Les animaux succèdent aux animaux
L’histoire des espèces végétales et animales est inscrite dans les fossiles. Quelques-uns montrent des « lignées » reconnaissables, qui nous permettent de suivre l’évolution de plusieurs animaux actuels. Mais pour établir une parenté, il faut faire le tri entre les ressemblances trompeuses et les véritables traits communs.
L’évolution des organes
Comme un poisson dans l’eau
C’est toujours un bonheur lorsqu’on est en bateau, dans le vent vif et les embruns salés, d’être entouré de dauphins jouant avec l’étrave ; une rencontre familière, pour Darwin l’attrape-mouche. Nous aimons spontanément les dauphins, peut-être à cause de quelques points communs évidents : nous sommes, nous aussi, des mammifères intelligents à peau nue (les cétacés sont des mammifères, nous l’avons appris à l’école).
D’autres animaux ressemblent étrangement à des poissons sans en être. Ainsi l’ichtyosaure, un reptile aquatique disparu ; et même un oiseau, le manchot, lorsqu’il file sous l’eau en se servant de ses ailes comme de nageoires. Tous atteignent la même perfection hydrodynamique, proche de celle du requin. Cette similitude apparente entre animaux de parenté lointaine s’appelle la convergence évolutive.
L’exemple du dauphin et de l’ichtyosaure est bien connue ; il en existe des milliers d’autres chez les plantes et les animaux. L’Australie, en particulier, est une mine de convergences. Les mammifères marsupiaux (dont les petits naissent à l’état d’embryon et finissent leur développement dans des poches dites marsupiales) y prennent des formes étonnamment semblables à celles des mammifères du reste du monde, qui sont presque tous placentaires (l’embryon est nourri par le placenta et naît à un stade beaucoup plus développé). Le thylacine, ou loup de Tasmanie, dont le dernier survivant connu fut abattu en 1961, ressemblait à un loup au dos rayé. Ce marsupial était doté de dents de carnivore et de pattes fines de coureur tout comme les vrais canidés. Sa silhouette était la même que celle d’un renard ou d’un loup, et la logique commune de leur anatomie de carnassiers correspond parfaitement aux préceptes de Cuvier. Le loup est pourtant plus proche du dauphin que du thylacine qui, lui, était un cousin des koalas. En Australie vivent encore des chats marsupiaux et des souris marsupiales, dont les noms sont des clins d’œil à la faune européenne. D’autres marsupiaux, comme les kangourous, ont une démarche parfaitement originale, mais montrent d’évidentes similitudes anatomiques avec des mammifères placentaires partageant les mêmes habitudes alimentaires.
La taupe marsupiale d’Australie présente de nombreux points communs avec la taupe européenne. Ces deux mammifères ont des mœurs souterraines. Ils creusent la terre ou le sable avec leurs pattes avant en forme de pelles et se nourrissent de vers de terre et d’insectes. Plus étonnant encore : la convergence se retrouve chez un insecte, la courtilière, encore appelée grillon taupe. Les pattes de la courtilière sont conformées de la même manière que celles des mammifères fouisseurs, en forme de pelle.

Convergence évolutive : les pattes antérieures en forme de pelle de la courtilière et de la taupe
Vous avez dit « adaptation » ?
Dans L’Origine des espèces, Darwin cite des convergences chez les animaux domestiques, tels le lévrier et le pur-sang, tous les deux splendidement « adaptés » à la course. Il souligne que ces convergences sont des bases trompeuses pour classer les espèces. Il s’agit d’« adaptation », et non d’une preuve de parenté.
Pourquoi ces guillemets au mot « adaptation » ? Parce qu’il est trompeur. Il sous-entend qu’une espèce animale s’« adapte » activement à son milieu, comme la girafe de Lamarck censée allonger son cou à force de le tirer. De fait, c’est le contraire : les contraintes du milieu ont sélectionné les animaux capables de survivre, tout comme les éleveurs ont sélectionné les chevaux performants pour arriver au pur-sang. Allez, je vous aide : cette question fait partie de l’interro surprise à la fin de ce livre.
Haut les mains !
La forme des baleines et des dauphins témoigne de leurs mœurs aquatiques, mais les unes et les autres sont comme nous des mammifères vertébrés. Pour le vérifier, il faudrait « déshabiller » l’une de leurs nageoires. Nous constaterions qu’il s’y cache une main étrangement semblable à la nôtre. Cette structure commune entre la main des différents vertébrés est un des arguments de Charles Darwin (et avant lui d’Erasmus Darwin et d’Étienne Geoffroy Saint-Hilaire) en faveur de leur parenté : « N’est-il pas très remarquable que la main de l’homme faite pour saisir, la griffe de la taupe destinée à fouir la terre, la jambe du cheval, la nageoire du marsouin et l’aile de la chauve-souris, soient toutes construites sur un même modèle et renferment des os semblables, situés dans les mêmes positions relatives1 ? »
Darwin appelle « unité de type » cette concordance des formes, indépendante des habitudes de vie (le terme actuel est homologie). Il l’explique par « l’unité de descendance ». Et d’ironiser au passage, comme il se plaît parfois à le faire, sur les naturalistes de son époque qui croient en une création unique : pourquoi, en effet, un créateur se serait-il évertué à inclure ces parentés inexplicables dans des anatomies aux fonctions si différentes ? Parentés inexplicables, sauf bien sûr par la théorie de la transmutation des espèces…
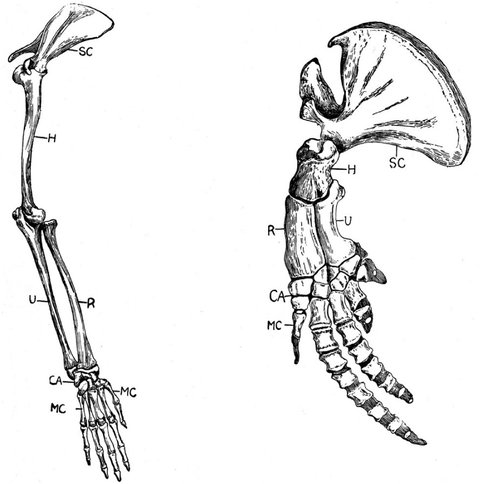
Comparaison entre les membres antérieurs d’un homme et d’un dauphin
L’apparition des pattes et des plumes
Un jour ou l’autre, un vertébré a bien dû s’aventurer hors de l’eau, pionnier de la conquête du monde terrestre. Parmi les candidats à cette sortie des eaux, le tiktaalik, dont des fossiles ont été exhumés en 2006 sur une île de l’Arctique canadien, est le dernier découvert. Cet animal encore un peu poisson et déjà un peu quadrupède, dont l’existence a été prévue par Darwin, avait une allure de salamandre. L’endroit où il vivait voici 375 millions d’années devait ressembler aux bayous de Louisiane, et le climat était équatorial. Les nageoires du tiktaalik comportaient un cubitus et un radius (os de l’avant-bras), un humérus (os du bras) et une ébauche de doigts. L’animal se servait plus souvent de ses poumons que de ses branchies.
D’autres êtres similaires, mi-poissons mi-quadrupèdes, ont porté quatre, sept ou huit doigts à certains membres. Visiblement, c’est la configuration à cinq doigts qui l’a emporté. Imaginez que les huit doigts aient conquis le monde, notre façon de compter aurait été bien différente. Quant à nos dons de pianistes…
Petite précision nécessaire : le tiktaalik n’est pas a priori notre ancêtre direct. Il donne juste une idée de l’aspect qu’ont eu ses collègues s’aventurant hors de l’eau, dont faisait partie notre ancêtre.
Autre précision : on pourrait croire que des pattes ont poussé aux poissons dans le « but » de leur permettre de marcher. En fait, c’est l’inverse qui se produit : une variation apparaît, et elle est utile ou non, retenue par la sélection ou non. L’exemple des plumes est révélateur : tout indique qu’elles sont apparues bien avant le vol, vraisemblablement pour maintenir constante la température de petits dinosaures non volants. Le vélociraptor de Jurassic Park en portait, et même les petits du terrible tyrannosaure2.
Aux origines, semble-t-il, un fin duvet est d’abord apparu sur le corps de certains dinosaures. Peut-être de petites plumes ont-elles ensuite servi lors des parades nuptiales, nul ne le sait. En tout cas, une fois suffisamment développé, cet attribut a fini par servir à voler. Les dinosaures emplumés nous l’affirment : ce n’est pas la fonction qui crée l’organe, comme le disait Lamarck, mais bien l’organe qui précède la fonction.
Un fossile mis au jour dans le désert de Gobi montre des prémices d’ailes qui pourraient faire de ce petit dinosaure nommé Mahakala omnogovae un bon prétendant au titre d’ancêtre des oiseaux. Mais que penser de Microraptor gui, découvert en Chine, qui portait quatre ailes, deux à l’avant, deux à l’arrière ?
L’algue qui a des visions
Imaginer que des nageoires se sont transformées en pattes, et des filaments en plumes, procède d’une mécanique relativement simple. Mais comment concevoir qu’un organe aussi sophistiqué que l’œil ait pu apparaître autrement que par intervention divine ? Conscient de la force de cet argument avancé par ces casse-pieds de créationnistes, Darwin a avancé dans L’Origine l’existence probable de nombreuses gradations de l’œil simple à l’œil complexe en parfaite concordance avec sa théorie.
Un nerf est apparu, qui était sensible à la lumière – d’autres l’ont été au son –, là est l’origine de la vision. Les fossiles étant muets à ce sujet, il prend l’exemple des yeux des crustacés, moins développés que ceux des vertébrés. Nous connaissons aujourd’hui des organes photosensibles plus simples encore que ceux des crustacés, chez les méduses, ou chez les vers de terre, que le naturaliste fréquentera si assidûment à la fin de sa vie. Il existe même l’« œil » primitif imaginé par Darwin, chez… une algue : l’euglène !
La gradation a donc bien commencé avec des organes de vision simples, Darwin l’avait vu. Repérer les lumières et les ombres, et donc l’apparition d’un prédateur, est un avantage adaptatif évident. Suivons le développement d’un organe photosensible chez les mollusques. Il part de la tache sensible des praires et d’autres coquillages, continue avec les yeux plus évolués des escargots jusqu’à la complexification des yeux des pieuvres, qui, bien que très éloignés des animaux vertébrés, ont des yeux étonnamment comparables aux nôtres. D’après des biologistes comme Richard Dawkins, quatre cent mille générations (soit moins de cinq cent mille ans) auront été nécessaires pour passer de la tache oculaire photosensible la plus simple, comme celle des unicellulaires, à l’œil abouti des humains et des pieuvres.
Le regard inexplicable de la sole
La position des yeux des poissons plats, comme les soles, soulève un autre problème. Ces poissons chassent à l’affût, camouflés sur le substrat, et la fonction de ces yeux surélevés, situés sur le même côté, présente aujourd’hui un avantage certain. À l’état de larves, ces animaux ont un regard « normal » avec un œil de chaque côté de la tête, mais au cours de leur croissance, l’un d’eux migre et se retrouve du même côté que son alter ego ! Quel intérêt auraient trouvé les ancêtres intermédiaires des poissons plats affublés d’un regard présentant un début d’asymétrie ? Attaqué sur le sujet (devinez par qui), Darwin y répond dès la seconde version de L’Origine avec ces deux arguments : un organe en développement peut avoir eu une fonction autre que celle qu’il occupe aujourd’hui, lié à un caractère avantageux différent.
L’origine exacte des caractéristiques des poissons plats n’a pas encore été trouvée, ce qui ne remet pas en cause l’ensemble de la théorie de l’évolution, mais la question passionne les biologistes. En 1982, une étude sur les flets du Pacifique, qui présentent des individus « droitiers » (les deux yeux à droite) ou « gauchers », a déterminé que cette migration oculaire dépendait de facteurs à la fois génétiques et comportementaux. La raison de l’amorce de la migration n’est pas totalement résolue, mais les fossiles de deux poissons vieux de quarante-cinq millions d’années, Amphistium et Heteronectes, qui présentent un cheminement partiel de l’œil, viennent d’être découverts… dans les tiroirs des muséums d’histoire naturelle de Paris et de Vienne, en Autriche. Comme l’archéoptéryx arrivait à point nommé pour contrer les adversaires des théories de Charles Darwin, ces fossiles montrant une migration graduelle des yeux sont un témoignage de poids contre les attaques actuelles des créationnistes.
Petite leçon de bricolage
La plume du vélociraptor, la patte du tiktaalik et l’œil de la pieuvre nous montrent que l’évolution bricole avec ce qu’elle a déjà, selon la fameuse métaphore du généticien François Jacob : l’évolution est « un bricoleur qui ne sait pas encore ce qu’il va produire, mais récupère tout ce qui lui tombe sous la main3 ». C’est seulement ce qui fait déjà partie de l’anatomie d’un animal qui peut être recyclé, transformé, développé : notre langue nous est nécessaire pour parler, mais elle n’a pas été « destinée » au langage.
Nous portons tous en nous un bricolage anatomique hérité de notre ascendance plus ou moins lointaine, tels les os de nos mains à cinq doigts… ou ceux de l’intérieur de nos oreilles. L’oreille moyenne des mammifères (la partie juste derrière le tympan) possède trois osselets : l’enclume, le marteau et l’étrier ; alors que celle des reptiles n’en comporte qu’un : l’étrier. Bien après la sortie des eaux du tiktaalik et de ses homologues sont apparus les dinosaures. Certains d’entre eux, les reptiles mammaliens, ont conduit aux premiers mammifères. Les deux petits os qui formaient l’articulation entre leur crâne et leur mâchoire inférieure ont migré. On peut suivre, de fossile en fossile sur près de cinquante millions d’années, le cheminement de ces deux osselets se détachant progressivement de la mâchoire inférieure pour rejoindre l’étrier et l’oreille moyenne, caractéristiques des mammifères accomplis. Ces trois petits os (l’étrier est le plus petit os du corps humain) transmettent les vibrations reçues par le tympan et permettent une bonne ouïe. Nous entendons bien grâce à des bouts de mâchoire de dino !
Les ancêtres du cheval et de la baleine
De la même manière que tatie Tiktaalik est sortie des eaux, les oiseaux ont pris leur envol, de petits mammifères se sont mis à galoper, et les ancêtres de la baleine ont réappris à vivre dans l’eau. Chacun des organes de ces individus a participé à leur évolution globale. Au cours des millénaires, ces animaux se sont entièrement transformés.
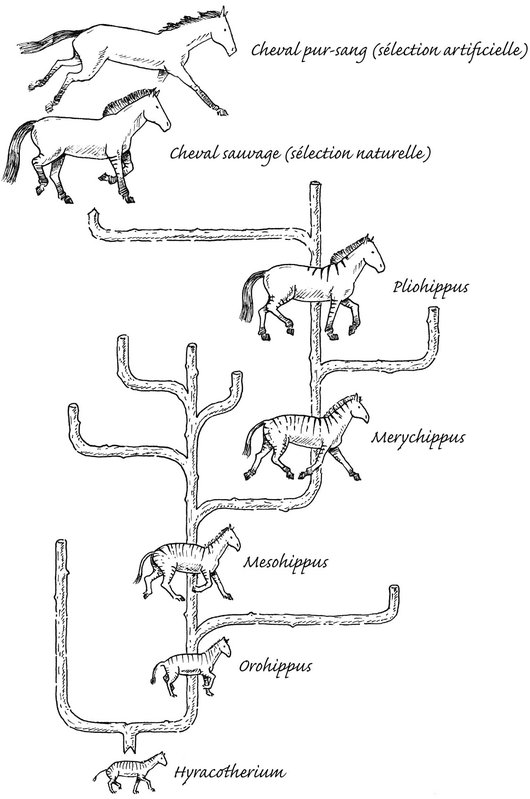
La « lignée » du cheval. Seuls les fossiles de la tendance ayant conduit au cheval sont représentés
Petit cheval deviendra grand
Les personnes peu familières des chevaux s’étonnent volontiers des « blessures » qu’elles voient sur leurs jambes (mais oui, pour la plus noble conquête de l’homme, on parle de jambes et non de pattes), une excroissance très visible, comme un petit moignon qui sortirait de la jambe. Un examen plus attentif en révèle quatre : une à chaque jambe. Ce sont les châtaignes. Il ne s’agit pas ici de végétal, mais de la relique d’un doigt perdu au cours de l’évolution. Les poulains naissent d’ailleurs parfois avec un ou deux doigts surnuméraires, ce qui peut s’interpréter comme un témoignage de l’aspect de leurs ancêtres.
L’évolution des ancêtres du cheval vers sa forme actuelle est bien connue, et le graphique en est très convaincant : on part de l’Hyracotherium (moins 55 millions d’années), de la taille d’un agneau, vers des animaux de plus en plus grands, dont les os des cinq doigts se soudent peu à peu jusqu’à former des pieds à un sabot. En réalité les choses sont plus compliquées, car l’évolution ne se fait pas en ligne droite comme si elle suivait une direction prévue. Il s’agit plus d’une ramification que d’une « lignée ».
Manœuvrer les pattes demande beaucoup d’énergie, et chez les animaux coureurs, on constate une fusion des muscles ou des os, sinon leur perte. Dans la lignée fossile des équidés, la manière dont les doigts latéraux ont disparu n’est pas progressive, mais ils ont bel et bien disparu, la châtaigne et l’anatomie interne du pied en sont des reliques ; l’os principal, le canon, est encore entouré de deux stylets, vestiges de deux doigts. Au bout du pied, le sabot est l’homologue de notre ongle : les chevaux actuels galopent sur leurs ongles, et font partie des ongulés. Les ongulés sont des herbivores, des proies potentielles, et doivent courir vite pour échapper à leurs prédateurs. Tout au long de son histoire naturelle, le petit cheval a grandi. Il s’est mis sur la pointe des pieds, toujours plus fin, toujours plus rapide, s’approchant toujours plus du ciel. Le pur-sang nous rappelle que grâce à la sélection artificielle, qui a pris le relais de la sélection naturelle, il est devenu une sorte de perfection. En toute modestie.
Comment la baleine s’est jetée à l’eau
Marcher sur ses ongles est un mode de déplacement « inventé » plusieurs fois par l’évolution. Des ongulés ont suivi des chemins bien différents de ceux de la ramification du cheval. Le Pakicetus, par exemple, ressemble vaguement à un chien, mais un chien qui aurait des petits sabots. Il vivait dans les marécages du Pakistan voici quelque 52 millions d’années. Pakicetus était à l’aise au sol, mais devait souvent plonger pour attraper ses proies. Cette bête étrange est un ancêtre possible des baleines et des dauphins. C’est en tout cas le plus ancien fossile connu, qui partage un petit os de son oreille interne avec les cétacés, ce qui indique une parenté. Un autre animal découvert plus récemment, l’Indohyus, possède lui aussi une oreille interne proche de celle des cétacés. Semblable à un chevreuil miniature, Indohyus vivait dans l’actuel Cachemire il y a 48 millions d’années. Contrairement à Pakicetus, il était herbivore. Il pourrait avoir été lui aussi un ancêtre des baleines et des dauphins, bien que les cétacés soient tous carnivores. Indohyus et Pakicetus se sont aventurés dans l’eau, l’un pour se nourrir, l’autre sans doute pour fuir des prédateurs. Un chevreuil aquatique ? c’est tout à fait possible : un petit cervidé africain gros comme un lièvre, le chevrotain aquatique, plonge en cas de danger.
De nombreux fossiles ont été découverts depuis Darwin et confortent ses écrits. Bien qu’ils restent insuffisants, on peut suivre avec des fossiles d’espèces de plus en plus récentes des animaux de plus en plus aquatiques jusqu’aux dauphins et aux baleines : Pakicetus, Ambolucetus, Rodhocetus… Les corps se sont fuselés et ont pris une position plus horizontale, les pattes avant sont devenues des nageoires, les pattes arrière se sont rétrécies jusqu’à disparaître, les narines sont remontées sur la nuque, vers la surface de l’eau. Le plus connu des cétacés fossiles est sans doute le Basilosaurus, découvert en 1832, époque à laquelle Darwin entamait son tour du monde à bord du Beagle. Le Basilosaurus était une baleine de vingt-cinq mètres de long très allongée, si allongée, que ses os ont servi à un canular sur un prétendu serpent de mer4.
En résumé, après être sortis des eaux, puis avoir évolué en mammifères, certaines lignées animales ont « fait demi-tour » et sont retournées à une vie totalement aquatique. Les baleines et les dauphins d’aujourd’hui naissent dans l’eau, et sont entièrement affranchis de toute vie terrestre. Darwin, avec son image de l’ours pêcheur se transformant en baleine, n’a pas eu tout à fait tort. Parant à l’avance l’inévitable attaque des créationnistes sur l’absence de formes intermédiaires, le naturaliste souligne dans L’Origine qu’on peut en trouver de parfaitement viables chez les espèces actuelles. Celles-ci, adaptées à la vie aquatique à des degrés variés, nous donnent une idée de la succession de ces formes passées. Bien sûr, ces animaux contemporains offrent juste une possibilité de comparaison, qui ne présume en rien de l’apparence des véritables ancêtres des cétacés.

Animaux actuels de plus en plus aquatiques : ils donnent une idée des espèces disparues à différentes étapes d’adaptation à la vie aquatique
Les pattes du serpent
Si le cheval exhibe des restes de doigts, la baleine porte en elle des traces de ses ancêtres plus secrètes : son squelette contient encore des reliques de bassin et d’anciennes pattes arrière. De tels vestiges – on parle aujourd’hui de structures vestigiales – sont fréquents. Aux Galápagos, Darwin et FitzRoy avaient été frappés par les moignons d’ailes des cormorans aptères, des oiseaux incapables de voler, mais ils n’avaient pas su expliquer ces ailes inutiles.
Les différentes espèces de brassemers, ou canards vapeur – souvent évoqués par Darwin sous les noms de canard à ailes courtes d’Amérique méridionale, et de bateau à vapeur pour l’espèce rencontrée aux Falkland –, en sont un autre exemple : alors que les autres canards sont d’excellents voiliers, généralement grands migrateurs, celui-ci est incapable de décoller. Le brassemer fuit les prédateurs en battant des ailes à la surface de l’eau, ce qui provoque des remous spectaculaires, qui évoquaient aux observateurs du XIXe siècle les bateaux à vapeur de leur époque. Darwin explique la disparition de ses capacités par le « défaut d’usage », point sur lequel il a été remis en question. Il en a déduit une loi d’économie : la nature ne dépense pas d’énergie à entretenir un organe sans fonction.
Ces ailes inutiles et ces os vestigiaux constituent avant tout, pour Darwin, un argument extrêmement convaincant contre la théorie d’une création spontanée unique, car la présence d’un organe dénué de fonction ne peut s’expliquer que par l’existence de changements passés. Il compare ces « organes rudimentaires aux lettres qui, conservées dans l’orthographe d’un mot, bien qu’inutiles pour sa prononciation, servent à en retracer l’origine et la filiation5 ». Ces structures vestigiales plaident non seulement en faveur de la « descendance avec modification », mais aussi pour une évolution par petites graduations.
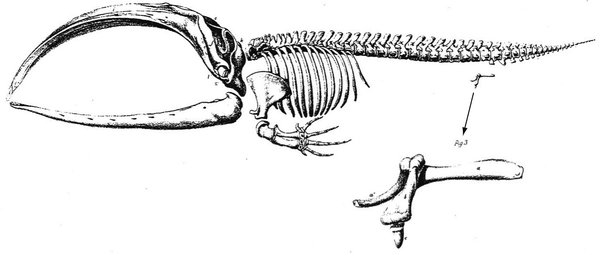
Vue latérale d’un squelette de baleine franche, avec, au-dessous, le détail des vestiges du membre inférieur rudimentaire
Darwin pensait que les organes rudimentaires, s’ils avaient perdu leur fonction première, pouvaient garder un rôle anatomique secondaire. Les vestiges de bassin des baleines auraient une fonction dans l’accouplement et la mise bas. Chez certains serpents, qui possèdent encore, comme les baleines, des os vestigiaux de pattes arrière, ce rôle se situe également au-dessous de la ceinture (quoiqu’une ceinture, pour un serpent…). Les boas portent deux petits moignons externes bien visibles dans la région cloacale, et ils savent s’en servir. Au cours de la parade nuptiale, les boas mâles agitent ces deux éperons pour chatouiller activement leur partenaire, et préparer ainsi l’accouplement. Faire l’amour comme un pied, c’est bien vu chez les serpents !
Les structures vestigiales existent chez les humains également, il n’y a pas de raison. Darwin en a énuméré un bon nombre dans La Filiation de l’homme : les poils corporels, les dents de sagesse, la membrane du coin de l’œil, les muscles de l’oreille, l’appendice, et bien sûr le coccyx, que l’on interprète comme un reste de la queue de nos ancêtres arboricoles.
Les moustaches du dauphin
Si les dauphins adultes ont la peau glabre et lisse comme un ballon de baudruche, leurs bébés portent à la naissance des poils autour du rostre (le museau). En remontant le cours de leur évolution individuelle, on s’aperçoit qu’à l’état d’embryon, les cétacés sont munis de quatre bourgeons de membres. Seules les deux esquisses de membres antérieurs se développeront pour donner des nageoires ; les deux autres régresseront et resteront les vestiges précédemment évoqués, sauf quelques cas où ils subsistent à l’âge adulte. Les fœtus de baleines, quant à eux, possèdent des bourgeons de dents qui disparaissent avant même leur naissance.
Darwin a consacré un chapitre de La Filiation de l’homme au développement embryonnaire, car celui-ci évoque étrangement l’évolution passée. Le fœtus humain porte des fentes branchiales rappelant l’ancêtre aquatique ; au sixième mois, il est couvert d’une fourrure, le lanugo, qui ressemble fort à un vestige de pelage. Évoquant la nuit des temps, notre histoire profonde semble gravée en nous dès la première étincelle de notre vie.
Fan de Darwin (qui ne le lui rendait pas) et grand diffuseur des idées transformistes en Allemagne, le naturaliste Ernst Haeckel (1834-1919) a caricaturé les choses par cette formule célèbre : « L’ontogénie récapitule la phylogénie. » Autrement dit, le développement de l’individu reprend les étapes de l’évolution de son espèce. C’est à moitié vrai et donc, vous l’avez compris, c’est aussi à moitié faux. D’abord, grosse nuance, l’embryon ne ressemble pas à ses ancêtres adultes, mais aux embryons des autres espèces au début de son développement. De plus, la formule d’Haeckel réintroduit une notion de hiérarchie dépassée6 qui ne correspond pas aux ramifications de l’évolution selon la théorie défendue par Darwin.
En revanche, le développement de l’embryon est bien une preuve de plus de la parenté ancestrale des êtres vivants. Le 12 septembre 1860, Darwin écrit à Lyell que pour défendre sa théorie sur la transmutation des espèces, l’argument embryologique est « le plus puissant de tous ».
Au fait, qu’est-ce qu’une espèce ?
Merci d’avoir posé cette question…
Attention, en lisant ce qui suit vous allez mettre les pieds – ou plutôt les yeux – dans un bourbier, car la réponse n’est pas claire. L’être humain tente de ranger méticuleusement la nature selon un ordre compréhensible, avec des limites bien rassurantes, mais elle se prête très mal au jeu.
Pourtant, à la base, tout semble simple.
Depuis l’enfance, nous savons nommer les animaux sans difficulté, et nous ne faisons pas de confusion entre une vache, un chat et une poule. Ces animaux sont classés en espèces. Celles-ci ont été déterminées avec précision au XVIIIe siècle par le naturaliste suédois Carl von Linné au moyen d’une appellation scientifique en deux termes, le nom de genre et le nom d’espèce, qui correspondent plus ou moins à nos nom et prénom7. Mais dès qu’on y regarde de plus près, les choses se compliquent.
Dès le début de L’Origine des espèces, Darwin souligne que la notion d’espèce est floue, ainsi que celle de variété (on préfère aujourd’hui le terme de sous-espèce). Évidemment, pour quelqu’un qui plaide en faveur de la modification des organismes, la notion d’espèce définie supposerait une fixité en contradiction avec ses idées.
Mais alors ?
Alors, qu’est-ce qu’une espèce ? Un ensemble d’animaux qui se ressemblent, certes, mais il y a quelquefois une telle différence entre mâles et femelles, ou entre adultes et petits, que l’aspect physique ne suffit plus à déterminer une parenté : une chenille ne ressemble vraiment pas au papillon, il s’agit pourtant de la même espèce… Des animaux qui descendent les uns des autres ? Oui, mais si l’on remonte dans le temps, tous les organismes vivants actuels descendent théoriquement d’un ancêtre commun : Luca (Last Universal Common Ancestor)8. Il faut donc affiner la définition avec l’interfécondité : deux animaux de la même espèce peuvent se reproduire, et leur descendance est elle-même fertile. Au contraire, la stérilité des descendants indique la séparation entre deux espèces. Les produits de l’accouplement entre un âne et une jument (le mulet) ou de celui d’un cheval et d’une ânesse (le bardot) sont stériles.
Cette définition de l’espèce comporte des exceptions – les plantes et les animaux ne respectent rien ! Le tigre et le lion, qui appartiennent à deux espèces bien séparées, non seulement peuvent se reproduire et donner des animaux étonnamment gros, mais les produits de ces hybridations, les tigrons (tigre + lionne) et les ligrons (lion + tigresse), théoriquement stériles, se sont récemment révélés féconds.
Il faut donc encore affiner les choses avec la notion de population. Elle désigne un groupe d’individus qui se perpétuent dans un habitat, avec des habitudes propres. Le biologiste évolutionniste Ernst Mayr a résumé ainsi la situation : « Les espèces sont des groupes de populations naturelles interfécondes qui se sont isolées d’autres groupes semblables sur le plan reproductif. » Pour la biologie actuelle, l’espèce reste l’unité fondamentale du monde vivant. Cette science est donc basée sur la notion scientifiquement floue d’espèce, qui n’est qu’un état génétique provisoire de l’évolution. Peut-être la biologie évoluera-t-elle bientôt vers une conception a-spécifique plus conforme à la fluctuation générale de la vie.
Les vacheries de la classification
Le grand arbre de l’évolution
Pour nommer les êtres vivants de manière universelle et donner un sens au foisonnement de la vie, les naturalistes occidentaux se sont attelés à les répertorier pour les classer. Dans cette classification, issue des travaux de Linné, les groupes zoologiques (ou taxons) sont assemblés selon une hiérarchie. La base en est l’espèce. Chaque espèce fait partie d’un genre, qui lui-même est inclus dans une famille, elle-même dans un ordre, puis une classe, puis un embranchement. Par exemple, l’espèce renard roux Vulpes vulpes est incluse dans le genre Vulpes, de la famille des canidés, de l’ordre des carnivores, de la classe des mammifères, de l’embranchement des cordés.
Pour définir des groupes zoologiques, les naturalistes ont associé les animaux en fonction de leurs points communs (principalement anatomiques), et de tout temps les discussions sont allées bon train pour déterminer des critères fiables. Sans cesse, certains taxons botaniques ou zoologiques sont renommés. Ils apparaissent ou disparaissent selon les découvertes et les idées en vigueur (pour ne pas dire les plus branchées, comme nous allons le voir).
Après Darwin, les animaux n’ont plus été seulement regroupés en fonction de leur anatomie, mais aussi de leur parenté évolutive. Les ressemblances dues à un héritage commun sont désormais prises en compte. L’approche évolutionniste cherche les caractères primitifs des espèces pour établir leur parenté ancienne (un caractère dit primitif est ancien, proche de l’ancêtre, alors qu’un caractère dit évolué est apparu plus récemment). Au final, la classification hiérarchique des espèces passées et présentes dessine un gigantesque arbre généalogique décrivant l’évolution du vivant depuis son apparition, de plus en plus stable et majestueux comme le chêne de la fable de La Fontaine, ou se ramifiant comme le corail récolté par Darwin au cours de son périple. Jusque-là, tout va bien.
Le dauphin est un peu vache
En 1950, l’entomologiste Willi Hennig (1913-1976) propose une approche nouvelle pour regrouper les animaux par parenté : la cladistique (du grec klados, « branche »). La méthode cladistique repose sur un principe de base assez simple (et sur un jargon beaucoup plus alambiqué que je vous épargne). Lorsqu’un caractère nouveau apparaît chez un organisme, celui-ci le transmet à ses descendants. Donc, si plusieurs êtres (ou groupes) partagent une même nouveauté évolutive, ils l’ont forcément héritée d’un ancêtre commun chez qui ce caractère nouveau est apparu.
Héritière de l’évolutionnisme darwinien, la cladistique se fonde sur de nouveaux critères de parenté, basés sur les caractères les plus évolués et non sur les plus anciens pour classer les êtres vivants. Et ça a tout bouleversé. L’exemple le plus significatif est celui de la classe des reptiles. Jusque-là, ces braves bêtes étaient bien séparées des oiseaux. Les oiseaux possèdent une nouveauté évolutive, les plumes, qui les différencie des reptiles, et là-dessus l’analyse cladistique ne contredit pas la classification traditionnelle. Hélas. Du côté des reptiles, les crocodiles sont – selon la cladistique, et malgré leur apparence – plus proches des oiseaux que des lézards ! Alors s’il faut inclure les crocodiles dans le même groupement que les reptiles, il faut y joindre tous les oiseaux, l’archéoptéryx et les dinosaures en prime : c’est le nouveau taxon des sauropsides.
Les analyses génétiques affinent chaque jour cette classification et bouleversent nos connaissances. Ainsi, dans le même ordre d’idées, on pense désormais que le brochet est plus proche du cheval qu’il ne l’est du requin, ou encore que la vache est plus proche du dauphin que du cheval. Les bovidés (qui sont des artiodactyles) et les cétacés sont donc maintenant regroupés dans la catégorie des cétartiodactyles. Du côté de nos cousins, le chimpanzé est plus proche de l’homme que du gorille, comme l’avait deviné Darwin. Nous sommes plus proches du saumon que celui-ci ne l’est du requin, ou encore plus proches des plantes à fleurs que des bactéries Escherichia coli qui vivent dans nos intestins…
Si la cladistique a dépoussiéré plusieurs branches de la classification traditionnelle en révélant des imperfections, elle comporte aussi les siennes, et connaît des détracteurs (ça, c’est classique !). En effet, la méthode cladistique comporte une certaine rigidité, et n’englobe pas la diversité évolutive. Par exemple, l’établissement des liens de parenté suppose que l’espèce mère disparaît systématiquement après la formation des nouvelles espèces, mais ce n’est pas toujours le cas, et cela remet en cause certains schémas. Bref, les animaux se moquent de l’ordre, et les systématiciens n’ont pas fini d’essayer de les mettre en rang.
1. Charles Darwin, L’Origine…, p. 491. Voir bibliographie.
2. Certains fossiles, en effet, sont entourés d’empreintes de plumes ; d’autres portent les traces des attaches.
3. Cette métaphore du bricoleur avait déjà été formulée par Darwin lui-même à la fin de son ouvrage sur la fécondation des orchidées, De la fécondation croisée des orchidées par les insectes : « Quoiqu’un organe n’ait pas été à son origine formé dans tel but spécial, s’il sert actuellement à la réalisation de ce but, on peut dire avec justesse qu’il est spécialement constitué pour lui. D’après le même principe, si un homme construit une machine dans une fin déterminée, mais emploie à cet effet, en les modifiant un peu, de vieilles roues, de vieilles poulies et de vieux ressorts, la machine, avec toutes ses parties, pourra être considérée comme organisée en vue de cette fin. Ainsi, dans la nature, il est à présumer que les diverses parties de tout être vivant ont servi, à l’aide de modifications légères, à différents desseins, et ont fonctionné dans la machine vivante de plusieurs formes spécifiques anciennes et distinctes », p. 329. Voir bibliographie.
4. Canular ? Pas pour tout le monde. Les cryptozoologues (spécialistes des animaux cachés, dont les serpents de mer) constatent que la plupart des témoignages de marins ne décrivent pas des serpents, mais des mammifères, car les animaux aperçus ondulent verticalement, alors que les poissons et les reptiles se meuvent horizontalement. Les cryptozoologues pensent que certains de ces animaux décrits pourraient, justement, être des baleines primitives serpentiformes, du type Basilosaurus, qui auraient survécu jusqu’à nos jours. L’absence de preuves n’est pas une preuve d’absence… En tout cas, l’ondulation verticale des dauphins et des baleines, due à la conformation de leur colonne vertébrale, est un héritage de leur passé de coureurs terrestres.
5. Charles Darwin, L’Origine…, p. 513.
6. La notion de continuité entre les êtres vivants, qui implique celle de « chaînon manquant », date d’Aristote. Aristote avait émis l’idée d’une « chaîne des êtres » où les organismes vivants étaient liés les uns aux autres, avec une hiérarchie allant du plus simple vers le plus complexe. Tout en haut, juste sous la divinité, l’homme en était le deuxième maillon, suivi par la femme, puis par les esclaves, les animaux, etc. (selon les époques, on ajoutera, au-dessus de l’homme, les anges et les archanges). À l’aube du XXe siècle, Haeckel situait toujours la femme blanche au-dessous de l’homme blanc, au même niveau que le nègre et le « type simiesque ». Ses écrits ont beaucoup plu à Hitler et à ses copains.
7. Par exemple, la mésange bleue est nommée Parus caeruleus : Parus indique son genre « mésange » et caeruleus, qui veut dire bleu, précise son espèce. Du même genre, mais pas de la même espèce, la mésange charbonnière s’en distingue par son nom entier : Parus major. La sous-espèce (ou race, ou variété) désigne une division au sein de la même espèce avec un troisième terme. Parus caeruleus ultramarinus, par exemple, désigne la sous-espèce nord-africaine de la mésange bleue, dont la calotte est d’un bleu plus foncé que les autres.
8. On appelle cet ancêtre Luca pour faire un clin d’œil à Lucy, lointaine parente nettement plus récente (l’australopithèque Lucy n’a que 3,5 millions d’années, alors que la petite cellule théorique Luca date de plus de 3 000 millions d’années). Dans Darwin dessine-moi les hommes, le biologiste Claude Combes l’explique ainsi : « Les êtres vivants partagent tant de caractères dans leur structure et leur fonctionnement qu’il est quasi impossible de faire l’économie d’un ancêtre commun. »