As BENTLEY ET AL. note in chapter 8 of this volume, Darwinian approaches to archaeology are rapidly gaining popularity, fueled in part by the recognition that cultural evolution can be fruitfully studied by adapting the analytical tools that evolutionary biologists and paleobiologists use to study biological evolution. The goal of this chapter is to illustrate how two of these methods, population genetics modeling and cladistic techniques of phylogenetic reconstruction, can be used to explain temporal and geographic variation in cultural phenomena as a result of transmission, innovation, and selection. We will begin by discussing the conceptual foundations of this novel approach. We will then outline some of the ways in which models rooted in population genetics theory have been used to investigate cultural evolution. Lastly, we will discuss some cultural evolutionary applications of the cladistic method of phylogenetic reconstruction.
Proponents of three schools of thought have applied population genetics modeling and cladistic techniques to cultural data. The first is evolutionary archaeology, which is also occasionally referred to as selectionist archaeology (Abbott et al. 1996; Dunnell 1980, 1989; Leonard and Jones 1987; Leonard 2001; Lipo et al. 1997; Lyman and O’Brien 1998; Neff 1992; O’Brien and Holland 1995; O’Brien and Lyman 2000, 2002, 2003a; Ramenofsky 1995). The second is gene culture coevolution theory or dual inheritance theory (Ames 1996; Bettinger 1991; Boyd and Richerson 1985, 1993; Cavalli-Sforza and Feldman 1981; Durham 1979, 1982, 1990, 1991, 1992; Pulliam and Dunford 1980; Richerson and Boyd 1992; Runciman 2002; Shennan 1991, 2000, 2002). The third is known as human behavioral ecology (Borgerhoff Mulder 1991; Cronk 1991; Shennan 2002; Smith 2000; Smith and Winterhalder 1992; Smith et al. 2001; Winterhalder and Smith 2000). These schools of thought differ over whether cultural evolution is effected primarily through differential reproductive success or primarily through phenotypic plasticity. The former simply involves some individuals outreproducing others; the latter is the ability of organisms to produce different phenotypes in response to different environmental conditions (Pigilucci 1996). The schools of thought also differ over the role played by population-level phenomena in cultural evolution. These differences impact the way proponents of the three schools of thought envisage the relationship between cultural evolution and genetic evolution, and also affect their attitude toward adaptively neutral and maladaptive cultural behaviors.
The central tenet of evolutionary archaeology is that artifacts are as much a part of the human phenotype as are bones, muscle, and skin (Dunnell 1989; Leonard 2001; Lyman and O’Brien 1998; O’Brien and Holland 1995). The corollary of this is that evolutionary archaeologists seek to describe the archaeological record using classes amenable to evolutionary explanation (Cochrane 2001; Dunnell 1978, 1995; Tschauner 1994) and then account for the class distributions in terms of what they consider to be the primary mechanisms of evolution, namely natural selection and drift (Abbott et al. 1996; Braun 1987; Dunnell and Feathers 1991; Leonard 2001; Lyman and O’Brien 1998; O’Brien and Lyman 2000; Ramenofsky 1995).
Explaining what evolutionary archaeologists mean by the terms “natural selection” and “drift” is not easy because the definitions they have put forward differ in subtle but potentially important ways. This point can be illustrated by comparing the definitions of natural selection and drift provided by Wilhelmsen (2001) and O’Brien and Lyman (2000), which seem to represent the extremes of a range of views. According to Wilhelmsen (2001), natural selection is “the differential reproduction of organisms which, at the scale of population, results in a directional change in trait frequencies over time that correlates to external environmental conditions” (p. 99), and drift is the differential reproduction of organisms as a result of “sampling error and the stochastic patterns of transmission” (p. 100). According to O’Brien and Lyman (2000), selection is the “process by which certain forms in a population that are better adapted to a particular environment increase in proportion to less well-adapted forms” (p. 404), and drift is “random changes in trait frequency in a population resulting from the vagaries of transmission” (p. 399). One of the differences between the definitions is that Wilhelmsen’s (2001) focuses on “organisms” whereas O’Brien and Lyman’s (2000) definition of selection and, by extension, their definition of drift, focuses on “forms.” The other difference concerns the mechanisms of change involved in selection and drift. Wilhelmsen’s (2001) definition of selection explicitly links it with differential reproduction. That is, his definition suggests that in the case of selection, changes in trait frequencies over time are the result of some individuals producing more offspring than others. In contrast, O’Brien and Lyman’s (2000) definition of selection does not mention differential reproduction. Indeed, no mechanism of change is specified. Selection is suggested to involve better adapted forms increasing in frequency at the expense of less well-adapted forms, but the manner in which this occurs is not specified. The situation is similar with drift; Wilhelmsen’s (2001) definition explicitly links drift with differential reproduction, whereas O’Brien and Lyman’s (2000) definition does not mention differential reproduction. Rather, for O’Brien and Lyman (2000) drift is the consequence of the “vagaries of transmission.”
These differences are potentially important because, together, they bear on the way in which evolutionary archaeologists conceptualize the relationship between genetic evolution and cultural evolution. By linking selection directly with differential reproduction of organisms, Wilhelmsen’s (2001) definitions of natural selection and drift imply that culture evolves in exactly the same manner as genes, and that cultural evolution is therefore reducible to genetic evolution. In contrast, O’Brien and Lyman’s (2000) focus on forms rather than organisms and their decision to leave the mechanism of change unspecified opens up the possibility that O’Brien and Lyman (2000) recognize that genetic evolution and cultural evolution involve different mechanisms and therefore potentially have different dynamics. Thus it is possible that O’Brien and Lyman (2000) and some other evolutionary archaeologists consider selection to involve not only increase in relative frequency of adapted forms where the forms are organisms and the increase is brought about through differential reproduction, but also increase in relative frequency of adapted forms where the forms are artifacts and the increase is brought about through cultural transmission. In other words, it is possible that some evolutionary archaeologists take seriously the possibility that change in the archaeological record may be the result of phenotypic plasticity as well as differential reproductive success rather than just the result of the latter. In keeping with this, O’Brien and Lyman (2000) have noted that explanations of artifact class distributions and human biological reproduction may be generated at different analytical scales. Likewise, Leonard and Jones (1987; Leonard 2001) have drawn a distinction between the replicative success of cultural traits and the reproductive success of individuals, and argued that the former should not be assumed to be reducible to the latter.
However, having reviewed a good deal of the work that has been produced by evolutionary archaeologists over the past thirty years, we are of the opinion that the differences between the definitions offered by Wilhelmsen (2001) and O’Brien and Lyman (2000) are not in fact significant. O’Brien and Lyman (2000) and most other evolutionary archaeologists normally dismiss the possibility of change occurring as a consequence of individuals copying behaviors and then modifying them in light of experience before passing them on. They also usually treat transmission as if it occurs solely from parents to offspring, ignoring the various other forms of cultural transmission, which, as noted earlier, can in principle lead to the oblique and horizontal movement of cultural traits among individuals and populations in the absence of gene flow (e.g., Abbott et al. 1996; Leonard 2001; O’Brien and Lyman 2000; Ramenofsky 1995). Thus in practice, evolutionary archaeologists treat cultural evolution as if it is effected primarily through differential reproductive success, and therefore do not allow for the possibility that genes and culture may have different evolutionary dynamics. Significantly, our characterization of evolutionary archaeology seems to be in line with that of O’Brien and Lyman (2002:35), who note that whereas “some behavioral ecologists argue that much of the change we see archaeologically is attributable to phenotypic plasticity . . . most evolutionary archaeologists would argue that much of it is attributable to selection.”
In contrast to evolutionary archaeology, dual inheritance theory treats cultural evolution as if it is primarily effected through phenotypic plasticity rather than differential reproductive success. In dual inheritance theory, genes and culture are viewed as two distinct but interacting systems of information transmission (e.g., Ames 1996; Bettinger 1991; Boyd and Richerson 1985, 1993; Cavalli-Sforza and Feldman 1981; Durham 1979, 1982, 1990, 1991, 1992; Pulliam and Dunford 1980; Richerson and Boyd 1992; Runciman 2002; Shennan 1991, 2000, 2002). They both involve the transmission of phenotype-influencing information but operate via different mechanisms. The genetic system is based on reproduction, while the cultural one involves social learning. With this difference in mind, dual inheritance theorists hold that genetic evolution and cultural evolution are similar in that they are both based on the process that Darwin referred to as descent with modification, but they also accept that the nature of social learning is such that cultural evolution is influenced by forces that have no obvious equivalents in genetic evolution. Most notably, individuals can choose to copy practices from non-kin, and they are also able to modify or discard practices in the light of experience. The significance of these processes is that cultural evolution cannot be assumed to be always in step with genetic evolution. Sometimes it will be, but frequently it will not. Our ability to learn from non-kin means that cultural patterns will often not coincide with genetic patterns. Likewise, our ability to learn from other individuals and to pass on those behaviors to yet other individuals throughout our lives means that cultural evolution will often be faster than genetic evolution. Dual inheritance theory even allows for the possibility that the transmission of some cultural traits might be maladaptive from a genetic point of view (Boyd and Richerson 1985; Cavalli-Sforza and Feldman 1981; Shennan 2002).
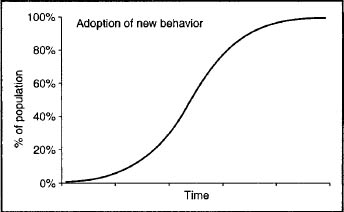
With respect to the mechanisms involved in cultural evolution, dual inheritance theorists foreground guided variation and biased transmission. Guided variation is the name given to the process in which one individual copies the behavior of another individual, modifies the copied behavior in the light of experience, and then passes on the modified behavior to a third individual. Biased transmission is the term used to refer to the process of model selection that dual inheritance theorists believe most individuals engage in when modifying their existing behaviors by copying others rather than relying entirely on their own experience. That is, dual inheritance theorists contend that individuals do not normally copy other individuals at random, but instead select models and behaviors on the basis of some criterion. Henrich (2001) has argued that this form of transmission is probably the basis for the S-curve that is characteristic of the diffusion of innovations in human populations, in which adoption is slow at first, rises to a peak, and then tails off as the innovation gradually reaches its ceiling in the population (figure 13.1). Multiple models of biased transmission have been developed (Boyd and Richerson 1985; Richerson and Boyd 1992). The main ones are direct bias, indirect bias, and frequency dependent bias. In direct bias, individuals evaluate alternative behaviors and choose among them. In indirect bias, individuals use some traits, such as those connoting health or prestige, to choose a cultural model, and then copy a range of the model’s behaviors. In frequency dependent bias, an individual copies a behavior on the basis of its frequency in the population; the most common form is thought to be conformist bias, which involves copying the behavior that is most widespread in the population. Significantly, guided variation and the various forms of biased transmission can, in principle, lead cultural evolution to be much faster than genetic evolution because they can affect the relative frequency of different cultural traits within a generation. In addition, the various forms of biased transmission can potentially lead to the “horizontal” or “oblique” movement of cultural traits between individuals and populations in the absence of gene flow. The corollary of this is that dual inheritance theory anticipates that the distributions of cultural traits and genes through time and space may differ in certain circumstances.
Figure 13.1. An S-curve, a theoretical plot of the shift of one behavior to another through time. It begins with the first instances of the new behavior, followed by an acceleration in the rate of change, gradual slowdown during the middle phases as the new behavior becomes the majority, and ends in the slowing of the conversion rate to zero as the new behavior asymptotically approaches 100 percent of the group.
As the above quote from O’Brien and Lyman (2002) indicated, human behavioral ecology also treats cultural evolution as if it is primarily effected through phenotypic plasticity. Linked to a substantial body of work in evolutionary biology (Krebs and Davies 1993), human behavioral ecology seeks to explain human behavior in terms of its ongoing adaptive significance (Borgerhoff Mulder 1991; Cronk 1991; Smith 2000; Smith and Winterhalder 1992; Smith et al. 2001; Shennan 2002; Winterhalder and Smith 2000). That is, human behavioral ecology attempts to understand how a particular behavior contributes to an individual’s reproductive success and/or that of his or her relatives given the prevailing environmental (including social) conditions. The key assumption of human behavioral ecology is that humans are sufficiently flexible that the vast majority of behavioral differences among them are primarily the result of diversity in environmental conditions rather than differences in genes or cultural inheritance (Shennan 2002; Smith 2000). Thus human behavioral ecologists concentrate on the relationship between behavioral strategies and ecological circumstances, and pay little attention to the mechanism(s) by which behavioral differences among individuals can potentially arise (Smith et al. 2001). One substantive consequence of this “black box” approach is that human behavioral ecology does not take into account population-level phenomena such as drift. Another is that human behavioral ecology makes no allowance for the possibility that some behavioral traits may not be adaptively significant because they were acquired as part of a package of traits. The first of these distinguishes human behavioral ecology from both dual inheritance theory and evolutionary archaeology. The second further distinguishes human behavioral ecology from dual inheritance theory.
This is not the place for an extended discussion of the pros and cons of the three schools of thought (see Bentley et al., chapter 8). Nevertheless, we believe it is important to make our theoretical preferences clear, and to briefly explain why we think the way we do. While the differences between dual inheritance theory, evolutionary archaeology, and human behavioral ecology are relatively small (Bettinger and Richerson 1996), we believe a combination of dual inheritance theory and human behavioral ecology (e.g., Shennan 2002) provides a more useful framework for studying cultural evolution than evolutionary archaeology, at least as it is practiced by the majority of its advocates. In our view, evolutionary archaeology’s contention that culture change can be explained solely in terms of natural selection and drift sensu stricto (i.e., where both processes involve differential reproductive success) is unconvincing. It may be the case that these processes were responsible for some of the changes observable in the archaeological record. However, as others have noted (Boone and Smith 1998; Runciman 2002), there is overwhelming evidence that cultural evolution often occurs too quickly for them to be the only processes involved. A good example highlighted by Boone and Smith (1998) is the shift from snowshoes to snowmobiles among the Boreal Forest Cree. Since this change is reported to have taken place in less than a generation (Winterhalder 1975), it simply cannot be the result of differential reproductive success, and natural selection and drift sensu stricto can therefore both be ruled out conclusively. Another good, and perhaps even more archaeologically relevant, example that was also highlighted by Boone and Smith (1998) is the adoption of the European horse by North American indigenous groups. Ramenofsky (1995) has argued that the horse spread because “individuals who owned horses reproduced in greater numbers than others” (p. 139), which in turn was a consequence of the horse outcompeting the dog as a means of transportation and also providing a significant advantage in hunting. While the notion that the horse conferred advantages with respect to transportation and hunting may be correct, the idea that the spread of horses was the consequence of differential reproductive success almost certainly is not. According to Ewers (1955), the horse was adopted by groups distributed across much of North America in less than two hundred years. Thus, even if we had no other information, a priori it is likely that the population was too large and dispersed, and the time frame too short, for natural selection sensu stricto to be the mechanism involved. Fortunately we do have other information available in the form of historical accounts, and it is clear from these that the primary processes involved in the spread of the horse among North American indigenous groups were trade, exchange, and raiding (Ewers 1955). As mechanisms of horizontal or oblique cultural transmission, trade, exchange, and raiding are aspects of phenotypic plasticity. As such, they do not fit well with evolutionary archaeology. In contrast, the spread of the European horse among Native Americans and the adoption of snowmobiles by the Boreal Forest Cree are easily accommodated within a dual inheritance/human behavioral ecology framework, since, as noted earlier, guided variation and biased transmission can in principle effect major cultural changes within a generation.
Evolutionary archaeology’s contention that culture change can be explained solely in terms of reproduction-based processes is also unconvincing when viewed in the light of empirical work on cultural transmission. Several studies suggest that copying from unrelated individuals can be an important process in cultural evolution. For example, in a classic experiment the psychologist Solomon Asch (1955) found that his subjects would agree with the conclusion of a group majority even when they could see clearly, with their own eyes, that the conclusion was incorrect (the experiment involved matching lines of different lengths drawn on paper). Asch’s (1955) study suggests that the urge to copy the majority view can be more powerful than one’s independent opinion. The importance of copying from unrelated individuals in human affairs is underscored by work reported by Simkin and Roychowdhury (2003) and Bentley (2006), which reveals clear patterns of such copying in one of the most unlikely places—academic publishing. This evidence too does not fit well with evolutionary archaeology but is easily accommodated within a dual inheritance framework/human behavioral ecology framework.
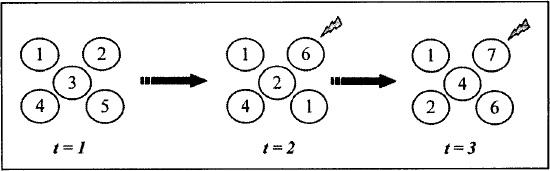
Figure 13.2. A simple representation of the neutral-trait model. Shown are five individuals per time step t for three successive time steps. At each time step, we refresh the population with new individuals, and each is given a new copy of a variant (represented by numbers inside the circles). Each variant is assigned a new value by either (1) copying a randomly selected individual from the previous time step, with equal probability of choosing any individual; or (2) inventing a new variant (gray lightning bolts) with probability µ, the mutation (innovation) rate per individual per time step t. After Bentley et al. (2007:fig. 1).
In the 1960s, geneticists made the surprising discovery that much DNA does not actually code for anything, and therefore evolves by drift rather than by natural selection. This discovery formed the basis of the neutral theory of evolution (Kimura 1983; Kimura and Crow 1964). Although it was originally intended as a theory for a particular form of genetic evolution, the neutral theory has been deployed in a variety of settings including the study of species distributions (Hubbell 2001), birdsong (Lynch and Baker 1994; Slater and Ince 1979), and cultural evolution (Bentley and Shennan 2003; Bentley et al. 2004; Bentley et al. 2007; Bettinger and Eerkens 1999; Brantingham 2003; Dunnell 1978; Eerkens and Lipo 2005; Hahn and Bentley 2003; Herzog et al. 2004; Kohler et al. 2004; Lipo 2001; Lipo et al. 1997; Meltzer 1981; Neiman 1995; Shennan and Wilkinson 2001).
In a simple version focusing on cultural evolution, the neutral model assumes there are N individuals, each characterized by a behavioral variant (figure 13.2). At each time step, each new individual copies a variant from an individual in the previous time step, such that any individual may be copied with equal probability. Innovation (analogous to genetic mutation) is added through the constant introduction of new, unique variants. The variable µ represents the innovation rate, in innovations per individual per time step. These two parameters, the number of individuals, N, and the mutation rate, μ, are the most important in the neutral model (Gillespie 1998). In the simple model, the number of individuals N per generation is kept constant and the innovation rate μ represents a fixed probability for each individual acquiring a new unique variant in a given generation. As determined simply by the product Nμ, the neutral model provides testable predictions concerning the change over time in the number and relative frequencies of different variants.
The neutral model can be easily simulated on a computer, with the variant frequencies being recorded over sample intervals of multiple generations or time steps. A set of these simulations focusing on cultural evolution has suggested three general characteristics in terms of how the frequencies of variants change through time:
1. A histogram of the relative popularity levels (frequencies) of variants yields a distribution that follows a mathematical function called a power law, for small values of the innovation rate μ (Bentley et al. 2004; Hahn and Bentley 2003).
2. If we follow a set of variants introduced in the same generation, over time the average of their frequencies stays the same, but the disparity (variance) in their frequencies increases (Hahn and Bentley 2003).
3. The set of most popular variants changes continually, at a rate that (to a first-order approximation) depends on the innovation rate but not significantly on the population size. This can be measured by the rate of turnover on any list of variants that are ranked according to popularity, as in “top 40” or “top 100” lists (Bentley et al. 2007).
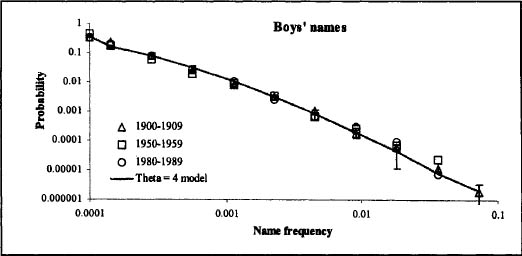
Figure 13.3. An example of a real-world data pattern that fits effect 1 described for the random-copying model (a histogram of the variant frequencies yields a power-law distribution, for small values of the innovation rate μ). The data are first names given to babies in the U.S. during the twentieth century. The plot shows frequencies of boys’ names versus expected frequencies based on the random-copying model. After Hahn and Bentley 2003: fig. 1a.
As an example of effect 1, figure 13.3 shows the fit of the random-copy model to the power law distribution of names for boys. Figure 13.4 shows how effect 2 can be seen among the first names given to babies in the 1900s in the United States, and following the frequencies of those names through the twentieth century. In a case study of registered purebred dog breeds in the United States, Herzog et al. (2004) demonstrated how Dalmatians could be identified as an exceptional case that cannot be explained by simple random copying. This was based on the fact that the popularity of Dalmatians exceeded the envelope of expected variations. Based on the rejection of the null hypothesis of simple random copying, Herzog et al. (2004) hypothesized that the sudden popularity increase of Dalmatians was due to the rerelease of the Disney movie 101 Dalmatians.
With regard to archaeological applications of the neutral theory, a formal distinction between artifact classes whose distributions are explained as a result of natural selection and those whose distributions are explained by drift (and hence neutral theory) was initially proposed by Robert Dunnell (1978). However, it was not until almost twenty years later that Neiman (1995) reported the first study in which neutral theory was formally applied to archaeological data. Neiman (1995) used the neutral theory to develop a series of predictions about the amount of variation to be expected in the decoration of a pottery assemblage if the motifs were neutral in terms of adaptation. He analyzed the variation among rim decorations from seven successive phases of the Woodland period in Illinois, and found that it matched the expectations of the neutral model. He concluded that the patterns of variation depended on changing levels of intergroup contact, which started low, increased, and then declined again. The time of highest interaction was also a time when exotic trade goods were widespread. Because the successful transmission of pottery-making traditions depends on long-lasting relationships between teacher and learner, Neiman (1995) suggested that the changing levels of intergroup contact related to changes in the level of long-term residential movement of potters between groups.
Since Neiman’s (1995) study appeared, a number of authors have reported work in which the neutral model has been applied to archaeological data. In view of the constraints on space, we will highlight just a few of these studies—Lipo et al.’s (1997) “Population Structure, Cultural Transmission, and Frequency Seriation,” Shennan and Wilkinson’s (2001) “Ceramic Change and Neutral Evolution: A Case Study from Neolithic Europe,” Kohler et al.’s (2004) “Vessels and Villages: Evidence for Conformist Transmission in Early Village Aggregations on the Pajarito Plateau, New Mexico,” and Eerkens and Lipo’s (2005) “Cultural Transmission, Copying Errors, and the Generation of Variation in Material Culture and the Archaeological Record.”
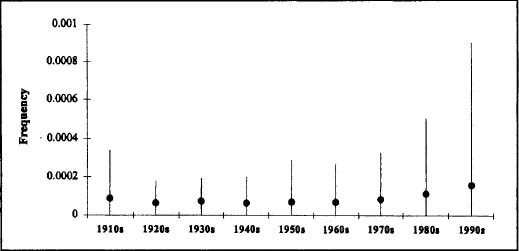
Figure 13.4. An example of effect 2 described for the random-copying model (the disparity in variants of the same age will increase over time, even as the average frequency stays the same). The plot shows the mean frequency (filled circles) and standard deviation (lines) over time of 99 male names that were new to the top 1,000 in the 1910s. After Hahn and Bentley 2003:fig. 3.
The goal of Lipo et al.’s (1997) article was to demonstrate the use of neutral traits to map patterns of interaction in the archaeological record. To begin with, they employed simulations to investigate the effects on the evolution of neutral traits of the differential interaction of individuals over space and time, and to explore the way in which these effects impact seriation analysis (see Webster, chapter 2). They found that the geographic distribution of a neutral trait through time is determined by the distribution of individuals in an environment and their frequency of interaction. They also found that when members of a population are free to interact equally over an area it is possible to produce a single, nearly perfect seriation for the whole area, whereas when the interactions among the members of a population are restricted it is impossible to produce such a seriation. With this in mind, they argued that seriation can be used as a tool to test hypotheses about the interaction of populations. Specifically, they argued that the failure of a set of assemblages to seriate should be taken as indicative of a lack of interactions among populations. Subsequently, Lipo et al. (1997) investigated the distribution of Mississippian-period decorated ceramics from the Memphis and St. Francis areas of the lower Mississippi valley with the aid of the large database of typed sherds published by Phillips et al. (1951). Lipo et al. (1997) began by seriating the assemblages into the largest groups possible. They then divided each group of assemblages that displayed a significant departure from unimodality into two groups. This process was repeated until all groups were unimodal, and therefore represented what Lipo et al. (1997) assumed to be the archaeological signatures of real communities. In order to test for the potentially confounding effects of sample size, Lipo et al. (1997) created new seriations by excluding assemblages that were judged too small. The resulting spatial groupings generally resembled the ones based on all of the assemblages, except in two cases where these new groupings overlapped and incorporated several of the former ones, which suggested that sample size had in fact partially determined the spatial grouping of assemblages. Lastly, Lipo et al. (1997) examined how estimates of interaction are affected by classification level. They accomplished this by collapsing classes to create more inclusive seriation groupings. Lipo et al. (1997) found increasingly larger groups patterned by distance, which is suggestive of interacting communities—in essence, that the degree of community interaction indicated by archaeological evidence depends on the scale of classification of the artifacts.
Shennan and Wilkinson (2001) applied Neiman’s (1995) neutral model in a study of patterns of pottery decoration from two settlements of the early Neolithic Linear Pottery culture in the valley of the Merzbach in western Germany. The Linear Pottery culture is thought to represent the archaeological trace of an early agricultural population that spread across much of Central Europe between about 5700 and 5400 B.C. It is especially well documented in the valley of the Merzbach as a result of excavations carried out in advance of strip mining. Shennan and Wilkinson (2001) found that the neutral model could predict the amount of variation in pottery decorations only in the early phases of occupation and not in the later phases. By using the neutral theory as their null hypothesis, Shennan and Wilkinson (2001) were able to identify the later Neolithic phases as an era of deliberate selection for novel decoration types, rather than the simple drift that characterized the early phases. Shennan and Wilkinson (2001) proposed that the later-phase potters were using this novelty to establish their own local identity with respect to neighboring groups.
Kohler et al. (2004) used the neutral model and previously compiled pottery data to examine cultural evolutionary processes at Burnt Mesa Pueblo, a Late Coalition period village in New Mexico (Gray 1992). Kohler et al. (2004) subdivided the pottery into eighty-three distinct styles, based on general descriptions of the surface designs (e.g., lines only, solid designs, hatched designs, then further data on line type, hatch type, curved line). They then calculated estimates of the product Nμ (population size times the innovation rate), which under the neutral model should be directly proportional to the diversity of variants. By calculating Nμ and comparing it to the diversity of variants, Kohler et al. (2004) were able to test whether the neutral model described stylistic change at Burnt Mesa Pueblo. The challenge is that for a typical archaeological assemblage, the population size of the community is difficult to estimate, and innovation rate is practically impossible to measure directly. Fortunately, as Shennan and Wilkinson (2001) pointed out, N can be estimated indirectly through the number of households (assuming one potter per household) and/or the number of vessels in each period, and the innovation rate μ in a chronological phase can be indirectly estimated by the number of new decorations divided by the total number of vessels for the phase. Using this method, Kohler et al. (2004) found less diversity in ceramic style than would be expected under the neutral model. Specifically, they found that the innovation rate at Burnt Mesa Pueblo declined steadily through the period of its occupation. Kohler et al. (2004) concluded from this that a conformist bias had developed in the way pottery designs were made.
Eerkens and Lipo (2005) developed models to account for variation in continuously measured variables in archaeological assemblages such as projectile point dimensions. They proposed that variation is created either by simple copying error or by cognitive process-dependent activities such as innovation or invention. They assumed that variation created through copying error should be random and relatively small in magnitude. With regard to the magnitude of copying error, they specifically assumed that it should be below the level at which people are aware there is a difference. They also assumed that variation created through cognitive mechanisms should be directional and of greater magnitude than copying error. The authors then modeled assemblage variation related to several transmission processes using Markov chain simulations where random processes are followed in successive generations or time steps. They found that in unbiased transmission the amount of variation increases with continued copying errors compounding variation over time, but the average value for the trait stays the same. They also found that biased transmission such as conformist and prestige-biased transmission acts to dampen the amount of variation expressed over time. Subsequently, Eerkens and Lipo (2005) used their simulation findings to interpret the results of analyses of morphological variation in projectile points from the Owens valley of California and Woodland period ceramics from Illinois. Using obsidian hydration ages associated with the projectile points, they examined the distribution of coefficient of variation values for basal width and thickness over time. They found that variation associated with basal width decreases over time, indicating that some variation-reducing process acted to diminish variation. Variation in thickness tended to increase over time, fitting an unbiased transmission model. With regard to the Woodland period ceramics, thickness and diameter measures were taken from assemblages associated with dated features, and coefficient of variation values calculated. It was found that during some periods the variation in thickness was consistent with the copying error model, while in others there were increases in sherd thickness beyond the simulated limits of copying error. Eerkens and Lipo (2005) interpreted the latter as evidence for the operation of selection. The most noteworthy finding with respect to pot diameter is that it showed an increase in variation of nearly 16 percent during the final four-hundred-year period of the sequence. This is much greater than the simulated variation due to copying error. Accordingly, Eerkens and Lipo (2005) suggested that a variation-increasing mechanism such as invention must have been operating.
In recent years researchers have also begun to use selection-based genetic models to cast light on archaeological problems. For example, Shennan (2001) used population genetic modeling to investigate the impact of population size on cultural evolution when innovations affect fitness. In this study Shennan (2001) employed two variations on a population genetics model developed by Peck et al. (1997) to assess the relative benefits of sexual and asexual reproduction. In Peck et al.’s model, mutations can be either beneficial or deleterious; there is a correlation between an allele’s fitness prior to mutation and its post-mutation fitness; and many mutations produce only very small changes in fitness. For the first version, Shennan (2001) altered Peck et al.’s (1997) model so that transmission was only possible from one “cultural parent” to one “cultural offspring.” To produce his second model, Shennan (2001) modified Peck et al.’s (1997) model to allow transmission between individuals belonging to different generations where the older individual is not the biological parent of the younger individual. In simulation trials Shennan (2001) found a marked increase in the mean fitness of the population as effective population size increased. In the trials of the first model there was a 10,000-fold increase in the mean fitness value of the population as effective population size increased from five to fifty. In trials of the second model in which cultural traits were adopted from nonbiological parents 5 percent of the time, the population’s mean fitness value increased a thousandfold as the effective population size increased from five to twenty-five, and then increased by around five times as effective population size increased from twenty-five to seventy-five. Shennan’s (2001) simulation studies showed that larger populations have a major advantage over smaller ones when it comes to cultural innovation due to the decreasing role of sampling effects as populations get larger. When effective population size is large, there is a far greater probability of fitness-enhancing cultural innovations being maintained and deleterious ones being lost than when effective population size is small. In the latter situation, innovations that are maintained tend to be less beneficial in terms of reproduction and also less attractive for imitators.
Recently Henrich (2004) has presented a rather different model of the impact of population size on cultural evolution, and in particular the opportunities it offers for cumulative change. His model assumes that when learning cultural behaviors, in particular complex skills, individuals will try to copy the best practitioner of that skill within their population. Most people will not do as well as the best, but the mean margin of failure is likely to be variable—small for easy things, larger for more difficult ones. On the other hand, there will be variability from one person to the next in attempts to imitate. Occasionally an individual may strike it lucky and, in a failed attempt to imitate, produce an outcome that gives a better result than the previous best. This then becomes the new goal for the rest of the population to aspire to. As a result, so long as the mean margin of failure does not increase, the level of the whole population will be improved. If the mean margin of failure is high and the variation between individuals is small, then improvement is unlikely. Thus the likelihood of cumulative cultural evolution is partly dependent on the ratio between mean margin of failure and the amount of interindividual variation. It is also partly dependent on population size, since in large populations even improbable events—in this case arriving at a behavior that gives a better result than the previous best—occur now and again, and obviously the larger the population the more likely this is. Depending then on the ratio between the mean margin of failure and the amount of interindividual variation, a larger or a smaller population size will be required for cumulative cultural evolution to take place. It follows that, for a given ratio, if the size of the interacting population changes for some external reason then this will affect the rate of cumulative cultural evolution. If population increases, then the probability of cumulative improvement increases. On the other hand, if it decreases then it is likely that a process of cultural devolution will take place, because the probability of someone improving on the existing situation, or even equaling the current best, is small. Thus in the next generation the best individual to copy is likely to be slightly worse than in the generation before, and this process will be repeated through the generations, until some equilibrium is reached. Henrich (2004) suggested that this model explains the apparent loss of cultural adaptations in Tasmania after it became separated from the Australian mainland with rising sea levels at the end of the last Ice Age, since this isolation meant that the Tasmanians were no longer part of a larger interacting continental population.
In our view, the studies described above demonstrate the considerable potential of population genetics models, when suitably adjusted, to shed light on cultural evolution. Crucially, such models provide baselines of great epistemological value. Where cultural patterns agree with the patterns predicted by a given model, we can invoke the principle of parsimony and discount processes that are more complicated than the modeled process. In contrast, where the modeled and actual patterns disagree, we can legitimately disregard the modeled process and seek a more complex explanation. As such, the application of population genetics-based cultural evolutionary models allows us to narrow the range of possible explanations for cultural patterns in a controlled manner.
First outlined in the 1950s (Hennig 1950, 1965, 1966), cladistics is currently the dominant method of phylogenetic reconstruction used in evolutionary biology and paleobiology (Kitching et al. 1998; Quicke 1993; Schuh 2000; Smith 1994; Wiley et al. 1991). Based on a null model in which new taxa arise from the bifurcation of existing ones, cladistics defines phylogenetic relationship in terms of relative recency of common ancestry. Two taxa are deemed to be more closely related to one another than either is to a third taxon if they share a common ancestor that is not also shared by the third taxon. Exclusive common ancestry is indicated by shared evolutionarily novel or derived character states. Two taxa are inferred to share a common ancestor to the exclusion of a third taxon if they exhibit shared derived character states that are not also exhibited by the third taxon.
In its simplest form, cladistic analysis proceeds via four steps. First, a character state data matrix is generated. This shows the states of the characters exhibited by each taxon. Next, the direction of evolutionary change among the states of each character is established. Several methods have been developed to facilitate this, including communality (Eldredge and Cracaft 1980), ontogenetic analysis (Nelson 1978), and stratigraphic sequence analysis (Nelson and Platnick 1981). Currently the favored method is outgroup analysis (Arnold 1981), which entails examining a close relative of the study group. When a character occurs in two states among the study group, but only one of the states is found in the outgroup, the principle of parsimony is invoked, and the state found only in the study group is deemed to be evolutionarily novel with respect to the outgroup state. Having determined the probable direction of change for the character states, the next step in a cladistic analysis is to construct a branching diagram of relationships for each character. This is done by joining the two most derived taxa by two intersecting lines, and then successively connecting each of the other taxa according to how derived they are (figure 13.5). Each group of taxa defined by a set of intersecting lines corresponds to a clade, and the diagram is referred to as a cladogram or tree. The lines that form a cladogram are usually called branches except for the one at the base, which is referred to as the root. The points of intersection between branches are called nodes. The last step of the process is to compile an ensemble cladogram from the character cladograms. Ideally, the distribution of the character states among the taxa will be such that all the character cladograms imply relationships among the taxa that are congruent with one another. Normally, however, a number of the character cladograms will suggest relationships that are incompatible. This problem is overcome by generating an ensemble cladogram that is consistent with the largest number of characters and therefore requires the smallest number of ad hoc hypotheses of character appearance or homoplasies to account for the distribution of character states among the taxa.
Recently a number of researchers have begun to apply cladistics and related biological phylogenetic methods to cultural data to shed light on events in antiquity (Atkinson and Gray 2005; Bryant et al. 2005; Buchanan and Collard, in press; Cochrane 2004; Darwent and O’Brien 2005; Eerkens et al. 2005; Foley 1987; Foley and Lahr 1997, 2003; Forster and Toth 2003; Gray and Atkinson 2003; Gray and Jordan 2000; Greenhill and Gray 2005; Harmon et al. 2005; Holden 2002, 2005; Holden et al. 2005; O’Brien and Lyman 2003b; O’Brien et al. 2001, 2002; Rexová et al. 2003; Robson-Brown 1996). This approach acknowledges that reconstructing human population history from linguistic and archaeological data shares the same problems as reconstructing the evolutionary history of a group of species (Foley 1987; Kirch and Green 1987). In both cases, the key challenge is to distinguish the similarities resulting from shared ancestry (homologies) from those due to mechanisms other than shared ancestry (homoplasies). While the processes that generate biological and cultural homologies and homoplasies are not the same (e.g., gene transfer versus cultural transmission), the epistemology and ontology of establishing ancestral relationships is general enough to warrant the application of phylogenetic methods to cultural data. Most significantly, in both cases a model is sought that explains the distribution of resemblances among a group of taxa in the absence of prior knowledge of how those resemblances arose. The bifurcating tree model is used because it is the simplest, and therefore the most defensible, way of linking taxa together. Once a tree model has been generated for a group of taxa, is it possible to classify the similarities among them as homologous or homoplastic. Homologous similarities support relationships that are compatible with the tree model, whereas homoplastic ones suggest relationships that conflict with the tree model.
Figure 13.5. An example of a tree of evolutionary relationships generated via cladistics, together with the character state data matrix from which it was derived. Trees of evolutionary relationships generated with cladistics are usually referred to as “cladograms.” They are read from the tips to the root. Thus, the cladogram shown indicates that taxa B and C form a monophyletic group to the exclusion of taxon A based on the shared possession of derived character states for characters 3 and 4. It also suggests that taxa A, B, and C form a monophyletic group based on the shared possession of derived character states for characters 1 and 2. Taxon C is the most derived taxon, having the derived states for characters 5, 6, and 7 in addition to the other derived characters. Character 7 is homoplastic as it is in a derived state in taxa A and C, even though taxa A and C are not directly related through one common ancestor. Two equally parsimonious solutions are available to resolve this character data matrix; in the cladogram shown above, taxa B and C are shown more closely related and forming a clade to the exclusion of taxon A. In the second solution taxa A and C form a clade to the exclusion of taxon B. From Buchanan and Collard (2007), copyright Elsevier (2007), reprinted with permission.
This approach is illustrated by the historical linguistic studies of Gray and Jordan (2000), Holden (2002), and Gray and Atkinson (2003), and the archaeological work of O’Brien and colleagues (O’Brien and Lyman 2003b; O’Brien et al. 2001, 2002) and Buchanan and Collard (in press). Gray and Jordan (2000) employed cladistic methods to assess the two main competing models regarding prehistory in the Pacific, the express train model and the entangled bank model (see Bellwood, chapter 14). The former suggests a rapid dispersal of Austronesian speakers from a homeland in Taiwan around 6,000 years ago through Island Melanesia and into the Polynesian islands of the remote Pacific, whereas the latter contends that the Polynesian colonizers derived from a population in Island Melanesia that had been there for tens of thousands of years. In the entangled bank model, the cultural and linguistic patterns among Polynesians are the complex result of not just their colonization, but also founder’s effects associated with original colonization, and the continued cultural contact between different islands during the subsequent millennia, with genetic, linguistic, and cultural traits transmitted at varying intensities between populations. In the entangled bank model linguistic patterns largely reflect human interaction and continued cultural transmission rather than the phylogenetic history of the language speakers described by the express train model. To test these conflicting models, Gray and Jordan (2000) used a cladistics computer program to produce a phylogeny of Pacific languages, onto which they then mapped the prehistoric events suggested by the express train model. They found a close fit between the chronological stages of the express train model and the branching pattern of their language phylogeny. In that languages that were closely related in the phylogeny were not necessarily close geographically. Gray and Jordan (2000) concluded that these linguistic patterns result predominantly from colonizing migrations of the language speakers rather than cultural contact since the time of initial colonization.
Holden (2002) conducted a comparable analysis in which she used maximum parsimony analysis to reconstruct the relationships among seventy-five Bantu and Bantoid African languages from ninety-two items of basic vocabulary. As in the Polynesian case, some researchers contend that the Bantu languages evolved rapidly during the Neolithic and Iron Age with the colonization of farmers into sub-Saharan Africa, while others hold that the evolution is more the result of diffusion of Bantu words among neighboring speech communities. Holden’s (2002) analysis returned a relatively small set of possible phylogenetic trees in support of the tree model of Bantu language history, and found these trees to be consistent with the model for the spread of farming in sub-Saharan Africa constructed by archaeologists through chronological analysis of pottery. Holden (2002) concluded, therefore, that the dispersal and diversification of the Bantu languages was linked to the expansion of farming during the Neolithic and Iron Age, and that since that time Bantu-speaking communities have not moved to any great extent.
Gray and Atkinson (2003) employed phylogenetic analysis of Indo-European languages to test the two main competing hypotheses for Indo-European origins, one that Indo-European languages spread with Kurgan pastoralists beginning around 6000 B.P. (Gimbutas 1973), and the other that the language family expanded with the spread of agriculture from Anatolia around 9500–8000 B.P. (Renfrew 1987; see Bellwood, chapter 14). Working with a restricted list of essential vocabulary words across Indo-European languages both past and present, Gray and Atkinson (2003) reconstructed a set of likely phylogenetic trees of their evolution, and then used statistical methods to estimate their divergence times. Since the overwhelming majority of these trees produced estimated dates between 7800 and 9800 B.P. for Indo-European origins, consistent with the estimates Renfrew (1987) obtained from the archaeology, Gray and Atkinson’s (2003) phylogenetic analysis strongly supports Renfrew’s (1987) Anatolian farming hypothesis.
O’Brien et al.’s (2001, 2002; O’Brien and Lyman 2003b) archaeological application of cladistics focused on the long-standing problem of the evolution of projectile point form in the southeastern United States during the Paleoindian period (ca. 11,500 to 10,000 14C yr B.P.). O’Brien et al. began by recording three qualitative and five quantitative characters on a sample of 621 specimens representing a range of projectile point types, including Clovis, Dalton, and Cumberland. They then subjected the specimens to paradigmatic classification in order to cluster them into taxa with unique combinations of character states. In the next part of the study, O’Brien et al. carried out a cladistic analysis of the seventeen taxa that contained at least four specimens. One of the seventeen taxa was selected as the outgroup on the basis of least-step occurrence seriations and chronological considerations; the remainder were treated as the ingroup. The cladistic analysis yielded a single most parsimonious cladogram. O’Brien et al. evaluated the fit between the cladogram and the data set with a goodness-of-fit index called the Consistency Index (CI). This index ranges between 1.0 and 0.0, with values close to 1 indicating a good fit between the cladogram and the data set and values close to 0 indicating a poor fit. The cladogram obtained by O’Brien et al. had a CI of 0.59, which suggests that it is a reasonable depiction of the relationships among the taxa. In the final part of their study, O’Brien et al. used the cladogram to investigate the character state changes that occurred in the course of the evolution of Paleoindian projectile point form.
Buchanan and Collard’s (in press) study also focused on the archaeology of the Paleoindians. Specifically, they applied cladistic techniques to qualitative and quantitative data from a North America–wide sample of projectile points in order to test competing models of Early Paleoindian colonization and adaptation. The archaeological cultures of the Early Paleoindian period, which include the well-known Clovis culture, represent the first well-documented indications of human occupation in North America. Buchanan and Collard (in press) employed three sets of tests to determine whether or not a phylogenetic signal was present in the size-corrected projectile point data. Results from a permutation tail probability test (p = 0.0001), goodness-of-fit statistics (CI = 0.56; RI = 0.67), and the phylogenetic bootstrap (producing clade support of 52–84 percent for branches) indicated that a significant phylogenetic signal exists in the data. Subsequently, the cladograms derived from the maximum parsimony and bootstrap analyses were compared with cladograms constructed to represent different colonization routes, cultural diffusion, and environmental adaptation hypotheses. The colonization models analyzed include the ice-free corridor model, the Northwest Coast model, and Stanford and Bradley’s (2002; see also Bradley and Stanford 2004) Solutrean model. The strength of fit between the observed and hypothetical cladograms was assessed using the Kishino-Hasegawa (K-H) test (Kishino and Hasegawa 1989). In this test, a p-value is calculated for the length difference between a hypothetical cladogram and an observed cladogram by comparing it to a distribution of length differences obtained from a randomly generated sample of cladograms. Buchanan and Collard (in press) found that the four most-parsimonious cladograms and the bootstrap cladogram were all significantly different from the hypothetical cladograms. Therefore, none of the hypotheses was as well supported by the data set as the most parsimonious cladogram. Comparison of cladogram lengths then was used to assess the fit of the hypothetical cladograms to the observed cladograms (Jordan and Shennan 2003). Using this approach, the ice-free corridor entry model was found to best account for the structure in the data.
Another group of researchers has applied cladistics and related phylogenetic methods to cultural data in an effort to resolve an ongoing debate about the processes involved in cultural evolution at the population level (Collard and Shennan 2000; Collard et al. 2005; Jordan and Shennan 2003, 2005; Moylan et al. 2005; Shennan and Collard 2005; Tehrani and Collard 2002). So far, the debate in question has concentrated on two competing hypotheses, which have been termed the branching hypothesis (also known as the demic diffusion or phylogenesis hypothesis) and the blending hypothesis (also known as the cultural diffusion or ethnogenesis hypothesis) (Bellwood 1996; Collard and Shennan 2000; Kirch and Green 1987; Moore 1994, 2001; Tehrani and Collard 2002). According to the former, the cultural similarities and differences among human populations are primarily the result of cultural assemblages dividing as the communities that produce them repeatedly grow and split. The branching hypothesis predicts that the similarities and differences among cultures can be represented by a cladogram, and that there will be a strong association between cultural variation and linguistic, morphological, and genetic patterns (Ammerman and Cavalli-Sforza 1984; Renfrew 1987). In contrast, supporters of the blending hypothesis (Dewar 1995; Moore 1994, 2001; Terrell 1988, 2001; Terrell et al. 1997, 2001) contend that it is unrealistic “to think that history is patterned like the nodes and branches of a comparative, phylogenetic, or cladistic tree” (Terrell et al. 1997:184). Instead, they argue that the biological, linguistic, and cultural evolution of our species is best characterized by “a constant flow of people, and hence their genes, language, and culture, across the fuzzy boundaries of tribes and nations” (Moore 2001:51). The blending hypothesis predicts that a reticulated graph can best represent the similarities and differences among cultures (Terrell 2001), and that there will be a close relationship between cultural patterns and the frequency and intensity of contact among populations. Other models have been proposed (e.g., Boyd et al. 1997), but to date these have received little attention in the literature.
The researchers who have applied cladistics to cultural data in the context of the phylogenesis/ethnogenesis debate have done so on the grounds that it enables a quantitative estimate of the relative contribution of the two processes to any given data set to be obtained (Collard and Shennan 2000; Collard et al. 2005; Jordan and Mace 2005; Jordan and Shennan 2003, 2005; Shennan and Collard 2005; Tehrani and Collard 2002). They have argued that in the absence of prior knowledge of how the resemblances among a group of cultural assemblages arose, a reasonable course of action is to fit the bifurcating tree model to characters derived from the assemblages, and then determine how many character states fit the tree and how many do not. Those that fit the tree model are assumed to have been transmitted vertically (in the sense of traits being passed from an ancestral group to its descendants, rather than necessarily from parents to children) and are therefore the result of phylogenesis, while those that do not are assumed to have been transmitted horizontally and are therefore the result of ethnogenesis. As was the case with using cladistics to investigate events in prehistory, this course of action is justified on the grounds that the bifurcating tree model is the simplest, and therefore the most defensible, way of linking a group of taxa together. An additional reason for employing cladistics to measure the relative contribution of vertical and horizontal transmission to cultural data sets as opposed to the regression-based approaches that conventionally have been used to investigate to cultural evolutionary processes (Guglielmino et al. 1995; Moore and Romney 1994, 1996; Roberts et al. 1995; Welsch 1996; Welsch et al. 1992) is that the latter may inflate the significance of ethnogenesis. As noted earlier, geographic proximity is usually employed as the proxy for ethnogenesis. However, phylogenesis can also be expected to correlate with geographic proximity, since sister groups are likely to be nearest neighbors. Thus the use of geographic proximity solely as a proxy for ethnogenesis will overstate the latter’s importance and understate the importance of phylogenesis.
Again, in view of the constraints on space, we will highlight just a few studies in which cladistics has been applied to cultural data with a view to shedding light on the phylogenesis/ethnogenesis debate—Collard and Shennan’s (2000) “Ethnogenesis Versus Phylogenesis in Prehistoric Culture Change: A Case-Study Using European Neolithic Pottery and Biological Phylogenetic Techniques,” Tehrani and Collard’s (2002) “Investigating Cultural Evolution through Biological Phylogenetic Analyses of Turkmen Textiles,” and Jordan and Shennan’s (2003) “Cultural Transmission, Language, and Basketry Traditions amongst the Californian Indians.”
Collard and Shennan’s (2000) study examined decorated pottery from seven multiphase early Neolithic Linear Pottery culture settlements in the valley of the Merzbach stream on the Aldenhovener Platte of western Germany. Their aim was to establish whether within this small area there was evidence for processes of cultural branching or blending. Their first set of analyses focused on the assemblages from the four settlements that have evidence for occupation throughout the whole of the ten-phase period. They conjectured that, if the phylogenesis hypothesis is correct, analyses of the assemblages should divide the settlements into the same groups in consecutive phases. On the other hand, if the ethnogenesis hypothesis is accurate, the analyses should place the settlements into different groups in consecutive phases. The four settlements were divided into the same groups in six of the instances in which consecutive phases could be compared. In the remaining three instances, the settlements were divided into different groups in consecutive phases. These results are not wholly compatible with either hypothesis. Rather, they indicate that phylogenesis and ethnogenesis were both involved in the generation of the pottery assemblages. Collard and Shennan’s (2000) second set of analyses focused on three instances in which a new pottery assemblage appears. They reasoned that, if the phylogenesis hypothesis is correct, then the newly founded assemblages should have a single parent assemblage in the preceding phase. Conversely, if the ethnogenesis hypothesis is accurate, then the newly founded assemblages should have multiple parent assemblages. The second set of analyses supported the phylogenesis hypothesis rather than the ethnogenesis hypothesis. A newly founded assemblage was strongly linked with a single parent assemblage in two of the analyses. The results of a third analysis were more ambiguous, but the simplest interpretation of them also supported the notion that the newly founded assemblages had a single parental assemblage in the preceding phase. Overall, therefore, Collard and Shennan’s (2000) analyses of the Merzbach valley Early Neolithic pottery supported the phylogenesis hypothesis more strongly than the ethnogenesis hypothesis. New settlements seem to arise as a result of the fissioning of a single existing settlement rather than from the amalgamation of people from several existing ones. While they do show evidence of influence from adjacent settlements in their pottery, there tends to be a microcultural continuity.
Tehrani and Collard’s (2002) study examined decorated textiles produced by Turkmen groups between the eighteenth and twentieth centuries. Two sets of cladistic analyses were carried out. The first focused on the period before the Turkmen were incorporated into the Russian empire. These analyses indicated that in the precolonial period the evolution of Turkmen textile designs was dominated by branching. A randomization procedure (the permutation tail probability test) suggested that the data contain a phylogenetic signal, and parsimony analysis indicated that the data fit the bifurcating tree model associated with cultural branching reasonably well. The fit between the model and data was not perfect, indicating that blending played a role in the evolution of Turkmen culture. However, goodness-of-fit statistics (CI, Retention Index) and a second randomization procedure (bootstrapping) suggested that blending was markedly less important than branching. According to the goodness-of-fit statistics, about 70 percent of the similarities among the assemblages are homologous, and approximately 30 percent are homoplastic. This is compatible with the borrowing of designs and motifs being responsible for a third of interassemblage resemblances, although the possibility of independent invention as a source of homoplastic similarities cannot be completely discounted. Tehrani and Collard’s second set of analyses dealt with weavings produced after the defeat of the Turkmen by the Russian military. These analyses suggested that the social and economic changes experienced by the Turkmen after their incorporation into the Russian empire led to a greater role for blending in Turkmen cultural evolution. Branching remained the dominant cultural evolutionary process, but the importance of blending increased. The goodness-of-fit statistics indicated that ~60 percent of the interassemblage resemblances are homologous, and ~40 percent are homoplastic. Thus there is a 10 percent increase in the number of homoplastic resemblances among the woven assemblages from the period of Russian domination. This is consistent with more intertribal borrowing of designs and motifs, but again independent invention cannot be entirely discounted as a source of the homoplasies. Tehrani and Collard concluded that the two sets of analyses supported the branching hypothesis more strongly than the blending hypothesis.
Jordan and Shennan (2003) reached a contrasting conclusion. These researchers used cladistics to examine variation in Californian Indian basketry in relation to linguistic affinity and geographic proximity. They carried out three sets of cladistic analyses. In the first, they used the permutation tail probability test to determine whether or not their basketry data sets (coiled baskets, twined baskets, all baskets) contain a phylogenetic signal. These analyses suggested that a significant phylogenetic signal is present in all three data sets. In the second set of analyses, Jordan and Shennan (2003) used the CI to assess the fit between the data sets and the bifurcating tree model. These analyses suggested that the phylogenetic signal detected by the permutation tail probability test is weak. In the third set of analyses, Jordan and Shennan (2003) used the K-H test to assess the fit between the data sets and trees reflecting linguistic relationships, geographic distance, ecological similarity, and adjacency. This test enabled them to distinguish between two different potential sources of homoplasy—independent invention and blending. In an analysis of the complete sample of baskets, the fit between the data set and the adjacency tree was considerably better than the fit between the data set and the other trees. This suggests that blending had a bigger impact on the distribution of similarities and differences among the basketry assemblages than branching or adaptation to local environments. In an analysis of just the coiled baskets, blending was also found to play a more significant role than branching or adaptation to local environments. The analysis of the twined baskets contrasted with the preceding analyses in that the language tree fitted the data set better than the other trees. This suggests that branching was more important in generating the twined baskets than blending or adaptation to local environments. Jordan and Shennan concluded on the basis of these results, and the results of a range of multivariate analyses, that the evolution of Californian Indian baskets is best explained by ethnogenesis.
In our view, the studies discussed in this section suggest that cladistics and related phylogenetic methods can be a useful tool for tackling certain problems concerning cultural evolution. It offers a well-characterized model that can be used to analyze material culture and linguistic data sets. Where the fit between a cultural data set and the tree model is close, we can invoke the principle of parsimony and legitimately conclude that the similarities and differences among the cultural units are best explained by vertical transmission. On the other hand, where there are numerous homoplasies and the fit between a cultural data set and the tree model is consequently poor, we can infer that horizontal transmission and/or convergent evolution played a more important role in generating the similarities and differences among the cultural units than branching. The instances of homoplasy can then be investigated with other methods that are not based on the bifurcating tree model, such as spectral analysis (Hendy and Penny 1992) or split decomposition (Bandelt and Dress 1992; Dopazo et al. 1993; Dress et al. 1996; Huson 1998).
Regarding further applications of phylogenetic methods to problems in archaeology, there are some cases where we can predict quite confidently that the bifurcating tree model is likely to be relevant and where the data patterns seem to point in this direction even though formal analyses remains to be done. Thus, as we mentioned earlier, the spread of farming into Central Europe seems to have involved a fast initial colonization process followed by more local population expansion, mirrored by the increasing regionalization of pottery styles. A similar pattern of initial uniformity followed by increasing regionalization seems apparent in the case of Bell Beakers, but it is by no means clear that the mechanisms are the same, since the pattern seems to be restricted to a limited part of the Bell Beaker cultural inventory, especially the Beaker vessels themselves. Here, and more certainly in subsequent periods that seem to have had large sedentary populations, it seems likely that cultural innovations relating to different aspects of social life and originating in different places would have spread through those populations more or less independently, resulting in multiple cultural lineages rather than a single cultural “core” (e.g., Boyd et al. 1997).
Lastly, we think it is worth reiterating that the phylogenesis/ethnogenesis debate is concerned with cultural evolution at the group level as opposed to cultural transmission within groups. There is undoubtedly a link between within-group cultural transmission and among-group cultural evolution, but it cannot be assumed to be a straightforward one. In principle, cultural transmission among individuals within a group can be vertical, oblique, and/or horizontal and still be compatible with both phylogenesis and ethnogenesis. For example, even if cultural transmission within each group in a sample is predominantly horizontal, cultural evolution at the population level may still be dominated by phylogenesis, providing the amount of trade, exchange, copying, and/or intermarriage among groups is limited. Conversely, even if within-group cultural transmission is predominantly vertical, cultural evolution at the population level may still be dominated by ethnogenesis, providing there is a large amount of trade, exchange, copying, and/or intermarriage among groups. Thus finding a good fit between the distribution of similarities and differences among a set of cultural assemblages and the bifurcating tree model does not necessarily mean that cultural transmission within the groups that produced the assemblages was dominated by vertical transmission. Likewise, a poor fit between the distribution of similarities and differences among a set of cultural assemblages and the bifurcating tree model does not necessarily mean that cultural transmission within the groups that produced the assemblages was dominated by horizontal and/or oblique transmission.
Applying evolutionary biological methods to cultural data with a view to shedding light on cultural evolution is a relatively new approach. Most work on cultural evolution involves identifying patterns in data sets and then trying to determine which processes are likely to have produced those patterns. Evolutionary biological methods are advantageous in this regard because they tend to be based on well-characterized process models. If the method indicates a good fit between the data and the model, we can invoke the principle of parsimony and discount processes that are more complicated. Conversely, if the method suggests that the fit between the data and the model is poor, we can legitimately disregard the modeled process and seek a more complex process to explain the data. Evolutionary biological methods, therefore, allow us to select process explanations for cultural patterns in a rigorous manner. A further important benefit of applying evolutionary biological methods to cultural data is that it allows archaeologists, anthropologists, and historical linguists to provide information about patterns and processes of cultural evolution that can be linked with contemporary genetic data and with the increasingly available evidence for past human mobility from studies of stable isotopes and ancient DNA. We believe this combination has the potential to give us unparalleled insights into the interrelations among populations and the genetic, linguistic, and nonlinguistic cultural attributes associated with them.
MC, SJS, and RAB thank the Arts and Humanities Research Council for supporting their work on cultural evolution. BB is grateful for the support provided to him by the National Science Foundation through its postdoctoral fellowship (# 0502293). We are grateful to Ethan Cochrane and Jamshid Tehrani for constructive criticisms of earlier drafts of the manuscript.
Abbott, Alysia L., Robert D. Leonard, and George T. Jones. 1996. Explaining the change from biface to flake technology: A selectionist application. In H. D. G. Maschner, ed., Darwinian archaeologies, 33–42. New York: Plenum.
Ames, Kenneth M. 1996. Archaeology, style, and the theory of coevolution. In H. D. G. Maschner, ed., Darwinian archaeologies, 109–131. New York: Plenum.
Ammerman, Albert J., and L. Luca Cavalli-Sforza. 1984. The Neolithic transition and the genetics of populations in Europe. Princeton: Princeton University Press.
Arnold, E. N. 1981. Estimating phylogenies at low taxonomic levels. Zeitschrift für zoologische Systematik und Evolutionsforschung 19: 1–35.
Asch, Solomon E. 1955. Opinions and social pressure. Scientific American 193: 31–35.
Atkinson, Quentin D., and Russell D. Gray. 2005. Are accurate dates an intractable problem for historical linguistics? In C. P. Lipo, M. J. O’Brien, M. Collard, and S. J. Shennan, eds., Mapping our ancestors: Phylogenetic approaches in anthropology and prehistory, 269–296. New Brunswick, NJ: Transaction.
Bandelt, Hans-Jürgen, and Andreas W. M. Dress. 1992. Split decomposition: A new and useful approach to phylogenetic analysis of distance data. Molecular Phylogenetics and Evolution 1: 242–252.
Bellwood, Peter. 1996. Phylogeny vs. reticulation in prehistory. Antiquity 70: 881–890.
Bentley, R. Alexander. 2006. Academic copying, archaeology, and the English language. Antiquity 80: 196–201.
Bentley, R. Alexander, Matthew W. Hahn, and Stephen J. Shennan. 2004. Random drift and culture change. Proceedings of the Royal Society B 271: 1443–1450.
Bentley, R. Alexander, Carl P. Lipo, Harold A. Herzog, and Matthew W. Hahn. 2007. Regular rates of popular culture change reflect random copying. Evolution and Human Behavior 28: 151–158.
Bentley, R. Alexander, and Stephen J. Shennan. 2003. Cultural evolution and stochastic network growth. American Antiquity 68: 459–485.
Bettinger, Robert L. 1991. Hunter-gatherers: Archaeological and evolutionary theory. New York: Plenum.
Bettinger, Robert L., and Jelmer W. Eerkens. 1999. Point typologies, cultural transmission, and the spread of bow-and-arrow technology in the prehistoric Great Basin. American Antiquity 64: 231–242.
Bettinger, Robert L., and Peter J. Richerson. 1996. The state of evolutionary archaeology: Evolutionary correctness, or the search for the common ground. In H. D. G. Maschner, ed., Darwinian archaeologies, 221–231. New York: Plenum.
Boone, James L., and Eric A. Smith. 1998. Is it evolution yet? A critique of evolutionary archaeology. Current Anthropology 39: S141–S173.
Borgerhoff Mulder, Monique. 1991. Human behavioral ecology. In J. R. Krebs and N. B. Davies, eds., Behavioral ecology: An evolutionary approach, 69–98. 3rd ed. Oxford: Blackwell.
———. 2001. Using phylogenetically based comparative methods in anthropology: More questions than answers. Evolutionary Anthropology 10: 99–111.
Boyd, Robert, and Peter J. Richerson. 1985. Culture and the evolutionary process. Chicago: University of Chicago Press.
———. 1993. Culture and human evolution. In D. T. Rasmussen, ed., The origin and evolution of humans and humanness, 119–134. Boston: Jones & Bartlett.
Boyd, Robert, Monique Borgerhoff Mulder, William H. Durham, and Peter J. Richerson. 1997. Are cultural phylogenies possible? In P. Weingart, S. D. Mitchell, P. J. Richerson and S. Maasen, eds., Human by nature, 355–386. Mahwah, NJ: Erlbaum.
Bradley, Bruce A., and Dennis J. Stanford. 2004. The north Atlantic ice-edge corridor: A possible Palaeolithic route to the New World. World Archaeology 36: 459–478.
Brantingham, P. Jeffery. 2003. A neutral model of stone raw material procurement. American Antiquity 68: 487–509.
Braun, David P. 1987. Coevolution of sedentism, pottery technology, and horticulture in the central Midwest, 200 B.C.-A.D. 600, In W. F. Keegan, ed., Emergent Horticultural Economies of the Eastern Woodlands, 153–181. Carbondale: Southern Illinois University Press.
Bryant, David, Flavia Filimon, and Russel D. Gray. 2005. Untangling our past: Languages, trees, splits, and networks. In R. Mace, C. J. Holden, and S. J. Shennan, eds., The evolution of cultural diversity: A phylogenetic approach, 67–83. London: UCL Press.
Buchanan, Briggs, and Mark Collard. 2007. Investigating the peopling of North America through cladistic analyses of Early Paleoindian projectile points. Journal of Anthropological Archaeology 26: in press.
———. In press. Testing models of Early Paleoindian colonization and adaptation using cladistics. In M. J. O’Brien, ed., Transmission and archaeology: Some fundamental issues and case studies. Washington, DC: Society for American Archaeology Press.
Cavalli-Sforza, L. Luca, and Marcus W. Feldman. 1981. Cultural transmission and evolution: A quantitative approach. Princeton: Princeton University Press.
Cochrane, Ethan E. 2001. Style, function, and systematic empiricism: The conflation of process and pattern. In T. D. Hurt and G. F. M. Rakita,, eds., Style and function: Conceptual issues in evolutionary archaeology, 183–202. Westport, CT: Bergin & Garvey.
———. 2004. Explaining cultural diversity in ancient Fiji: The transmission of ceramic variability. Ph.D. diss., University of Hawai’i.
Collard, Mark, and Stephen J. Shennan. 2000. Ethnogenesis versus phylogenesis in prehistoric culture change: A case-study using European Neolithic pottery and biological phylogenetic techniques. In C. Renfrew and K. Boyle, eds., Archaeogenetics: DNA and the population prehistory of Europe, 89–97. Cambridge: McDonald Institute for Archaeological Research.
Collard, Mark, Stephen J. Shennan, and Jamshid J. Tehrani. 2005. Branching versus blending in macroscale cultural evolution: A comparative study. In C. P. Lipo, M. J. O’Brien, M. Collard, and S. J. Shennan, eds., Mapping our ancestors: Phylogenetic approaches in anthropology and prehistory, 53–63. New Brunswick, NJ: Transaction.
Cronk, Lee. 1991. Human behavioral ecology. Annual Reviews in Anthropology 20: 25–53.
Darwent, John, and Michael J. O’Brien 2005. Using cladistics to construct lineages of projectile points from Northeastern Missouri. In C. P. Lipo, M. J. O’Brien, M. Collard, and S. J. Shennan, eds., Mapping our ancestors: Phylogenetic approaches in anthropology and prehistory, 185–208. New Brunswick, NJ: Transaction.
Dewar, Robert E. 1995. Of nets and trees: Untangling the reticulate and dendritic in Madagascar’s prehistory. World Archaeology 26: 301–318.
Dopazo, Joaquin, Andreas Dress, and Arndt von Haesler. 1993. Split decomposition: A new technique to analyze viral evolution. Proceedings of the National Academy of Sciences USA 90: 10320–10324.
Dress, A. W. M., D. H. Huson, and V. Moulton. 1996. Analyzing and visualizing sequence and distance data using SplitsTree. Discrete Applied Mathematics 71: 95–109.
Dunnell, Robert C. 1978. Style and function: A fundamental dichotomy. American Antiquity 43: 192–202.
———. 1980. Evolutionary theory and archaeology. Advances in Archaeological Method and Theory 3: 35–98.
———. 1989. Aspects of the application of evolutionary theory in archaeology. In C. C. Lamberg-Karlovsky, ed., Archaeological thought in America, 35–49. Cambridge: Cambridge University Press.
———. 1995. What is it that actually evolves? In P. A. Teltser, ed., Evolutionary archaeology: Methodological issues, 33–50. Tucson: University of Arizona Press.
Dunnell, Robert C., and James K. Feathers. 1991. Late Woodland manifestations of the Malden plain, southeast Missouri. In M. S. Nassaney and C. S. Cobb, eds., Stability, transformation, and variation: The Late Woodland Southeast, 21–45. New York: Plenum.
Durham, William H. 1979. Toward a coevolutionary theory of human biology and culture. In N. Chagnon and W. Irons, eds., Evolutionary biology and human social behavior, 39–59. North Scituate, RI: Duxbury.
———. 1982. Interactions of genetic and cultural evolution: Models and examples. Human Ecology 10: 289–323.
———. 1990. Advances in evolutionary culture theory. Annual Review of Anthropology 19: 187–210.
———. 1991. Co-evolution: Genes, culture, and human diversity. Stanford, CA: Stanford University Press.
———. 1992. Applications of evolutionary culture theory. Annual Review of Anthropology 21: 331–355.
Eerkens, Jelmer W., Robert L. Bettinger, and R. McElreath. 2005. Cultural transmission, phylogenetics, and the archaeological record. In C. P. Lipo, M. J. O’Brien, M. Collard, and S. J. Shennan, eds., Mapping our ancestors: Phylogenetic approaches in anthropology and prehistory, 169–183. New Brunswick, NJ: Transaction.
Eerkens, Jelmer W., and Carl P. Lipo. 2005. Cultural transmission, copying errors, and the generation of variation in material culture and the archaeological record. Journal of Anthropological Archaeology 24: 316–334.
Eldredge, Niles, and Joel Cracraft. 1980. Phylogenetic patterns and the evolutionary process. New York: Columbia University Press.
Ewers, John C. 1955. The horse in Blackfoot culture. Bureau of American Ethnology Bulletin 159. Washington, DC: Bureau of American Ethnology.
Foley, Robert. 1987. Hominid species and stone-tool assemblages: How are they related? Antiquity 61: 380–392.
Foley, Robert, and Marta M. Lahr. 1997. Mode 3 technologies and the evolution of modern humans. Cambridge Archaeological Journal 7: 3–36.
———. 2003. On stony ground: Lithic technology, human evolution, and the emergence of culture. Evolutionary Anthropology 12: 109–122.
Forster, Peter, and Alfred Toth. 2003. Toward a phylogenetic chronology of ancient Gaulish, Celtic, and Indo-European. Proceedings of the National Academy of Sciences USA 100: 9079–9084.
Gillespie, John H. 1998. Population genetics: A concise guide. Baltimore: Johns Hopkins University Press.
Gimbutas, Marija. 1973. The beginning of the Bronze Age in Europe and the Indo-Europeans 3500–2500 BC. Journal of Indo-European Studies 1: 163–214.
Gray, M., 1992. Appendix: Application of stylistic codes for Pajarito black-on-white ceramics. In T. A. Kohler and M. J. Root, eds., Bandelier archaeological excavation project: Summer 1990 excavations at Burnt Mesa Pueblo and Casa del Rito, 175–180. Pullman: Washington State University.
Gray, Russell D., and Quentin D. Atkinson. 2003. Language-tree divergence times support the Anatolian theory of Indo-European origin. Nature 426: 435–439.
Gray, Russell D., and Fiona Jordan. 2000. Language trees support the express-train sequence of Austronesian expansion. Nature 405: 1052–1055.
Greenhill, Simon J., and Russell D. Gray. 2005. Testing population dispersal hypotheses: Pacific settlement, phylogenetic trees, and Austronesian languages. In R. Mace, C. J. Holden, and S. J. Shennan, eds., The evolution of cultural diversity: A phylogenetic approach, 31–52. London: UCL Press.
Guglielmino, C. R., C. Viganotti, B. Hewlett, and L. L. Cavalli-Sforza. 1995. Cultural variation in Africa: Role of mechanisms of transmission and adaptation. Proceedings of the National Academy of Sciences USA 92: 7585–7589.
Hahn, Matthew W., and R. Alexander Bentley. 2003. Drift as a mechanism for cultural change: An example from baby names. Proceedings of the Royal Society B 270: S120–S123.
Harmon, Marcel J., Todd L. VanPool, Robert D. Leonard, Christine S. VanPool, and Laura A. Salter. 2005. Reconstructing the flow of information across time and space: A phylogenetic analysis of ceramic traditions from prehistoric western and northern Mexico and the American Southwest. In C. P. Lipo, M. J. O’Brien, M. Collard, and S. J. Shennan, eds., Mapping our ancestors: Phylogenetic approaches in anthropology and prehistory, 209–229. New Brunswick, NJ: Transaction.
Hendy, Michael D., and David Penny. 1992. Spectral analysis of phylogenetic data. Journal of Classification 10: 5–24.
Hennig, Willi. 1950. Grundzüge einer Theorie der phylogenetischen Systematik. Berlin: Deutscher Zentralverlag.
———. 1965. Phylogenetic systematics. Annual Review of Entomology 10: 97–116.
———. 1966. Phylogenetic systematics. Urbana: University of Illinois Press.
Henrich, Joseph. 2001. Cultural transmission and the diffusion of innovations: Adoption dynamics indicate that biased cultural transmission is the predominate force in behavioral change and much of sociocultural evolution. American Anthropologist 103: 992–1013.
———. 2004. Demography and cultural evolution: How adaptive cultural processes can produce maladaptive losses: The Tasmanian case. American Antiquity 69: 197–214.
Herzog, Harold A., R. Alexander Bentley, and Matthew W. Hahn. 2004. Random drift and large shifts in popularity of dog breeds. Proceedings of the Royal Society B 271: S353–S356.
Holden, Clare J. 2002. Bantu languages trees reflect the spread of farming across sub-Saharan Africa: A maximum-parsimony analysis. Proceedings of the Royal Society B 269: 793–799.
———. 2005. The spread of Bantu languages, farming, and pastoralism in sub-Equatorial Africa. In C. P. Lipo, M. J. O’Brien, M. Collard, and S. J. Shennan, eds., Mapping our ancestors: Phylogenetic approaches in anthropology and prehistory, 249–267. New Brunswick, NJ: Transaction.
Holden, Clare J., Andrew Meade, and Mark Pagel. 2005. Comparison of maximum parsimony and Bayesian Bantu language trees. In R. Mace, C. J. Holden, and S. J. Shennan, eds., The evolution of cultural diversity: A phylogenetic approach, 53–65. London: UCL Press.
Hubbell, Stephen P. 2001. The unified neutral theory of biodiversity and biogeography. Princeton: Princeton University Press.
Huson, D. H. 1998. SplitsTree: Analyzing and visualizing evolutionary data. Bioinformatics 14: 68–73.
Jordan, Peter, and Thomas Mace. 2005. Tracking culture-historical lineages: Can “descent with modification” be linked to “association by descent”? In C. P. Lipo, M. J. O’Brien, M. Collard, and S. J. Shennan, eds., Mapping our ancestors: Phylogenetic approaches in anthropology and prehistory, 149–167. New Brunswick, NJ: Transaction.
Jordan, Peter, and Stephen J. Shennan. 2003. Cultural transmission, language, and basketry traditions amongst the Californian Indians. Journal of Anthropological Archaeology 22: 42–74.
———. 2005. Cultural transmission in indigenous California. In R. Mace, C. J. Holden, and S. J. Shennan, eds., The evolution of cultural diversity: A phylogenetic approach, 165–196. London: UCL Press.
Kimura, Moto. 1983. The neutral allele theory of molecular evolution. Cambridge: Cambridge University Press.
Kimura, Moto, and James F. Crow. 1964. The number of alleles that can be maintained in a finite population. Genetics 49: 725–738.
Kirch, Patrick V., and Roger C. Green. 1987. History, phylogeny, and evolution in Polynesia. Current Anthropology 28:431–456.
Kishino, Hirohisa, and Masami Hasegawa. 1989. Evaluation of the maximum likelihood estimate of the evolutionary tree topologies from DNA sequence data, and the branching order in Hominoidea. Journal of Molecular Evolution 29: 170–179.
Kitching, Ian J., Peter L. Forey, Christopher J. Humphries, and David M. Williams. 1998. Cladistics: The theory and practice of parsimony analysis. Oxford: Oxford University Press.
Kohler, Timothy A., Stephanie Van Buskirk, and Samantha Ruscavage-Barz. 2004. Vessels and villages: Evidence for conformist transmission in early village aggregations on the Pajarito Plateau, New Mexico. Journal of Anthropological Archaeology 23: 100–118.
Krebs, John R., and Nicholas B. Davies. 1993. An introduction to behavioural ecology. Oxford: Blackwell.
Leonard, Robert D. 2001. Evolutionary archaeology. In I. Hodder, ed., Archaeological Theory Today, 65–97. Cambridge: Polity.
Leonard, Robert D., and George T. Jones. 1987. Elements of an inclusive evolutionary model for archaeology. Journal of Anthropological Archaeology 6: 199–219.
Lipo, Carl P. 2001. Science, style, and the study of community structure: An example from the central Mississippi River valley. Oxford: BAR International Series.
Lipo, Carl P., M. E. Madsen, Robert C. Dunnell, and T. L. Hunt. 1997. Population structure, cultural transmission, and frequency seriation. Journal of Anthropological Archaeology 16: 301–334.
Lyman, R. Lee, and Michael J. O’Brien. 1998. The goals of evolutionary archaeology: History and explanation. Current Anthropology 39: 615–652.
Lynch, A., and A. J. Baker. 1994. A population mimetics approach to cultural evolution in chaffinch song: Differentiation among populations. Evolution 48: 351–359.
Meltzer, David J. 1981. A study of style and function in a class of tools. Journal of Field Archaeology 8: 313–326.
Moore, Carmelia C., and Romney A. Kimball. 1994. Material culture, geographic propinquity, and linguistic affiliation on the north coast of New Guinea: A reanalysis of Welsch, Terrell, and Nadolski (1992). American Anthropologist 96: 370–396.
———. 1996. Will the “real” data please stand up? Reply to Welsch (1996). Journal of Quantitative Anthropology 6: 235–261.
Moore, John H. 1994. Putting anthropology back together again: The ethnogenetic critique of cladistic theory. American Anthropologist 96: 370–396.
———. 2001. Ethnogenetic patterns in native North America. In J. E. Terrell, ed., Archaeology, language, and history: Essays on culture and ethnicity, 30–56. Wesport, CT: Bergin & Garvey.
Neff, Hector. 1992. Ceramics and evolution. Archaeological Method and Theory 4: 141–193.
Neiman, Fraser D. 1995. Stylistic variation in evolutionary perspective: Inferences from decorative diversity and interassemblage distance in Illinois Woodland ceramic assemblages. American Antiquity 60: 7–36.
Nelson, Gareth. 1978. Ontogeny, phylogeny, paleontology, and the biogenetic law. Systematic Zoology 27:324–345.
Nelson, Gareth, and Nelson Platnick. 1981. Systematics and biogeography: Cladistics and vicariance. New York: Columbia University Press.
O’Brien, Michael J., John Darwent, and R. Lee Lyman. 2001. Cladistics is useful for reconstructing archaeological phylogenies: Palaeoindian points from the southeastern United States. Journal of Archaeological Science 28: 1115–1136.
O’Brien, Michael J., and T. D. Holland. 1995. The nature and premise of a selection-based archaeology. In P. A. Teltser, ed., Evolutionary archaeology: Methodological issues, 175–200. Tucson: University of Arizona Press.
O’Brien, Michael J., and R. Lee Lyman. 2000. Applying evolutionary archaeology: A systematic approach. New York: Kluwer.
O’Brien, Michael J., and R. Lee Lyman, eds. 2003a. Style, function, transmission: Evolutionary archaeological perspectives. Salt Lake City: University of Utah Press.
———. 2003b. Cladistics and archaeology. Salt Lake City: University of Utah Press.
O’Brien, Michael J., R. Lee Lyman, Youssef Saab, Elias Saab, John Darwent, and Daniel S. Glover. 2002. Two issues in archaeological phylogenetics: Taxon construction and outgroup selection. Journal of Theoretical Biology 215: 133–150.
Peck, J. R., G. Barreau, and S. C. Heath. 1997. Imperfect genes, Fisherian mutation, and the evolution of sex. Genetics 145: 1171–1199.
Phillips, Philip, James A. Ford, and James B. Griffin. 1951. Archaeological survey in the lower Mississippi alluvial valley, 1940–1947. Papers of the Peabody Museum of American Archaeology and Ethnology 25. Cambridge: Harvard University Press.
Pigliucci, Massimo. 1996. How organisms respond to environmental changes: From phenotypes to molecules (and vice versa). Trends in Ecology and Evolution 11: 168–173.
Pulliam, H. Ronald, and Christopher Dunford. 1980. Programmed to learn: An essay on the evolution of culture. New York: Columbia University Press.
Quicke, Donald J. 1993. Principles and techniques of contemporary taxonomy. Glasgow: Blackie.
Ramenofsky, Ann F. 1995. Evolutionary theory and native artifact change in the post contact period. In P. A. Teltser, ed., Evolutionary archaeology: Methodological issues, 129–147. Tucson: University of Arizona Press.
Renfrew, Colin. 1987. Archaeology and language: The puzzle of Indo-European origins. London: Cape.
Rexová, Katerina, Daniel Frynta, and Jan Zrzavy. 2003. Cladistic analysis of languages: Indo-European classification based on lexicostatistical data. Cladistics 19: 120–127.
Richerson, Peter J., and Robert Boyd. 1992. Cultural inheritance and evolutionary ecology. In E. A. Smith and B. Winterhalder, eds., Evolutionary ecology and human behavior, 61–92. New York: Aldine de Gruyter.
Roberts, J. M., C. C. Moore, and A. K. Romney. 1995. Predicting similarity in material culture among New Guinea villages from propinquity and language: A log-linear approach. Current Anthropology 36: 769–788.
Robson-Brown, Kate A. 1996. Systematics and integrated methods for the modelling of the pre-modern human mind. In P. Mellars and K. Gibson, eds., Modelling the early human mind, 103–117. Cambridge: McDonald Institute for Archaeological Research.
Runciman, W. G. 2002. Heritable variation and competitive selection as the mechanism of sociocultural evolution. Proceedings of the British Academy 112: 9–25.
Schuh, Randall T. 2000. Biological systematics: Principles and applications. Ithaca, NY: Cornell University Press.
Shennan, Stephen J. 1991. Tradition, rationality, and cultural transmission. In R. W. Preucel, ed., Processual and post-processual archaeologies: Multiple ways of knowing the past, 197–208. Carbondale, IL: Center for Archaeological Investigations.
———. 2000. Population, culture history, and the dynamics of culture change. Current Anthropology 41: 811–835.
———. 2001. Demography and cultural innovation: A model and some implications for the emergence of modern human culture. Cambridge Archaeological Journal 11: 5–16.
———. 2002. Genes, memes, and human history: Darwinian archaeology and cultural evolution. London: Thames & Hudson.
Shennan, Stephen J., and Mark Collard. 2005. Investigating processes of cultural evolution on the north coast of New Guinea with multivariate and cladistic analyses. In R. Mace, C. J. Holden, and S. J. Shennan, eds., The evolution of cultural diversity: A phylogenetic approach, 133–164. London: UCL Press.
Shennan, Stephen J., and J. Richard Wilkinson. 2001. Ceramic change and neutral evolution: A case study from Neolithic Europe. American Antiquity 66: 577–593.
Simkin, Mikhail V., and Vwani P. Roychowdhury. 2003. Read before you cite! Complex Systems 14: 269.
Slater, P. J. B., and S. A. Ince. 1979. Cultural evolution in chaffinch song. Behaviour 71: 146–166.
Smith, Andrew B. 1994. Systematics and the fossil record: Documenting evolutionary patterns. Oxford: Blackwell.
Smith, Eric A. 2000. Three styles in the evolutionary analysis of human behavior. In L. Cronk, N. Chagnon, and W. Irons, eds., Adaptation and human behaviour, 27–46. New York: Aldine de Gruyter.
Smith, Eric A., Monique Borgerhoff Mulder, and Kim Hill. 2001. Controversies in the evolutionary social sciences: A guide to the perplexed. Trends in Ecology and Evolution 16: 128–135.
Smith, Eric A., and Bruce Winterhalder (eds.). 1992. Evolutionary ecology and human behavior. New York: Aldine de Gruyter.
Stanford, Dennis, and Bruce A. Bradley. 2002. Ocean trails and prairie paths? Thoughts about Clovis origins. In N. G. Jablonski, ed., The first Americans: The Pleistocene colonization of the New World, 255–271. Memoirs of the California Academy of Sciences 27. San Francisco.
Tehrani, Jamshid J., and Mark Collard. 2002. Investigating cultural evolution through biological phylogenetic analyses of Turkmen textiles. Journal of Anthropological Archaeology 21: 443–463.
Terrell, John E. 1988. History as a family tree, history as a tangled bank. Antiquity 62: 642–657.
———. 2001. Introduction. In J. E. Terrell, ed., Archaeology, language, and history: Essays on culture and ethnicity, 1–10. Westport, CT: Bergin & Garvey.
Terrell, John E., Terry L. Hunt, and Chris Gosden. 1997. The dimensions of social life in the Pacific: Human diversity and the myth of the primitive isolate. Current Anthropology 38: 155–195.
Terrell, John E., Kevin M. Kelly, and Paul Rainbird. 2001. Foregone conclusions? In search of “Papuans” and “Austronesians.” Current Anthropology 42: 97–124.
Tschauner, Harmut. 1994. Archaeological systematics and cultural evolution: Retrieving the honour of culture history. Man 29: 77–93.
Welsch, Robert L. 1996. Language, culture, and data on the north coast of New Guinea. Journal of Quantitative Anthropology 196: 209–234.
Welsch, Robert L., John E. Terrell, and John A. Nadolski. 1992. Language and culture on the north coast of New Guinea. American Anthropologist 94: 568–600.
Wiley, E. O., D. Siegel-Causey, D. R. Brooks, and V. A. Funk. 1991. The compleat cladist: A primer of phylogenetic procedures. Lawrence: University of Kansas Museum of Natural History.
Wilhelmsen, Kris H. 2001. Building the framework for an evolutionary explanation of projectile point variation: An example from the central Mississippi River valley. In T. L. Hunt, C. P. Lipo, and S. L. Sterling, eds., Posing questions for a scientific archaeology, 97–144. Westport: Bergin & Garvey.
Winterhalder, Bruce. 1981. Foraging strategies in the boreal environment: An analysis of Cree hunting and gathering. In B. Winterhalder and E. A. Smith, eds., Hunter-gatherer foraging strategies, 66–98. Chicago: University of Chicago Press.
Winterhalder, Bruce, and Eric A. Smith. 2000. Analyzing adaptive strategies: Human behavioral ecology at twenty-five. Evolutionary Anthropology 9: 51–72.