


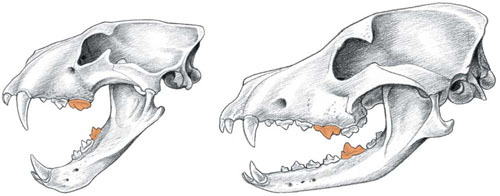
Skull of leopard (LEFT) and dog (RIGHT), with the carnassials indicated (in orange). The shortened felid jaw accommodates only specialised meat-eating teeth, whereas the dog retains a more diverse dentition with a full battery of premolars in front of the carnassials and two pairs of crushing molars behind the carnassials.
All cats are carnivores – they eat meat – but the label signifies far more than their diet. Any animal that eats meat can be described as carnivorous (even some plants are carnivorous), but to a biologist, this does not make it a carnivore. That designation belongs solely to the members of the order Carnivora. There are approximately 4 630 living mammals, which are classified into 26 different orders based on their evolutionary relationships. Our own, the order Primates, includes some 256 species ranging from bushbabies to humans and, although many primates eat meat, this does not warrant their being classified as carnivores. The order Carnivora comprises the true carnivores, some 270 mainly predatory species which, despite astonishing variation in size and shape, are united by their shared evolutionary origins. The smallest carnivore in the world, the tiny least weasel of North America and Eurasia, is slender enough to squeeze through a wedding ring and weighs 10 000 times less than the largest terrestrial species, the polar bear. The carnivores make up one of the largest mammalian orders, are found on every major landmass, including Antarctica, and occur from the hyper-arid interior of the Sahara to the polar snows of the Arctic.
What makes them all carnivores? The answer lies in their unique cheek teeth, a modification of the mammalian molar (technically, the last upper pre-molar and the first lower molar) into a pair of flattened cutting shears called the carnassials. Like the blades of scissors, the carnassials slide closely past one another, slicing off chunks of meat as efficiently as a knife. To taxonomists, the scientists who classify species, the carnassials provide an unmistakable diagnostic attribute of the true carnivore. A number of modern species such as giant pandas, sea otters and aardwolves lack obvious carnassials (explained below) but their ancestors possessed them, as do some of their living relatives; they are clearly carnivores, even if some of them no longer eat meat. As we will see in this chapter, the carnassial not only connects all carnivores to one another but, far more importantly, it allowed them to become the dominant group of predatory mammals on the planet.

Reconstruction of a miacid, the earliest carnivore type. Miacids were similar in anatomy and lifestyle to primitive carnivores living today, such as the African palm civet and probably shared a similar diet of small vertebrate prey, invertebrates, eggs and some plant matter.

A ‘primitive’ carnivore, the African palm civet. Here, primitive does not imply inferiority but rather indicates that the civet has retained many features of anatomy and ecology that arose early in carnivore evolution. Their persistence demonstrates that, in fact, such features are well suited for survival.
The carnivore carnassial made its appearance in a rather inconspicuous group of small predators called the miacids. First recorded from the fossil record of the northern hemisphere around 60 million years ago, miacids were slender, small-bodied forest dwellers with skeletons very similar to those of modern genets. They had supple spines, long tails, retractile claws and a somewhat opposable first digit on the front foot, a set of characters well suited to the dense forests that dominated the Earth at the time. They were equipped for an arboreal lifestyle and were able to hunt in the tree-tops, though most likely they also spent considerable time foraging on the ground. Miacid teeth were very similar to those of the modern African palm civet and, like the civet, miacids were generalists. Their carnassials efficiently processed small vertebrate prey such as small mammals, birds and reptiles, but their dentition had not progressed so far along the carnivoran specialisation that they could not also cope with a more omnivorous selection that included invertebrates, eggs, fruit, nectar and seeds.
In fact, while the emergence of the carnassial would be fundamental to the predatory success of the carnivores, most species – fossil and living – actually consume a varied diet like that of their miacid ancestors. This dietary spread reflects an evolutionary plasticity thought to be the reason the carnivores have proliferated while other mammals that followed a predatory path declined and, in most cases, died out. Again, the key lies in their carnassial dentition, yet numerous other mammalian groups have evolved a similar battery of meat-shearing teeth. Why is the carnassial so special?
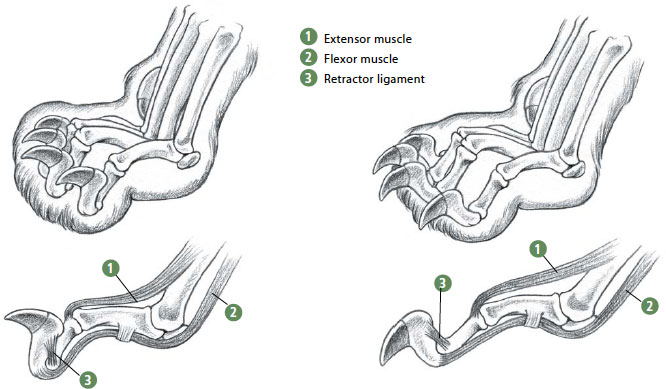
ABOVE AND BELOW: Retractile claws. Retraction is actually the relaxed condition (as in this sleeping lioness’ foot, BELOW LEFT), in which the elastic retractor ligament automatically withdraws the claw when it’s not in use. To extend the claws, the flexor muscles contract powerfully against the action of this ligament and contraction of the extensor muscles holds the digit firmly in the extended position. The action is more accurately termed protractile, meaning actively extended. Cheetahs (BELOW RIGHT) retain the ability to protract their claws but it is poorly developed and they lack the protective claw sheaths seen in other cats, so their claws protrude conspicuously even when relaxed.


A feeding leopard displays the action of the carnassials. When eating, cats turn their heads to the side, employing the carnassials in a slicing action that shears off chunks of meat to be swallowed with little additional chewing.
Evolutionary biologists think it has to do with the position of the shearing teeth. In true carnivores, the carnassials are located in the middle of the tooth row, in front of the rear crushing teeth, the posterior molars. In other mammals that experimented with carnivory as a way of life, it was these rear molars that were modified into the flattened slicing form suitable for dealing with meat.
The advantage for the carnivores was adaptability. With the carnassials responsible for shearing, the rear molars were free to follow various evolutionary routes closed to those predators that used them for processing meat. The molars of carnivores could become more robust, with reduced shearing edges and greater crushing surfaces, enabling bones, insects, seeds, fruit and plant matter to be exploited. The result, a dual-purpose dentition that both shears and crushes (using the carnassials and molars respectively), is typified in dogs, and in fact most modern carnivores have elements of both. From this arrangement, the transition to an even more herbivorous diet is possible: the carnassials themselves gradually become less blade-like and more like molars, well equipped to deal with tough vegetation. In a handful of species such as the giant panda and red panda (which actually belong to different carnivore families; see text box ‘Carnivore families’ below), the carnassial shear has been lost entirely and they are completely herbivorous.
In the alternative evolutionary route, the rear molars undergo gradual reduction until there is little or no crushing ability and the dentition is dominated by the carnassials; in other words, teeth that can handle only meat. This hyper-carnivorous type is most developed in the cats (but is not exclusive to them) and indeed modern cats have lost all their rear molars or retain them only as residual, useless pegs.
The flexibility of the carnassial-molar combination was the evolutionary catalyst enabling the carnivores to rise to dominance over other, early mammals that had committed their rear molars to carnivory. Ironically, it was the carnivores’ ability to exploit food sources other than meat that probably catapulted them ahead in the evolutionary race and indeed, the more generalist, dual-purpose dentition as seen in dogs has always been the most widespread combination in carnivore evolution. The hyper-carnivorous specialisation in the cats makes them the most predatory of all carnivore families (discussed in Chapter 2), and, as we will see later, perhaps the most vulnerable to extinction.

The needle-like canines of this black-footed cat are suited for puncturing the skulls and vertebral columns of small prey such as rodents and small birds.
Carnivore families

TOP ROW, LEFT TO RIGHT: tiger, dwarf mongoose, grey wolf, white-nosed coati, Cape fur seal, walrus.
BOTTOM ROW, LEFT TO RIGHT: aardwolf, large-spotted genet, brown bear, Cape clawless otter, harbour seal.
All modern carnivores trace their origins back to the miacids, with the carnassial specialisation an unambiguous and unbroken connection. Today’s 270 species of carnivores are classified into 11 families. Each family comprises a group of species more closely related to one another than to the members of the other families which, in most cases, is obvious; all cats belong to one family, the dogs to another and so on. The 11 families are further grouped into two major divisions (known as sub-orders) reflecting a major separation that occurred early in miacid evolution. The dog sub-order (Caniformia) is made up of dogs, bears, raccoons, weasels, seals, sea lions and walruses, and the cat sub-order (Feliformia) comprises cats, hyaenas, genets and mongooses. This illustrates, for example, that despite their dog-like appearance, hyaenas are actually more closely related to cats than to the canids.
The number of carnivore species and how they are classified is still hotly debated. Seals, sea lions and walruses are sometimes accorded their own full order, the Pinnipedia, though they clearly belong on the Caniformia branch of carnivore evolution. A few carnivore families are contentious; walruses are sometimes grouped with fur seals, and three groups have recently and controversially been accorded their own family: Malagasy civets and mongooses (Eupleridae), skunks (Mephitidae) and red pandas (Ailuridae). Even the number of species remains unclear, with the cat family a case in point. Depending on the authority, there are between 36 and 40 cats; see the cat phylogeny chart on page 44 for details.

Proailurus is the earliest recognisable felid, with 30 million-year-old fossils from France marking the beginnings of the cat family. Proailurus is also the earliest cat known from the New World, though the oldest fossils (from modern-day Nebraska) are only 16 million years old.
The exact origin of the cats from their miacid ancestors is obscured by millions of years without fossils. Nonetheless, we can be fairly certain that while carnivores arose about 60 million years ago, cats have been on the scene for only about half as long. Miacids had been following a variety of evolutionary routes that would ultimately lead to all modern families of carnivores for at least 30 million years before a species appeared that was clearly a cat. It was called Proailurus, the ‘dawn cat’. The oldest fossils of Proailurus come from modern-day Eurasia which, together with a handful of European miacids that are clearly the ancestors of the cat sub-order Feliformia, places the origin of the cats firmly in the Old World.
The robustly built dawn cat was about the size of a male caracal but it had some features unlike any modern felid. Chief among them, Proailurus had more teeth; although its carnassials were characteristically feline, the dawn cat had not yet lost the rear molars of its miacid ancestors. Similarly, the dawn cat’s body plan falls somewhere between miacids and cats, with a suite of adaptations suggesting it was more arboreal than most felids: supple ankles and wrists capable of wide rotation to grasp branches, a sinuous, elongated back suited for vertical as well as horizontal movement, and a stance that used the heels as well as the toes, increasing the area of the foot in contact with branches. Although there is no cat alive today with the same combination of features, the closest modern analogue is a distant cat relative, the fossa. Largest of the viverrid family and restricted to Madagascar, the fossa looks feline enough to have originally been classified as a cat. Fossas hunt on the ground but are equally adept at climbing and leaping from branch to branch in pursuit of tree-living prey such as lemurs. Proailurus probably did likewise, pursuing a lifestyle more arboreal than that of any cat today.
Following the appearance of the dawn cat, there is little in the fossil record for 10 million years to suggest that cats would prosper. In fact, although Proailurus persisted for at least 14 million years, there are so few felid fossils towards the end of the dawn cat’s reign that palaeontologists refer to this period as the ‘cat gap’ (between 23 and 17 million years ago). Filling the vacuum, other carnivore families fielded their own experiments in hyper-carnivory. Dogs, bears and hyaenas as well as two extinct carnivore families, the nimravids or paleo-cats (family Nimravidae) and the bear-dogs (family Amphicyonidae), all evolved cat-like forms to occupy the vacant felid niche. The turning point for the cats came about with the appearance of a new genus of felids, Pseudaelurus.

All cats are digitigrade, meaning they walk on their toes. In comparison, plantigrade carnivores such as this brown bear walk using the entire foot – similar to the way humans place their feet.
Pseudaelurus (a misnomer meaning ‘false cat’) is probably descended directly from the dawn cat but had lost some of its progenitor’s more ancient features. Like modern cats, Pseudaelurus’ rear molars were greatly reduced and it walked on its toes (digitigrade), perhaps slightly less so than most modern species but just enough for a palaeontologist to discern the difference. It still had the dawn cat’s elongated spine but otherwise, it would have been very reminiscent of a small leopard.
African sabre-tooths
Based on the relative numbers of fossils, sabre-toothed cats dominated conical-toothed species in Africa for over 15 million years. At least eight species of large sabre-tooths are known from Africa, ranging in size from slightly smaller than a leopard to as large as a male lion. One of the most specialised was Megantereon, about the size of a large, powerfully built leopard. Megantereon had proportionally the longest canines of any African sabre-tooth and is thought to have killed its prey with a canine shear bite – inflicting a deep, massive wound to the throat or abdomen to sever vital blood vessels. Ungulates were probably held with the bite until they succumbed to blood loss or suffocation (much as modern big cats kill their prey), but the technique might also have been effective against juvenile mega-vertebrates such as elephants, rhinos and their extinct relatives. By speedily inflicting a devastating shear bite and retreating in the face of irate parental defence, Megantereon could simply wait until the injured calf died of blood loss before moving in to claim its meal. Megantereon and at least two other species of African sabre-tooths occurred alongside modern cheetahs, leopards and lions for two million years before dying out around 1.5 million years ago.

The sabre-toothed cat Megantereon with its kill, a juvenile hippo.

The African large carnivore guild around three million years ago. Living species: lion 2 , spotted hyaena 6 , leopard 7 , cheetah 8 and striped hyaena 9 . Note – the modern African wild dog and brown hyaena did not appear on the scene until more recently. Extinct species: the sabre-toothed cats Homotherium 1 , Megantereon 3 , Dinofelis 4 , the giant hyaena Pachycrocuta 5 and the hunting hyaena Chasmaportethe 10 .
The emergence of Pseudaelurus heralded a new era in the evolution of the cats. From the mid-Miocene around 17 million years ago, the numbers of felid species appearing in the fossil record increased rapidly. At the same time, the number of cat-like species from other carnivore families declined. It is not clear why the true cats suddenly became so good at being cats, but they evidently out-shone all other families in the competition for hyper-carnivory. Almost certainly, global climate change contributed to their success. The world had been undergoing a gradual cooling and drying for millions of years at the time Pseudaelurus arrived, so that the humid, dense forests during the time of Proailurus were giving way to open savanna woodlands and plains. Ungulates, primates, rodents and birds emerged from the forests, evolving into a multitude of new forms to exploit the abundant grasses, and tubers that arose in open habitats. Pseudaelurus followed them and, as its prey species evolved larger body size and greater speed to evade predators in open habitats, so too did Pseudaelurus. By around 15 million years ago, Pseudaelurus had evolved into a number of species with a global distribution.
The phylogeny of the cats
The exact evolutionary relationships of modern cats are imprecisely known. Most biologists agree on a total of 36 cats but three species are sometimes sub-divided, inflating the number to 40. An isolated island population of the Asian leopard cat, called the Iriomote cat, is sometimes accorded its own species, and some biologists believe that the South American pampas cat actually comprises three species. Finally, taxonomists argue over whether the domestic cat is the same species as its wild progenitor, the African wildcat, or a full species in its own right. None of the ‘splitting’ arguments is supported by genetic analyses. Evaluating genetic differences between cats also reveals clusters of related species called lineages; the members of a lineage are more closely related to one another than to the species of another lineage. The living cats fall into eight distinct lineages which began separating from their shared Pseudaelurus ancestors around 12 million years ago.
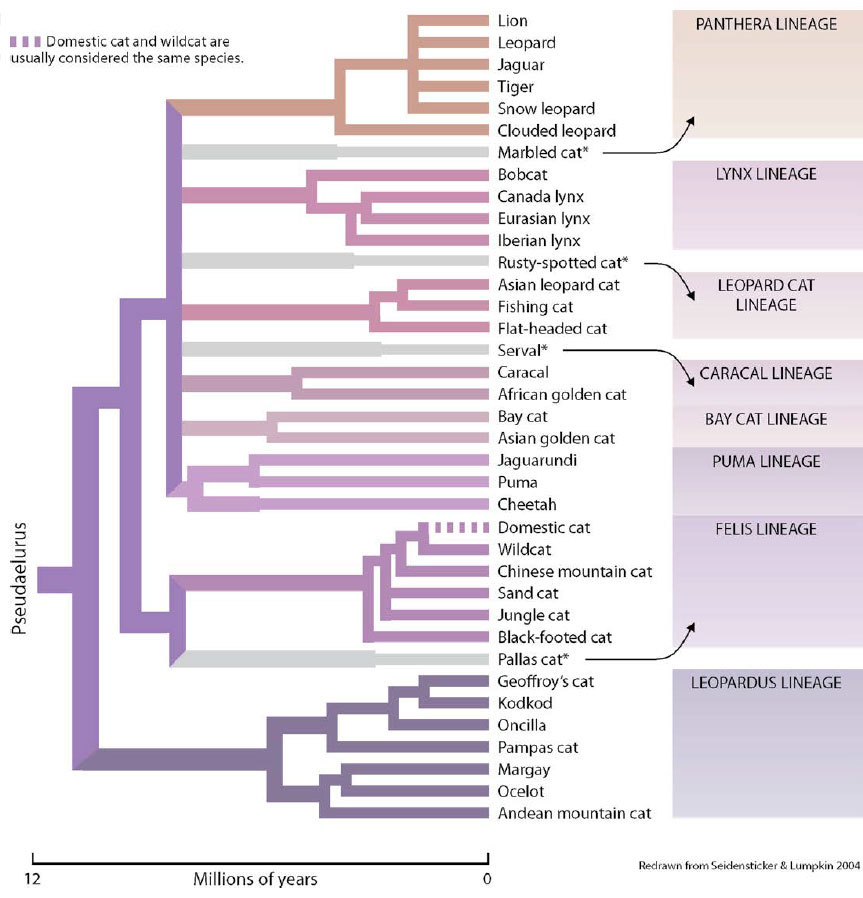
* The position in the felid phylogeny is still unclear for the marbled cat, rusty-spotted cat, serval and pallas cat; arrows indicate the lineage to which each is thought to be most closely related.
The rapid radiation of Pseudaelurus laid the evolutionary foundations that would lead to all true cats, including every species living today and many extinct ones. Pseudaelurus was also responsible for seeding Africa with felids, with the earliest evidence of cats on the continent coming from modern-day Libya. As everywhere, the fossil record of cats in Africa is patchy, but it is clear that, until very recently, it was dominated by the sabre-tooths (see text box ‘African sabre-tooths’, pages 42–43). Sabre-toothed cats belong with all modern species in the family Felidae, but they represent an ancient lineage which diverged from the evolutionary route that would lead to today’s cats. Both lineages arose from a shared Pseudaelurus ancestor about 15 million years ago, but only one of them developed the spectacularly elongated canine teeth known as sabres. The sabre-tooth lineage spawned a host of remarkable species, which are grouped together in their own felid sub-family, the Machairodontinae. The other group retained the conical tooth shape of its Pseudaelurus forebears, leading eventually to all modern cats (sub-family Felinae).
Compared to their sabre-toothed cousins, fossil evidence of the conical-toothed cats in Africa is sparse and we know very little about the immediate ancestry of today’s African cats. Surprisingly, given that large species usually leave more fossils, Africa’s three largest cats – the lion, leopard and cheetah – are the most mysterious. There are no obvious ancestors in the fossil record, until all three species appear at the same time in Tanzanian deposits that date back around 3.5 million years. Some older fragments may represent earlier appearances or possibly even evidence of their immediate ancestors, but none of the material is intact enough to be sure. Except for their ancient beginnings, we do not know how Africa’s cats arose.

The serval (ABOVE LEFT) is thought to be most closely aligned with the caracal (ABOVE RIGHT) lineage but the relationship is a distant one. Molecular analyses suggest the serval has been a discrete species for 10 million years.
Physical adaptations: being a cat
As the most carnivorous of carnivores, cats are supremely well adapted to procuring their meals. Compared to abundant, accessible foods such as insects, grass or leaves, meat is scarce; so cats have to be well equipped to locate their prey without being detected themselves. Once found, the potential meal is usually highly mobile, armed with teeth, horns or antlers to defend itself; and it is often much larger than its hunter. Cats must capture and overpower their prey quickly and efficiently to avoid going hungry or, in the worst scenarios, incurring potentially fatal injuries from formidable prey. The physical challenge of being a cat is answered by a suite of anatomical and morphological specialisations that makes them the most exquisite of stealth hunters.
The framework: skeleton and skull
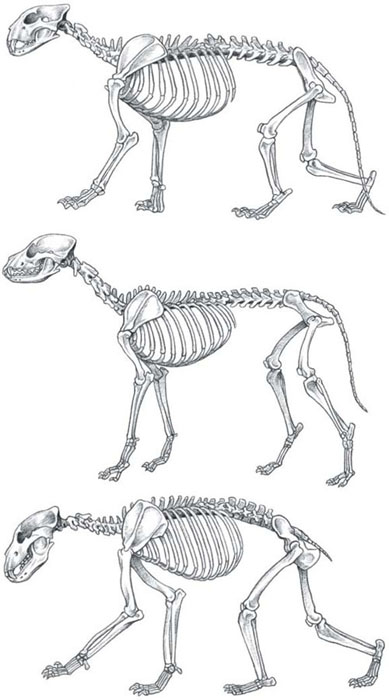
TOP TO BOTTOM: The skeletons of cat, dog and bear. With long limbs, digitigrade stance and a relatively stiff spine, the dog is lightly built for endurance running. The bear’s heavy limbs and robust build reveal a combination of great strength and endurance but poor speed. The cat skeleton is the efficient middle ground, with adaptations for explosive bursts of both speed and strength but lacking in endurance.
Africa’s cats are remarkably varied. They range from the tiny black-footed cat, with a top weight of 2.5 kg, to the lion, 100 times as heavy. They can be tall and slender like the cheetah and serval, small and squat like the sand cat, or athletically robust like the leopard and golden cat. They vary in size, colour, build and even the relative length of their tails; sinuously long and ‘typically’ feline in most species, but oddly truncated in jungle cats, servals and caracals.
In spite of their apparent differences though, all cats are built along very similar lines. Discounting size, the skeletons of cats are surprisingly uniform, with the differences between lion and black-footed cat, or sand cat and cheetah, representing minor variations on a successful theme. Regardless of species, the feline skeleton delivers speed to run down prey and strength to subdue it. However, this presents an immediate biomechanical impasse, as adaptations for speed tend to counter those for strength. The feline design is an efficient compromise between the two, meaning that most cat species tend to be very fast and very strong for short bursts, but lack endurance in either. Cats are built for explosive effort. They cannot run down prey over long distances as can members of the dog family, nor can they spend hours excavating it, as do bears or badgers.
Compared to many mammals, cats have long, slender and relatively light legs, an adaptation for speed; long, lightweight legs translate to longer strides, increasing the ground covered for any given step. Like other cursorial carnivores such as hyaenas and dogs, the main bones of the felid limb are long and the bones of the foot (collectively called metapodials) are greatly elongated. As discussed earlier, all modern cats are digitigrade, which further increases the length of the limbs; to provide stability in this stance, the metapodials are fused together in a rigid assemblage. As one would expect, the cheetah shows the greatest sophistication of these refinements. It is the only cat in which the main bones of the front leg, the humerus and radius, are the same length; a leopard’s radius is about 85 per cent the length of its humerus, furnishing it with considerably greater strength but a shorter stride than the cheetah.
All cats have lost the clavicle (collarbone) or retain it as a vestigial ‘floating’ bone, so that the front limbs hang free in sturdy, muscular slings supported from the spine and ribs, able to pivot unhindered on the scapulae (shoulder blades). This permits a greatly increased range of movement in the front legs, which different cats put to different use; in cursorial species like the cheetah, greater movement enhances the length of the stride, whereas for more arboreal species such as the leopard, it allows greater mobility for vertical movement. The shape of the scapula – elongate in the cheetah, broad and shallow in the leopard – reflects how their different lifestyles put the front limbs to use.

Although similar in size, cheetahs (TOP) and leopards (ABOVE) have strikingly contrasting builds, reflecting adaptations geared towards speed and power respectively. Cheetahs have longer, slimmer legs, a longer tail for balance, and a longer, flexible spine to furnish a speed that exceeds 100 km/h. The leopard’s shorter, less flexible but more powerfully built limbs and spine limit its top speed to around 60 km/h but deliver far greater strength for tackling larger prey and climbing.

The caracal’s stoutly built skull and robust dentition are adaptations for taking prey that is large relative to its size. Unique among African felids, the conspicuous ear tufts are thought to enhance facial expressions used for communicating with other members of the species.

As typified in the robust back of the male lion (TOP), large cats have proportionally shorter and stiffer spines than small cats such as the serval (TOP, ABOVE). The modifications in large cats arise from a shortening and thickening of each vertebra, not from a reduction in their number.
The feline spine displays variation along a similar scale. Small cats have a long, flexible spine, helping to augment their stride length while running on the ground but also increasing their ability to climb and negotiate obstacles vertically. Here the cheetah resembles small cats, with proportionally a longer and more supple spine than any cat larger than a male caracal, though of course it is for speed, not tree climbing, in the cheetah. In this, they differ from all other large cats, which have undergone a gradual shortening and stiffening of the spine from their distant Pseudaelurus roots. This is primarily to support larger body weights but it also serves to yield spectacular strength. The Panthera cats (represented in Africa by the lion and leopard) have the shortest, stoutest backs of all the felids. In the leopard, the spine has retained some flexibility reflecting its more arboreal habits, while the lion’s massively robust spine delivers Herculean leverage for wrestling large prey to the ground.
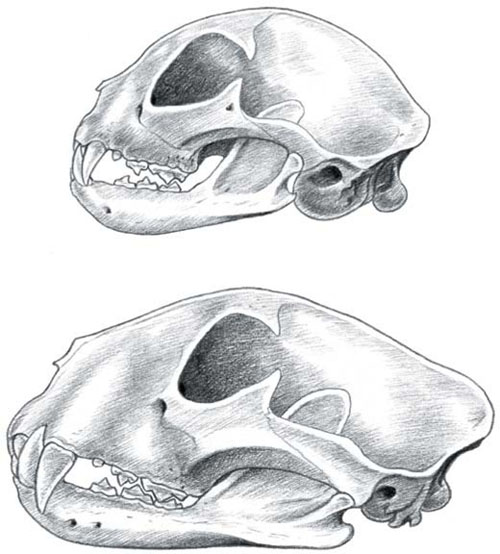
The skulls of serval (TOP) and leopard (ABOVE). Compared to small cats, large species have relatively longer muzzles, smaller orbits (eye sockets), smaller braincases and a well-developed sagittal crest.
As anyone who has seen their pet cat yawn can testify, the skull of the cats carries its own suite of adaptations for killing. Cats have shortened faces compared to most carnivores, a consequence of their reduced tooth count. The shortened mandible (lower jaw) enhances the action of the temporalis muscle which closes the jaw, increasing the force of the killing bite. However, as the size of the cat increases, the relative space on the braincase for anchoring this muscle declines. Proportionally, large-bodied cats have smaller brains than small cats, meaning that, as the jaw increases in size to cope with larger prey, the braincase remains relatively small. To provide space for muscle attachments, large cats have evolved a great flange of bone on top of the skull called the sagittal crest. The larger the cat, the larger the crest and, in fact, it differs between the sexes in the same species. Male leopards have considerably more prominent sagittal crests than females, leading some authors to speculate that males have markedly different diets.
Despite its greater size, the cheetah’s skull resembles that of the small cats, a consequence of its high-speed lifestyle. The skull is relatively small with a foreshortened face and jaw to reduce weight, and a poorly developed sagittal crest. It also has proportionally the smallest canines of all large cats, creating space for enlarged nasal passages that allow the cheetah to inhale great gulps of air through the nose while maintaining a suffocating throat-hold on its prey. The cheetah’s diminutive skull and jaws work well to subdue medium-sized prey with slim necks (as in their main prey, antelopes) but do not have the structure and strength to overcome larger species. With only slightly larger body size, the leopard’s solidly built skull helps it to hold and dispatch prey over four times the size of the top weight manageable by the slight cheetah. A similar relationship exists between the skulls of the caracal and serval. Although similar in size, the serval’s skull is far more lightly built, reflecting a diet dominated by small rodents and birds. The caracal’s heavily constructed skull can deal with prey up to the size of a female impala.

Compared to other large cats, the cheetah’s skull is small, with proportionally shorter jaws and a smaller sagittal crest to reduce weight for the high-speed sprint. Cheetahs also have relatively small canines, creating space for enlarged nasal passages to improve the intake of oxygen while maintaining a suffocating throat-hold on the prey.
We have already seen the importance of the carnassial teeth in the life of cats, but the kill is dependent upon the canines. Compared to the long, elliptical canines of the sabretooths, the broad, conical shape of the canines in living cats reinforces them against the risk of breakage during the deep, forceful killing bite. Most cats kill large prey by a suffocating bite at the throat but occasionally use a stabbing bite to the skull or into the nape, dislocating the cervical vertebrae and piercing the spinal cord. The latter technique is used frequently by small cats killing relatively small prey, and indeed the canines of small cats are well innervated – able to ‘feel’ the gaps between vertebrae to line up for the puncturing bite (this may also be true of large cats but further investigation is required). The clash of canines against bone in such a bite would probably have shattered the elongated canines of the sabre-tooths, but the stout canines of modern cats can usually withstand the impact. Intriguingly, there is a living cat that may be an exception to the rule. The clouded leopard of Asia has proportionally the longest canines of any felid, and possibly represents the nascent emergence of a sabre-toothed form among modern cats. The species is so little known that biologists have yet to discover how it kills, but its canines are long and just flattened enough to suggest that they would risk breakage in any bite that struck bone.
The senses


Light reflected from the tapetum lucidum enhances the nocturnal vision of cats, and also gives rise to ‘eye-shine’. Of all African cats, the nocturnal vision of cheetahs is probably the least developed, a reflection of its mostly diurnal habits. Even so, cheetahs see far more efficiently in darkness than humans, and occasionally hunt when there is sufficient moonlight.
The world of the cat is one of sight and sound. Although felids have a relatively highly developed sense of smell (detailed on page 55), they rely chiefly upon superb vision and hearing to do their hunting. Cats are opportunists and kill at any time of day, but, except for the diurnal cheetah, hunting is mostly a crepuscular (dawn and dusk) or nocturnal activity. To accommodate all possible lighting conditions, cats’ eyes have a sophisticated muscular mechanism for controlling the pupils. Unlike human eyes, where these muscles (called the ciliary muscles) are arranged around the pupil like the spokes of a wheel, allowing limited control, the ciliary muscles of a cat’s eyes are drawn across each other like a braid. The ability is greatest in small cats, which are generally more nocturnal than large species, and are able to reduce their pupils to a characteristically feline vertical slit as protection against intense light during the day. Large cats have the same mechanism but it is somewhat less developed, probably reflecting their more round-the-clock habits and resulting in pupils that are oval shaped.
Once light reaches the eye, cats have an extraordinary ability to utilise it. A reflective sheet of cells called the tapetum lucidum redirects unabsorbed light into the retina for a second chance at imaging (resulting in the characteristic ‘eye-shine’ of nocturnal species). Cats have as many as 15 layers of mirrored cells stacked in their tapetum, reflecting up to 130 times as much light as that bounced around inside the human eye. Beyond the ability to capture light, night vision needs rhodopsin or ‘visual purple’, a light-sensitive pigment assembled from two fundamental building blocks, vitamin A and beta-carotene. However, unlike many mammals, cats cannot synthesise their own vitamin A nor can they access the main source of beta-carotene, plants. Ironically, their prey provides the solution. Herbivores are able to process the compounds and store them in the liver, lungs and fat. In doing so, prey unintentionally equips predator with an accessible source of both, ensuring that cats can see in the dark. Indeed, domestic cats deficient in vitamin A suffer night blindness. No animal can see in total darkness, but domestic cats can see in light one-seventh as dim as the absolute limit of human night vision.
The cat’s visual prowess is rounded off by its ability to judge distance and detail. Feline binocular vision is the most highly developed of all the Carnivora and, among mammals, is second only to that of primates. To improve detail over distance, nerve cells are clustered in the centre of the eye but, rather than as a central spot of improved vision (such as in human eyes), the nerves are arranged in a band. The result is known as a ‘visual streak’, a strip of highly acute vision that increases the ability of cats to discern their prey in the horizontal plane. Among cats that have been tested, the visual streak is most concentrated in the diurnally hunting cheetah, which can spot a moving gazelle on the horizon at two kilometres.

Cats’ ears (CLOCKWISE, FROM TOP LEFT): Caracal, sand cat, lion and serval. Small felids have proportionally larger ears than big cats, reflecting their greater reliance on searching out small, cryptic prey such as rodents hidden in grass. Hair lining the inside leading edge of the ear helps to keep out dust and dirt.
All cats have superb vision but some species rely more heavily than others on their hearing to hunt. Small cats are particularly well adapted to hearing high-pitched sounds, up to two octaves higher than the upper limit detectable by humans. It enables cats to hear with ease the vocalisations of their rodent prey, which typically fall between 20–50 kHz; the limit of human hearing is 20 kHz, so we are deaf to such ultrasonic ‘squeaks’. Sensitivity to high-pitched noise is only half the challenge, though; such sounds are also extremely soft. To overcome this, rodent-hunting cats have very large external ears (called the pinna) to enhance sound gathering. Relative to body size, the serval has the largest pinna of any felid, assisting it to hear the sounds of rodents moving through grass and even detect their ultrasonic calls from subterranean burrows. Desert-dwelling sand cats also have large pinnae to listen for rodents, and have a further refinement that seems to be specific to arid habitats: sand cats have a very enlarged bulla, the bony chamber which houses the fragile ear bones in all mammals. An oversized bulla is thought to reduce air resistance inside the ear, enhancing sensitivity to both high and low frequency sounds in an environment that is spectacularly quiet. Somewhat enlarged bullae are also found in the black-footed cat, a denizen of semi-arid habitats.

Cats’ whiskers (CLOCKWISE, FROM TOP LEFT): lion, leopard, African wildcat and serval. The vibrissae (whiskers) of all cats are well served by nerves and muscles. Cats are able to extend their whiskers forwards to feel their surroundings or for the accurate placement of a killing bite.
Secondary to sight and hearing, cats employ two further senses to survive: touch and smell. The whiskers, or vibrissae, of cats are richly served by nerves, intensifying their sensitivity to stimulation and enabling them to negotiate their world in complete darkness. When moving at night, a cat fans its muzzle whiskers forwards in order to feel its way, assisted by clusters of vibrissae on the eyebrows, cheeks and throat. Cats even have vibrissae on the wrists to help them place their feet when stalking prey without looking down. Although less noticeable than vibrissae, a second type of long, tactile hairs called tylotriches are scattered thinly over the entire body hidden among the ‘ordinary’ fur. Working in unison with hearing, a cat’s full-body tactile precision enables it to hunt in complete darkness. Domestic cats experimentally blindfolded were easily able to negotiate a large room and find, catch and kill mice with a precise bite to the neck.
Of all the senses, smell plays the least part in the cat’s effort to feed itself (but is very important in felid social life, see Chapters 3 and 4). Nonetheless, cats have a sense of smell estimated to be 30 times finer than that of humans. A leopard easily follows the scent trail of a dragged carcass for up to two kilometres and, although the ability has never been fully assessed, cats clearly find some meals by smelling them from afar.