robust Paranthropus boisei, originally named “Zinjanthropus” boisei, OH5. It was recovered from 1.8-million-year-old rocks in Olduvai Gorge, Tanzania. (Photo courtesy National Museums of Kenya) (D) Richard Leakey’s most famous discovery, the best skull of a primitive member of our own genus, Homo rudolfensis (originally referred to as Homo habilis), KNM-ER 1470, from beds dated at 1.88 million years old on the eastern shore of Lake Turkana. (Photo courtesy National Museums of Kenya)
The Pliocene saw an even greater diversity of hominins (fig. 15.3), with a number of archaic species overlapping in time with the radiation of more advanced hominins. Archaic relicts of the Miocene included Ardipithecus ramidus ramidus, found in Ethiopia in 1992 from rocks 4.4 million years in age, which had human-like reduced canine teeth and a U-shaped lower jaw (instead of the V-shaped lower jaw of the apes). Nearly complete skeletal material of this species has now been discovered and reported by Tim White and his colleagues, making it the oldest known fossil hominin skeleton. Rocks in Kenya about 3.5 million years in age yield primitive forms like Kenyapithecus platyops. By 4.2 million years ago, however, the first members of the advanced genus Australopithecus, the most diverse member of our family in the Pliocene, are also found. The oldest of these fossils is Australopithecus anamensis from rocks near Lake Turkana in Kenya ranging from 3.9 to 4.2 million years in age. These creatures were fully bipedal, as shown not only by their bones but also by hominin trackways near Laetoli, Tanzania. The most famous of these early australopithecines is A. afarensis (from rocks 3.0–3.4 million years ago near Hadar, Ethiopia), better known as “Lucy” by its discoverers Don Johanson and Tim White (figs. 15.2 and 15.6A). When it was discovered in the 1970s, Australopithecus afarensis was the first early hominin to clearly show a bipedal posture (based on the knee joint and pelvic bones) but was not as upright as later hominins. These were still small creatures (about 1 meter or 3 feet tall) with small brains, and very apelike in having large canine teeth and a large overhung jaw.

FIGURE 15.6. Two of the oldest known relatively complete hominid skeletons. (A) The skeleton known as “Lucy,” the nearly complete specimen of Australopithecus afarensis from rocks 3.4 million years old in Ethiopia. (Courtesy D. Johanson, Institute of Human Origins) (B) The famous “Nariokotome Boy,” KNM-WT 15000, a nearly complete skeleton of Homo ergaster (formerly referred to Homo erectus), from rocks 1.6 million years old on the western shores of Lake Turkana, Kenya. (Courtesy A. Walker)
By the late Pliocene, hominins had become very diverse in Africa (fig. 15.3). These included not only the primitive forms Australopithecus garhi (dated at 2.6 million years) and A. bahrelghazali (dated at 3.4 million years) but also the best-studied australopithecine, Australopithecus africanus. Originally described by Raymond Dart in 1924 based on a juvenile skull (the “Taung Baby”), for decades the Eurocentric anthropology community refused to accept it as ancestral to humans. But as more South African caves yielded better specimens to paleontologists like Robert Broom (especially the adult female skull known as “Mrs. Ples”), it became clear that Australopithecus africanus was a bipedal, small-brained African hominin, not an ape. This went contrary to all the accepted notions, which postulated that human evolution was driven by brain size, that bipedalism was secondary, and that it had occurred in Europe or Asia. The Piltdown forgery was deliberately set up to reinforce this bias, but by the 1950s, when Piltdown was exposed as a fraud, the evidence from Australopithecus africanus became undeniable. Australopithecus africanus was a rather small, gracile creature, with a dainty jaw, small cheek teeth, no skull crest, and a brain only 450 cc in volume. On the basis of its gracile and very human-like features, Australopithecus africanus is also the best candidate for ancestry of our own genus Homo.
In addition to Australopithecus africanus, the late Pliocene of Africa also yields a number of highly robust hominins. For a long time, they were lumped into a very broad concept of the genus Australopithecus, either as distinct species or even dismissed as robust males of Australopithecus africanus. In recent years, however, paleoanthropologists have come to regard them as a separate robust lineage, now placed in the genus Paranthropus. The oldest of these is the curious “Black Skull,” discovered in 1975 by Alan Walker and his crew on the shores of West Lake Turkana, Kenya, from rocks about 2.5 million years in age (fig. 15.5B). Although its brain is small, and it would have had a small body as well, the skull is robust with large skull crest and massive molars and an advanced dish-shaped face. Currently, scientific opinion places the Black Skull as the most primitive member of Paranthropus, P. aethiopicus. It was followed by the most robust of all hominins, P. boisei, from rocks in East Africa ranging from 2.2 to 1.2 million years in age (fig. 15.5C). This fossil was nicknamed “Nutcracker Man” for its huge thick-enameled molars, robust jaws, wide flaring cheekbones, and strong crest on the top of its head, indicating a diet of nut or seed or bone cracking. Originally found by Mary Leakey at Olduvai Gorge in 1959, it was named “Zinjanthropus boisei” by Louis Leakey, who made his reputation from it. The rocks of South Africa between 1.6 and 1.9 million years in age yield the type species of Paranthropus, P. robustus. These too had massive jaws, large molars, and large skull crests but were not as robust as P. boisei. Paranthropus robustus lived side by side in the same South African caves as A. africanus. It was not only more robust but also larger than that species as well, with some individuals weighing as much as 150 pounds.
Finally, the latest Pliocene saw the first members of our own genus Homo, which are easily distinguished from contemporary Australopithecus and Paranthropus by a larger brain size, flatter face, no skull crest, reduced brow ridges, smaller cheek teeth, and reduced canine teeth. The first of these to be described was Homo habilis (“handy man”), discovered in the 1960s by Louis and Mary Leakey in Olduvai Gorge, Tanzania, from beds about 1.75 million years in age. Originally, all of the early Homo specimens were shoehorned into the species H. habilis, but now paleoanthropologists recognize that this material is too diverse to belong to one species, so several are now recognized. These include the very modern-looking skull (fig. 15.5D) of H. rudolfensis (from beds ranging from 1.9 to 2.4 million years in age), which made Richard Leakey’s reputation, and the very advanced but short-lived Homo ergaster (fig. 15.6B), from beds 1.6–1.8 million years in age. These species are known not only from bones but also from their primitive tools, choppers and hand axes of the “Olduwan culture.”
Many of these archaic Pliocene taxa persisted into the early Pleistocene (as recently as 1.6 million years ago), including Paranthropus robustus and P. boisei, Homo ergaster, and H. habilis (fig. 15.3). The best-known fossil of H. ergaster is a nearly complete skeleton found on the shores of West Lake Turkana in 1984, and known as “Nariokotome Boy” (fig. 15.6B), which would have been 2 meters tall if fully grown.
By 1.9 million years ago, however, a new species had appeared: Homo erectus (fig. 15.7). This creature was not only bipedal and erect (as its species name implies) but also almost as large in body size as we are. Some individuals reached 190 centimeters (6 feet) in height. Its brain capacity was about 1 liter, only slightly less than ours. Like earlier species of Homo, they made crude choppers and hand axes (“Acheulian culture” tools) and certainly knew how to make and use fire and how to construct stone and wooden dwellings and small villages. Originally, Homo erectus was confined to Africa, where all of our other ancestors had long lived. By around 1.8 million years ago, we have evidence that H. erectus finally escaped the African homeland, for specimens from Indonesia (originally described as “Pithecanthropus erectus” or “Java Man”) have been dated at that age (Swisher et al. 2000), and specimens are also known from elsewhere in Eurasia, such as Romania, that are almost as old. By about 500,000 years ago, we have abundant fossils of H. erectus in many parts of Eurasia, including the famous specimens from the Chinese caves at Zhokoudian, known as “Peking Man” and dated as old as 460,000 years ago. The latest dating suggests that H. erectus may have persisted as recently as 27,000 years ago, outlasting even the Neanderthals and overlapping with modern H. sapiens (Swisher et al. 2000). Homo erectus was thus the first member of our family to live outside Africa, and it roamed through the entire Old World (except Australia and the glaciated regions). Homo erectus was not only the first widespread hominin species but also one of the most successful and long-lived species, spanning more than 1.8 million years in duration between 1.9 and 0.03 million years ago. During most of that long time, it was the only species of Homo on the planet and changed very little in brain size or body proportions. If longevity is a measure of success, then it could be argued that it was even more successful than we are.
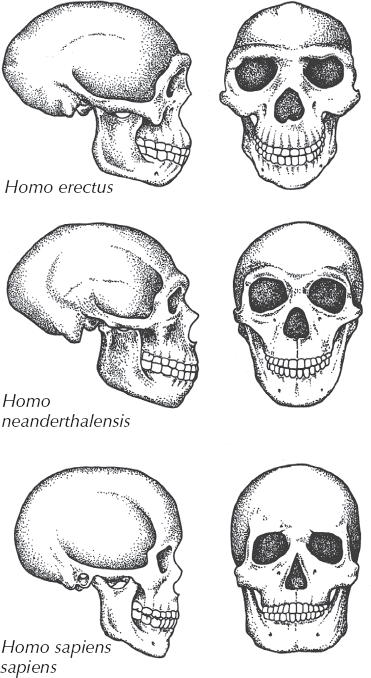
FIGURE 15.7. Comparison of the skull of Homo erectus (including specimens known as “Java Man” and “Peking Man”), Neanderthal, and modern Homo sapiens. Homo erectus had a brain capacity about half of ours, much heavier brow ridges, and a more protruding face. Yet it had upright posture and an essentially modern skeleton; it was clearly a member of our genus, and not “just an ape” as creationists allege. On the other hand, it is clearly not a modern human, but a transitional form between australopithecines and ourselves. (Drawing by Carl Buell)
By about 300,000 years ago, another species was established in western Europe and the Near East: the Neanderthals. These were the first fossil humans to be discovered, although they were originally dismissed as the remains of diseased Cossacks that had died in caves. The first complete descriptions of skeletons were based on a specimen that suffered from old age and disease, so for decades Neanderthals were thought to be stoop-shouldered and primitive, the classic stereotypical grunting “cavemen.” Modern research (Stringer and Gamble 1993) has shown that Neanderthals were very different from this stereotype. Although their skulls are distinct from ours in having a protruding face, large brow ridges, no chin, and a flatter skull that sticks out in the back (fig. 15.8), they had a slightly larger brain capacity than we do, and they practiced a complex culture. The famous discoveries at Shanidar Cave in Iraq showed that Neanderthals buried their dead with elaborate religious rituals and even flowers, suggesting that they had at least some kind of religious beliefs and possibly belief in an afterlife. Their bones (and presumably bodies) were robust and muscular and slightly shorter than the average modern human, but they also lived exclusively in the cold climates of the glacial margin of Europe and the Middle East, where their stocky build (similar to a modern Inuit or Laplander) would be an advantage. Their tool kits and culture were also more complex, with Mousterian hand axes, spearheads, arrowheads, and other complex devices, as well as bone and wooden tools. Some of these tools show complex working and simple carving, so they were artistic as no hominin before had ever been.

FIGURE 15.8. Comparison of the profiles of a Neanderthal and a modern Homo sapiens. Although Neanderthals had brains slightly larger than ours, their skulls were shaped very differently, with a much flatter cranium that protruded in the back, prominent brow ridges, a bulging face, and no real chin. Their skeletons were also distinctive, with much more heavy robust bones than any living human. Contrary to creationist claims that they are just deformed modern humans, they are very distinctive and clearly not a member of our species. (Drawing by Carl Buell)
For decades, anthropologists treated Neanderthals as a subspecies of Homo sapiens, but recent work suggests that they were a distinct species. The best evidence of this comes from the Skhul and Qafzeh caves in Israel, where layers bearing Neanderthal remains are interbedded and alternate with layers containing early modern humans. In addition, Neanderthals appeared later than the earliest archaic Homo sapiens, so they could not be our ancestors but rather an extinct European side branch. Recently their DNA has been sequenced and they are clearly not Homo sapiens, but now there is some evidence that all modern humans have a bit of Neanderthal DNA in them, so there must have been some interbreeding between the two species. DNA analysis of just two tiny bones from Denisova Cave in Siberia has revealed the existence of yet another species of humans, the Denisovans, who are poorly known at this time.
Finally, we see the first fossil skulls and skeletons that look almost indistinguishable from our own species. Some of these “archaic Homo sapiens” are known from deposits in Africa dating as far back as 500,000 years ago. About 90,000 years ago, skulls from Africa (such as Klasies Mouth Cave in South Africa) are almost completely modern in appearance and are universally regarded as Homo sapiens sapiens (our species and subspecies). Like Homo erectus, early Homo sapiens spent most of its history in Africa, and finally migrated to Eurasia about 45,000 years ago. There it came into contact with Neanderthals, and for about 9,000 years they coexisted. Mysteriously, Neanderthals vanished 36,000 years ago. Whether they were wiped out by Homo sapiens or by some other cause is highly controversial. Whatever happened, modern Homo sapiens soon took over the entire Old World, developing complex cultures (the “Cro-Magnon people”) that produced the famous cave paintings of Europe and many kinds of weapons and tools.
This brief review of the hominin fossil record hardly does justice to the richness and quality of the specimens or to the incredible amount of anatomical detail that has been deciphered. If it all seems like too much to absorb, just gaze at the faces of the skulls in the Fig. 15.5. They look vaguely like modern human skulls, but they definitely show the progression from more primitive hominins that creationists want to call “mere apes” (even though they were all completely bipedal and had many other human characteristics) up through forms that creationists want to call “modern humans” (even though they had many distinctive anatomical features, like those found in Neanderthals, that make them a distinct species). Anyone without advanced anthropological training and without biases or prejudices can glance at these fossils and see the hallmarks of human ancestry.
The next time you visit a zoo, make a point of walking by the ape cages. Imagine that the apes had lost most of their hair, and imagine a cage nearby holding some unfortunate people who had no clothes and couldn’t speak but were otherwise normal. Now try guessing how similar those apes are to us in their genes. For instance, would you guess that a chimpanzee shares 10 percent, 50 percent, or 99 percent of its genetic program with humans?
—Jared Diamond, The Third Chimpanzee
If the fossil record of human evolution was not proof enough, the clinching evidence is found in every cell in your body. When molecular biologists first began comparing the molecules from apes, monkeys, and humans back in the 1960s, they made a startling discovery: we are extremely similar to chimps and gorillas at the molecular level. The first evidence of this came when Vince Sarich’s famous experiments showed by the relatively crude method of comparing immune responses that there was little difference between the genes of chimps and humans. Then the DNA-DNA hybridization method came along and got better results still. This technique took the DNA of chimps and humans and separated the strands in a solution by warming it up. As the solution cooled down, the individual unpaired strands of DNA link up again, with some combining a strand of chimp and a strand of human DNA. When you heat up that hybrid DNA again, it unzips and separates. The more genes the two strands have in common and the more similar they are, the harder it is for them to unzip, so that the temperature of separation is directly proportional to the number of genes in common.
When these results were first published by Sibley and Ahlquist in 1984, they caused a shock in the scientific community. It turns out that human and chimpanzee DNA is 97.6 percent identical! Less than 3 percent of our DNA is different from that of a chimpanzee. Likewise, our DNA shares about 96 percent of its genes with gorillas, and it is 94.7 percent identical with that of gibbons, 91.1 percent identical with that of rhesus monkeys (an Old World monkey, of family Cercopithecidae), 84.2 percent identical with that of the capuchin monkey (a New World monkey, of family Cebidae), but only 58 percent identical with that of a primitive “prosimian” such as a galago. Not only did the sequence of similarity match exactly with the branching sequence of primates (fig. 15.4), which made sense, but the real shock was just how similar our genes are to those of most of the great apes. Molecular biologists pointed out that the genetic similarity between humans and chimps was closer than between any other two species they had studied, such as two closely related species of rats or two kinds of frogs. Since these experiments, the actual sequence of the mitochondrial DNA and the nuclear DNA have been determined for both chimps and humans, and the results are the same (within a fraction of a percent). There’s no two ways about it. The chimps are our “kissing cousins” among the animal kingdom, just a few genes removed from being fully human.
Jared Diamond puts it all in perspective with an interesting analogy. Suppose you were a molecular biologist from another planet and you were given just DNA samples of humans and the two chimpanzee species (the common chimp, Pan troglodytes, and the pygmy chimp or bonobo, Pan paniscus), plus a sampling of DNA from the primates and other groups of animals. You sequence the genes, plot the results, and conclude that humans are just a third species of chimpanzee, genetically closer than just about any other three species of animals on the planet. For example, the genetic similarity between lions and tigers is only 95 percent, yet they can even interbreed in zoos. Without seeing the differences in our bodies and behavior, the conclusion is unavoidable: we are very closely related.
What do we make of this? Although there are many different ideas suggesting what is going on here, the basic idea is that the 1–2 percent of the genome that differentiates us from chimps must be the regulatory genes that turn on and turn off the structural genes (which make up most of the 97.6 percent that is the same). We have the genes for most parts of the ape body, and the monkey body too, and every once in a while there is a genetic mistake or atavism, and humans express the long-repressed genes that we still carry to make a tail (fig. 15.9).
FIGURE 15.9. Every once in a while, a human is born with an atavistic tail, a throwback to our evolutionary past, when the regulation that normally shuts down our genes for tails fails to operate. The human tail comes complete with fully developed vertebrae, muscles, and other features of animal tails. (A) X-ray of a human with well-developed tail vertebrae. (B) Image of two humans with fully developed tails. (From Bar-Maor et al. 1980; used with permission of the Journal of Bone and Joint Surgery)
In fact, since the 1920s, many biologists and anthropologists have argued that much of what differentiates us from the chimpanzee is neotenic retentions of juvenile ape characters. If you look at a juvenile chimpanzee (fig. 15.10), its skull is much like that of a human, with a large brain, small brow ridges, short snout, and upright posture. Then, during development to an adult, the chimpanzee develops the larger snout with long canines, big brow ridges, and forward slouching posture of the head. If regulatory genes tweak our embryonic development a tiny bit, we can make most of the characteristics that mark us as human just by becoming juvenile apes that reach sexual maturity without every truly growing up.

FIGURE 15.10. (A) Juvenile chimpanzees have many characteristics of the skull found in adult humans, including upright posture, relatively large brain, small brow ridges, and a less protruding snout. (B) As they grow into adult chimps, these features all become more apelike. Since the 1920s, many anthropologists have argued that much of what makes us human is retention of juvenile ape characteristics into adulthood (neoteny). (From Naef 1926)
You may have read some of the fascinating works of Aldous Huxley, the famous novelist and author of the dystopian classic Brave New World (a high school reading list favorite). He was also the brother of the famous evolutionary biologist Julian Huxley, and both were grandsons of Darwin’s “bulldog,” Thomas Henry Huxley. Aldous knew these ideas about human neoteny very well because of his brother’s influence. In 1939, he published a novel entitled After Many a Summer Dies the Swan. The theme of the novel is immortality, and how humans are always striving to find a way to extend their lives beyond what nature intended. The main character is a millionaire (modeled after William Randolph Hearst, whom Huxley met when he was a Hollywood screenwriter in the 1920s) named Jo Stoyte, who is attempting to live forever by hiring a classic “mad scientist,” Dr. Obispo, to do research on extending his life. Dr. Obispo discovers that the third earl of Gonister in England had lived several centuries without any signs of aging, apparently by ingesting carp guts. Archival records showed that he had fathered children when he was over 100 years old. (This is a science fiction novel, remember!). Dr. Obispo seduces the millionaire’s mistress (modeled on Hearst’s real mistress, actress Marion Davies), and the millionaire accidentally kills the scientist’s assistant in a jealous rage. He and Dr. Obispo have to flee the law, so they run to England and try to find out what happened to the third earl of Gonister. Finally, they break into his castle, and in the basement they find him, still alive and over 300 years old—all grown up to become an adult ape. I’ve spoiled the punch line for an excellent novel, but it’s worth reading for the richness of detail and the amazing ironies that Huxley was so great in capturing.
In recent years, even more startling breakthroughs have occurred in molecular genetics. For centuries, the scientific community was deeply racist, and treated non-white people as inferior, or even a different species from white people. But the molecules paint a completely different picture. When you put the DNA of all of the human “races” in the mix with the DNA of Neanderthals and most of the living species of chimps and gorillas, a surprising result emerges. As shown in figure 15.11, the genetic differences among all the human “races” are tiny. All humans are far more genetically similar to one another than the populations of West African chimps are to one another, and the same is true of other populations of chimps and gorillas. As anthropologists have been saying for years, human “races” are genetically meaningless, and their basis is only a tiny part of our genome. In fact, evidence shows that most of the “racial differences” such as skin color and the shape of the eyes are a very recent change in human evolution, occurring sometime after nearly all modern human lineages emerged from Africa about 70,000 years ago. This is an important thing to think about when we find issues of race coming up in society.
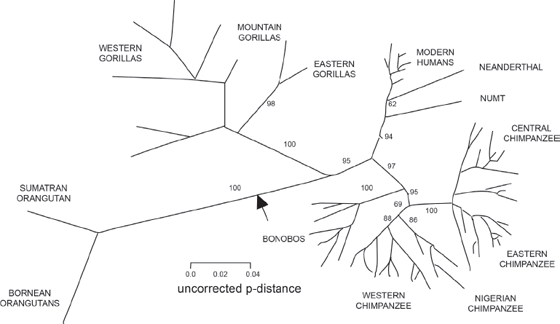
FIGURE 15.11. Molecular phylogeny of apes and humans, showing their genetic distance from one another based on mitochondrial DNA. All human “races” are much more similar to one another than two populations of gorillas or chimpanzees are to each other. (Modified from Pascal Gagneux et al., “Mitochondiral Sequences Show Diverse Evolutionary Histories of African Hominoids,” Proceedings of the National Academy of Sciences USA 93 [1999], fig 1b: ©1999, National Academy of Sciences USA)
Some people may find it unsettling that we are 97.6 percent genetically identical to the chimpanzee, but no amount of creationist propaganda can change the truth found in every cell in your body. Instead, it is better to accept that the evidence of science has shown us just how much an integral part of nature we really are; we are part of creation, and we cannot treat the rest of creation as something to be abused. So the next time you see a chimpanzee in a zoo, have pity on your close relative and think hard about the implications of that handful of genes that differentiate you from it.
As Smith and Sullivan (2007:100) put it:
Do people come from monkeys? Not at all. We do share a common ancestor with chimpanzees, and before them, with the group that became monkeys. But to say we come from monkeys is simply wrong, and evolution has never claimed it…. Unless we want to live in a web of lies, we can’t pick and choose what to believe, not when the raw data of genetics, fossil studies, and anatomical studies are laid before us.
Anyone is free to argue whether they like being a primate or whether they like being related to chimpanzees—but that’s not the question. The question is whether or not we’re descended from monkeys, and the evidence is in: we’re not, but we are related to them.
For Further Reading
Beard, K. C. 2004. The Hunt for the Dawn Monkey: Unearthing the Origin of Monkeys, Apes, and Humans. Berkeley: University of California Press.
Conroy, G. C. 1990. Primate Evolution. New York: Norton.
Delson, E. C. 1985. Ancestors: The Hard Evidence. New York: Liss.
Diamond, J. 1992. The Third Chimpanzee: The Evolution and Future of the Human Animal. New York: HarperCollins.
Johanson, D., and B. Edgar. 1996. From Lucy to Language. New York: Simon & Schuster.
Lewin, R. 1987. Bones of Contention: Controversies in the Search for Human Origins. Chicago: University of Chicago Press.
Lewin, R. 1988. In the Age of Mankind: A Smithsonian Book on Human Evolution. Washington, D.C.: Smithsonian Institution Press.
Lewin, R. 1998. Principles of Human Evolution: A Core Textbook. New York: Blackwell.
Marks, J. 2002. What It Means to be 98 Percent Chimpanzee. Berkeley: University of California Press.
Pääbo, S. 2014. Neandertal Man: In Search of Lost Genomes. New York: Basic.
Prothero, D. R. 2016. The Princeton Field Guide to Prehistoric Mammals. Princeton, N.J.: Princeton University Press.
Sibley, C. G., and J. E. Ahlquist. 1984. The phylogeny of hominoid primates, as indicated by DNA-DNA hybridization. Journal of Molecular Evolution 20:2–15.
Stringer, C., and C. Gamble. 1993. In Search of Neanderthals: Solving the Puzzle of Human Origins. London: Thames and Hudson.
Swisher, C. C., III, G. H. Curtis, and R. Lewin. 2000. Java Man. New York: Scribner.
Tattersall, I. 1993. The Human Odyssey: Four Million Years of Human Evolution. Upper Saddle River, N.J.: Prentice Hall.
Tattersall, I. 2015. The Strange Case of the Rickety Cossack and Other Cautionary Tales from Human Evolution. New York: St. Martin’s.
Tattersall, I., and J. Schwartz. 2000. Extinct Humans. New York: Westview.