Figure 5.1: The anatomy of smell.
The nose, for example, of which no philosopher has ever spoken with veneration and gratitude—the nose is, albeit provisionally, the most delicate instrument at our disposal. It is an instrument capable of recording the most minimal changes that escape even spectroscopic detection.
FRIEDRICH NIETZSCHE, Twilight of the Idols, 1889
They appear to be bringing us a certain message from a material reality.
GASTON BACHELARD, La Formation de l’esprit scientifique, contribution à une psychoanalyse de la connaissance objective, 1938
TUCKED WITHIN the arms of a deadly sea anemone that is fastened to a coral reef off the coast of Isla Verde in the Philippines is a pair of small orange-and-white striped fish known as common clownfish or, more properly, anemonefish, or, even more properly, Amphiprion ocellaris. One of the pair, a female, has led a more interesting life than most vertebrates, because she has not always been female. Like all anemonefish, she started life as a smaller male who had been subordinate to the one female in the group of fish inhabiting this particular anemone. Anemonefish have a rigid social structure, and as a male this one had competed with the other males until eventually becoming dominant and enjoying the honor of mating with the sole female. But when its mate was eaten by a passing eel, the ovaries that had lain dormant in its body for several years matured, its testes ceased to function and the male clownfish became the queen female ready to mate with the next male in the pecking order.
Anemonefish are common inhabitants of coral reefs from the Indian Ocean to the western Pacific, feeding on plants, algae, plankton and animals such as mollusks and small crustaceans. Their small size, bright colors and absence of spines, sharp fins, barbs or spikes make them easy prey for the eels, sharks and other predators that rove the reef. When threatened, their principal means of defense is to dash between the tentacles of their host anemone, from whose poisonous sting they are protected by a thick layer of mucus covering their scales. In turn, the anemone benefits from its colorful tenants who chase off unwelcome intruders, such as grazing butterflyfish.
It was in this setting that the anemonefish came to be most familiar to us in the animated film Finding Nemo.*1 The challenge facing Nemo’s dad, Marlin, was to find his son, who had been abducted from his home in the Great Barrier Reef and carried all the way to Sydney. But the challenge that besets real anemonefish is to find their way back home.
Each anemone may be host to an entire colony of anemonefish that contains a dominant male and female together with several juvenile males vying with one another for the role of queen’s consort. The unusual capability of the dominant male to change sex on the death of the queen fish, a capability known as protandrous hermaphroditism, may be an adaptation to life in the dangerous reef, as it allows the colony to survive the demise of the single reproductive female without ever having to leave the protection of the host anemone. But although an entire colony of fish may remain resident on a single anemone for many years, the progeny of those fish must leave the safety of their home. And, eventually, they will need to find their way back.
A full moon is the cue for spawning of most coral fish.*2 As the moon begins to wane over the ocean, the female of the pair busies herself laying a clutch of eggs to be fertilized by the dominant male. Thereafter, her work is done; guarding the eggs and chasing away carnivorous reef fish is the job of the male anemonefish. After about a week of his custodianship, the eggs hatch and hundreds of larvae are launched into the currents.
Larval anemonefish are only a few millimeters in length and almost completely transparent. For about a week they drift in the pelagic currents, feeding on zooplankton. As anyone who has dived off coral reefs will know, drifting in an ocean current will soon take you far from your starting point; so anemonefish larvae can be carried many kilometers from their natal reef. Most are eaten, but some survive; after about a week, these lucky few swim to the sea floor and, within a day, metamorphose (like our frog in chapter 3) into their juvenile form, a smaller version of the adult fish. Lacking the protection of the poisonous anemone, the brightly colored juvenile is very vulnerable to predators that cruise the benthic waters. If it is to survive, it must quickly find a coral reef where it can gain sanctuary.
It was thought that larval reef fish drifted with the ocean currents and that they relied on mere chance to be washed up close to a suitable reef. But that explanation didn’t really make complete sense since it was known that most larvae are strong swimmers and there is no point in swimming if you don’t know where to go. Then in 2006 Gabriele Gerlach, a researcher from the famous Marine Biological Laboratory in Woods Hole, Massachusetts, carried out genetic fingerprinting of fish living on reefs separated by between three and twenty-three kilometers within the complex that forms the Great Barrier Reef of Australia. She discovered that fish inhabiting the same reef were much more closely related to one another than they were to those inhabiting more distant reefs. Since all juvenile reef fish larvae disperse across large distances, the finding only makes sense if most adults return to the reef on which they were born. Somehow, each larval reef fish must be imprinted with a signature that identifies its spawning area.
But how do larval or juvenile anemonefish that have drifted so far from their home know which direction to swim in? The sea floor doesn’t provide any useful visual cues. It lacks reference points and so looks the same in all directions: a sandy desert decorated with scattered pebbles, boulders and the occasional wandering arthropod. The distant coral reef is unlikely to provide any auditory signals that could travel several kilometers. The currents themselves are an additional problem, as the direction of flow varies with depth and it can be very difficult to determine whether the body of water is moving or stationary. There is no evidence that anemonefish possess the kind of magnetic compass that helped to guide our robin on her winter migration. So how do they find their way?
Fish do possess a keen sense of smell. Sharks, two-thirds of whose brains is devoted to olfaction, can famously smell a drop of blood from more than a kilometer away. Perhaps reef fish smell their way home? To test this theory, in 2007 Gabriele Gerlach designed a “two-channel olfactory choice flumes test” in which larval reef fish were placed downstream of two flumes of seawater: one collected from the reef on which they hatched and the other from a distant reef. She then measured which water flume the larvae preferred: home or away.
Invariably, the larval fish swam toward the flume filled with water from the reef on which they had been spawned. They could clearly discriminate between foreign and native reef waters, presumably by their different smells. Michael Arvedlund, a researcher from James Cook University in Queensland, Australia, used a similar experimental setup to demonstrate that anemonefish could smell their host anemone species and distinguish it from others that they do not colonize. Even more remarkably, Daniella Dixson, also from James Cook University, found that anemonefish can distinguish between water collected from their preferred habitat of reefs that lie beneath vegetated islands and the less favored offshore reef water. It really seems that Nemo and other reef fish sniff their way home.
The ability of animals to navigate with their sense of smell is legendary. Every year, along ocean coasts around the world, millions of salmon assemble into large schools at the mouths of rivers before venturing inland to battle against the flow of current, rapids, waterfalls and sandbanks to reach their spawning grounds. As with the anemonefish, it was thought that the salmon’s selection of a suitable river was pretty much down to chance. But then in 1939 the Canadian Wilbert A. Clemens tagged 469,326 young salmon caught in a particular tributary of the Fraser River system. Years later, he caught 10,958 of those he had tagged that had returned to that same tributary. Not a single marked salmon was caught in any other tributary of the river. None had lost its way on its journey from the ocean to its home stream. How they manage to navigate through ocean and stream remained a mystery for many years. Then Professor Arthur Hasler from the University of Wisconsin-Madison suggested that the young salmon follow a scent trail, and tested his theory in 1954 by catching several hundred returning fish upstream of a fork (the confluence of two streams) in the Issaquah River near Seattle and then transporting them downstream to below the fork. The salmon invariably returned to the same branch of the fork in which they had been captured. But when he blocked their nostrils with cotton-wool stoppers before release, they swam up to the fork in the river tacking this way and that and could not decide whether to go right or left.
The olfactory sense is perhaps even more remarkable on land because the volume of the atmosphere, in which odorants are diluted, is even vaster than that of the ocean. The atmosphere is also subject to a greater degree of turbulence, owing to the weather, so odorant molecules are dispersed more quickly in air than in water. Yet the sense of smell is vital to the survival of most land animals, used not only to find the way home but also to catch prey, escape from predators, find a mate, provide alarm signals, mark territory, trigger physiological changes and communicate. This whole smellscape of olfaction is much less obvious to humans, who often harness the keener olfactory senses of their companion animals to detect these signals and signs. Dogs are, of course, famously interested in smell and the bloodhound, whose olfactory epithelium (more on this later) is forty times the size of ours, is rightly famous for its ability to follow the scent trail of a single individual. We have all seen those movies featuring a keen tracker dog that needs only a quick sniff of the discarded shirt of an escaped convict to be able to track the villain across moorland, forest and stream. And though the stories may be fiction, the ability of the hound is entirely real. Dogs can tell from a track which way the person or animal was traveling and can follow a scent trail that is several days old.
The startling power of animals’ sense of smell can be appreciated if we reflect upon the feats that a bloodhound or an anemonefish routinely accomplishes. Consider first the bloodhound; its sense of smell is tuned to detect tiny quantities of organic chemicals, such as butyric acid, that are shed by humans and other animals; and the sensitivity of its nose is extraordinary. If just a single gram of butyric acid were allowed to evaporate in a room, then we humans would be just about capable of detecting its sweet, rancid odor. But a dog is able to detect that same gram of chemical if its vapors were diluted to fill the air above an entire city to a height of 100 meters. And consider again those anemonefish or salmon that detect the scent of their distant home, kilometers away, diluted in the vastness of the ocean.
But the animal sense of smell isn’t remarkable only for its sensitivity. There is also its highly developed discriminatory power. Dogs are routinely used by customs officers to detect a wide range of odorants, from drugs such as marijuana and cocaine, to chemicals in explosives such as C-4—often through dense packaging and a suitcase. They can also distinguish between the scents of individuals, even identical twins. So how do they do this? Surely the butyric acid shed by one of us is the same butyric acid that is released by everyone? Of course it is; but, alongside the butyric acid, each of us sheds a delicate and complex cocktail of hundreds of organic molecules that provides a signature of our presence that is as individual as our fingerprints. Dogs can “see” our olfactory fingerprint as easily as we can see the color of a person’s shirt. Anemonefish or salmon must similarly recognize the scent of their home, just as we might recognize our street or spot the color of our front door.
But dogs, salmon or anemonefish aren’t the supreme athletes of olfaction. A bear’s sense of smell is over seven times as sensitive as even a bloodhound’s; and it can smell a carcass 20 kilometers away. A moth can detect a mate at a distance of some 10 kilometers; rats smell in stereo and snakes smell with their tongues. All these olfactory skills are essential for animals that must seek out food, find mates and/or avoid predators; they have evolved a sensitivity to volatile cues that betray the proximity of these resources or dangers, whether in air or water. The sense of smell is so important to animal survival that behavioral responses to odors appear to be hard-wired in a number of species. Experiments with Orkney Island voles demonstrated that they avoided traps baited with the secretions of predatory stoats, even though stoats have been absent from the island for five thousand years!
Humans are said to have a much poorer sense of smell than our relatives. When, several million years ago, Homo erectus lifted his upper body off the forest floor to walk upright, he also raised his nose from the ground and its rich source of aromas. Thereafter, sight and sound, both more efficient from a higher vantage point, became his principal sources of information. So the human snout became shorter, the nostrils narrowed and mutations accumulated in most of the thousand or so ancestral mammalian genes that encode olfactory receptors (more on these later). We also, perhaps sadly, lost an auxiliary olfactory sense found in other animals and conferred by the vomeronasal organ (VNO) or Jacobson’s organ, whose role is to detect sex pheromones.
Yet, despite our diminished genetic repertoire of only about three hundred olfactory receptor genes and our altered anatomy, we have retained a surprisingly good sense of smell. We may not be able to sniff out a mate or our dinner from several miles away, but we can discriminate between around ten thousand different scents and, as Nietzsche noted, can outperform “even spectroscopic detection” of odorous chemicals. Our ability to appreciate scents has inspired some of our greatest poetry (“A rose by any other name would smell as sweet”) and plays a crucial role in our sense of well-being and contentment.
Our sense of smell has also played a surprisingly active role in human history. The earliest texts record a reverence for pleasant aromas and an abhorrence of foul smells. Places of worship and meditation were frequently scented with perfumes and spices. In the Hebrew Bible, God instructs Moses to build a place of worship and tells him: “Take to you sweet spices, stacte, and onycha, and galbanum; these sweet spices with pure frankincense: of each shall there be a like weight. And you shall make it a perfume, a confection after the art of the apothecary, tempered together, pure and holy.”1 The ancient Egyptians even had a god of perfume, Nefertum, who was also a god of healing, a kind of mythical aromatherapist.
The association of health with pleasant aromas, and, conversely, disease and decay with foul smells, led many to believe that the causal direction led from odor to health or disease, rather than the reverse. For example, the great Roman physician Galen taught that malodorous sheets, mattresses and blankets could accelerate the pollution of the body fluids. Nauseating exudations (miasmas) coming from sewers, charnel houses, cesspools and marshes were considered to be the sources of many fatal diseases. Conversely, pleasant smells were thought to ward off illness so that, in medieval Europe, physicians would insist that, before they entered a plague victim’s house, it had to be thoroughly aired and perfumed by lighting fragrant fires scented with incense, myrrh, roses, cloves and other aromatic herbs. Indeed, the profession of perfumery was originally dedicated to the disinfection of houses, rather than personal grooming.
The importance of the sense of smell is not of course limited to detecting odorants breathed in through our nostrils. Remarkably, our sense of taste is generally considered to be about 90 percent smell. When we taste food, the taste receptors on our tongue and palate detect chemicals dissolved in saliva; but the receptors come in only five varieties, able to identify combinations of only five basic tastes—sweet, sour, salty, bitter and umami (a Japanese word that means “pleasant savory taste”). But volatile odorants evaporating from our food and drink gain entry to the nasal cavity from the back of our throat to activate combinations of hundreds of different smell receptors. These provide us with a far greater ability, compared with taste, to distinguish between thousands of different aromas and to enjoy the rich flavors (mostly scents) of fine wine, aromatic food, spices, herbs or coffee. And even though we have lost the vomeronasal sense enjoyed by most of our fellow mammals, the huge perfume industry is evidence of the role that scent continues to play in human courtship and sex. Freud even saw a connection between sexual repression and the sublimation of the sense of smell in most of us, but nevertheless claimed that “there exist, even in Europe,*3 peoples who are highly appreciative of the strong odor of the genitalia.”2
So how do humans, dogs, bears, snakes, moths, sharks, rats or anemonefish detect these messages “from a material reality”? How do we distinguish between such a wide variety of odorants?
Unlike our senses of sight and hearing, which capture information indirectly via electromagnetic waves or sound waves carried to us from an object, both taste and smell receive information directly from contact with the object detected (a molecule), bringing messages “from a material reality.” Both appear to work through rather similar principles. The molecules they detect are either dissolved in saliva or float through the air and are then picked up by receptors either on the tongue (taste) or in the olfactory epithelium in the roof of our nasal cavity (smell). This requirement for volatility means that most odorants are fairly small molecules.
The nose itself plays no direct role in smelling, other than channeling air toward the olfactory epithelium, which is at the back of the nose (figure 5.1). This tissue is quite small, measuring only 3 square centimeters (about the size of a postage stamp) in humans, but it is lined with both mucus-secreting glands and millions of olfactory neurons. These are a type of nerve cell that are to the sense of smell what retinal rods and cones in the eye are to the sense of sight. The front end of the olfactory neuron is shaped a bit like a broom, with a many-pronged head where the cell membrane is folded into lots of hair-like cilia. This broom with its brush of cilia pokes out of the cell layer where it can capture passing odor molecules. The back end of the cell is like the broom’s handle, forming the cell’s axon or nerve that extends through a small bone at the back of the nasal cavity to enter the brain, where it connects with a region called the olfactory bulb.
The remainder of this chapter may be best read with an orange in front of you, perhaps chopped into segments so that the tangy aromas are released and travel through your nose to reach the nasal epithelium. You might even slip one of the segments into your mouth to allow its volatile odorants to find their way through the retronasal route to that same tissue. Like all natural scents, the smell of an orange is very complex and made up of hundreds of volatile compounds, but one of the most fragrant is called limonene,*4 whose course we will now follow from molecule to fragrance.
Limonene, as its name implies, is abundant in citrus fruits such as oranges and lemons and is largely responsible for their tangy aroma and flavor. The chemical belongs to a class of compounds known as terpenes, which are the smelly constituents of the essential oils of many plants and flowers, generating the rich aromas of pine, roses, grapes and hops. So feel free to exchange the orange for a glass of beer or wine if you prefer. This chemical is produced in many parts of the citrus plant, including its leaves, but is most abundant in the skin of its fruit, which can be squeezed to yield nearly pure limonene.
Limonene is a volatile liquid that will gradually evaporate at room temperature, so your orange will be releasing millions of limonene molecules into the surrounding air. Most of these will float off into the room and out through the door and windows, but a few will be carried close to your nose by the air currents. Your next inhalation will suck in several liters of this odorant-laden air that will pass through your nostrils and across the nasal epithelium, which is lined with approximately ten million olfactory neurons.
When limonene molecules waft past the olfactory epithelium brushes, some of them are ensnared by the olfactory neurons. The capturing of a single limonene molecule is sufficient to trigger the opening of a tiny channel in the neuron’s cell membrane that allows a flow of positively charged calcium ions into the cell from outside. When about thirty-five limonene molecules have been captured, the subsequent flow of ions into the cell amounts to a tiny electric current of about one picoamp*5 in total. This level of current acts like a switch to fire an electrical signal, called an action potential (we will learn much more about these in chapter 8) down the handle of the brush-like cell, its axon. This signal travels all the way to the olfactory bulb in your brain. After further neural processing you experience this “message from a material reality” as the tangy aroma of oranges.
The key event in this whole process is of course the capturing of the odor molecule by the olfactory neuron. So how does it work? By analogy with sight and the eye’s light-sensitive cone and rod cells (also types of neurons), it was expected that the sense of smell would similarly be conferred by some kind of surface-located olfactory receptors. But, in the 1970s, the nature and identity of olfactory receptors were completely unknown.
Richard Axel was born in Brooklyn, New York, in 1948, the first child of immigrant parents who had fled Poland before the Nazi invasion. His childhood was typical for the neighborhood: running errands for his father, a tailor, between playing stickball (a kind of street baseball, with manhole covers as the bases and a broom handle as bat) or basketball in the local roads and courtyards. His first job, aged eleven, was as a messenger, delivering false teeth to dentists; at twelve he was laying carpets, and at thirteen serving corned beef and pastrami in a local delicatessen. The chef was a Russian who used to recite Shakespeare while slicing cabbage heads, providing the young Richard with his first real exposure to the cultural world beyond delis and basketball courts and inspiring a deep and abiding love of great literature. Axel’s intellectual talents were spotted by a local high-school teacher who encouraged him to apply, successfully, for a scholarship to Columbia University in New York to read literature.
As a freshman, Axel threw himself into the intellectual maelstrom of university life in the 1960s. But to support his party-going lifestyle he took a job washing glassware in a molecular genetics laboratory. He became fascinated by this emerging science, but remained hopeless at glass washing, so was sacked from that job and rehired as a research assistant. Torn between literature and science, he eventually decided to enroll in a graduate genetics course but then switched to studying medicine to escape the Vietnam draft. He was apparently as bad at medicine as he’d been at glass washing. He couldn’t hear a heart murmur and never saw the retina; his glasses once fell into an abdominal incision and he even managed to sew a surgeon’s finger to his patient. He was eventually allowed to graduate only on condition that he promise never to practice medicine on living patients. He returned to Columbia to study pathology, but after a year the chairman of the department insisted that he should never practice on dead patients either.
Realizing that medicine clearly lay beyond his talents, Axel eventually managed to return to research at Columbia University. Thereafter he made rapid progress and even invented a novel technique for getting foreign DNA inside mammalian cells that became a mainstay of the genetic engineering/biotech revolution of the late twentieth century and earned Columbia University hundreds of millions of dollars of revenue in licensing agreements: a generous return on their scholarship investment.
By the 1980s Axel was wondering whether molecular biology could help to solve that mystery of mysteries: how the human brain works. He switched from studying the behavior of genes to studying the genes for behavior, with the long-term aim of “dissecting how higher brain centers generate a ‘percept,’ say, of the scent of a lilac, or coffee, or a skunk.…”3 His first foray into neuroscience was investigating egg-laying behavior in a marine snail. It was at about this time that a very talented researcher, Linda Buck, joined his lab. She had trained as an immunologist at the University of Dallas before becoming fascinated by the emerging field of molecular neuroscience and moving to Axel’s laboratory to be at the forefront of this research. Together, Axel and Buck devised an ingenious series of experiments to probe the molecular basis of smell. The first question they addressed was the identity of the receptor molecules that were presumed to exist on the surface of olfactory neurons and to capture and identify different odorant molecules. Extrapolating from what was known about other sensory cells, they guessed that the receptors were some kind of proteins poking out of the cell membrane where they could bind passing odor molecules; but, at the time, nobody had ever isolated any of these odor receptors, so no one had a clue what they looked like or how they worked. All the team had to go on was an inkling that the elusive receptors might belong to a family of proteins called G-protein-coupled receptors that were known to be involved in detecting other kinds of chemical signals, such as hormones.
Linda Buck managed to identify an entirely new family of genes encoding this kind of receptor that were only expressed*6 in olfactory receptor neurons. She went on to demonstrate that these genes did indeed encode the elusive odor-capturing receptors. Further analysis showed that the rat’s genome encoded about a thousand of these newly identified receptors, each a little different from the others, and each presumably tuned to detect a single odorant. Humans have a similar number of olfactory receptor genes, but two-thirds of them have degenerated into what are called pseudogenes, which are a kind of gene fossil that have accumulated so many mutations that they no longer work.
But whether there are three hundred or a thousand receptor genes, this is very far from the figure of ten thousand different scents that humans can identify. There clearly wasn’t a one-to-one mapping between types of odor receptors and types of odors; so how the signals received by olfactory receptors are transformed into smells remained a mystery. It also wasn’t clear how the job of detecting all the variety of odor molecules was shared out between different cells. The genome of each cell has the complete set of olfactory receptor genes, so could potentially detect every odor. Or is there some kind of division of labor? To answer these questions the Columbia University team devised an even more ingenious experiment. They genetically altered mice so that all olfactory neurons expressing one particular odor receptor were dyed blue. If all the cells stained blue, that would indicate that they all expressed this receptor. The answer was clear when the team examined the olfactory cells of the engineered mice: approximately one in every thousand cells was dyed blue. It seemed that each olfactory neuron was not a generalist but a specialist.
It wasn’t long before Linda Buck moved from Columbia to set up her own laboratory at Harvard, and the two groups continued working in parallel to dissect many of the remaining secrets of olfaction. They soon devised techniques to isolate individual olfactory neurons and directly probe their sensitivity to particular odorants, such as the limonene of your orange. They discovered that each odorant chemical activated not just one but several neurons; also, that single neurons responded to several different odorants. These findings appeared to solve the conundrum of how only three hundred olfactory receptors can identify ten thousand different smells. Just as only twenty-six letters can be combined in lots of different ways to write every word in this book, so a few hundred olfactory receptors can be activated in trillions of different combinations to provide the vast array of scents.
Richard Axel and Linda Buck were awarded the Nobel Prize in 2004 for their pioneering discoveries of “odorant receptors and the organization of the olfactory system.”
The initiating event in the detection of an odor, such as that of an orange, a coral reef, a mate, a predator or prey is now understood to be the binding of a single molecule of odorant to a single olfactory receptor on the surface of the brush end of one of those broom-like olfactory neurons. But how does each receptor recognize its own set of odorant molecules, such as limonene, and not capture and bind to any one of the chemical ocean of other possible odorants that might float past the olfactory epithelium?
This is the central mystery of smell.
The conventional explanation is based on what is known as the lock-and-key mechanism. The odor molecules are thought to fit into the olfactory receptors like a key in a keyhole. For example, the limonene molecule was thought to slip snugly into a specialist olfactory receptor. Somehow, in a process that remains unclear, this binding event was thought to turn the lock of the receptor and trigger the release of a protein, called a G protein, that is normally tethered to the inner surface of the receptor, rather like a torpedo tethered to the hull of a ship. Once the torpedo protein is fired into the cell it makes its way to the cell membrane where it opens a channel that allows electrically charged ions to flow into the cell. This electric current flowing through the membrane triggers the neuron to fire (more on this in chapter 9) and send a nerve signal that travels all the way from the olfactory epithelium to the brain.
The lock-and-key mechanism proposes that the receptor molecules are complementary in shape to the odor molecules, which fit inside them. A simple analogy is the shape-fitting puzzles that toddlers enjoy, in which a block cut into a particular shape (say, a circle, square or triangle) has to fit into a wooden board with the complementary shape cut out. We can think of each odor molecule as one of the block shapes—so that, perhaps, an orange odorant such as limonene is a circle, an apple odorant is a square, and a banana odorant a triangle. We can then imagine each olfactory receptor as possessing an odorant binding pocket that is molded into the ideal shape for the olfactant molecule to fit neatly inside.
Of course, real molecules rarely come in such neat shapes, so real receptor proteins are presumed to have much more complex binding pockets to fit the more intricate shapes of real odorant molecules. Most are probably highly complex shapes similar to the active sites of enzymes that, as you will recall from chapter 3, bind substrate molecules. Indeed, odorant molecules are believed to interact with binding pockets in a fashion akin to the way that enzyme substrates are tethered into the active sites of enzymes (figure 3.4), or even the way that drugs interact with enzymes. Indeed, it has been argued that understanding the role that quantum mechanics plays in the interaction between olfactants and their receptors could eventually lead to more efficient drug design.
In any event, a clear prediction of the shape theory is that there should be some kind of correlation between the molecular shape of an odorant and its smell: similarly shaped odorant molecules should smell alike and very differently shaped molecules are likely to have sharply distinct odors.
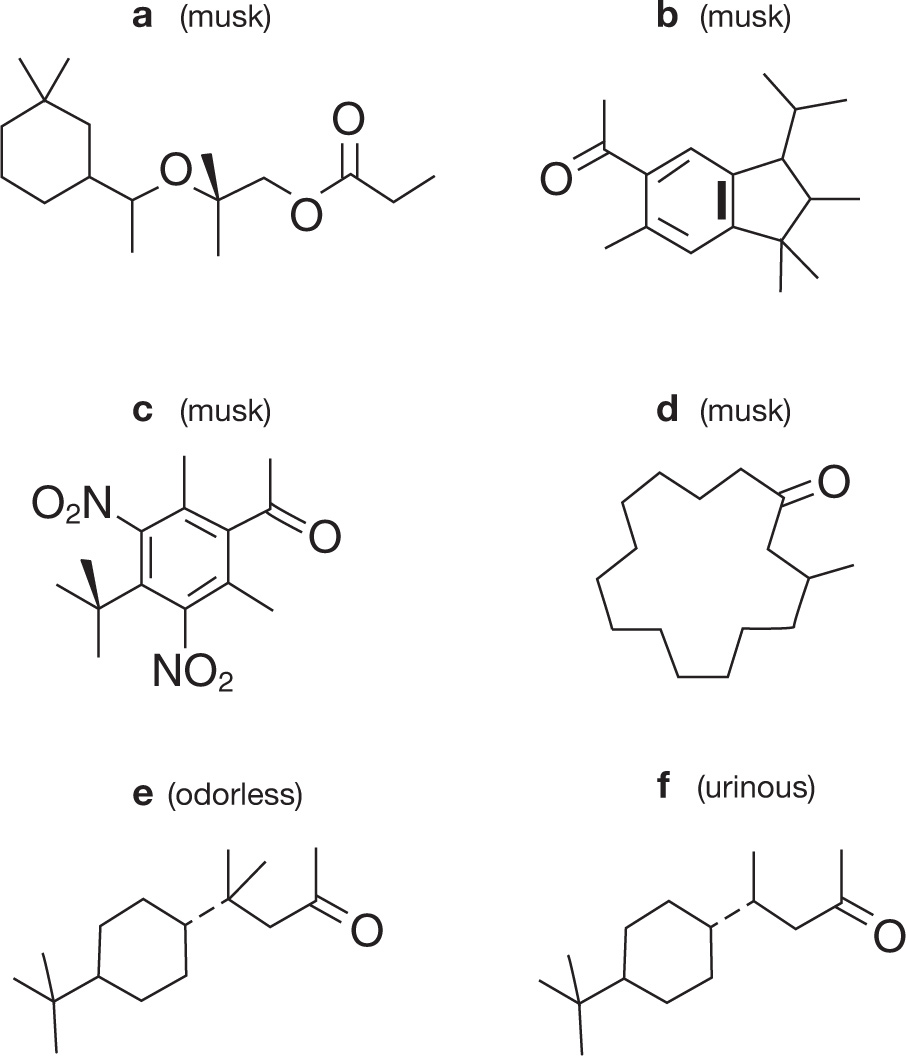
One of the most feared scents in human history was the smell of mustard or rotten hay in the trenches of the Great War. Invisible gases would float across no-man’s-land and just the faintest whiff of mustard (mustard gas) or musty hay (phosgene) might be sufficient to give a soldier a few precious seconds in which to don his mask before the deadly substance filled his lungs. The chemist Malcolm Dyson survived a mustard gas attack, and maybe it was this insight into the survival value of a keen nose that led him to ponder the nature of scent, because after the war he went on to synthesize many industrial compounds and to use his nose to smell the products of his synthetic reactions. But Dyson was puzzled by the apparent absence of any obvious relationship between the shape of a molecule and its smell. For example, many molecules that have very different shapes, such as the compounds in figure 5.2 a–d, smell the same—in this case, they all smell musky.*7 Conversely, compounds that have very similar structures (such as compounds e and f in the figure) often have very different smells—in this case compound f smells like urine whereas e has no smell at all.4
This far from straightforward connection between the shape of a molecule and its odor was, and still is, a major problem for the industrial manufacturers of perfumes, flavors and fragrances. Instead of being able to design a perfume in the way they might design the shape of the perfume bottle, perfumers are forced to rely on chemical synthesis by brute force and trial-and-error sniff tests by chemists such as Dyson. But Dyson noticed that odor groups (chemicals that smell the same) were often composed of compounds that incorporated the same chemical groups, for example, the oxygen atom linked to a carbon atom by a double C=O bond in the musky-smelling chemicals in figure 5.2. These chemical groups are the component parts of any large molecule and determine many of its properties, apparently including, as Dyson noted, its scent. Another set of compounds with a similar smell is the large number of chemicals, with diverse molecular shapes, that possess a sulphydryl (S–H) chemical group, in which a hydrogen atom is attached to a sulphur atom, and that have the characteristic rotten-egg smell. Dyson went on to propose that what the nose detects is not the shape of an entire molecule but rather a different physical feature, namely the frequency at which the molecular bonds between its atoms vibrate.
Figure 5.2: Molecules (a)–(d) have very different shapes but smell pretty much the same. Molecules (e) and (f) have nearly identical shapes but have very different smells.
In the late 1920s, when Dyson first made these claims, no one had any idea how to detect molecular vibrations. But on a voyage to Europe in the early 1920s the Indian physicist Chandrasekhara Venkata Raman was enchanted by “the wonderful blue opalescence of the Mediterranean sea” and speculated that “the phenomenon owed its origin to the scattering of light by molecules of water.” Normally, when light bounces off an atom or molecule it does so “elastically,” that is, without losing any energy, rather like a hard rubber ball bouncing off a rigid surface. Raman suggested that on rare occasions light can scatter “inelastically,” rather like a hard ball hitting a wooden bat and transferring some of its energy into the bat and the batsman (think of Bugs Bunny whacking a fast baseball so hard he sets both bat and bunny vibrating). In inelastic scattering, photons similarly lose energy to the molecular bonds they bump into, causing them to vibrate; the scattered light therefore emerges with less energy. Reducing the energy of light lowers its frequency and shifts its color toward the blue end of the spectrum, providing Raman with his “wonderful blue opalescence.”
Chemists utilize this principle to probe molecular structure. Essentially, light is shone on a chemical sample and the difference in color or frequency (hence energy) between the input and output light is recorded as a Raman spectrum for a particular chemical, which provides a kind of signature of its chemical bonds. The technique bears its inventor’s name, Raman spectroscopy, and earned him a Nobel Prize. When Dyson heard about Raman’s work he saw that it could provide a mechanism by which the nose might probe the molecular vibrations of odor molecules. He proposed that the nose “may be a spectroscope” capable of detecting the signature frequencies at which different chemical bonds vibrate. He even identified common frequencies in the Raman spectra of compounds that correlated with their odors. For example, all mercaptans (compounds that contain a terminal sulphur–hydrogen bond) share a particular Raman peak with frequency of 2567–2580. And they all stink of rotten eggs.
Dyson’s theory did at least account for the analytical nature of odors, but no one had the slightest idea how anything like Raman spectroscopy could be harnessed by our nose to provide the sense of smell. After all, not only would the scattered light need to be captured and analyzed by any biological spectroscope, there would also need to be a source of light in the first place.
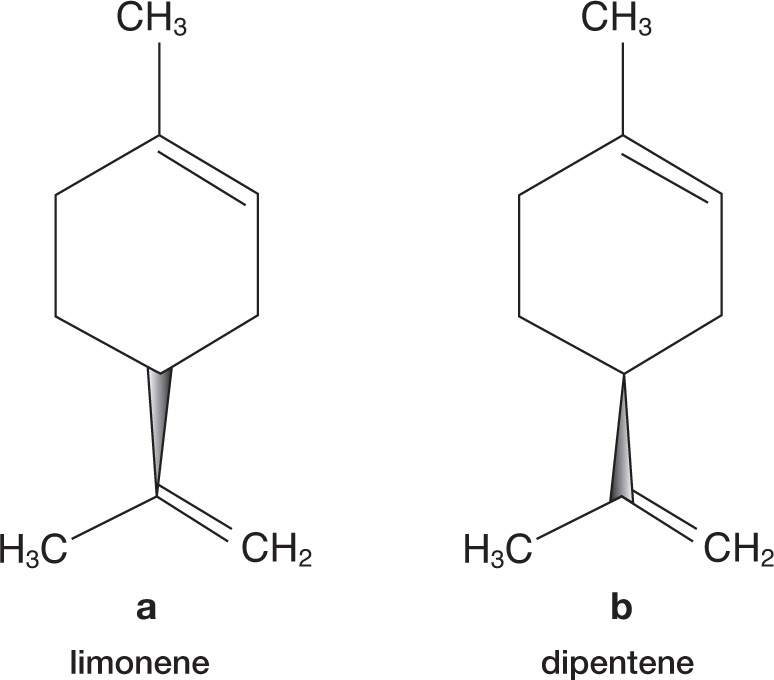
An even more serious deficiency with Dyson’s theory became apparent when it was discovered that the nose can easily differentiate molecules that have exactly the same chemical structure and identical Raman spectra, but are mirror images of each other. For example, the limonene molecule that is largely responsible for the smell of your orange can be described as a right-handed molecule. But there is a nearly identical molecule called dipentene that is its “left-handed” mirror image molecule (see figure 5.3, where the pointed, shaded area at the bottom of each part represents the carbon–carbon bond that points either below (a) or above (b) the page). Dipentene has the same molecular bonds as limonene and thereby gives an identical Raman spectrum but its odor is very different: it smells like turpentine. Molecules that come in left- and right-handed forms are described as being chiral,*8 and they often have quite different odors. Another chiral compound is carvone, the chemical found in seeds such as dill and caraway, which is responsible for the caraway scent; its mirror-image molecule smells like spearmint. A Raman spectroscopist would be unable to distinguish these compounds with a spectrometer, but a simple sniff does it easily. Clearly, smell cannot rely, at least not solely, on detecting molecular vibrations.
Figure 5.3: Limonene (a) and dipentene (b) are mirror-image molecules with very different smells. The molecules differ only in the orientation of the lower chemical group, which is represented as pointing below the page (bond pointing down) in limonene but above the page (bond pointing up) in dipentene. Of course, the dipentene molecules could be flipped over so that its chemical group points under the page like limonene, but then its double bond would flip round to the left, rather than the right, so it would still be different. The molecules are like left- and right-handed gloves.
These seemingly fatal flaws in the vibration theory of olfaction led to its eclipse by the lock and key theory through most of the latter half of the twentieth century, despite the best efforts of a few molecular vibration enthusiasts, such as the Canadian chemist Robert H. Wright, who provided a potential solution to the problem of left- and right-handed molecules that possess the same bonds but have different smells. He pointed out that the olfactory receptors are themselves likely to be chiral (coming in left- or right-handed forms), so they would hold an odorant molecule in a left-handed or right-handed way that would then present its bonds to the vibration detector differently. To take a musical analogy, the left-handed Jimi Hendrix (representing the olfactory receptor) generally held his guitar (the chiral odorant molecule) with its neck pointing to the right; whereas the right-handed Eric Clapton held his guitar (representing the mirror-image molecule) with the neck pointing left.*9 Both musicians could play the same riff (generate the same vibrations) on mirror-image guitars; but the sound picked up by a fixed microphone (representing the vibration detector part of the olfactory receptor) placed, say, just to the left of each musician, would be subtly different because their strings (the molecular bonds) are in different locations relative to the microphone. Wright proposed that chiral olfactory receptors detect the vibration frequencies of chemical bonds, but only when the bonds are in the right position: he claimed that the receptors come in right- or left-handed forms, just like guitar players. But with still no idea of how the biological vibration detector would actually work, the vibration theory remained on the margins of olfactory science.
However, the shape theory also has its problems. As we have already discussed, it has difficulty explaining odorant molecules with very different shapes but the same odor, and vice versa. To tackle these problems, Gordon Shepherd and Kensaku Mori came up with what is sometimes called the “weak shape,” or odotope, theory in 1994.5 The key difference between this and the classical shape theory lies in Shepherd and Mori’s proposal that, rather than the shape of the entire molecule being recognized by olfactory receptors, the receptors need only identify the shape of the component chemical groups. For example, as we have already pointed out, all the musky-smelling compounds in figure 5.2 have an oxygen atom linked to a carbon atom by a double bond. The odotope theory proposes that it is the shape of these chemical substructures, rather than that of the entire molecule, that is recognized by olfactory receptors. This theory makes better sense of the analytical nature of scent, but it suffers many of the same problems as the vibration theory when dealing with molecules containing the same chemical groups but arranged differently. Thus, neither odotope nor vibration theory can explain how pairs of chemicals can have different odors despite possessing the same chemical groups arranged differently on the same molecular scaffold. For example, both vanillin (which forms the primary component of natural vanilla) and isovanillin consist of a six-carbon ring with three identical chemical groups attached at different positions (figure 5.4). The odotope theory would predict that the identical chemical groups should smell the same. Yet vanilla smells, well, like vanilla, but isovanillin has a nasty phenolic (sweet medicinal) scent. Molecules with the same chemical basic parts—such as vanillin and isovanillin, shown here—can, nevertheless, have very different smells.
To deal with these problems, shape theorists generally propose a combination of odotope theory and some sort of overall chiral shape recognition mechanism. Nevertheless, this still cannot explain the equally common situation of mirror-image molecules actually having the same smell.*10 It suggests that they are being recognized by the same receptor, which is the molecular equivalent of having the kind of hand that would fit both a left-handed and a right-handed glove. It just doesn’t seem to make complete sense.
Figure 5.4: Molecules with the same chemical basic parts—such as vanillin and isovanillin, shown here—can, nevertheless, have very different smells.
Shape recognition is intuitively easy to understand: we routinely deal with shape complementarity every time we slip on a glove, turn a key in a lock or use a wrench to tighten a nut. Enzymes (which we saw in action in chapter 3), antibodies, hormone receptors and other biomolecules are also known to interact primarily via the geometric arrangement of their atoms and molecules; so it is not surprising that the shape theory of olfaction has received strong support from many biologists, including the olfaction receptor Nobel Prize winners, Richard Axel and Linda Buck.
Vibration-based communication is much less familiar to us, despite the fact that it is fundamental to at least two of our senses, vision and hearing. But whereas the physics of how the eye detects the vibration frequency of light and the ear records the vibration frequency of air are pretty well understood, no one had any idea, until recently, how the nose might detect the frequency of a molecular vibration.
Luca Turin was born in Lebanon in 1953 and studied physiology at University College London. After graduating, he moved to France to work at the National Center for Scientific Research, and it was in Nice that he experienced an olfactory epiphany on a visit to the Galeries Lafayette. In the middle of the perfume room was a display mounted by the Japanese company Shiseido of their new perfume, Nombre Noir, which Turin describes: “It was halfway between a rose and a violet, but without a trace of the sweetness of either, set instead against an austere, almost saintly background of cigar-box cedar notes. At the same time, it wasn’t dry, and seemed to be glistening with a liquid freshness that made its deep colors glow like a stained-glass window.”6 Turin’s encounter with the Japanese scent was to inspire a lifelong quest to discover the secret of how molecules floating into the nose could create such evocative experiences.
Like Dyson before him, Turin was convinced that the correlations between vibrational spectra and scent couldn’t be mere coincidence. He was persuaded by Dyson’s argument that the olfactory receptors must somehow be detecting molecular vibrations. But, unlike Dyson, Turin proposed a speculative, yet plausible, molecular mechanism by which biomolecules could detect the vibrations of chemical bonds via quantum tunneling of electrons.7
Tunneling, you may remember from chapter 1, is the peculiar quantum mechanical property that arises from the ability of particles such as electrons or protons to behave as waves of probability capable of seeping through barriers that are impenetrable via any classical route. We discovered in chapter 3 how it plays a crucial role in many enzymes’ reactions. While Turin was puzzling over the secret of scent, he came across a paper describing a new analytical chemical technique called inelastic electron tunneling spectroscopy (IETS). In IETS, two metal plates are placed very close to each other, separated by a tiny gap. If a voltage is applied between the plates, electrons will gather on one plate, making it negatively charged (the donor), and will experience an attractive force from the other, positively charged, plate (the acceptor). Considered classically, the electrons lack the energy to jump across the insulating gap between the plates; but electrons are quantum objects and, if the gap is small enough, they can quantum tunnel across from donor to acceptor. This process is called elastic tunneling because the electrons do not gain or lose energy in the process.
However, there is a crucial additional condition: an electron can tunnel elastically from its donor site across to the acceptor site only if there is an empty slot available for it at the exact same energy. If the nearest available gap in the acceptor is at lower energy, then the electron must lose some of its energy to make the jump. This process is called inelastic tunneling. But the dumped energy needs to go somewhere, otherwise the electron can’t tunnel. If a chemical is placed in the gap between the plates, then an electron can tunnel across so long as it is able to donate its excess energy to the chemical—which it can do so long as the molecules in the gap have bonds capable of vibrating at just the right frequency, corresponding to that of the dumped energy. Having passed on their excess energy in this way, these “inelastically” tunneling electrons arrive on the acceptor plate with slightly lower energy; so by analyzing the energy differences between electrons leaving the donor site and arriving at the acceptor site, inelastic electron tunneling spectroscopy probes the nature of a chemical’s molecular bonds.
To return to our musical analogy, if you have ever played a stringed instrument you will know that it is possible to get a note out of a string without even touching it, by resonance. Indeed, this trick can be used to tune a guitar. If you fold a tiny scrap of very light paper over one of the strings, then pluck the same note on an adjacent string, you can make the paper scrap pop off without that string being touched at all. This is because, once you get the tuning just right, the plucked string sets the air vibrating, and the vibrating air passes the vibration on to the unplucked string, setting off its vibration in resonance with the plucked string. In IETS, the electron only pops off the donor site if the chemical between the two plates has a bond similarly tuned to just the right frequency for it to make the jump. In effect, the tunneling electron loses energy by plucking a molecular bond on its quantum journey across the plates.
Turin proposed that olfactory receptors work in a similar way but with a single molecule—the olfactory receptor—taking the place of the IETS plates and gap. He envisaged an electron located first at a donor site in the receptor molecule. As in IETS, the electron could potentially tunnel to an acceptor site in the same molecule but, he proposed, it is prevented from doing so by an energy discrepancy between the two sites. However, if the receptor captures an odorant molecule that possesses a bond tuned to just the right vibrational frequency, then the electron can pop from donor to acceptor via tunneling while simultaneously transferring just the right amount of energy to the odorant, effectively plucking one of its molecular bonds. Turin proposed that the tunneled electron, now sitting in the acceptor site, causes the release of the tethered G protein molecular torpedo, causing the olfactory neuron to fire and thereby send a signal off toward the brain, allowing us to “experience” the scent of the orange.
Turin managed to amass a lot of circumstantial evidence for his quantum vibrational theory. For example, as already mentioned, sulphur–hydrogen compounds usually have a strong rotten-egg smell, and they all possess a sulphur–hydrogen molecular bond that vibrates at around 76 terahertz (76 trillion oscillations per second). His theory makes a strong prediction: any other compound associated with a bond vibrational frequency of 76 terahertz should also have a rotten-egg smell, irrespective of its shape. Unfortunately, very few other compounds have that same vibration band in their spectra. Turin searched through the spectroscopy literature for a molecule with the same vibration. Finally, he discovered that the terminal boron–hydrogen bonds in chemicals called boranes have vibrations centered at 78 terahertz, which is quite close to the 76 terahertz S–H vibration. But what did boranes smell like? That information wasn’t available in the spectroscopy literature, and the chemicals were so exotic that he couldn’t get hold of any to have a sniff. But he found an old paper that described them as smelling repulsive, which is a term often used to describe a sulphurous smell. In fact, it turns out that boranes are the only known non-sulphur molecules that have the same rotten-egg stink as hydrogen sulphide: for example decaborane, which is made up of only boron and hydrogen atoms (chemical formula B10H14).
This discovery that among the literally thousands of chemicals that have been sniffed, the only one to stink like hydrogen sulphide is a molecule that shares the same vibrational frequency provided strong support for the vibrational theory of smell. Remember that perfumiers have been trying for decades to unlock the molecular key to scent. Turin had contrived to do what none of their chemists had managed: predict a scent from theory alone. It was the chemical equivalent of predicting the scent of a perfume from the shape of its bottle. Turin’s theory also provided a biologically plausible quantum mechanism that would allow a biomolecule to detect a molecular vibration. But a “plausible mechanism” is not enough. Was it correct?
The vibration theory had scored some encouraging successes, as with decaborane, but it still suffered from similar problems to the shape theory, such as the mirror-image molecules (e.g. limonene and dipentene) having very different scents, but identical vibrational spectra. Turin decided to test another prediction of his theory. You may remember that the tunneling theory of enzyme action (chapter 3) was tested by replacing the most common form of hydrogen with one of its heavier isotopes, such as deuterium, in order to make use of the kinetic isotope effect. Turin tried a similar trick with an odorant called acetophenone, described as having a “pungent sweet odor … resembling that of hawthorn or a harsh orange blossom.” He purchased a very expensive batch of the chemical in which each of the eight hydrogen atoms in its carbon–hydrogen bonds had been replaced with deuterium. The heavier atoms, like heavier guitar strings, will vibrate at lower frequencies: a normal carbon–hydrogen bond vibrates with a high note frequency of between 85 and 93 terahertz, but if deuterium is substituted for the hydrogen then the vibration frequency of the carbon–deuterium bond drops down to about 66 terahertz. The “deuterated” chemical therefore has a very different vibrational spectrum from the hydrogen one. But does it smell any different? Turin locked the door of his laboratory before gingerly sniffing both compounds. He was convinced that they “smelled different, the deuterated one less sweet, more solvent-like.”8 Even after carefully purifying each compound he was convinced that the hydrogen and the deuterated forms smelled very different. His theory, he claimed, was vindicated.
Turin’s research brought him to the attention of investors who provided the financial backing needed to set up a new company, Flexitral, devoted to exploiting his quantum vibration ideas to manufacture new fragrances. The author Chandler Burr even wrote a book describing Turin’s quest for the molecular mechanisms of smell;9 and the BBC filmed a documentary about his work.
But many were still far from convinced, particularly the shape theory enthusiasts. Leslie Vosshall and Andreas Keller from the Rockefeller University repeated the sniff tests with normal and deuterated acetophenone but, rather than relying on Turin’s highly tuned nose, they asked twenty-four naïve subjects whether they could distinguish between the compounds. The results were unequivocal: no difference in smell. Their paper, published in Nature Neuroscience in 2004,10 was accompanied by an editorial in which the vibration theory of scent was described as having “no credence in scientific circles.”
But, as any medical researcher will tell you, trials in humans can be confounded by all sorts of complications, such as the expectations of the subjects and their experiences prior to the experiment. To avoid these problems, a team led by Efthimios Skoulakis of the Alexander Fleming Institute in Greece and including researchers from MIT, among them Luca Turin, decided to switch to a much better-behaved species: laboratory-bred fruit flies. The team devised the fruit-fly equivalent of Gabriele Gerlach’s flume choice experiment with coral reef fish, described earlier in this chapter. They called it the fly “T maze” experiment. The flies were introduced into a T-shaped maze through the stem and encouraged to fly to the junction where they would have to make a choice whether to go left or right. Scented air was pumped into each arm, so by counting the number of flies that went in each direction the researchers could work out whether the flies could distinguish between the odorants loaded respectively into the left and right airflows.
The group first investigated whether the flies could smell acetophenone. Indeed they could: just a dab of the chemical in the end of the right-hand arm of the maze was sufficient to persuade nearly all the flies to fly toward its fruity odor. The group then substituted deuterium for the hydrogen atoms in the acetophenone; but, in a new twist, they replaced either three, five or all eight hydrogen atoms with deuterium and tested each version of the chemical separately, with the undeuterated compound always in the other arm of the maze. Their results were remarkable. With only three deuterium atoms, the flies lost their preference for turning right at the junction and randomly went left or right. But when the researchers loaded the right arm with the five- or eight-atom-substituted chemical the flies resolutely turned left, away from the deuterated odorant. It seemed they could smell the difference between the normal and the heavily deuterated form of acetophenone, and now they didn’t like what they smelled. The team tested two additional odorants and found that the flies could easily distinguish between hydrogen and deuterium forms of octanol, but not between the corresponding forms of benzaldehyde. To demonstrate that the flies were using their sense of smell to sniff out the deuterium bonds, the researchers also tested a mutant strain of fly that lacks functional olfactory receptors. As expected, these anosmic*11 mutants were completely unable to distinguish between the hydrogen and deuterated odorants.
Using a Pavlovian conditioning setup, the researchers even managed to train the flies to associate certain forms of chemicals with punishment: a mild electric shock to their feet. The team was then able to perform an even more remarkable test of the vibration theory. They first trained flies to avoid compounds with the carbon–deuterium bond, with its characteristic vibration at 66 terahertz. They then wanted to discover whether this avoidance could be generalized to very different compounds that happened to possess a bond vibration at the same frequency. And it could. The team discovered that the flies trained to avoid compounds with the carbon–deuterium bond also avoided compounds called nitriles whose carbon–nitrogen bond vibrates at the same frequency, despite being chemically very different. The study provided strong support for a vibration component of olfaction, at least in flies, and was published in the prestigious science journal Proceedings of the National Academy of Science in 2011.11
The following year Skoulakis and Turin teamed up with University College London researchers to return to the delicate question of whether humans can also smell by vibrations. Rather than relying solely on Turin’s highly sensitive sense of smell, the team recruited eleven sniffing subjects. They first confirmed the Vosshall and Keller result: their test subjects could not sniff out the carbon–deuterium bonds in acetophenone. But the team reckoned that with only eight carbon–hydrogen bonds, the signal from the deuterated form of the chemical might be rather weak and thus indistinguishable to the average nose; so they decided to investigate more complex musky-smelling molecules (like those in figure 5.2) that have up to twenty-eight hydrogen atoms, all of which could be replaced with deuterium. This time, in contrast to the acetophenone trial, all of their eleven subjects could easily distinguish between normal and fully deuterated musk. Maybe humans really can sniff out molecules tuned to different frequencies after all.
One of the criticisms leveled at the quantum vibration theory was that its theoretical foundation was all rather vague. This has now been addressed by a team of physicists from University College London who, in 2007, carried out the “hard-nosed” (if you’ll pardon the pun) quantum calculations behind the tunneling theory and concluded that it was “consistent both with the underlying physics and with observed features of smell, provided the receptor has certain general properties.”12 One of the team, Jenny Brookes, even went on to propose a solution to that niggling problem of mirror-image molecules such as limonene and dipentene (figure 5.3) that have the same vibrations but very different odors.
In fact, it was Jenny’s supervisor and mentor, the late Professor Marshall Stoneham, who first came up with what is sometimes called the swipe-card model. Stoneham was one of the leading UK physicists of his generation with interests that ranged from nuclear safety to quantum computing, biology and, appropriately for this chapter, music: he played the French horn. Their theory is a quantum mechanical elaboration on Robert H. Wright’s idea that both the shape of the olfactory receptor and the bond vibrations of the odorant molecule play a role in smell. They proposed that the binding pocket of the olfactory receptor works like a swipe-card machine. Swipe cards have a magnetic strip that is read to generate an electric current in the swipe-card machine. But not everything fits into a swipe-card reader: the card has to be the right shape and thickness, with its magnetic strip in the right place, before you can even use it and check whether the machine recognizes it. Brookes and her colleagues proposed that olfactory receptors work in a similar way. An odorant molecule, the team postulated, must first fit into a left- or right-handed chiral binding pocket, rather like a credit card fitting in a card reader. So odorants with the same bonds but different shapes, such as a left-handed and a right-handed version of the same molecule, will be picked up by different receptors. Only after either odorant has fitted into its complementary receptor does it have the potential to stimulate the vibration-induced electron tunneling event to make the receptor neuron fire; but because the left-handed molecule will be firing a left-handed receptor, it will smell different from a right-handed molecule firing a right-handed receptor.
If we return to our musical analogy one final time, with the guitar acting as the odorant molecule and the guitar strings as the molecular bonds that need to be plucked, then the receptors come in Eric Clapton or Jimi Hendrix forms. Both can play the same molecular notes, but right- or left-handed molecules have to be picked up by right- or left-handed receptors, just as right-handed guitars have to be played by right-handed guitarists. So although limonene and dipentene have the same vibrations, they have to be held by left- or right-handed olfactory receptors. The different receptors will be wired to different regions of the brain and will thereby generate different smells. This combination of shape and quantum vibration recognition at last provides a model that fits nearly all the experimental data.
Of course, the fact that this model fits the data doesn’t itself prove that there is a quantum basis for olfaction. The experimental data provide strong evidence for any theory of olfaction that involves both shape and vibration. No experiment has yet directly tested whether quantum tunneling is involved in smell. However, so far at least, inelastic quantum tunneling by electrons is the only known mechanism that provides a plausible explanation for how proteins can detect vibrations in odor molecules.
The vital piece of the olfaction puzzle that is still missing is the structure of olfactory receptors. Knowing this would make it easier to find the answer to key questions such as how snugly the binding pockets are tailored to each odorant molecule, whether mirror-image molecules bind to the same receptors, and whether the receptor molecules possess electron donor and acceptor sites suitably positioned to promote inelastic electron tunneling. Yet despite many years of effort by some of the top structural biology groups around the world, no one has yet managed to isolate olfactory receptor molecules susceptible to study in the same way that allowed quantum mechanical mechanisms to be elucidated in enzymes (chapter 3) or photosynthetic pigment proteins (chapter 4). The problem is that in its natural state the olfactory receptor is embedded in the cell’s membrane, a bit like a jellyfish floating on the surface of the sea. Taking the receptor protein out of the membrane is like taking a jellyfish out of the ocean: it is just not going to retain its shape. And no one has yet found a way of determining the structure of proteins while they remain embedded in cell membranes.
So, although considerable controversy remains, the only theory that provides an explanation of how flies and humans can distinguish the smells of normal and deuterated compounds is based on the quantum mechanical mechanism of inelastic electron tunneling. Experiments have recently shown that, as well as flies and humans, other insects and even fish are able to sniff the differences between hydrogen and deuterium bonds. If quantum smelling is found in such a range of creatures it is likely to be very widespread. Humans, fruit flies, anemonefish and a host of other animals are probably harnessing the ability of an electron to vanish from one point in space and instantly materialize in another so that they can capture that “message from a material reality” and find food, or a mate—or their way home.
*1 Sadly, that popularity now threatens the animal in the wild, as it has become a favorite of poachers who over-collect the fish to feed the burgeoning aquarium market in anemonefish. So don’t keep Nemo in your home; he or she belongs on real coral reefs!
*2 The stronger tides at this time are thought to aid dispersal.
*3 Note the implicit racism.
*4 1-methyl-4-(1-methylethenyl)-cyclohexene.
*5 A picoamp is one trillionth (10−12) of an amp.
*6 In this context “expressed” means a gene that is active in the sense that its information is copied into RNA, which then feeds into the protein synthesis machinery to make the protein encoded by that gene, such as an enzyme or particular olfactory receptor.
*7 Traditionally, musk was obtained from a number of natural sources, including the sex glands of the musk deer, the face glands of the musk ox, the feces of the pine marten and the urine of the rock badger. But nearly all perfume musk is now synthetic.
*8 A chiral molecule has a non-superposable mirror image.
*9 In fact Hendrix generally played a right-handed guitar upside down but he reversed the strings so that high E would be in the same position as if he were playing a left-handed guitar.
*10 For example, (4S,4aS,8aR)-(K)-geosmin and its mirror-image molecule (4R,4aR,8aS)-(C)-geosmin, which both smell “earthy, musty.”
*11 From anosmia, the inability to detect odors: a condition that, in humans, is usually associated with trauma to the nasal epithelium, though rare genetic forms are known.