The physiology of the ferret has been extensively described in the comparative biomedical research literature due to the use of ferrets as animal models of human physiology and disease. Ferrets have both similar and unique anatomical and physiological features that offer valuable comparisons to human physiology. The popularity of ferrets as pets has produced numerous clinical reports describing parameters of normal physiology contrasted to common clinical syndromes of heart and endocrine disease, among others. This chapter will provide an overview of normal ferret physiology, and the reader will find additional descriptions of normal physiology in other chapters focusing on normative biology, diseases of specific organ systems, and use of the ferret as an animal model.
Cardiovascular Physiology
The ferret has been used extensively to model cardiovascular physiology, particularly isolated whole heart and muscle strip preparations for in vitro study (see Chapter 27). Cardiac disease in pet ferrets is common, including dilated cardiomyopathy, arrhythmias, endocarditis, heartworm, neoplasia, acquired valvular disease, and congestive heart failure (see Chapter 18). Clinical data on normal heart rate, blood pressure, ECG, and cardiac size as established by thoracic radiography and ultrasound have been published [1–3]. In these studies, gender and body weight were identified as important contributors to variance in cardiovascular parameters. Electrocardiographic Holter monitoring has been described for the ferret [4].
Heart Rate and Blood Pressure
Truex and colleagues reported that the sinoatrial node is discrete (3 × 1 × 1 mm), easily identifiable, and the conduction system well differentiated [5]. Anatomy of the ferret sinoatrial node more closely resembles the human anatomy compared with other models [6]. Heart rate measurements have been reported for ferrets under anesthesia, under simple restraint, and unrestrained [7], acknowledging the impact of various anesthetics on heart physiology and in the case of conscious animals, the relative activity or stress level at the time of measurement [8]. Older studies reported heart rate either under barbiturate or urethane anesthesia, and the data should be interpreted accordingly. Under urethane anesthesia, the heart rate was 387 ± 54 beats/min (bpm) (X ± SD) [7], whereas under barbiturate anesthesia considerably lower values of 230 ± 26 bpm were recorded [9]. In the barbiturate-anesthetized mink, the heart rate was 324 ± 9 bpm with a blood pressure of 198/143 ± 37/26 mm Hg [10]. In conscious ferrets, heart rates of 341 ± 39 bpm and 200–255 bpm have been reported [7,9]. The mean heart rate in conscious ferrets was higher in female ferrets (300 ± 58 bpm) compared with males (260 ± 34 bpm) using “hanging restraint” in clinically normal ferrets (see Fig. 4.1 in the section “Normal Electrocardiogram”) [11].
Fig. 4.1. Restraining a ferret for ECG examination by the “hanging” method.
Blood pressure measurements have been made in both anesthetized and conscious animals. Under barbiturate or urethane anesthesia, mean systolic blood pressure values between 140 and 164 mm Hg have been reported, with diastolic values of 110–125 mm Hg [7,12]. Study of blood pressure in the conscious animal generated mean systolic values of 161 mm Hg in males and 133 mm Hg in females [9]. In both conscious and anesthetized ferrets, individual animals with pressures as high as 190 mm Hg were encountered, but it is not known whether this represents the extreme end of the normal range or ferrets with a pathologic degree of hypertension.
Control of Heart Rate and Blood Pressure
The nervous system, intrinsic cardiovascular factors, and pharmacologic agents influence heart rate and blood pressure. Most studies have been conducted under urethane anesthesia because urethane induces a long-lasting narcosis (8–10 hours), with minimal cardiovascular or respiratory depression [13], and has minimal effect on neurotransmission [14].
Nerves
In the urethane-anesthetized ferret, cervical vagotomy has little effect on the heart rate, indicating that little vagal tone is present. Electrical stimulation of the peripheral cut end of the cervical vagus at voltages sufficient to activate small-diameter myelinated fibers produces a profound frequency-related bradycardia blocked by atropine. At 50-Hz stimulation, cardiac arrest was produced within a few seconds, and with prolonged stimulation (>15 seconds), vagal escape can be demonstrated [7].
The high heart rate in urethane-anesthetized ferrets (387 ± 54 bpm [7]) may be due to a high tonic sympathetic discharge. Compared with the rat, guinea pig, rabbit, and cat, the ferret has the highest levels of atrial norepinephrine (11.11 ± 1.43 µg/g) and dopamine (0.222 ± 0.088 µg/g) [15]. The ferret heart was used to demonstrate toxic effects of exogenous norepinephrine, often clinically elevated in heart failure, on sympathetic nerve terminals. Norepinephrine toxicity was mediated by oxygen free radicals and antioxidants, such as carotene and ascorbic acid, and α-tocopherol can prevent myocardial β-adrenergic receptor downregulation and postreceptor abnormalities [16]. Exogenous norepinephrine at a dose which produced no significant increase in blood pressure or myocyte hypertrophy caused cardiomyocyte apoptosis in intact animals. This effect was associated with an increase in oxidative stress, upregulation of Bax protein and downregulation of Bcl-2 protein and was prevented by antioxidant vitamins [17]. The findings suggest that norepinephrine-induced myocyte apoptosis is mediated by oxidative stress, and that antioxidant vitamins may be beneficial in heart failure in which cardiac norepinephrine release is increased.
Previous studies have shown that nitric oxide (NO) has a significant facilitator effect on the actions of the vagus nerve on several aspects of cardiac function in the ferret and other mammals. Using an n-NOS inhibitor, 1-(2-trifluoromethylphenyl) imidazole (TRIM), the vagally evoked, frequency-dependent bradycardia was significantly attenuated and could be reversed by infusion of l-arginine [18]. These effects could also be demonstrated in guinea pigs but not in rabbits. TRIM also attenuated the increase in A-V conduction time stimulated electrically, irrespective of pacing the heart, suggesting that vagal stimulation directly caused an A-V delay. These studies reinforce the hypothesis that NO from neurons facilitates vagal control of the heart.
Sinoatrial Node Pacemaker
Using fluorescent in situ hybridization (FISH) to detect transcripts of the slow and cardiac forms of troponin I (TnIS and TnIC, respectively), the location of the central region, and presumably the origin of pacemaker activity in the ferret sinoatrial node, was described [6]. Heterogeneous expression of 33 different cation channel transcripts demonstrated the complexity of electrical activity in the node and suggests that ion channel heterogeneity has an important role in pacemaker function.
The reflex control of the circulation in the ferret has seldom been investigated, but the following brief observations are relevant. In anesthetized animals, distension of the biliary system to noxious levels evokes a reflex increase in blood pressure mediated by sympathetic nerves [19]; similar responses have been produced by distension of the duodenum and gastric antrum. It has been suggested that changes in blood pressure may be used to distinguish between innocuous and noxious levels of stimulation of the abdominal viscera [19]. These studies indicate that the urethane-anesthetized ferret may be a useful model for the investigation of visceral pain mechanisms.
The von Bezold–Jarisch reflex, a vagally mediated response induced by cardiopulmonary chemoreceptive afferents, causes bradycardia, and a paradoxical vasodilatation and decreased cardiac contractility in the ferret [20] as it does in other species [21]. The reflex is mediated by cardiopulmonary vagal afferents and cardiac vagal efferents. The von Bezold–Jarisch reflex results in sudden arterial hypotension and can be induced by injection of a variety of chemical stimuli including 5-hydroxytryptamine 3 receptor agonists such as serotonin or 2-methyl 5HT which induces a prompt onset, dose-related bradycardia lasting about 30 seconds [22]. This response, which is blocked by selective 5HT3 receptor antagonists, has been used as a pharmacological screen for the selection of 5HT3 receptor antagonists (see Chapter 31 on emesis research). In contrast to the rat which develops shallow respiration, the bradycardia in ferrets is accompanied by a tachypnea and increased depth of ventilation. Preliminary studies indicate that this reflex is a response to activation of nonvagal thoracic afferents.
Electrical stimulation of abdominal vagal afferents and greater splanchnic afferents in urethane-anesthetized ferrets induces a rise in arterial pressure [23]. Activation of the abdominal vagal afferents at higher frequencies can induce retching, with each retch characterized by a large decrease in intrathoracic pressure which modulates the blood pressure increase. A rise in blood pressure can also be induced by the intragastric irritant copper sulfate [24] and markedly reduced or abolished by hexamethonium and atropine methyl bromide, indicating involvement of the autonomic system. Lesion studies have implicated pathways other than the vagus.
Pharmacologic Agents
In the ferret, acetylcholine (0.1–1 µg/kg IV) and the acetylcholinesterase inhibitor neostigmine (1 mg/kg IV) produce bradycardia and a concomitant fall in blood pressure. Adrenalin (0.5–50 µg/kg IV), norepinephrine (1–20 µg/kg IV), angiotensin II, and vasopressin produce increases in systemic blood pressure [7,25]. Atropine has little effect on blood pressure, but guanethidine (5 mg/kg IV), propranolol (2 mg/kg IV), and phentolamine (2 mg/kg IV) all cause a decrease in blood pressure, indicating the importance of a tonic sympathetic adrenergic drive in the maintenance of blood pressure in the ferret, as in other species. The nicotinic cholinergic receptor antagonist d-tubocurarine produces a biphasic change in blood pressure; at low doses (0.1–1 mg/kg IV) hypotension results whereas higher doses (4 mg/kg IV) produce hypertension. Because ganglionic blockade was not produced by low doses, it has been suggested that the characteristic hypotensive response to d-tubocurarine is caused by the release of vasodilator agents [26]. Detailed pharmacologic investigations of the cardiovascular system have also been undertaken in the mink [10].
Normal Electrocardiogram
The electrocardiogram measured from standard limb leads shows the characteristic P, QRS, and T waves, with the QRS complex recorded at 1.84 ± 0.61 mV in lead II and becoming progressively smaller in leads I and III [7]. Heart rate and the direction, amplitude, and duration of the different ECG waves were evaluated in 43 clinically normal, hand-restrained ferrets with comparison to previous data generated from anesthetized ferrets [11].
The mean heart rate in conscious ferrets was higher in female ferrets (300 ± 58 [sd]) compared with males (260 ± 34 bpm). P waves were mostly positive in Einthoven I, II, III, and in the aVF leads (always positive in II and III), mostly negative in the aVR, and variable in the aVL leads. Typically, a single R wave was only visible during ventricular depolarization using the Einthoven leads and aVF, and in 25% of ferrets, it was preceded by a small Q wave. S waves were observable in a small percentage of ferrets using caudal leads. The QRS-axis pointed levocaudally (40–60°). Both positive and negative T waves occurred in all horizontal leads, but biphasic configuration was not found. The T wave direction was more commonly (but not exclusively) concordant with the QRS-axis in all limb leads. Most ferrets have sinus rhythm, with sinus arrhythmia (defined as more than 10% variations in the RR intervals) detected in 10% of the animals.
Cardiac Size
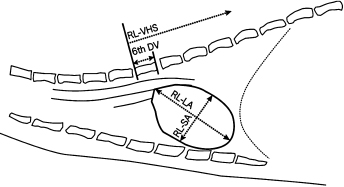
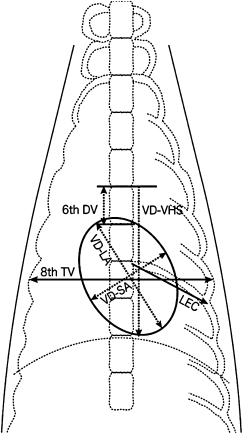
A radiographic survey to determine normal cardiac size used 64 clinically normal male and female ferrets sedated with butorphanol (Table 4.1) [1]. Standard radiography positions (right lateral [RL] and ventrodorsal [VD]) were used to compare ratios between the length of the RL-sixth dorsal vertebra (6th DV), RL- and VD-long axis (LA) and RL- and VD-short axis (SA), RL and VD-vertebral heart size, VD-length of the eighth costal (LEC) and the VD-thoracic width at the eighth thoracic vertebra (8th TV) (Fig. 4.2 and Fig. 4.3). Standard values for ferrets weighing <1 kg and ≥1 kg were established based on the 6th DV and a relational expression obtained by the regression coefficient of the ratio of VD-SA to VD-8th TV such that VD-8th TV = 2.887 + (0.769 × VD-SA). In clinical application, determining the ratio of the 6th DV to other measurements or using the ratio of VD-8th TV to VD-SA both showed similar gender- and body weight-related differences. Given that gender and body weight are correlated in mature adult ferrets, the mean and median for ratios VD-8th TV to VD-SA were 1.94 and 1.85, respectively, in ferrets weighing <1 kg and were 1.97 and 1.89, respectively, in ferrets weighing ≥1 kg. Therefore, the 8th TV measurement should be approximately 1.9-fold greater than the VD-SA. A diagnosis of cardiac hypertrophy is characterized by a lower ratio, and greater values indicate cardiac reduction or dilatation of the thoracic cavity.
Table 4.1. Radiographic Measurements of Cardiac Dimensions for 64 Clinically Normal Ferrets
Fig. 4.2. Right lateral (RL) radiographic view of a normal ferret thorax. Radiographic view shows measurement of the cardiac silhouette. LA, long axis; SA, short axis; 6th DV, sixth dorsal vertebra; VHS, vertebral heart size. With permission from Onuma et al. [1].Fig. 4.3. Ventrodorsal (VD) radiographic view of a normal ferret thorax for measurement of the cardiac silhouette. LA, long axis; SA, short axis; 6th DV, sixth dorsal vertebra; VHS, vertebral heart size; LEC, length of the left eighth costa; 8th TV, thoracic width at the eighth thoracic vertebra. With permission from Onuma et al. [1].
Circulation in Specific Regions
The vasculature and circulation of several specific organs in the ferret have been investigated. In their study of barbiturate-anesthetized ferrets, Kempf and Chang [12] measured a cardiac output of 139 mL/min (range 82–200 mL/min) and circulation times of 6.8 ± 1.2 seconds (fluorescein) and 4.5 ± 0.7 seconds (cyanide). Doppler flow on a limited number of hand-restrained ferrets indicated that the mean of the maximum velocities of blood flow was 1.1 ± 0·2 m/s in the aorta and 1.1 ± 0·3 m/s in the pulmonary trunk. Normal values for two-dimensional, M4-mode, and color flow Doppler echocardiography were reported for 29 clinically healthy ferrets anesthetized with isoflurane [27], and measurements in male and female ferrets were similar.
Heart
The left coronary artery is dominant and supplies the left ventricular myocardium and the nodal regions. The small body size and lack of extensive myocardial collateral circulation supports the use of the ferret to model cardiac ischemia and reperfusion [28–31]. These studies demonstrated that cardiac preconditioning in the ferret has a lower threshold than in rats, rabbits, or dogs. Unlike the dog and pig, the beneficial effects of preconditioning were lost when ischemic intervals were extended to 75 minutes or longer.
Brain
One anatomic feature of the ferret cardiovascular system requires clarification. Several authors have reported a single innominate artery arising from the arch of the aorta, instead of the more usual arrangement of two carotid arteries. In fact, the innominate artery divides into two carotid arteries within the thorax, and these continue on either side of the neck, closely applied to the vagi and internal jugular vein, to supply the brain [7]. The impression given by some authors that the ferret possesses a single direct ascending artery supplying the brain is misleading. It has often been suggested that the presence of an innominate artery would be advantageous in maintaining cerebral perfusion as the animal turns its head while maneuvering in a confined space [32], but it is difficult to envisage this as the innominate artery is located entirely within the thorax.
Testes
The male ferret testicle undergoes stimulation from luteinizing hormone when day length increases in the spring [33]. Seasonal variations in testicular blood flow have been investigated [34], and testicular blood flow is at a minimum in July (21.3 ± 1.5 mL/min/100 g), and increases to a maximum of 35.2 ± 1.2 mL/min/100 g between January and March. Spermatozoa are present in the seminiferous tubules between January and June. These changes are presumably regulated by seasonal fluctuations in levels of reproductive hormones.
Gut
Measurements of systemic blood pressure following administration of drugs into the celiac artery via the abdominal aorta provide insight into some influences on the mesenteric circulation. Vasoactive intestinal polypeptide (VIP) (1 µg/kg) and substance P (100 ng/kg) produce a fall in blood pressure, bradykinin (50 µg/kg) produces a biphasic change, and a delayed increase was noted with high doses of neurotensin (1 µg/kg) [35]. Stable analogues of ATP produce a marked vasoconstriction in the ferret gut in contrast to the vasodilator action usually associated with ATP. Stimulation of the peripheral cut end of the greater splanchnic nerve produces a large increase in blood pressure because of a direct effect on the mesenteric bed and the release of adrenalin from the adrenal glands. Norepinephrine and dopamine have been detected in the mesenteric artery and spleen [15].
Data are not available on the blood flow to each region of the gut, nor on the relative distribution of blood flow between the gut muscle and mucosa.
Pituitary
Direct measurements of the blood flow to the pituitary have not been made, but the pituitary vasculature has been the subject of detailed anatomic studies. The hypothalamic-pituitary portal system has also been investigated because of its involvement in the endocrine functions of the pituitary [36].
Pulmonary Vasculature
It has been suggested that the sensitivity to hypoxia in the ferret may be an adaptation to burrowing [37]. Compared with other species (e.g., rat, rabbit, cat, dog, pig), the ferret has the most marked pulmonary vasoconstriction response to hypoxia, with a maximum response occurring at about 25 mm Hg [38–40]. Unlike lungs from other laboratory animal species, the ferret lung has a stable and reproducible hypoxic response in isolated preparations [41]. In isolated ferret lung preparations, there is a biphasic response to hypoxia; the vasopressor response to anoxia is characterized by an intense initial vasoconstriction, followed by marked vasodilation as the partial pressure of inspired oxygen drops. The mechanism of biphasic response to hypoxia may be explained by a decrease in ATP concentration which opens ion channels resulting in increased potassium ion (K+) efflux, hyperpolarization, and transition of the initial vasoconstrictor response to vasodilation [42]. The extreme sensitivity of the ferret pulmonary vasculature to hypoxia makes it a very useful model for investigating this important component of pulmonary hypertension in humans [43].
Respiratory System
The ferret is used as an animal model for influenza research, particularly with renewed interest in H1N1 as a human pathogen continuing to pose a public health threat [44] (see Chapter 25). Additionally, a ferret model of cystic fibrosis generated by somatic cell nuclear transfer has a phenotype consistent with cystic fibrosis seen in human newborns [45].
A review of the anatomy and physiology of the ferret respiratory system has been published [46]. The ease of making physiological measurements, the comparatively large lung volume in relation to body weight, and the large cross-sectional area of the airways [47], distinguish the ferret as a useful animal model for studies of pulmonary physiology and inhalation toxicology. The ferret has a total lung capacity that exceeds the predicted value by 297%. The only comparable animal is the sea otter, an aquatic mustelid, in which the large lung capacity (462% of predicted) is a probable adaptation for diving [47]. The disproportionately large lungs of the ferret are advantageous for studies involving uptake, clearance, and deposition of aerosols, as well as the mechanics of pulmonary blood flow and diffusion. The anatomy of the left lower lobe of the ferret lung is particularly advantageous for isolated perfusion [41].
The ferret is useful for study of pulmonary pharmacology because the large lung and long trachea allow several preparations to be made from a single animal. This allows the animal to serve as its own control for simultaneous measurement of physiologic responses to varying experimental conditions in vitro. In addition to the large lung capacity, the chest wall of the ferret is very compliant, an apparent adaptation for subterranean hunting [37]. Compared with the dog lung, the ferret has more submucosal glands in the bronchial wall and an additional generation of terminal bronchioles [48], making the ferret lung closer anatomically to the human lung. The ferret's size and long thin body shape requires lengthening of the plethysmograph. Ex vivo, isolated, perfused lung preparations that can remain viable for extended periods are commonly used for metabolic studies and for regional lavage to study markers of lung injury.
Pulmonary Mechanics
The resting respiratory rate in the conscious ferret was reported to be 33 to 36 breaths/min with similar values of 27 to 44 breaths/min in anesthetized animals, with minor variability attributable to the animal size as well as the type and dose of general anesthesia used [7,37,49]. In nine pentobarbital-anesthetized, tracheotomized, supine male ferrets weighing 576 ± 12 g, tidal volume was 6.06 ± 0.030 mL and frequency was 26.7 ± 3.9 breaths per minute, with a resulting minute volume of 157.0 ± 14.8 mL/min. Total lung capacity, vital capacity, and expiratory reserve volume were 89 ± 5, 87 ± 5, and 16 ± 2 mL, respectively. It is notable that the total lung capacity of these 0.6 kg ferrets was similar to that of a much larger, 2.5 kg rabbit. Functional residual capacity of 17.8 ± 2.0 mL was determined by gas freeing the lungs in vivo [47]. Dynamic lung compliance (2.48 ± 0.21 mL/cm H2O) and pulmonary resistance (22.56 ± 1.61 cm H2O/L second) studies have revealed that ferrets have almost infinitely compliant chest walls such that lung and total respiratory system curves are essentially identical. Maximum expiratory flow-volume curves showed peak flows of 10.1 vital capacities (VC)/s at 75% VC and flows of 8.4 and 5.4 VC/s at 50% and 25% VC, respectively.
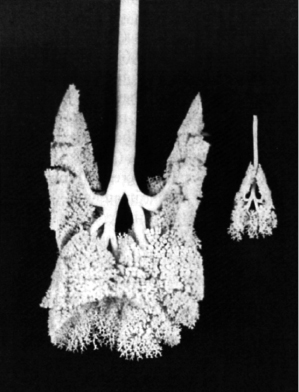
The trachea of the adult male ferret has an average length of 10 cm which is only slightly less than the average of 12 cm in the human [50]. The number of generations of terminal bronchioles in the ferrets (1 to 2) is intermediate between that of the dog (0 to 1) and the human (3 to 4). Like humans, the airways of the young ferret grow in length and diameter in proportion to body length [51] such that body length at a given age is a reliable predictor of airway measurements [52]. Oldham and colleagues [51] reported airway lengths and diameters to be linearly correlated with body length and curvilinearly correlated to age and body weight as a function of body length. They established appropriate mathematical formulas for modeling particle deposition in the ferret lung. These authors also made a first attempt at establishing the lung mass to body weight relationship in ferrets by examining a young male ferret aged 57 days and weighing 599 grams. The lung lobe to body weight ratios calculated for the six anatomically separate lung lobes of the ferret varied from 0.0007 in the smallest lobe, the right cardiac, to 0.003 in the largest lobe, the left apical. Because only one animal was examined, it is not known if the lung-to-body weight ratio remains constant as the animal grows. Silicone rubber casts of the tracheobronchial tree were made to establish the average number of generations or pathways between the trachea and the terminal bronchioles using growing ferrets varying in age from 14 hours to 56 days (Fig. 4.4). The average path ended in a terminal bronchiole within 17 generations. There was insufficient data to establish if the number of nonalveolarized airways increases or decreases between birth and maturity.
Fig. 4.4. The young adult and newborn ferret tracheobronchial casts. The silicone rubber casts were prepared in the thorax to maintain anatomical accuracy. Reprinted with permission from Laboratory Animal Science [51].
The ferret lung functionally and anatomically adapts to changing atmospheric pressure. When exposed to atmospheric conditions creating hypoxia, ferrets have intense pulmonary vasoconstriction, similar to that of humans who develop high altitude pulmonary edema (see previous discussion under “Pulmonary Vasculature”). Using isolated ferret lungs, this vasoconstrictive response was shown to be regulated by ATP-dependent K+ channels [53]. In a study examining the effect of positive airway pressure on postnatal lung growth and capacity, young ferrets exposed to a 6 cm of water increase in atmospheric pressure experienced as much as a 40% increase in lung volume with significant remodeling of the lung parenchyma [54]. Ferrets that underwent right-sided pneumonectomy at 8 weeks of age experienced a 65% increase in left lung volume by maturity [55]. Central airway cross-sectional areas at multiple locations were 12% larger in the postpneumonectomy animals compared with controls. Central and peripheral conducting airways had an increase in cross-sectional area to similar degrees, but this airway growth was less than the compensatory increase in lung volume.
Only limited studies have examined pulmonary mechanics in the ferret and clearly, additional studies are needed.
Airway Secretion
Studies have focused on the mechanisms and control of tracheal secretion in the ferret because it yields a large sample size and contains a large number of submucosal glands. Submucosal glands in the ferret are estimated to occupy a much larger proportion of their bronchial wall than dogs (1%) and are more similar to humans (11%) [56]. Histologic and histochemical studies of the ferret tracheobronchial tree indicate that the epithelium is composed of ciliated cells, nonciliated dark cytoplasmic cells, with or without secretory granules, and a few goblet cells [48,57]. The number of goblet cells increases toward the bronchus where approximately 60 cells/mm can be found, many of which contain acidic material, possibly glycosaminoglycans. Glands containing acidic material are more numerous in the distal airways. Surface epithelial cells in postnatal ferret trachea have been shown to be involved in proteoglycan secretion [58].
Various techniques have been used for studying tracheal secretion in the ferret, including the in vivo construction of tracheal pouches [59], in vitro collection of secretions from sectioned tracheas [60], studies of ion fluxes from tracheal segments mounted in modified Ussing chambers [61], and mucus secretion from individual tracheal glands with time-lapse optical imaging of mucous droplets under an oil layer [62]. The following results are from in vitro studies.
Fluid Secretion and Ion Fluxes
Because of the large number of submucosal glands, it is probable that fluid collected from the respiratory tract is primarily glandular in origin, a hypothesis supported by functional studies. The volume of fluid appearing on the luminal surface of the trachea depends on the competing rates of fluid secretion and fluid absorption. Under resting conditions, little if any spontaneous secretion is present [61]. Under basal conditions, there is a net reabsorption of fluid, which is consistent with ion flux studies demonstrating a net absorption of sodium with little or no active chloride secretion [63]. This transepithelial short circuit current was reduced to zero by ouabain (a cardiac glycoside), reduced by amiloride (a potassium-sparing diuretic), and unaffected by bumetanide (a loop diuretic).
Nerve stimulation, phenylephrine, and acetylcholine enhance fluid secretion, and it has been proposed that the autonomic innervation of the submucosal glands regulates fluid secretion via α-adrenergic and muscarinic cholinergic receptors. Ion flux studies demonstrate stimulation of chloride secretion by isoproterenol and stimulation of sodium and chloride ion secretion by methacholine [61]. Like other tissues, the movement of these ions probably provides the mechanism by which the tracheal fluid is secreted; however, involvement of macromolecules cannot be excluded [64,65]. Using the whole trachea, in vitro basal secretory rates of 0.22 ± 0.03 µL/min were obtained; stimulation with methacholine increased the rate to 3.9 ± 0.33 µL/min. An increased stimulation of secretion was also produced in this preparation by phenylephrine and histamine, while decreased secretion was produced by salbutamol and substance P [66]. Substance P has been shown to be a potent stimulator of airway mucus secretion in isolated canine, ferret, and human airways [67–69]. Substance P can cause a dose-dependent decrease in mucus viscoelasticity and dose-dependent increase in mucus cleavability. Although humans, dogs, and ferrets differ in airway size, and experiments differ by type of anesthesia used and collection times, most measurements of the viscoelastic and transport properties of mucus are within one standard deviation of each other, indicating the usefulness of the ferret model for clinical studies of mucus rheology.
Secretion of Macromolecules
Using 35SO4 as a marker, electrical field stimulation, acetylcholine, phenylephrine, terbutaline, and norepinephrine produced an increased in vitro secretion of macromolecules of more than 106 molecular weight, suggesting that secretion of macromolecules in respiratory secretions is regulated by tracheal innervation, muscarinic cholinergic receptors, and by both α- and β-adrenergic receptors [70]. A non-noradrenergic, noncholinergic (NANC) component has also been identified. Because VIP enhances the output of sulfated macromolecules in the ferret trachea, it may be responsible for the NANC component of the neural response [71]. VIP does not, however, appear to directly stimulate ion transport in this tissue.
VIP has been implicated in the regulation of mucus secretion in the respiratory tract [65], a process involving release of macromolecules from exocrine cells and transport of ions and water across the airway mucosa. However, because airway glands and mucosa both consist of mixed cell populations, it has been unclear which specific cells contained VIP receptors and mediated the VIP-evoked responses. The responding cell type was identified using immunocytochemistry to monitor concentration changes in adenosine 3′,5′-cyclic monophosphate (cAMP), the intracellular second messenger believed to mediate VIP responses. Both serous and mucous cells of ferret tracheal submucosal glands and ciliated and basal cells of dog tracheal mucosa increased in cAMP content in response to VIP stimulation, suggesting that these cell types possess VIP receptors and thus participate in VIP-stimulated responses [65]. In contrast, ferret tracheal epithelium and dog epithelial goblet cells had little or no reactivity after VIP exposure, suggesting that these cells lack VIP receptors.
Immunohistochemical studies have localized lysozyme in the secretory granules of serous but not mucous cells in the ferret trachea [72]. Thus, by monitoring lysozyme secretion, factors influencing the control of serous cell secretion can be studied. It has been established that the total secretion volume and rate of lysozyme release are independent of each other. Inflammatory mediators such as PGE1 decrease methacholine-stimulated lysozyme release by 70% without decreasing the total secretion volume [73]. Lysozyme release in incubated tracheal segments is stimulated by cholinergic and α-adrenergic receptor agonists, whereas β-adrenergic agonists have only a small effect [72]. Autoradiographic studies have demonstrated the presence of α1- and β-adrenergic receptors on both epithelial and submucosal gland cells [74,75]. A morphometric study demonstrated that serous cell granules are only discharged by α-adrenergic and cholinergic stimuli, an observation consistent with the results of the aforementioned studies on lysozyme release [76].
In vitro studies using ferret respiratory tissue have contributed to delineating the role of various mediators of respiratory tract inflammation and the pulmonary response to irritants and carcinogens. Tachykinins, including substance P, are contained in the sensory nerves of ferret airways [77]. Sensory nerve stimulation causes release of the tachykinins, thus producing a pattern of responses (smooth muscle contraction, submucosal gland secretion, increased vascular permeability, neutrophil adhesion, and cough) collectively referred to as “neurogenic inflammation.” The responses to either exogenously or endogenously released tachykinins are modulated selectively by neutral endopeptidase (NEP), an enzyme that exists on the membranes of cells that contain tachykinin receptors (e.g., submucosal glands, smooth muscle, postcapillary venous endothelium). By cleaving and thus inactivating the tachykinins, NEP limits their action on receptors. The reduced NEP activity associated with respiratory viral infections and inhaled irritants (e.g., toluene diisocyanate, cigarette smoke) potentiates neurogenic inflammatory responses. Exogenously delivered human recombinant NEP reduces responses to tachykinins. Thus, reduced NEP activity in tissues, by exaggerating inflammatory responses resulting from sensory nerve stimulation, may play an important role in the pathogenesis of inflammatory diseases in airways and in other tissues.
Platelet activating factor (PAF), a mediator of anaphylaxis, stimulates secretion of mucin by ferret tracheal explants in organ culture. Enhanced secretion is believed to result from PAF-stimulating peptidyl leukotriene biosynthesis within the epithelial cells of the respiratory mucosa [78]. This was supported by demonstration that exogenous arachidonic acid increased cholinergic secretory responsiveness of ferret tracheal glands [79]. PAF also induces relaxation of pulmonary arteries but contraction of pulmonary veins in the ferret. This vasoactivity is dependent on the presence of endothelium and does not appear to be mediated by metabolites of arachidonic acid or by endothelin [80].
Proton magnetic resonance imaging (MRI), used in conjunction with the intravascular contrast agent albumin-(Gd-DTPA), has been used to show that histamine causes an increase in vascular fluids in the nasal turbinates while methacholine (a nonspecific, muscarinic receptor agonist) does not [81]. These pharmacologic agents are associated with the asthmatic state. MRI in the ferret represents a new means to investigate airway pharmacology and the pathophysiology associated with various pharmacological agents, allergens, or viral infections.
Bradykinin and related kinins have been proposed as important mediators of rhinorrhea, and the ferret has been useful in modeling bradykinin binding site distribution and secretory responses in tissues maintained in Ussing chambers [82]. Influenza A infection of the ferret stimulates increased kinin levels in nasal secretions in correlation with the severity of clinical signs, while there is a notable absence of changes in histamine levels in nasal lavage fluid [83].
Good tissue viability of explanted ferret respiratory epithelium maintained in vitro has allowed for examination of the toxic effects of various compounds on the respiratory system. Using this method, the effect of nicotine on ciliary function in the ferret can be studied in the absence of effects related to the presence of goblet cells or submucosal glands. Using the ferret, it was shown that nicotine increases ciliary beat frequency through a direct effect on respiratory cilia [84]. Oxidant air pollutants exacerbate pulmonary disease via airway smooth muscle hyperresponsiveness and potentially, by affecting airway secretion. Chronic exposure to NO2 in the developing ferret lung results in significant changes in alveolar dimensions with thickened alveolar walls, increased cellularity, and collagen deposition, all resulting in increased lung size [85]. The appearance of lesions indicative of oxidant damage even at low concentrations of NO2 during lung development suggests this pollutant may have adverse consequences for adult lung function. Experiments examining the effects of in vivo ozone exposure on the in vitro secretory responsiveness of ferret tracheal glands suggest that oxidant injury not only increases basal secretion of respiratory glycoconjugates but also increases tracheal gland sensitivity to carbachol, a cholinergic agonist. Tracheal gland responsiveness to α- or β-adrenergic agonists is not changed by oxidant exposure [86].
Airway Smooth Muscle Activity
Most examinations of smooth muscle function in the ferret airway have been performed using tracheal smooth muscle. The cells had a resting membrane potential of 58.31 ± 2.10 mV and an input resistance of 21.3 ± 5.31 mV [87]. Control of the tracheal smooth muscle has been investigated in two different preparations, discussed later.
In vitro increases in luminal pressure have been produced in the whole trachea by methacholine, phenylephrine, histamine, bradykinin, and kallidin histamine [66]. The last two substances have no effect on fluid secretion, whereas the remainder stimulate secretion. Autoradiographic studies have revealed a differential distribution of α-1 and β-adrenergic receptors and muscarinic cholinergic receptors: α-receptors are numerous in small bronchioles but sparse in large airways; β receptors are present in the highest density in the bronchioles, although their density is relatively high throughout the airway; and cholinergic receptors are most dense in the bronchial muscle and decrease in density toward the distal bronchioles [74,75,88].
Contraction of tracheal smooth muscle in the ferret appears to be cholinergically mediated through stimulation of only the nonmyelinated efferent axons; stimulation of myelinated axons had no effect [89]. After administration of atropine and elevation of trachealis muscle tone with 5-hydroxytryptamine, stimulation of the vagosympathetic trunk produced a decrease in tracheal pressure. These inhibitory responses were blocked by bretylium tosylate. Because contraction of tracheal smooth muscle in the ferret appears solely dependent on activity in nonmyelinated vagal efferent axons and norepinephrine released from sympathetic nerve endings mediates inhibition, a non-noradrenergic inhibitory system appears to be absent in the ferret.
Studies using ferret trachealis muscle have demonstrated that stretch of the muscle sarcolemma induces substance P release from capsaicin-sensitive C fibers [90]. Substance P released from C fibers influences smooth muscle ionic mechanisms controlling the time for reversal of stretch-induced membrane depolarization and shortening. The ferret was the first animal model used to demonstrate stretch-induced tachykinin release in the airways, supporting the hypothesis that tachykinin-containing primary afferent neurons have local effector functions mediated by tachykinin release [91,92]. Local effects of tachykinins may be important for bronchial reflexes such as the cough response, bronchial muscle contraction, as well as inflammatory responses including changes in capillary permeability.
Ferret trachea has a strong contractile response to bradykinin, a potent bronchoconstrictor, and unlike the guinea pig trachea, has no appreciable basal tone which can complicate measurement of responses to agonists [93]. Contractions induced by bradykinin are potentiated by inhibitors of neutral endopeptidases and angiotensin-converting enzyme, indicating that peptidase action may degrade peptides as a mechanism of modulating tissue sensitivity. The effect of bradykinin to induce contraction of ferret trachea is mediated by a direct action on airway smooth muscle B2 receptors in contrast to the guinea pig trachea where response to bradykinin is markedly attenuated by cyclooxygenase inhibitors [94].
Contraction of smooth muscle in ferret lung lobes responds in a concentration-dependent manner to methacholine and histamine [95]. In this study, the 50% effective doses of methacholine and histamine were 6.41 × 10−6 M ± 1.38 × 10−6 (SEM) and 2.39 × 10−6 M ± 0.53 × 10−6 (SEM), respectively. The maximum level of bronchoconstriction developed in response to methacholine (2.42 mm Hg/mL/min ± 0.28 SEM) was six times greater than that found for histamine (0.42 mm Hg/mL/min ± 0.05 SEM). Responses to both agonists were less pronounced in the ferret lung preparation than those in a similar lung preparation of guinea pig. Compliance changes in both animals were also evaluated. The ferret did not demonstrate a compliance change in response to histamine as was seen for methacholine, suggesting that resistance changes precede compliance changes, or that the ferret airways are particularly resistant to histamine.
Evidence suggests that non-noradrenergic airway relaxation may be controlled by VIP [96]. The magnitude and duration of smooth muscle relaxation in response to VIP may be influenced by rates of peptide degradation after release from efferent peptidergic neurons. The mast cell proteases tryptase and chymase counteract the smooth muscle relaxant effects of VIP in ferret trachea and suggest a potential role for mast cell proteases in the modulation of non-noradrenergic neural control of airway tone by VIP.
Pituitary adenylate cyclase-activating peptide (PACAP) is a VIP-like peptide implicated in the regulation of smooth muscle tone and glandular secretion [97]. In the ferret as well as in several other species, immunostaining of nerve fibers for PACAP identified the distribution of PACAP containing fibers in the tracheobronchial wall distributed around blood vessels and seromucous glands and among bundles of smooth muscle. In the lungs, the immunoreactive fibers were observed close to small bronchioli.
Respiratory Reflexes
Historically, defensive respiratory reflexes have been studied in the pentobarbital-anesthetized ferret. Under anesthesia, coughing is evoked by mechanical stimulation of the bronchi and larynx, but not the trachea. Sulfur dioxide also elicits coughing, but this is weaker than that evoked by mechanical stimulation. The absence of a cough reflex in response to tracheal stimulation under anesthesia may be related either to the depressant effects of the anesthetic (pentobarbital) or to the relative paucity of epithelial nerves in the trachea [48]. In the conscious, free-breathing animal, a tracheal cough reflex secondary to mechanical irritation is apparent. Ferrets have been used as a nonrodent model to develop a method for repeated administration of a liquid surfactant suspension into the lung for the purpose of safety testing [98]. After tracheal catheters were surgically implanted into juvenile ferrets, as much as 12 mL/kg of liquid was administered via the tracheal catheters on multiple occasions for maximum period of 1 month, with only 3% mortality. Although almost all ferrets had an occasional nonproductive reflex cough due to tracheal stimulation by the catheter, they continued to gain weight, play, and otherwise display normal behavior.
Other reflexes that have been studied include aspiration, expiration, and the sneeze reflex [99]. The deflation reflex is present in the urethane-anesthetized ferret and persists in the presence of vagotomy [100]. The reflex respiratory changes accompanying retching and vomiting are discussed in Chapter 31. Further studies of the reflex control of respiration are required, particularly reflex control of airway secretion.
Eustachian Tube Function
The ferret eustachian tube functions as a small-scale model of the eustachian tube of the rhesus monkey and human [101]. The ferret eustachian tube is more similar anatomically and functionally to the human eustachian tube than that of the chinchilla, an established model for studies of the auditory apparatus. The ferret equilibrates less pressure per swallow than humans at equal applied middle ear pressures due to the smaller luminal radius of the eustachian tube. Because ferrets are highly susceptible to infection with human influenza A with resulting disease expression similar to humans [83] (see Chapter 25), the ferret has been an important model for examining the pathophysiologic effects of upper respiratory infections on eustachian tube function. The ferret develops negative intratympanic pressures following influenza infection [101].
Gastrointestinal Tract
The gross morphology of the gastrointestinal tract has been described [102] and is detailed in Chapter 2. Physiologic studies of the ferret gastrointestinal tract were undertaken as early as 1904 when muscle activity of the ferret colon was described [103]. As with other aspects of its physiology, the gut has been investigated largely to determine the potential of the ferret for use as an alternative carnivore to the cat and dog in biomedical research. The majority of studies on ferret gastrointestinal physiology have concentrated on function.
Salivary Glands
The gross and microscopic anatomy of the major and minor salivary glands has been described [104]. Ferrets possess parotid, submandibular, sublingual, molar, zygomatic, and lingual glands. Because ferrets tend to ingest their food quickly, it is unlikely that salivary enzymes play a significant role in digestion, as evidenced by demonstration that parotid and submandibular saliva lack amylase activity [105]. For the ferret, it is probable that the lubricant function of saliva is most important, particularly when the animals are fed a pelleted diet in the laboratory or as pets. No sexual dimorphism in gland morphology, as noted in other species such as the mouse, has been observed in any of the glands examined from male and female ferrets.
The mucosubstances of the major salivary glands of the ferret have been analyzed using different histochemical reactions [106]. Almost all the parotid acinar cells had sialic acid-containing mucus with only a minority of cells staining positive for sulfated mucins. In contrast, submandibular gland mucus was sulfated in the major parts of the gland with occasional cells staining positive for carboxylated mucin. The sublingual and the molar glands had tubules which had sulfated mucins in the central cells and carboxylated mucins at the periphery. The mucus in the tubules of the zygomatic gland was predominantly sulfated. In the sublingual, molar, as well as in the zygomatic gland, there were granulated cells scattered among the tubules. Some of these granules were found to contain carboxylated mucin while others contained neutral mucins.
Although salivary secretion has been most studied in the parotid and submandibular glands, the submucosal zygomatic and molar glands are particularly large in the ferret. A study comparing acetylcholine synthesis in ferret salivary glands found levels three to four times higher in the mucous-type glands (sublingual, zygomatic, and molar glands) compared with the serous parotid and seromucous submandibular glands [107]. Choline acetyltransferase was a critical enzyme, as an inhibitor of acetylcholine synthesis (bromoacetylcholine) reduced synthesis by 95–99% in all five types of glands. Robust production of acetylcholine by the zygomatic and parotid glands was stimulated by the buccal and auriculotemporal nerves, respectively, and parasympathetic denervation reduced the acetylcholine synthesis by 95–96%, respectively. Zygomatic saliva is very viscous, and the flow rate in response to electrical stimulation (20 Hz) of the buccal nerve (zygomatic gland), expressed per gland weight, was one-third of the measurement from stimulation of the auriculotemporal nerve (parotid gland) or the chorda-lingual nerve (submandibular gland). Approximately 25% of the parasympathetic secretory response of the zygomatic gland depended on nonadrenergic, noncholinergic transmission mechanisms, likely mediated by substance P and VIP and possibly calcitonin gene-related peptide. VIP and substance P were found in high concentrations in the sublingual and molar glands (VIP), and in the submandibular, zygomatic, and molar glands (substance P) notably, the concentration of calcitonin gene-related peptide of the sublingual gland was not detectable. Five muscarinic receptor subtypes were detected in all ferret salivary tissues by immunoblotting with similarly high levels of M3, low M2, and mid-range levels of M1, M4, and M5, with serous glands (parotid and submandibular) having more M5 receptors than the mucin-secreting glands. This detailed account of the ferret salivary glands supports that ferrets could be used to study pharmacologic interventions into human salivary disorders, such as xerostomia (dry mouth).
The final saliva in the ferret is usually hypotonic relative to plasma, and the role of the duct system in the modification of the primary secretion has therefore been investigated [108,109]. In both the submandibular and parotid glands, the duct system is impermeable to water both in the resting and stimulated states; thus, changes in the composition of the primary secretion must result from transductal movement of ions. The ducts involved are striated, and they bring about a net reabsorption of sodium and chloride (lumen to blood) and a net secretion of potassium (blood to lumen) in both glands, although the sodium and chloride fluxes are larger in the submandibular gland. In the parotid gland there is some reabsorption of bicarbonate ions.
During stimulation of secretion with pilocarpine (a parasympatholytic alkaloid), net transductal fluxes from the parotid and submandibular salivary gland of Ca++, Na+, K+, Cl–, and HCO3– [108,109] have been characterized by analysis of ductal fluid samples obtained simultaneously from the intercalated, lobular, and main ducts of these glands using micropuncture and microanalytical, as well as standard, physiological techniques. These studies have revealed an interesting difference between the parotid and submandibular glands. In the unstimulated state, both glands produce a primary secretion in their secretory segment (acinus plus intercalated duct) that is isotonic with plasma and has a similar ionic composition in regard to levels of Na+, K+, Cl−, and HCO3− [108,109]. When stimulated by pilocarpine (100 µg/kg IV), which evokes secretory rates of up to 600 µL/min/g wet weight gland, the parotid gland secretes a hypertonic fluid, whereas the submandibular gland secretion remains isotonic. The increase in osmolarity of the parotid fluid is mainly due to Na+ and HCO3− ions. Although the submandibular fluid remained isotonic, its composition changed: HCO3− decreased from 24.2 ± 3.8 to 11.5 ± 3.3 mEq/L, whereas Cl− increased from 106.1 ± 6.5 to 134.5 ± 6.2 mEq/L. These observations suggest that a chloride–bicarbonate exchange mechanism may be involved, as suggested in other salivary glands. The production of an isotonic primary secretory fluid by one gland and a hypertonic one by another in the same animal may be unique to the ferret; hence, this animal could provide a very useful model for investigating the factors influencing each type of secretion.
Most calcium in the saliva from the parotid gland is nonionized, and there is little variation in nonionized calcium concentration when changes in salivary flow between 17 and 363 µL/min/g wet weight tissue are evoked by pilocarpine (2 mg/kg IV) [110]. In contrast, the ionized calcium concentration is related to flow rate, suggesting that some ionized calcium from the primary secretion is reabsorbed in the duct system.
Less is known about the control of secretion. In most species, including the ferret, the major salivary glands are supplied with parasympathetic (cholinergic) nerve fibers, which when stimulated evoke a copious flow of saliva. The salivary secretion in response to stimulation of the parasympathetic innervation is not always completely abolished by the muscarinic receptor blocker, atropine [111]. The extent of the sympathetic (adrenergic) innervation varies considerably not only between different species but also between the glands of the same species. Sympathetic stimulation produces either a small flow of protein-rich saliva or no flow at all.
Electrical stimulation of the parasympathetic auriculotemporal nerve in the pentobarbital-anesthetized ferret provokes secretion (190 mg/min) in the parotid gland [112]. This secretion is markedly reduced (25 mg/min) but not abolished by atropine (2 mg/kg IV). Secretion is also stimulated by methacholine (0.1–2 µg/kg) and substance P (0.05–0.1 µg/kg), but not by adrenalin (20 µg/kg), phenylephrine (100 µg/kg), isoprenaline (100 µg/kg), or VIP (100 µg/kg). These studies demonstrate a potent cholinergic stimulation of gland secretion and, in addition, provide evidence for a noncholinergic stimulation of secretion, similar to that observed in other species. This finding has attracted considerable interest because of the possible involvement of cotransmission.
Electrical stimulation of the sympathetic innervation evokes secretion of submandibular and parotid saliva with higher volumes and protein concentration measured in saliva from the submandibular gland [113]. Fluid responses to sympathetic nerve stimulation are about 1–2% of those occurring with parasympathetic stimulation [114]. Extracts of submandibular and parotid saliva lack amylase activity which may be due to the absence of true serous cells in these salivary glands of the ferret [104]. Lack of amylase is also observed in dogs and cats, but this enzyme is present in the saliva of rabbits and rats [115]. Secretory cells of both gland types are supplied with α(α1)-adrenoceptors and β-adrenoceptors as are the salivary glands of other species [113]. In the submandibular gland of the ferret, α(α1)-adrenoceptors are predominantly involved in fluid secretion and β-adrenoceptors are predominantly involved in protein secretion. In parotid glands, fluid secretion appears to be solely dependent on α(α1)-adrenoceptors, while β-adrenoceptors are almost solely involved in protein secretion.
Secretion of saliva in the ferret may be elicited not only by cholinergic or adrenergic agonists but also by peptides, injected directly into the bloodstream. Intravenous administration of the tachykinins, substance P, and neurokinin A, promote salivary secretion from the ferret parotid and submandibular glands, suggesting that the neuronal release of tachykinins is probably important in the nerve-evoked secretory response of these salivary glands [105]. Tachykinin antagonists can reduce the parasympathetic nerve-evoked secretion of parotid and submandibular saliva by 15–20% and 35–40%, respectively.
PACAP, a VIP-like neuropeptide, was found to be present in neurons in the submandibular gland of the ferret, where PACAP-immunoreactive nerve fibers were distributed around blood vessels, acini, and ducts [116]. Most of the PACAP-immunoreactive fibers were distinct from those storing VIP, suggesting different roles for the two peptides in gland function. PACAP exerted vasodilator effects on the submandibular vasculature which were independent of muscarinic receptor activation and which resulted in a greater decrease in vascular resistance than administration of VIP. While neither VIP nor PACAP alone evoke any flow of saliva, both enhance the flow of saliva as well as the output of protein in response to parasympathetic nerve stimulation, with VIP more potent than PACAP. Similar stimulatory effects occurred in vitro despite adrenoceptor and muscarinic blockade and degeneration of sympathetic nerves. Supersensitivity to both VIP and PACAP resulted after denervation of both the sympathetic and parasympathetic systems.
Esophagus
The entire supradiaphragmatic portion of the esophagus is composed of striated muscle, a pattern found in various other species, including the dog [117]. In humans, the esophagus is composed of striated muscle in the upper portion and smooth muscle in the lower portion, and in this respect, the ferret is not an ideal model. The esophageal lining is composed of keratinized stratified squamous epithelium, which is patchy in the lower portion and absent in the subdiaphragmatic continuation [102]. Submucosal glands are present, but their secretion and control have not been characterized.
The anatomy and functional properties of the sphincters at either end of the esophagus in the ferret are of interest due to use of the ferret in emesis and reflux research (see Chapter 31). Pharmacologic effects and vagal/sympathetic influences on the lower esophageal sphincter have been examined [118,119]. NO was implicated as an inhibitory transmitter in modulating vagus-mediated contractions, and VIP was proposed to be the transmitter responsible for sustained relaxation, as is the case for the adjacent gastric corpus (see later discussion). Sympathetic stimulation had a β-adrenergic inhibitory effect on the sphincter but tachykinins released from axon collaterals were also implicated. Vagal sensory and motor neurons innervate the crural diaphragm and phrenoesophageal ligament. The crural diaphragm vagal afferents show mechanosensitivity to distortion of the gastroesophageal junction, while vagal motor neurons innervate both the crural diaphragm and distal esophagus and likely exert motor control, preventing reflux [120]. Surgical studies in the ferret have demonstrated that it is possible to perform a “Nissen” type fundoplication with animals maintained for up to 6 months postoperatively. This model may help to understand how surgical intervention is of benefit in the treatment of gastroesophageal reflux disease (see Chapter 31).
It was proposed that in contrast to adult humans, the epiglottis of other mammals and infant humans is situated close to the soft palate and that this posture is maintained during swallowing, with food passing laterally around an intact airway. The movement of the epiglottis during swallowing was studied in ferrets and pigs by placing radiopaque markers on the epiglottis and soft palate and using videofluoroscopy to monitor the animals while they fed on hard and soft foods, liquids, and food mixed with barium sulfate [121]. Analysis of the images showed bolus formation and downward movement of the epiglottis away from the soft palate for all tested foods in both species. The duration of the epiglottic movement was approximately 0.3 second for liquids and slightly longer for solids. Because swallowing never occurred past an upright epiglottis, the results of this study do not support the hypothesis that adult animals maintain a patent airway during swallowing. Instead, the epiglottis in ferrets and swine, and presumably in other nonhuman mammals, downfolds similarly to that of adult humans during swallowing.
Preliminary studies of the esophagus have shown that its intrathoracic portion is innervated by the vagus [122]. A single vagal stimulus produces twitch-like contractions of the esophagus and, with increasing frequencies of stimulation (>25 Hz), these contractions fuse to produce a sustained tetanic contraction. The vagal efferent fibers involved are small myelinated axons with a conduction velocity of <20 m/s. The contractions are blocked by tubocurarine, indicating that they are mediated by activation of nicotinic cholinergic receptors on the muscle; this is also the case in the dog. In urethane-anesthetized ferrets, basal lower esophageal sphincter pressure increased after bilateral vagotomy, implicating control of sphincter relaxation by vagal tone [123]. This was demonstrated when peripheral vagal nerve stimulation resulted in sphincter relaxation, often followed by a brief contraction and subsequent prolonged inhibition of sphincter pressure.
Megaesophagus is an idiopathic clinical syndrome in adult ferrets that shares some features with myasthenia gravis. Animals present with a history of regurgitation, difficulty in swallowing, partial anorexia, ptyalism, and lethargy [124]. Cachexia, dehydration, and weakness develop secondarily. Symptomatic treatment is only palliative, with death usually resulting from bronchopneumonia and hepatic lipidosis. Additional information is contained in Chapter 16.
Stomach
Natural infection of the ferret stomach with Helicobacter mustelae as a model for studying disease mechanisms and strategies for prevention and therapy of Helicobacter pylori-associated diseases in humans has firmly established the ferret's value in comparative gastroenterology and pathology [125,126]. The ferret stomach, both in the fasted and fed states, is similar to that of humans in gross morphologic appearance and function. As in other carnivores, the stomach has a considerable storage capacity, as illustrated by the observation that adult ferrets can drink over 100 mL of milk in 10 minutes. Following a meal, about 80% of the food is stored in the proximal stomach, with the circular muscle layer undergoing the largest change in length [127].
The gastric mucosa has been studied both at light and electron microscopic levels in normal ferrets [128], ferrets naturally and experimentally infected with H. mustelae [125,126,129], and those treated with caffeine [130] or cinchophen [131]. The histochemistry of the glands has also been investigated [132]. A detailed description of the mucosal histology is beyond the scope of this chapter, but the main conclusion relevant to the physiology of gastric secretion is that the structure of the mucosa at the cellular level is remarkably similar to that of humans [125].
It is apparent that the ferret stomach is anatomically similar to that of humans, as evidenced by the physiologic investigations described in the following.
Gastric Secretion
The gastric secretions of both hydrochloric acid and proteolytic enzymes have been studied, although the control of the former has been studied more extensively.
Hydrochloric Acid
The study of gastric secretion in the ferret was greatly enhanced by the surgical preparation of ferrets with chronic gastric fistulae [133]. In the fasting state, the ferret is a basal secretor of hydrochloric acid with a fasting pH that ranges from 1.5 to 3.5 [134]. The secretory volume has been reported to be 0.72 ± 0.088 mL/kg body weight/15 minutes, the pH 3.0 ± 0.23, the titratable acidity 35.6 ± 3.09 meq/L, and the acid output, 17.0 ± 2.33 µEq/15 minutes. The urethane-anesthetized ferret also is a basal secretor of acid. The magnitude of basal acid secretion is not influenced by the estrus state of the animal [135]. Gastric acid secretion has also been studied in vitro in the fundic mucosa of 8- to 15-week-old ferrets [135,136]. The mucosa secreted acid spontaneously (0.6–1.8 µEq/cm2/h), and this increased to between 3 and 4.6 µEq/cm2/h when acetylcholine (1 × 10−4 M), pentagastrin (6.5 × 10−7 M), and histamine (1.6 × 10−6 M) were added to the system.
Histamine has an important role in the physiologic control of gastric acid secretion in humans [137], and in this context, a biochemical study of histamine metabolism in the ferret is of interest. Acid secretion is stimulated by histamine. An optimal dose of 67.3 µg/kg IP produces an acid output 30 minutes after administration of 177.0 ± 39.14 µEq/15 minutes. The stomach contains relatively low concentrations of free histamine (12 µg/g) and lacks the histamine-forming enzyme, specific l-histidine decarboxylase, although histamine-destroying activity is present. The histamine H2 receptor antagonist cimetidine abolishes the acid secretory response to exogenous histamine, and reduces the response to pentagastrin. The response to histamine (67.3 µg/kg IP) is reduced 30% by atropine and abolished by ethylenediaminetetraacetic acid (EDTA) [138]. The cholinomimetic bethanecol (0.4 mg/kg IP) evokes a stimulation of acid output comparable to that of histamine (67.3 µg/kg) but, in a volume of about 5 mL/15 minutes, approximately double that produced in response to histamine [138].
The peptide pentagastrin has its maximal effect on acid secretion after 30 minutes at a dose of 10 µg/kg, and produces 490 ± 50 uEq/15 minutes. The response is reduced by atropine and abolished by EDTA. Gastric acid secretion is also stimulated by calcium, a response that is abolished by atropine; this suggests an action via acetylcholine. Because calcium infusion in the ferret stimulates gastrin release [139], it is unclear whether the effect of atropine blocks the release of gastrin or prevents its action on the secretory cells. Gastrin cryptic peptide B has been identified in extracts of ferret gastric antral and duodenal mucosa, with larger amounts being present in the antral mucosa (708 ± 278 pmol/g) [140,141].
The role of the vagus in the control of acid secretion has also been investigated. Hypoglycemia induced by insulin (1.5 units/kg IP) produces a sustained stimulation of acid secretion that is markedly reduced by atropine and vagotomy [138]. Unilateral (dorsal) vagotomy reduces the basal acid secretion by about 60% and also reduces the response to pentagastrin and histamine, with the former being affected to a greater extent [142]. Studies in the urethane-anesthetized ferret have demonstrated that vagal activation by reduced intracellular glucose, whether induced by 2-deoxy-d-glucose (120 mg/kg IV) or electrical stimulation, both evoke a significant increase in acid secretion that is markedly reduced by atropine [142].
Proteolytic Enzymes and Mucus
In addition to being a spontaneous secretor of acid, the ferret stomach also secretes proteolytic enzymes under basal conditions. Histamine produces an increase in the secretion of proteolytic enzymes from a basal value of 757 ± 274.6 µg/15 minutes to 1698 ± 575.3 µg/15 minutes [133,138,143]. A histamine dose of 1080 µg/kg, however, is required to produce this doubling of output in contrast to the considerably lower dose (67.3 µg/kg) required to produce a 10-fold increase in acid output 30 minutes after histamine injection. A pentagastrin dose of 2 µg/kg produces a maximal response in protease secretion, in contrast to the 10 µg/kg required to provoke a maximal acid secretory response. Studies in the anesthetized ferret have demonstrated that vagal stimulation also provokes proteolytic enzyme secretion.
The physiology of gastric mucus in the ferret has been described in a study of the effect of carbenoxolone on gastric glycoprotein synthesis [144]. Carbenoxolone promotes the healing of gastric ulcers in humans. The ferret study demonstrated an increased rate of incorporation of N-acetyl glucosamine, glucosamine, galactosamine, and N-acetyl neuraminic acid into glycoproteins. No effect was observed on the incorporation of galactose or of threonine into the acid-precipitable glycoproteins. It was suggested that carbenoxolone may stimulate specific glycosyl transferases, leading to the formation of a modified glycoprotein with superior protective properties.
Surface mucous cells have been studied at the ultrastructural level [128]. Histochemical studies [132] have demonstrated that the surface mucous cells are PAS-positive. In the pyloric region, compound tubuloacinar glands are seen with foamy cytoplasm, giving PAS- and PAS-AB positive reactions that indicate the presence of neutral mucosubstances. Two features are of particular note: (1) the pyloric and duodenal Brunner's glands are similar, whereas in most other mammals, they are markedly different in their mucosubstances, and (2) of the species examined to date, only the glands in humans and ferrets do not contain acid mucosubstances. These observations, in combination with the natural infection of the ferret stomach with H. mustelae, indicate that the ferret is a good model for the study of pyloric mucous secretion in H. pylori-infected humans.
The prevalence of H. mustelae infection in adult ferrets approaches 100% and, like H. pylori infection of humans [125,145], persistent colonization of the gastric mucosa is acquired at an early age and is closely associated with development of chronic active gastritis [146], mucosal ulceration [126], and experimentally, with promotion of carcinogenesis [147,148]. The H. mustelae-infection ferret model is unique, as the ferret is the only domestic animal with natural Helicobacter-induced gastritis that develops secondary duodenal and gastric ulceration, an important clinical sequelae of H. pylori infection of humans, as the result of the host–pathogen interaction [126,146,149].
Integrity of the Gastric Mucosa
Before the recognition of H. pylori as the etiologic agent responsible for chronic gastritis and mucosal ulceration in humans, studies focused on permeability changes in the gastric mucosa in an effort to establish the etiology of gastric ulceration. Gastric transmural potential difference (pd) decreases following mucosal damage, thus providing a convenient index of mucosal integrity. In the ferret, the resting pd is 39 ± 7 mV (lumen – Ve [negative]), which decreases as the concentration of a damaging solution of acetic acid (50–200 mM) increases [150]. Flux studies reveal that as the permeability of the mucosa increases by the effect of acetic acid, H+ ions are lost from the lumen while Na+ ions enter the lumen. The ferret stomach therefore appears to be a convenient system for the study of substances that influence the permeability of the gastric mucosa and of the possible involvement of permeability in the production of gastric ulcers.
Gastric Motility
The ferret stomach possesses an inner circular smooth muscle layer and an outer longitudinal layer. It has been reported that no oblique muscle layer is present, but this awaits confirmation [102]. Interstitial cells of Cajal which are involved in the genesis of slow waves have been described in the ferret stomach and colon [151]. Historically, gastric motility had only been systematically investigated in anesthetized ferrets. Gastrointestinal motility measured with force transducers in conscious ferrets [152] indicated that the gastrointestinal motility pattern in both the interdigestive and digestive states was similar to that reported for humans. The activity front, phase III contractions of the migrating motor complex, occurred cyclically in the antrum and migrated to the duodenum and ileum in the interdigestive state, and relatively low-amplitude contractions were sustained in the antrum, duodenum, and ileum in the digestive state. Colonic motility was characterized by basal, relatively low-amplitude contractions, and a single high-amplitude contraction preceding defecation. Cisapride (0.3–3 mg/kg sc.) enhanced antral and colonic motility.
Studies of gastric motility have concentrated on the role of extrinsic autonomic innervation in the regulation of the overall gastric pressure (tone) and of the amplitude of the rhythmic contractions. The experimental approaches that have been used include direct electrical stimulation of the nerves, the effect of nerve lesions on gastric motor reflexes, and pharmacologic interference with neuromuscular function.
From a clinical perspective, upper gastrointestinal examinations using barium contrast performed in unsedated ferrets and ferrets sedated with ketamine and diazepam indicated gastric emptying may be longer in sedated ferrets (130 ± 40 minutes vs. 75 ± 54 minutes); however, this was not statistically significant. Small intestinal transit time was less than 2 hours with optimal barium contrast in the small bowel at 20 and 40 minutes post barium dosing. Longitudinal colonic mucosal folds were well visualized and aided in distinguishing small intestine from large intestine [153].
Influence of the Vagus
A comparison of the vagal innervation of the stomach in humans and the ferret demonstrated that the gross innervation of the ferret stomach generally represents a simplified version of that in humans, and the fiber composition of the nerve trunks is similar, with unmyelinated nerve fibers predominating [154]. Electrical stimulation of the peripheral cut end of the cervical or abdominal vagi evokes a frequency-related increase in the amplitude of the gastric contractions, with a maximal response being produced at 10 Hz. Responses are only evoked with stimulus parameters sufficient to activate unmyelinated axons. With stimulation periods >10 seconds, multiple contractions are produced at a frequency similar to those occurring spontaneously. The excitatory effects of vagal stimulation are mimicked by close intra-arterial injection of acetylcholine (10–100 µg/kg) and are blocked by atropine (1 mg/kg IV), as is the response to acetylcholine [155]. Further studies have revealed that the pattern of vagal stimulation also plays a complex role in determining motility and secretory responses [150].
In the anesthetized ferret, the stomach may be divided into corpus and antral regions while retaining their extrinsic innervation. Using this preparation, it was demonstrated that the vagus stimulates activity in both regions; in the presence of atropine; however, differences are observed. In the antrum, vagal stimulation inhibits the ongoing spontaneous antral contractions, whereas in the corpus, a profound long-lasting decrease in the mean pressure is observed. Following a 10-second period of stimulation at 10 Hz, it takes longer than 10 minutes for the corpus pressure to recover. These and other experiments demonstrate that the vagal preganglionic fibers activate cholinergic, excitatory, and noncholinergic non-noradrenergic inhibitory intramural neurons in both regions of the stomach. The nature of the inhibitory transmitter is unknown, but it has been suggested that VIP is involved in the corpus and neurotensin in the antrum [35].
The ease with which the stomach can be divided in the ferret greatly facilitates our understanding of the functions of the two gastric regions by highlighting differences between them in their responses not only to nerve stimulation but also to drugs such as the prokinetic cisapride [156].
Motor Reflexes.
The vagal efferents driving the intramural NANC inhibitory neurons can be activated reflexly by stimulation of gastric mucosa either mechanically or chemically, suggesting that the gastric motor response to a meal is determined both by the volume and the chemical nature of the food [157,158]. The adaptive motor responses to gastric distension have been most extensively studied, although the role of the chemical environment of the lumen in the regulation of motility has also been investigated.
The capacity of the stomach has been measured by monitoring the volume of milk ingested in a single continuous session so that the motor responses observed in response to experimental gastric distension could be related to normal gastric physiology. In ferrets with a mean body weight of 784 ± 60 g (n = 11), 94.5 ± 7.5 mL of milk was ingested at a rate of 13.0 ± 0.7 mL/min [159].
Two components of gastric motility have been studied: tone and rhythmic contractions.
Tone.
Inflation of the stomach with 50 mL of 154 mM NaCl produces a stable tone of 6.3 ± 0.85 cm H20 (relative to atmospheric pressure) [160]. Most (80%) of the fluid is accommodated in the corpus region, and pressure studies in the divided stomach demonstrate that this region is mainly responsible for determining the overall gastric tone in response to physiologic levels of distension.
Lesion studies reveal that the major reflex responsible for the relaxation of the corpus muscle in response to distension is a vagovagal reflex, involving vagal afferent activation of the intramural, NANC, inhibitory neurons described previously [35]. In the pylorus, NO has been implicated as an inhibitory transmitter modulating phasic contractile activity but raising pyloric tone [161]. Tone is maintained at low levels in the corpus by vagal activation of NANC inhibitory neurons and also by tonic sympathetic inhibition of intramural cholinergic activity [162]. Frequency of contractions is also low due to tonic inhibition of cholinergic neurons by the vagus but not the sympathetic nervous system. There appears to be little vagal involvement in contraction amplitude but there is sympathetic inhibition of this parameter, again via inhibition of cholinergic neurons. In the antrum, there is no vagally driven inhibition of tone, but a sympathetic inhibition of cholinergic neurons tends to reduce tone in the intact animal. Frequency of contractions does not appear to be extrinsically modulated. The vagus is tonically excitatory with regard to contraction amplitude in the antrum whereas the sympathetic nervous system is inhibitory, again via inhibition of cholinergic neurons. After chronic vagotomy, some adaptation appears to take place within the surviving control systems in both the corpus and the antrum. In the absence of the vagus, a splanchno-splanchnic adrenergic reflex is also demonstrated, although its characteristics are markedly different from the vagovagal reflex [163]. Studies have shown that this vagovagal inhibitory reflex may be inhibited at a central site by the GABAB receptor agonist baclofen [164].
In ferrets induced to form autoantibodies against VIP by immunization [165], the level of spontaneous corpus motility in the period immediately after surgical preparation was 15 times higher in immunized versus control animals, but there was no deficit in the ability of the corpus to accommodate fluid. This supported the hypothesis that the NANC inhibitory mechanisms regulating corpus tone and motility are different and that VIP acts primarily to regulate phasic contractile activity. Because of plasticity in the mechanisms controlling corpus tone, the effect of VIP may have been superseded during the time course of the immunization procedure. In a follow-up study [166], corpus responses to low-frequency vagal stimulation were maintained in immunized animals despite the absence of a response to exogenous VIP. This suggested that gastric reservoir function adapts to the loss of the vagal inhibitory innervation by an upregulation of intrinsic reflex pathways controlling myenteric inhibitory neurons, which are non-VIPergic.
Contractile Activity.
One of the most striking features of the ferret stomach is that it is spontaneously rhythmically active under urethane anesthesia, as is the rest of the gut immediately following surgery. The type of anesthetic regime employed during measurement of gastric motility can have significant artifactual effects [167]. Under urethane, the overall frequency of contraction is 7.1 ± 0.005 contractions/min, although the antrum and corpus contract at different frequencies when surgically uncoupled [155]. The amplitude of the contractions is reduced but not abolished by vagotomy or atropine, indicating that both cholinergic and noncholinergic systems are involved in their production.
The best defined reflex involving contraction amplitude is the corpo-antral reflex. Using the divided stomach preparation, it has been demonstrated that inflation of the corpus evokes an increase in the amplitude of antral contractions. This reflex is mediated by the vagus, and it may be involved in the initiation of gastric emptying. Distension of the duodenum produces a splanchnosplanchnic adrenergic reflex inhibition of gastric motility by a direct effect on the muscle and by modulation of cholinergic ganglionic transmission [163]. This enterogastric reflex is implicated in the regulation of gastric emptying.
Pancreas and Biliary System
Both the exocrine and endocrine portions of the ferret pancreas have been investigated, particularly with the introduction of the cystic fibrosis knockout ferret which develops multiorgan dysfunction as do human patients [168]. Cystic fibrosis kits develop progressive exocrine and endocrine pancreatic inflammatory lesions associated with onset of hyperglycemia, glucose intolerance, and abnormal insulin responses.
Under urethane anesthesia, the pancreas has a very sparse spontaneous secretion of fluid (0.05 g/10 minutes), which can be increased by secretin to about 0.5 g/10 minutes in a dose-related manner. The ferret pancreas has been used for the bioassay of the stability of secretin and its analogues [169]. Vagal stimulation and pancreozymin-cholecystokinin stimulate the secretion of amylase in animals receiving a submaximal infusion of secretin.
A comparative study of the islets of Langerhans, including those of the ferret, revealed capillary connections between the islets and exocrine tissue [170]. The direction of blood flow appears to be from the islets to the exocrine tissue, thus supporting the hypothesis that the endocrine pancreas influences the exocrine portion, possibly in a trophic manner via this vascular link [171].
A glycogenolytic factor has been extracted from the ferret pancreas [172], but comprehensive studies have not been undertaken to identify the hormones present in the islets. A small quantity of gastrin cryptic peptide B has been identified in pancreatic extracts [139]. The ferret, similar to the human, has a bile salt-dependent lipase which is expressed in adult pancreas and in the lactating mammary gland of the jill and may represent the main lipolytic system in the adult [173]. In situ hybridization using an mRNA bile salt-dependent lipase probe detected low expression in newborn ferret pancreas which suggests that lipid digestion in the newborn is dependent on milk bile salt-dependent lipase to hydrolyze lipids. In support of this, lipase activity is only two times lower in the gastric mucosa of the newborn than in the adult ferret. Bile salt-stimulated lipase activity is 10 to 20 times higher in ferret milk than in human milk and constitutes a significant portion (1–2%) of total milk protein [174]. There is an 86% homology of the N-terminal amino acid sequence between the ferret and human milk lipase homologues, indicating that the ferret is an ideal animal model for the study of the synthesis and secretion of this digestive lipase.
One of the most common clinical endocrine problems in the domestic ferret presents as episodic hypoglycemia from hyperinsulinemia produced by functional pancreatic tumors, known as insulinomas (see Chapters 17 and 24). Ferrets fed water and a regular diet were reported to have blood insulin concentrations of 19–22 µU/mL [175]. Diabetes mellitus has been documented in the ferret in multiple case reports [176–179].
The gross and microscopic anatomy of the biliary system has been described [180], but little is known of the composition of the bile, its rate of secretion, or the enterohepatic circulation. In a study concerned with biliary tract pain [19], it was noted that the resting biliary pressure is 1.5–4 mm Hg, and increasing the pressure by 5–7 mm Hg evoked a reflex opening of the sphincter of Oddi. Gallbladder contraction can be mediated by cholecystokinin which is found in abundance within the alimentary tract and has major effects on the gastrointestinal smooth muscle motility, inhibition of gastric emptying, and stimulation of small intestine and colonic motility [181]. The contractile response of the gall bladder appears to occur via direct effects on smooth muscle cells and/or neurons, whereas its effects on colonic motility are mediated by a neural mechanism with the cholecystokinin receptor subtype not yet identified.
Reconstructive gastric surgery to treat ulcers is accompanied by a change in bile reflux, gastritis, and an increased incidence of gastric cancer in humans. Surgical models in the ferret have been developed to simulate maximal and minimal bile reflux [182].
Small Intestine
The gross morphology of the small intestine has been described in Chapter 2. In their histologic study, Poddar and Murgatroyd [102] identified villi and goblet cells in all regions of the small intestine. Brunner's glands are found in the submucosa of the duodenum proximal to the opening of the bile duct, and these contain only neutral mucosubstances, as in humans [132]. The physiology of the small intestine has been less extensively investigated than the stomach, but a number of interesting features are present.
Hormone Release
The mucosa of the upper part of the small intestine contains a number of gastrointestinal hormones, in particular those involved in the control of pancreatic function such as secretin and pancreozymin-cholecystokinin (PZ-CCK). Although the mechanisms underlying the release of antral gastrin are relatively well defined, this is not the case for PZ-CCK as demonstrated in an in vitro study of segments of ferret jejunum [183]. Ferret jejunum released PZ-CCK in response to 30 mM tryptophan or phenylalanine, 7.5 mM Ca++ or 50 mM K+. The response to phenylalanine was reduced by the removal of Ca++ but was unaffected by atropine (10−6 M) or theophylline (10−3 M). Interestingly, an unidentified vasoconstrictor substance (probably not 5-HT) was also released from the mucosa by phenylalanine. These data demonstrate that this preparation has the potential for use in the study of the factors influencing the release of substances from the intestinal mucosa.
VIP and substance P have been localized to both nerve cell bodies and nerve fibers in the ferret intestine by immunohistochemistry [184]. In response to vagal stimulation in the ferret, neuronally released substance P excites, and VIP inhibits, jejunal motor activity. However, both peptides increase water secretion by the jejunal epithelium. Concentrations of these two neuropeptides are similar to those in human intestine, and the muscular layer contains much higher concentrations of these substances than the epithelial layer. The increase and decrease in jejunal motility mediated by substance P and VIP, respectively, is unaffected by cholinergic blockade (atropine).
Secretion and Absorption
Using changes in transmural pd as an index of electrogenic intestinal secretion, the relationship between jejunal motility and pd in the urethane-anesthetized ferret has been studied [185]. These studies reveal that spontaneous bursts of jejunal motility (see later discussion) are associated with increases in the pd with the lumen becoming more negative, and that vagotomy, atropine, or tetrodotoxin abolish these responses.
Motor and pd changes are evoked by stimulation of the peripheral end of the cervical vagus. These results suggest that the spontaneous bursts of motor activity and pd changes are closely linked, but whether the two events are produced by the vagus influencing both the mucosa and muscle or whether the vagus evokes the motor changes, which in turn stimulates secretion, is unknown. Other studies provide some evidence that the vagus can influence secretion directly in this tissue by both cholinergic and noncholinergic mechanisms. This linkage between motor and secretory events in the intestine has been reported in humans, and thus the ferret appears to be a useful animal model for the study of this interaction.
Very little is known of the absorptive processes in the ferret gut. The ferret kit is able to absorb maternal IgG in milk much longer than humans [186]. Closure of the gut occurs relatively late, with values from 28 to 42 days having been reported [187,188]. Corticosteroids cause premature gut closure in the rat but not in the ferret, suggesting different mechanisms of closure in rodents and carnivores [188]. The ferret can absorb intact β-carotene and convert it to retinoic acid which then enters the systemic circulation via the portal vein and, as such, has been a useful model for examining the role of β-carotene in cancer prevention [189–191] (see Chapter 32). After intestinal perfusion of β-carotene in the ferret in vivo, it was demonstrated that a substantial amount of retinoic acid is formed via an excentric cleavage pathway in the ferret intestine [190].
In addition to an understanding of these transport processes, a full description of intestinal absorption in the ferret requires information on the mucosal enzymes responsible for digestion at the brush border. Disaccharidase activity (e.g., maltose, sucrose, lactose, and trehalose) is present in extracts of ferret jejunal mucosa [192]. The activities of amylase, total proteases, monoglyceride lipase, glycyl-leucine dipeptidase, and sucrase were investigated in mucosa from five consecutive parts of the small intestine of the ferret [193]. In comparison with rats, the activity gradient of carbohydrases and total proteolytic activity were shifted in the distal direction with the conclusion that the ferret, as other predators also examined in the same study, lack the proximal to distal gradient in some enzymes characteristic of omnivores and herbivores. This observation may relate to the higher transit time of digesta in predators such as the ferret compared with the rat, with the subsequent need for continued exposure of nutrients to digestive enzymes during gastrointestinal transit.
Motility
The duodenum in the urethane-anesthetized ferret is spontaneously active, frequently showing periodic bursts of contractions. Motility is increased by local injection of cholecystokinin octapeptide [194] and by L-NAME [161]. Under urethane anesthesia, the ferret jejunum is spontaneously active, with characteristic bursts of contractions that last about 1 minute each [195]. This activity is abolished by atropine or vagotomy and mimicked to a large extent by vagal stimulation. In the presence of atropine, the immediate response to vagal stimulation is converted to a small, amplitude-delayed response. These observations indicate that the vagus influences jejunal motility mainly by a cholinergic pathway, although there is also some suggestion of a noncholinergic vagal excitatory pathway. The periodicity of the bursts is probably organized within the myenteric plexus, with tonic vagal activity having a permissive role.
Colon
A major problem in studying the physiology of the colon in the ferret involves delineation of its limits. No external division is visible between the ileum and colon, but it is possible to identify the approximate site of this junction by the vascular anatomy (see Chapter 2). Histologic studies have identified subserosal lymphoid formations that mark the most rostral limit of the colon, and electromyographic studies (see later discussion) confirm this as the region of the ileocolic junction. Also at this point, which coincides with the splenic flexure, villi are no longer present. Tubular glands with goblet cells containing predominantly or exclusively sulfated mucosubstances are found in the colon [196]. The wall of the colon is formed from circular and longitudinal muscle layers but, in contrast to several other species (e.g., guinea pig), taenia coli are not present. The only aspect of colonic physiology studied to date is motility.
Motility
Using conscious ferrets [197], electrical activity of the intestine has been used to identify the location of the colon. With electrodes placed anterior to the subserosal lymphoid tissue, activity is characterized by continuous slow wave activity (3.06 ± 0.6 cycles/min). Irregular and regular spiking activities of 37.2 ± 6.7 minutes and 5.9 ± 0.4 minutes, respectively, have also been recorded, resembling that seen in the dog ileum. Tissue activity typical of that recorded in the colon of related species is observed at recording sites in the 8–10 cm of intestine caudal to the subserosal lymphoid tissue; slow wave activity with a frequency of 9–13 cycles/min is present only 7% of the time. Long and short spike bursts have been recorded; the former had a duration of 21.3 ± 7.8 seconds and the latter 4.6 ± 2.5 seconds. Long spike bursts are propagated aborally, whereas short bursts are not. Histologic studies reveal a strip of fibrous connective tissue that disrupts the continuity of the muscle layers between the intestinal region with ileal electrical activity and the region with colonic electrical activity. This strip of connective tissue prevents the electrotonic spread of the migrating myoelectric complex from the ileum to the colon, a function served by the ileocolonic sphincter in the dog.
Under urethane anesthesia, colonic pressure activity changes have been observed with a similar temporal pattern to the electrical activity recorded in the conscious animal [198–200] This spontaneous activity is markedly reduced by atropine (1 mg/kg IV) or by vagal cooling, suggesting that much (but not all) of the colonic contractile activity is due to a vagal-cholinergic pathway. Electrical stimulation of the peripheral cut end of the cervical or abdominal vagus evokes a frequency-related increase in colonic motility, with a peak response occurring at 10 Hz. This response is markedly decreased by atropine, and is replaced by a delayed noncholinergic, non-noradrenergic excitatory response. Similar colonic responses are observed when the vagal efferents supplying the colon are activated reflexly by vagal afferent stimulation. These results demonstrate that the motor activity of the ferret colon is largely vagus-dependent, mediated by both cholinergic and noncholinergic transmitter systems, with the former predominating [198–200]. The vagal noncholinergic excitation of colonic motility is of particular interest in view of the increasing importance attached to this type of control system.
The influence of sacral innervation on the colon has been the subject of preliminary investigation [103]. The colonic excitatory fibers are confined to the first sacral segment, and the excitatory effect is mainly the result of an effect on the longitudinal muscle coat. Two aspects are of interest in this paper: (1) the importance of lymphoid tissue for distinguishing the ileum from the colon was recognized and (2) retroperistalsis was evoked in the colon [103]. This type of activity has been observed in other gut regions prior to vomiting. A study of the mechanisms underlying this phenomenon in the ferret colon would therefore be of interest, because such disordered motility has been implicated in the genesis of vomiting.
Preliminary studies of gut transit time measured values between 148 and 219 minutes for animals fed a meat-based diet [200]. Further studies are required using different diets and markers to confirm these rather rapid times.
Blood
Hematologic and serum chemistry data for the ferret have been reported [201–203] and are reviewed in Chapter 7 of this volume. Hematologic data are also available for the mink [204].
Blood Groups
Evidence for existence of blood groups in domestic ferrets was sought by testing serum samples for naturally acquired or experimentally induced erythrocyte antibodies capable of causing direct agglutination, antiglobulin-enhanced agglutination (Coombs test), or lysis [205]. There was no evidence of naturally acquired blood group antibodies or antibodies formed in response to repeated transfusion or after serial inoculation subcutaneously with red cells. Consequently, blood groups either do not exist in domestic ferrets or are only weakly immunogenic such that transfusion without cross matching appears to have minimal risk in ferrets [206].
Hematocrit and Erythrocyte Physiology
The hematocrit is in general higher than in other common laboratory species, as is the erythrocyte count itself. To maximize recovery of plasma or serum, it is advisable to take a blood sample three times the volume of the desired plasma sample and spin it for about 20% longer than for samples from other species (P. Andrews, unpublished data).
Isoflurane has been shown to rapidly decrease the hematocrit, hemoglobin, plasma protein concentration, and red cell and white blood cell count after induction of anesthesia [203]. After the initial decrease, all values partially recovered to preanesthetic baseline by 45 minutes after anesthesia. Although these alterations are apparently well tolerated in healthy ferrets, anemic, geriatric, or debilitated ferrets require close monitoring if isoflurane anesthesia is indicated (see Chapter 12).
Membranes
Ferret red blood cells have been used as a convenient and plentiful source of cell membranes for the investigation of the transport and permeability characteristics of biologic membranes in general. Preliminary studies performed on ferret red cells established its potential as an alternative source to the cat and dog for carnivore red cells. Intracellular concentrations of cations are as follows (mean ± standard error per liter of original cells); Na+, 95.7 ± 1.1 mmol/L; K+, 3.9 ± 0.2 mmol/L; Mg++, 3.01 ± 0.12 mmol/L; Ca++, 8–10 µmol/L [207]. The ATP concentration was also measured, and found to be 0.60 ± 0.04 mmol/L original cells, sufficient to drive sodium or calcium pumps, although the ferret red cell does not appear to have an active sodium pump [207].
There is evidence that ferret red cells may be divisible into two populations based on potassium transport characteristics. Ionic flux studies [208] revealed that the red cell membranes are highly permeable to potassium, in contrast to cat and dog red cells, and this can be resolved into a rapid component that accounts for 70–90% of the movement and a slow component that accounts for the remainder. The high-ceiling loop diuretic bumetanide (0.1 mM) inhibits potassium influx by 80–90% and sodium by 60–70%, suggesting that these ions move into the cell using a sodium-potassium cotransport system. The ferret red cell membrane has a number of transport characteristics not found in other carnivores, which makes further studies of these cells of great interest.
Hemoglobin
Ferret erythrocytes contain two hemoglobins that share a common β chain but have α chains that differ by one amino acid (Asp15—-Gly15), consistent with a high degree of homology among members of the order Carnivora [209]. Red cell content of hemoglobin is higher in the male than the female ferret, and the values in both sexes are higher than those in the dog and rat [9]. No data are available on the oxygen transport characteristics of ferret hemoglobin, although measurements of the blood oxygen content indicate that they are likely to be similar to those of other species. Ferret erythrocytes have been used to study the effects of deoxygenation on red blood cell kinase-mediated phosphorylation of ion transporters [210].
Blood Gases and pH
These parameters have been measured in anesthetized ferrets. Using urethane anesthesia in separate studies, the arterial blood pH was 7.283 ± 0.025, with a bicarbonate concentration of 15.7 ± 0.7 mM, PaCO2 was 30.4 ± 0.6 mm Hg and PaO2 was 95.0 ± 1.4 mm Hg (P. Andrews, unpublished data). These observations demonstrate the stability of the urethane-anesthetized ferret. In the barbiturate-anesthetized animal, Kempf and Chang [12] reported that the oxygen content of arterial blood was 15.7 ± 2.6 mL/100 mL blood and that of venous blood was 10.2 ± 1.8 mL/100 mL blood, giving an arteriovenous difference of 5.5 ± 1.7 mL/100 mL of blood and an oxygen consumption of 7.2 ± 2.5 mL/min (body weight, 823 ± 114 g; surface area, 0.51–0.67 m2). Medetomidine-midazolam-ketamine anesthesia induced relative hypoxia requiring supplemental oxygen during anesthesia [211]. Respiratory depression associated with elevated PaCO2 was greater in ferrets than rats under isoflurane anesthesia [212]. Additional information on the effects of anesthesia on gas exchange is found in Chapter 12.
Thrombosis
The ferret may be a useful species for evaluating antithrombotic drugs in a model in which aspirin is efficacious [213]. A 10-minute anodal electrical stimulation of 1 mA delivered to the external surface of the carotid artery to induce vascular injury produced an occlusive thrombus within 41 ± 3 minutes with an average weight of 8 ± 1 mg. Aspirin (10 mg/kg, IV) or ifetroban, a thromboxane receptor antagonist, administered at 1 mg/kg followed by 1 mg/kg per hour, IV, each given 15 minutes before electrical injury, reduced the thrombus weight by 58% and 74%, respectively. The ferret was more sensitive than the rat to aspirin inhibition of collagen-induced platelet aggregation and was intermediate between the rat and humans in response to thromboxane receptor agonists.
The blood coagulation profile of ferrets has been compared with coagulation in rats [214]. The ferret-activated partial thromboplastin time was slightly longer, and in contrast, the prothrombin time and thrombin time were profoundly shorter in ferrets compared with rats. Fibrinogen levels were two times higher in plasma from ferrets than rats. Although heparin prolonged blood coagulation times, higher doses of heparin were required to double the activated partial thromboplastin time in ferrets compared with effective doses in the rat.
Serum Chemistry
Serum chemistry data for the ferret have been reported [201–203] and are reviewed in Chapter 7 of this volume. Most serum biochemistry parameters that have been measured in the ferret are consistent with data obtained from other common laboratory species except for higher serum glucose and ammonia, and lower blood urea nitrogen (BUN) (see later discussion). Hypergammaglobulinemia, interpreted to reflect chronic antigenic stimulation, is commonly associated with Aleutian disease [215].
Glucose
In the fasting state, the ferret has a relatively high blood glucose level (Chapter 7). This has led to some difficulties in adapting the 2-deoxy-d-glucose autoradiographic technique to the ferret, but it has been used successfully nonetheless [216]. Hypoglycemia is a common clinical finding in ferrets with insulinoma [217,218].
Osmolarity
In the conscious ferret, plasma osmolarity is 328 ± 1 mOsm/kg and, following water deprivation for 24 hours, is increased by 12% to 366 ± 11 mOsm/kg (PLR Andrews, pers. comm.).
Plasma Proteins
In the ferret, the total serum lipoprotein concentration is similar to values measured for humans and dogs but differs in composition of lipids [219]. The major lipoprotein fraction is of high density (1.063–1.21 g/mL). The very low-density lipoprotein contains proportionately less triglyceride and more phospholipid than in humans, and the low-density lipoprotein contains proportionately more triglyceride and less cholesterol than in humans. Marked similarities have been observed in the gel electrophoretic pattern of human and ferret lipoprotein apoproteins, and in the solubility of apoproteins in tetramethylurea. These similarities between human and ferret lipoproteins may make it a useful animal model for the study of lipid metabolism.
Ammonia
The resting serum ammonia level in the ferret is 200–400 µg/dL [220] and is therefore considerably higher than in humans (17–80 µg/dL). Hyperammonemia (2000–7000 µg/dL) is produced 2–3 hours after feeding a diet containing less than 0.3% arginine, or by an intraperitoneal injection of jack bean urease. These results are important for several reasons. First, ferrets appear to tolerate very high levels of ammonia and remove ammonia from their system rapidly. Second, the ease with which hyperammonemia can be induced makes the ferret an ideal animal model for the study of ammonia metabolism, which is of particular relevance because of the association between hyperammonemia and Reyes syndrome [221]. Toxicity from arginine-free diet-induced hyperammonemia appears related to decreased methylation activity in the liver and transamidination activity in the kidney [222]. Elevated levels of toxic intermediates of arginine metabolism such as guanidinosuccinate, coupled with deficient hepatic creatinine synthesis, may play a role in the pathophysiology of hyperammonemia.
Nervous System
The role of the nervous system in general, and of the hypothalamus in particular, in the regulation of reproduction in the ferret has been the subject of study for over 60 years and is certainly the best characterized CNS function in this animal. Use of the ferret in the study of reproductive neuroendocrinology is reviewed elsewhere in this book, as is the influence of sex hormones on behavior (see Chapter 8). The general anatomy of the ferret brain has been reviewed by Lockard [223] and in Chapter 3 of this volume. The hypothalamic–pituitary relationship in the ferret and other animals has been described in detail. This section will concentrate on select neurophysiologic studies which highlight areas of research for which the ferret, with its relatively large gyrencephalic brain, may be of particular use.
Central Nervous System
For convenience, the central and autonomic components of the nervous system are discussed separately with subdivisions made on both functional and anatomic grounds.
Special Senses
The neurophysiology of the special senses is the major area of study. These investigations originally began because of the influence of photoperiod on reproduction. More importantly for developmental studies, the ferret nervous system at birth is relatively immature, particularly when compared with the cat, and thus provides a system in which the development of the CNS can be studied postnatally at stages that occur in the cat in utero. The “rewired” neonatal ferret, in which axons of the special senses, such as the visual system, are encouraged to grow and connect to alternative sites within the CNS, has yielded insight into CNS development and regeneration of nervous tissue (see Chapter 30).
Audition
Use of the ferret in audition science research is described in Chapter 29. The ear canals are closed until near the end of the first month of life at which point a startle response to loud sounds is observable [224]. Postnatal development of hearing progresses from birth through day 40 when adult-like thresholds can be observed in auditory brainstem measurements [225]. During development through 28–30 days after birth, axonal projections from the lateral superior olivary nuclei, the dorsal cochlear nucleus, and the dorsal nucleus of the lateral lemniscus converge in frequency-ordered layers in the central nucleus of the inferior colliculus where they distribute among different synaptic compartments and become functional for hearing at approximately 1 month of age [226].
Because it is relatively easy to train, there is a large literature base on using behavioral responses of the ferret to establish auditory capabilities. Although little is known about the function of the auditory nerve in ferrets, recent data on audition variables such as frequency tuning indicate that the auditory nerve and cochlea in the ferret are most similar to that of guinea pigs and chinchillas. The ferret has good low-frequency hearing similar to humans, but this capability is thought to result from auditory nerve fibers that are characteristically sensitive to higher frequencies and thus may not be the best model for human hearing loss [227].
Relative pitch perception in ferrets has been demonstrated in ferrets trained to discriminate between two-tone sequences that rise or fall over a four-octave frequency range [228].
Vision
The ferret has historically been a popular animal model for vision research. Because the eyes in the ferret do not open until about 28 days after birth, the ferret provides an ideal animal model for studying not only the general development of the visual system, but also how it is influenced by visual stimuli. In addition to the advantage of an immature visual system at birth, the albino ferret has been used to evaluate abnormal organization at various levels of the visual system from the retina through to the primary visual cortex that are found in albino mammals, including humans [229]. The ferret, therefore, provides an experimental system in which both the genetic and developmental influences on the visual system can be investigated separately and in combination.
The reader is referred to an extensive description of ferret vision in Chapters 2 and 30.
Olfactory System
The olfactory epithelium of the ferret was described histologically as early as 1938 as part of investigations of influenza infection [230]. The ferret, like many other carnivores, has an extensive turbinate system [231]. Turbinate growth in young ferrets is characterized by cartilaginous and osseous thickening as a normal developmental feature of growing ferrets [232]. A detailed description of the olfactory cortex noted that the lateral olfactory tract (LOT) projects to most of the pyriform cortex [233]. Latency measurements indicate that fine collateral branches of the LOT connect with the posterior olfactory cortex, whereas mainly larger diameter (faster conducting) fibers connect with the anterior olfactory cortex. Studies of sex discrimination and partner preference in male and female ferrets have shown that volatile anal scent gland odorants from male and female ferrets activate clusters of glomeruli located in the ventral–caudal portion of the main olfactory bulb of breeding ferrets of both sexes without activation in the accessory olfactory bulb [234]. Ferrets appear to lack glomeruli with dense acetylcholinesterase (AChE) activity as noted in rodents stimulated by body odors from conspecifics.
In the ferret as well as in cats and monkeys, development of visual cortical columns, barrels in somatosensory cortex, and glomeruli in the olfactory bulb, appear to arise from precise, rapid establishment of initial connections that are not as plastic as in later responses to sensory activity during the postnatal critical period [235].
Hypothalamus
The vast majority of studies on the hypothalamus have investigated its interactions with the pineal gland and the pituitary, and their roles in reproduction. These aspects are discussed elsewhere (see Chapters 8 and 17), and this section will briefly discuss the levels of neurotransmitters in the hypothalamus.
5-HT was detected by radioenzymatic assay in the septum, preoptic area, suprachiasmatic area, medial forebrain bundle, pineal and lateral (LHA), and anterior hypothalamic (AHA) areas; the pineal has the highest levels [236]. The levels of 5-HT in the pineal region and of the LHA and AHA vary slightly with time of day. Only the LHA and pineal levels are influenced by the prevailing photoperiod. Differential rhythms in pineal and anterior hypothalamic 5-HT levels reported by [236] were not confirmed [237]. Although 5-HT is probably a neurotransmitter in the ferret hypothalamus, the proposal that the pineal influences the hypothalamic-pituitary axis via 5-HT requires further studies before definitive answers can be given regarding its role in the modulation of reproduction.
In homogenates of the entire hypothalamus, activities of adrenalin (163 ± 11 pmol/g), norepinephrine (3912 ± 330 pmol/g), dopamine (701 ± 99 pmol/g), and norepinephrine N-methyl transferase (NMT; 246 ± 30 pmol/h/g) have been identified. The levels of norepinephrine, dopamine, and NMT are closer to those for the dog than the cat, but the adrenalin level is substantially lower than either the dog or cat [238].
Cerebellum
Two types of study on the physiology of the ferret cerebellum have been undertaken, one type directed toward understanding its role in jaw reflexes [239] and the other toward elucidation of its role in locomotion [240–242].
Field potentials were evoked in lobules 4 and 5 (ipsilateral > contralateral) in response to stimulation of the inferior alveolar nerve. The conduction velocity of the inferior alveolar axons in their peripheral course was 52 m/s, and further calculations of conduction time within the CNS indicated that the projection to the granular layer is direct. The function of such a rapid direct first-order projection needs further evaluation.
In decerebrate ferrets, recordings have been made from cerebellar Purkinje cells in lobules 5 and 6 during treadmill walking [240]. A complete description of this study is beyond the scope of this chapter, but it was shown that during locomotion, the Purkinje cell discharge is not very well modulated. If locomotion is suddenly perturbed, however, the discharge is clearly modulated, and this is particularly associated with an increase in complex spikes. This report suggests that the ferret may be a suitable alternative to the cat for this type of locomotor research. In addition, the ferret has been used for investigation of the effects of spinal compression injuries on locomotion [241,242]. A combination of these two types of locomotor studies may provide insight into the problems of locomotion in persons with selective cord lesions.
Purkinje cell firing in the cerebellum is an important conditioned response responsible for motor responses such as eye blink. Conditioned Purkinje cell responses have a specific temporal profile that underlies the time course of overt response behaviors, and it is believed that the temporal properties of the conditioned Purkinje cell response are determined by the temporal pattern of the parallel fiber impulses generated by the conditioned stimulus. In decerebrate ferret preparations [243], conditioned stimulus as brief as one or two impulses in the mossy fibers can be sufficient to elicit a full conditioned Purkinje cell response with normal time course, suggesting that parallel fiber input to the Purkinje cell influences the firing rate several hundred milliseconds later. These findings contrast to the traditional understanding of how parallel fiber impulses are coordinated temporally in conditioned responses.
Brain Stem
The brain stem has been investigated in the ferret mainly to determine the location and characteristics of the cells of origin of the abdominal vagus. The anatomy of the vagal brainstem connections have been described in Chapter 3, and the vagal afferents were the subject of a detailed study [244]. Recordings from the dorsal medulla in the region of the obex reveal respiratory or cardiovascular phased multiunit activity. Ventral to this layer, in the region of the dorsal motor vagal nucleus, single units have been identified and demonstrated by collision (an electrical stimulation technique to demonstrate afferent and efferent fibers [245]) to project axons to the abdomen [246]. The conduction velocity of the axons is in the range for that of unmyelinated fibers. Indirect evidence suggests that some of the abdominal vagal efferents may be reflexly activated by vagal afferents via monosynaptic or polysynaptic pathways [247]. Electrophysiologic studies of brainstem auditory-evoked responses are discussed later (see Reye's syndrome).
The levels of various neurotransmitters have been measured in homogenates of whole brain stem: adrenalin 45 ± 5 pmol/g; norepinephrine, 2125 ± 109 pmol/g; and dopamine, 174 ± 19 pmol/g. These values lie midway between those for the cat and dog measured in the same study [238]. The levels of norepinephrine N-methyl transferase (191 ± 37 pmol/h/g tissue) are comparable to those in the dog but very low in comparison to those in the cat (895 ± 53 pmol/h/g tissue).
Stimulation of abdominal vagal afferents produces an increase in the uptake of [3H]-2-deoxyglucose by the area postrema [248]. This provides evidence that vagal afferents may evoke vomiting by direct activation of the vomiting chemoreceptor trigger zone located within the area postrema (see Chapter 31).
Cortex
The ferret has a large, gyrencephalic brain, making it a good candidate for comparing the conservation among mammals of the molecular ultrastructure of the neocortex, variously established using cytoarchitecture to be organized in six layers in the dorsal telencephalon of mammals, including marsupials and monotremes [249]. Evaluating expression of 15 orthologs for layer-enriched molecular markers previously demonstrated in rodent neocortex, it was determined that most layer-specific markers had similar gene expression patterns between mouse and ferret cortex, supporting layer-specific cell-type identity conservation across mammalian superorders [250]. Expression of deep layer markers (ER81/ETV1, SULF2, PCP4, FEZF2/ZNF312, CACNA1H, KCNN2/SK2, SYT6, FOXP2, CTGF) indicated stratification of layers 5 and 6 into 5a, 5b, 6a, and 6b, also conserved between rodents and carnivores. Investigators found that layer-specific gene expression varied more across areas of ferret cortex than between homologous areas of mouse and ferret cortex, particularly in superficial layer markers such as SERPINE2, MDGA1, CUX1, UNC5D, RORB/NR1F2, and EAG2/KCNH5. Layer 4 markers RORB and EAG2 identified molecular sublamination to ferret visual cortex and molecular dissociation among the so-called agranular areas of the neocortex.
Disease Models
The ferret has been used in neurophysiologic studies to elucidate the mechanisms of several neurologic disorders.
Lissencephaly
The ferret has a gyrencephalic brain, and the onset of gyrification has been studied by MRI [251]. Cortical folding develops in a lateral to medial gradient during the period of most rapid cerebral cortical growth and is largely complete prior to myelination of cortical axons. Although lissencephaly is a rare syndrome in humans, the importance of an animal model in which this syndrome can be readily produced lies in two areas. In humans, lissencephaly is associated with epileptic seizures. Thus, a study of the lissencephalic cortex may provide insight into the genesis of such seizures in patients with far less extensive lesions. An understanding of how lissenchephaly may be induced experimentally will yield important information about the mechanisms of normal cortical histogenesis, and reveal how this may become disordered to produce various disturbing CNS birth defects.
Treatment of pregnant ferrets with the alkylating agent methylazoxymethanol acetate on gestation day 32 or 33 leads to the production of offspring with lissencephalic hydrocephalic cerebral hemispheres and a brain weight that is 28% less than normal [252]. Behaviorally, the animals have a markedly impaired learning ability when tested in several types of maze. The spectrum of EEG activity is modified, particularly during sleep. Although the stimulation current required to evoke an epileptiform after-discharge is no different from that for control animals, the duration of after-discharges and seizures is longer. This effect indicates a generally enhanced sensitivity in the lissencephalic animals. Also, these animals respond to treatment with antiepileptic drugs, although less well than gyrencephalic animals [252]. Neurochemical studies have revealed an increase in indices of cholinergic and catecholaminergic nerve terminals and little if any change in GABAergic systems. Further studies are required to determine whether neurochemical changes can be related to the EEG changes, which would suggest new approaches for the design of antiepileptic drugs.
Reye's Syndrome
This often fatal childhood syndrome of unknown etiology is characterized by a number of CNS problems, including coma, seizures, cerebral edema, and increased intracranial pressure. Although the cause is unknown, its appearance has been associated with hyperammonemia, salicylates, and viruses, particularly influenza. Ferrets are particularly susceptible to influenza B, and hyperammonemia may be readily induced by an arginine-deficient diet [253]. Combining these treatments with acetylsalicylic acid administration results in animals with several clinical signs of Reye's syndrome. Electrophysiologic studies have revealed that wave 1 (eighth nerve generator potential) of the auditory brainstem-evoked response is delayed in latency compared with that of controls, and wave 4 (brainstem nuclei response potential) is initially delayed. By 10 days after treatment, however, the latency returns to near normal values [254]. It was suggested that the central components of the auditory system recover more quickly than the peripheral components. Because of its sensitivity to all three of the factors predisposing to Reye's syndrome, the ferret is an excellent model for the study of its etiology and possible treatments (see Chapter 5 for further discussion).
High-Pressure Neurologic Syndrome (HPNS)
The profound neurologic effects (e.g., convulsions) of exposure to high hydrostatic pressures have attracted interest in connection with deep sea diving and as a research method for producing reversible changes in the excitability of the nervous system. In a comparative study of HPNS, the ferret had a high susceptibility to HPNS comparable with that of the mouse, guinea pig, squirrel, and rhesus monkey [255]. These preliminary observations indicate that the ferret may be a useful animal model for the study of HPNS.
Autonomic Nervous System
A complete anatomic description of the autonomic nervous system in the ferret has not been published, but the innervation of several organs has been reported: trachea [48,256,257]; heart [258]; stomach [259]; kidney [260], and bladder and colon [103,261,262].
The influence of the autonomic innervation on a number of visceral functions is discussed in the cardiovascular, respiratory, gastrointestinal, and urogenital sections of this chapter. This section will describe electrophysiologic studies of the autonomic nervous system that were concerned with the trachea and gut.
Tracheal Innervation
Because of its length and the large number of submucosal glands, the ferret trachea is popular for use in studies of the neural control of airway secretion. In addition, the parasympathetic nerves contract the tracheal smooth muscle, and the neural control of secretion and motility can thus be investigated in the same tissue. Associated with the ferret trachea are two chains of paratracheal ganglia separated by interganglionic nerve trunks [256,257]. The inputs to the ganglia appear to be both cholinergic and noncholinergic. The extrinsic innervation is from the laryngeal nerve. The ganglia each contain 10 to 20 cells, with cell body diameters ranging from 15 to 40 µm. Based on electrical characteristics, two cell types may be distinguished: AH cells with a resting membrane potential of −37 ± 7 mV, which have a marked hyperpolarization following an action potential, and nonspiking B cells with a resting potential of −52 ± 11 mV. Stimulation of the nerve trunks evokes fast excitatory and inhibitory postsynaptic potentials in the AH cells and in some of the B cells. The interganglionic nerve trunks are composed mainly of unmyelinated nerves, as are the postganglionic axons supplying the smooth muscle cells (calculated conduction velocity, 0.1–0.2 m/s). The interganglionic nerve trunks are important not only in distributing the extrinsic input to separated ganglion cells but also in the dissemination of postganglionic fibers to the smooth muscle cells [87,256]. The paratracheal ganglia may not act solely as simple relay stations, but may also play a role as peripheral integrators of respiratory reflexes. Electrophysiological studies have now been performed on ferret nodose ganglion cells in vitro (the cell bodies of vagal afferents). Substance P was shown to induce depolarization via an NK1 receptor, activating a calcium-dependent, outward flow potassium current [263].
It has been demonstrated that transmission between the pre- and postganglionic nerves can be modulated by norepinephrine acting on presynaptic α-adrenergic receptors [264]. The sympathetic nerves influence tracheal function not only by direct action on the muscle and submucosal glands but also by modulating the effects of the extrinsic parasympathetic drive. A similar mechanism has been proposed for the influence of the greater splanchnic (sympathetic) nerves on gastric motility [163]. Ganglionic transmission in the trachea is also reduced by barbiturates at doses comparable to those producing general anesthesia [265]. This observation suggests that barbiturates may not be the best anesthetics for use in studies of the respiratory system in the ferret; unfortunately, reports of pulmonary mechanics in this species commonly use barbituate anesthesia [37,49].
The neural mediators associated with the tracheal plexus have been characterized [266]. Choline acetyltransferase (ChAT), nitric oxide synthase (NOS), VIP, and substance P (SP) in the intrinsic neurons of this plexus were measured by distribution of double- and triple-labeled neurons as quantified in cryostat sections and in whole mounted specimens. Eighty-five percent of nerve cell bodies with ChAT immunoreactivity were located in ganglia of the longitudinal trunks, or the closely associated bridge ganglia, with the balance (15%) of ChAT-positive neurons located in ganglia of the superficial muscular plexus. Conversely, VIP-positive neurons were most frequent in the superficial muscular plexus (>75%) and <10% were observed in the longitudinal trunks or bridge neurons. Most NOS-and SP-positive neurons were also located in the superficial muscular plexus. The superficial muscular plexus had neurons with the following neurochemical profiles: 11% only NOS, 20% only VIP, 5% only SP, 67% NOS and VIP, and 40% VIP and SP. NOS, VIP, and SP were frequently localized in the same nerve cell body. The occurrence of nerve terminals containing only SP located around the borders of individual NOS/VIP/SP-containing neurons suggests possible sensory innervation to the airway neurons. The results demonstrated that most cholinergic nerves do not contain VIP, NOS, or SP, which are predominantly located in the superficial muscular plexus ganglia, cholinergic neurons are predominantly located in the longitudinal trunk ganglia, and nerve terminals containing exclusively SP, suggesting possible sensory origin, are closely associated with some neurons in the plexus.
Neurons within the nucleus tractus solitarius (nTS) release glutamate and express c-Fos protein and the AMPA subtype of glutamate receptor when activated by airway sensory systems that reflexly induce airway constriction, such as in response to aerosolized histamine or capsaicin [267]. Associated glutamate release within the nTS and blockade of the AMPA receptor subtype inhibited cholinergic outflow to the airways, indicating that glutamate and AMPA receptor signaling pathways participate in transmission of afferent inputs from the airways to the nTS and induce reflex airway constriction. Nicotinic acetylcholine receptors (nACh-R) also have a role in the transmission of afferent constricting inputs from bronchopulmonary receptors to the nTS and in reflex airway constriction in ferrets [268]. When activation of airway sensory receptors by inhalation of aerosolized histamine induced c-Fos expression in a subset of nTS neurons, the nAch-R α3 receptor was activated within the commissural subnucleus by nicotine and the result was increased cholinergic outflow to the airways. Hexamethonium (nACh-R blocker) blocked these effects but had no significant effects on airway reflex constrictions induced by lung deflation. These results suggest that the nACh-R is expressed by nTS neurons in response to inputs from airway sensory receptors, and activation by compounds such as nicotine will increase cholinergic outflow to the airways. Nonetheless, the nACh-R pathways are not required for reflex bronchoconstriction.
Gastrointestinal Innervation
Afferent activity has been recorded in vagal fibers supplying the esophagus and stomach [122,159] and in greater splanchnic nerves innervating the gallbladder and biliary tree [19]. Recordings have been made from vagal afferent mechanoreceptors supplying the mid-thoracic striated muscle portion of the esophagus. The discharge rate is related to the degree of esophageal distention, and stretch of the circular muscle produces a more marked response than longitudinal distention, although the discharge is relatively slow-adapting in both cases. The afferent axons are small diameter myelinated fibers, with conduction velocities between 7.5 and 15 m/s [122].
In contrast to the esophageal mechanoreceptors, the vagal afferents innervating the gastric smooth muscle are unmyelinated, with a mean conduction velocity of 0.91 ± 0.21 m/s [159]. The most interesting feature of the gastric mechanoreceptors is that their precise response to gastric distention depends on their location in the stomach. During gastric filling, most fluid is accommodated in the gastric body, and the discharge in the mechanoreceptors with receptive fields in this region gradually increases with increasing volume. The antrum is little distended but an increase in the level of contractile activity is reflexly evoked [158], and this is signaled by the antral mechanoreceptors. The receptors in both regions are “in-series” tension receptors. The nature of their response is largely determined by differences in the properties of the muscle in the two gastric regions. The “in-series” nature of the corpus mechanoreceptors has been confirmed [269].
One study of visceral afferents traveling with the abdominal sympathetic nerves investigated the role of biliary system afferents in the production of visceral pain [19]. In response to distension of the gallbladder and biliary ducts, two types of afferent activity were evoked: (1) low-threshold afferents activated by levels of distension but not producing reflex increases in blood pressure (innocuous stimulation) and (2) high-threshold afferents activated by degrees of distension that evoked changes in blood pressure (noxious stimulation). The latter type was most commonly found in the biliary tree. These results indicate that specific nociceptors may exist in the gallbladder, and the ferret may thus provide a suitable animal model for the investigation of visceral nociception.
Descriptions of the vagal afferent projections to the brain stem and various aspects of the immunochemistry of the dorsal vagal complex in the ferret have been published [270–274]. In addition, the functional consequences of vagal afferent activation have been studied using immunohistochemistry to detect expression of c-Fos [275,276].
Endocrine System
Most studies of ferret endocrinology have investigated aspects of the reproductive system; these are reviewed in Chapters 8 and 17 and therefore will not be discussed in detail here. For convenience, references are given to studies in which the levels of reproductive hormones were measured. Many of the investigations of the ferret endocrine system have been performed in the context of the high prevalence of adrenal-associated endocrinopathy and insulin-secreting pancreatic islet tumors. It is of note that (1) adrenal gland disease is a significant clinical problem in both pet and research colony ferrets; (2) the pathophysiology of this condition differs significantly from classical Cushing's or Addison's disease encountered in humans and other species; (3) insulin-secreting tumors are of unknown etiology and may occur together with adrenal-associated endocrinopathy; and (4) thyroid disease, either hyper- or hypothyroidism, has not been reported in the ferret (see later discussion). Apart from these investigations there is sparse information on the endocrine system. Preliminary studies have been performed and will be briefly described.
Reproductive Endocrinology
Testis
The effect of luteinizing hormone (LH), hypothalamic lesions, the amygdala, development, season, and time of day on testosterone secretion have all been investigated [277–280]. In addition, the influence of LH-RH (releasing hormone) has also been studied. Testosterone has been detected in plasma from spayed and intact females [280] and is elevated in mature intact males that develop Sertoli cell tumors [281].
Ovary
Progesterone levels in plasma have been measured before and during implantation, and in pregnant and pseudopregnant animals [282,283]. The influence of day length on the secretion of progesterone, estradiol, and estrone has also been studied [284]. Progesterone metabolites can be measured in feces and are highest in lactating females [285]; elevated systemic levels likely prevent nursing ferrets from returning to estrus. In the ferret, progesterone has been shown to increase the level of glycogen in the liver (but not in muscle), and a more marked response is observed in pseudopregnant animals [286]. Hyperestrogenism is caused by persistent estrus in the intact female ferret (coital stimulation induces ovulation), or in the spayed female, by remnants of ovarian tissue. In addition, any of the sex steroids, particularly estradiol, may be elevated in hyperadrenocorticism [287].
Vaginal cytology of the black-footed ferret (Mustela nigripes), Siberian polecat (Mustela eversmanni), and domestic ferret (Mustela putorius furo) is similar [288]. Proestrus is characterized by an increasing percentage of superficial epithelial cells and enlargement of the vulva. During estrus, 90% of the cells collected by vaginal lavage are superficial keratinized epithelial cells. Neutrophils are more commonly found during any stage of the estrus cycle in ferret vaginal cytology compared with that of other species. Large cells, probably of uterine symplasma origin, may be observed in vaginal lavages following whelping or pseudopregnancy.
Anterior Pituitary
The influence of LH-RH, day length, estrus and anestrus, ovariectomy, the hypothalamus, and development, on the secretion of LH have been investigated [289,290]. Follicle-stimulating hormone (FSH) secretion has been studied in relation to development, hypothalamic stimulation, LH-RH, day length, and ovariectomy [289–291]. The half-life of FSH and LH in estrus, anestrus, and ovariectomized animals has been measured by Donovan and Gledhill [292].
Dose-related increases in circulating LH in response to exogenously administered LH-RH has been demonstrated in both prepubertal (9-week-old) and postpubertal (32-week-old) male ferrets [293]. A significant effect of age on LH response was observed, with the prepubertal animals demonstrating significantly greater serum LH values than the postpubertal males. Prepubertal ferrets also exhibited a significant increase in endogenous LH pulse amplitude in sampling periods following exogenous administration of LH-RH compared with baseline pulse amplitudes in periods prior to the LH-RH infusions. These results suggest that the low frequency of endogenous LH pulses previously observed in prepubertal ferrets is not due to unresponsiveness of the pituitary gland to LH-RH. Thus, suppression of the hypothalamo-hypophyseal axis observed in the prepubertal ferret is probably mediated at the level of the hypothalamus.
The male ferret displays renewed testicular activity when the days lengthen, and this has been associated with stimulation of the pulsatile liberation of LH reflecting the pulsatile activity of the gonadotropin-releasing hormone (GnRH) system when daylight was equal to or in excess of 8 hours light/16 hours dark [33]. This photoperiod coincides precisely with the phase of photosensitivity known to be essential for gonadal stimulation in this species [294].
Adrenal Glands
The gross morphology, histology, and vasculature of the adrenal glands have been described by Holmes [295]. The adrenal cortex has the three main zones usually encountered in other mammals (glomerulosa, fasciculata, and reticularis) and, in addition, a zona intermedia and juxtamedullaris are also found. The adrenal glands are drained by relatively large veins which facilitate local sampling of circulating metabolites but also can complicate adrenalectomy (see Chapters 2 and 17). The adrenal weight increases when the female ferret transitions from late proestrus to estrus compared with anestrus or early proestrus. There is also some indication that the stainable lipid in the cortex increases in estrus, but this finding varies [295].
Effects of ACTH and dexamethasone stimulation on plasma concentrations of cortisol were studied in two groups of adult male ferrets [296]. The mean basal plasma cortisol values were not significantly different between ACTH stimulation (1.29 ± 0.84 µg/dL) and dexamethasone suppression test (0.74 ± 0.56 µg/dL) experiments. Intravenous injection of 0.5 IU of ACTH/ferret induced a significant increase in plasma cortisol concentrations by 30 minutes (5.26 ± 1.21 µg/dL), which persisted until 60 minutes (5.17 ± 1.99 µg/dL) after injection. Following IV injection of 0.2 mg of dexamethasone/ferret, plasma cortisol values significantly decreased as early as 1 hour (0.41 ± 0.13 µg/dL), and had further decreased by 5 hours (0.26 ± 0.15 µg/dL).
Sex steroid-producing adrenocortical tumors are common in gonadectomized ferrets [297,298]. It is believed that LH binding to a G-protein coupled receptor, LHR, redirects differentiation of multipotential stem/progenitor cells in the subcapsular region of the adrenal from corticoid-producing cells to sex steroidogenic cells [297]. The adrenal glands of healthy ferrets produce only limited amounts of androgenic steroids, while adrenocortical neoplasms that arise in neutered ferrets typically secrete androgens or their derivative, estrogen. Whereas normal adrenal gland tissue from healthy ferrets lacks staining, these ferret adrenal tumors are typically positive by immunohistochemistry for cytochrome b5 which is a regulator of an androne biosynthetic enzyme (cytochrome P450 17α-hydroxylase/17, 20-lyase) and therefore can be used as a biomarker of androgen biosynthesis by neoplastic adrenal tissue [299].
Hyperadrenocorticism in the ferret has been reviewed [300]. The clinical signs that develop in ferrets with adrenal gland hyperplasia or functional adenoma or adenocarcinoma (see Chapter 17) apparently are caused by excessive secretion of one or more steroids other than cortisol, although conflicting data have been published. One group reported that the ratio of urinary cortisol and creatinine may be useful for clinical diagnosis [301]. In 51 clinically normal ferrets, ratios ranged from 0.04 × 10−6 to 1.66 × 10−6, with a median value of 0.22 × 10−6. The values were significantly higher in ferrets with adrenal tumors, with a range of 0.5 × 10−6 to 60.13 × 10−6 and a median of 5.98 × 10−6. In contrast, ferrets with adrenal disease commonly have pre- and post-ACTH cortisol concentrations within reference range limits established for the ferret [287,302]. Plasma concentrations of several sex steroids including androstenedione, 17-hydroxyprogesterone, and estradiol are commonly elevated singly or in combination in clinically affected ferrets, so it is advisable to measure more than one [287]. Ultrasound may have equal or more diagnostic utility than measurement of serum hormone levels [303].
Interestingly, there have been no published case reports of pituitary-dependent adrenal hyperplasia or of spontaneous adrenal insufficiency in ferrets.
Thyroid
Serum thyroxine and triiodothyronine radioimmunoassay values have been established for the normal ferret [302]. Serum thyroxine (T4) and 3,3′,5-triiodothyronine (T3) values were obtained from intact and neutered males and females and assayed using a commercial radioimmunoassay. Serum T4 values ranged from 1.01 to 8.29 µg/dL for males (3.24 ± 1.65 µg/dL) and 0.71 to 3.43 µg/dL for females (1.87 ± 0.79 µg/dL). Serum T4 values of adult female ferrets, juvenile ferrets (less than 1 year old) of either sex, and castrated males were similar to the normal T4 values of the cat, 1.20–3.80 µg/dL. Intact adult male ferrets had higher serum T4 values which were more comparable to those of the normal dog, which range from 1.52 to 3.60 µg/dL. Serum T3 values ranged from 0.45 to 0.78 ng/mL for males (0.58 ± 0.09 ng/mL), and 0.29 to 0.73 ng/mL for females (0.53 ± 0.13 ng/mL). These values are comparable to those of dogs and cats which range from 0.50 to 1.50 ng/mL.
Effects of thyroid-stimulating hormone (TSH) and thyrotropin-releasing hormone (TRH) on plasma concentrations of thyroid hormones were examined in a study of 13 ferrets [296]. Plasma T4 values increased as early as 2 hours post injection (from 2.69 ± 0.87 µg/dL to 3.37 ± 1.10 µg/dL) and remained high until 6 hours (3.45 ± 0.86 µg/dL) after IV injection of 1 IU of TSH/ferret. In contrast, IV injection of 500 µg of TRH/ferret did not induce a significant increase until 6 hours (from 1.81 ± 0.41 µg/dL to 2.75 ± 0.79 µg/dL) after injection, and induced side effects of hyperventilation, salivation, vomiting, and sedation. There was no significant increase in triiodothyronine (T3) values following TSH or TRH administration. There have been no published case reports of hyper- or hypothyroidism in the ferret.
Posterior Pituitary
Preliminary studies have demonstrated the presence of bioactive antidiuretic hormone (ADH) in the plasma of anesthetized and conscious ferrets, and the level increases as a result of water deprivation. The control of ADH secretion has not been studied in detail, but large increases in its output are produced by the D2 receptor agonist apomorphine, and by electrical stimulation of the central cut end of the abdominal vagus [304].
Anterior Pituitary
Growth hormone physiology has received limited attention in the ferret. Light and electron microscopic immunocytochemistry of somatotropes in the anterior pituitary gland localized somatotropes in the pars distalis with no immunoreactive cells detected in the pars tuberalis [305]. Ultrastructural studies classified somatotropes into two subtypes on the basis of size of the secretory granules. Type I cells, with less developed organelles and small granules (mean diameter, 192 nm), were considered to be the immature somatotrope, while Type II cells, which were much more predominant in number and had a distinct Golgi zone, many mitochondria, and comparatively larger secretory granules (mean diameter, 257 nm), were considered to be the matured form of Type I cells. Several intermediate cell types were also encountered and may represent different transitional phases during the conversion of Type I to Type II. Protein A-gold immunoelectron microscopy localized growth hormone over the granules, with no labeling over any other cytoplasmic organelles of the two somatotrope subtypes.
Gastrointestinal Tract
Although the gut is usually regarded as the largest endocrine gland, this area has seldom been investigated in the ferret. The features studied have been previously discussed in this chapter (see “Gastrointestinal Tract” and its subsections on “Gastric Secretion,” “Small Intestine,” and “Pancreas and Biliary System”).
Urinary System
Kidneys
Glomerular Filtration Rate
Three methods of determining glomerular filtration rate (GFR) were performed in adult ferrets, aged 9 months to 7 years old [306]. Endogenous creatinine clearance was determined using serum and urine creatinine values obtained during 24- and 48-hour collection periods from 27 ferrets housed in metabolic cages. Creatinine and radiolabeled inulin were administered to 12 female ferrets by constant IV infusion during isoflurane anesthesia. Mean ± SD endogenous creatinine clearance in ferrets for metabolic cage collections was 2.50 ± 0.93 mL/min/kg of body weight. There were no significant differences between the 24- and 48-hour clearance rates. Mean inulin clearance was 3.02 ± 1.78, and mean exogenous creatinine clearance was 3.32 ± 2.16 mL/min/kg. Infused inulin clearance, the generally preferred method for GFR calculation in mammalian species, was significantly higher in younger (3.65 mL/min/kg) compared with older ferrets (2.29 mL/min/kg). Results of this study indicate that both inulin and creatinine clearance are adequate measures of GFR in ferrets as in other species. It is important to note that clinical biochemical evidence of renal failure in the ferret may not closely parallel that routinely observed in other domestic animals and humans. The increase BUN, if renal-based, may not be paralleled by an increase of serum creatinine of the same magnitude. Mean serum creatinine concentrations are lower in ferrets, and the range of normal values is narrower, compared with values in dogs and cats (see Chapter 7). It is not yet clear if glomerular filtration is the sole mechanism for elimination of creatinine in ferrets.
Morphologically the kidneys are similar to those in other species, but histologic studies have revealed two interesting features. First, ectopic glomeruli are present [307] and second, the intrarenal arteries have relatively thicker walls than in other laboratory animals [9]. The functional significance of these observations is unclear.
The autonomic innervation of the kidney is more complex than that reported for other species. In summary, the innervation is from two main sources: the aorticorenal ganglion and the direct renal artery. In the region of the adrenal gland, the aorticorenal ganglion gives rise to 2 to 4 fine branches. These course into the kidney in proximity to the renal artery and, in addition, the lower lumbar sympathetic chain gives rise to a single nerve that runs directly to the kidney, termed the direct renal nerve. No ganglion was observed along the course of the nerve [260,308]. Reports of such a direct renal nerve in other species have not been found. Electron microscopic studies of the direct and indirect renal nerves reveal that they both contain small myelinated and unmyelinated axons, with the latter predominating.
Physiologic studies have demonstrated the presence of efferent activity in both nerves. Of particular interest is the finding that in response to hemorrhage, some nerves decrease their discharge rather than increase it as reported in other species. It has been speculated that these units may represent sympathetic dopaminergic vasodilator nerves known to supply the kidney in some species, although the dopamine concentration in the ferret renal cortex (0.014 ± 0.004 µg/g) is low [15]. Of the rat, guinea pig, cat, and rabbit, only the guinea pig has a higher level of norepinephrine (1.44 ± 0.154 µg/g) than the ferret, suggesting a predominant role for norepinephrine in the regulation of renal function in these species.
As observed in other species [309], the kidney of the Inactin-anesthetized ferret (120 mg/kg IP) produces urine, and lesion of the renal nerves leads to a denervation natriuresis [36]. This demonstrates a tonic, sympathetically driven reabsorption of sodium ions.
Although little is known of the details of urine production, bladder urine osmolarity may reach 2000 mOsm/L, indicating a considerable concentrating ability and supporting the hypothesis that the ferret may have originated from a desert-dwelling animal. Urine-specific gravity values in 69 clinically healthy pet ferrets less than 1 year old were reported to average 1.051 for intact males (range 1.034–1.070) and 1.042 for intact females (range 1.026–1.060) [310]. Conscious ferrets fed a commercial dog diet with water ad libitum had a urine output of 8–140 mL/24 hours, containing sodium, 0.2–6.7 mmol/24 hours, potassium, 0.9–9.6 mmol/24 hours, and chloride, 0.3–8.5 mmol/24 hours [9]. These values depend on the nature of the diet and, in particular, water intake is directly related to the salt content of the diet. In view of the effect of high levels of sodium intake on the development of hypertension, it is possible that some of the high blood pressure values reported in this species were produced by excess dietary sodium. In this study, most animals were proteinuric, with urine values for protein of up to 33 mg/dL, but the reason for this is unknown [9].
Bladder and Ureters
Under anesthesia, the ureters show spontaneous rhythmic contractions propagating toward the bladder, but nothing is known of the control of this activity. The bladder in a large ferret (2 kg) can accommodate about 10 mL of urine at a relatively low pressure (5 cm H2O). The control of the bladder is of particular interest, because both the hypogastric (sympathetic) and sacral (parasympathetic) nerves cause contraction of the entire bladder, whereas in other animals (e.g., cat), the sympathetic nerves usually produce relaxation. The bladder also contracts in response to adrenalin, and the contractions to sympathetic stimulation and adrenalin are converted to a relaxation state by chrysotoxin (an ergot derivative). This experiment was performed in 1905, and it is important because it was one of the first studies to demonstrate that the effects of sympathetic nerve stimulation are mimicked by adrenalin, and more significantly, that both responses are similarly modified by an antagonist (chrysotoxin) [263,311]. These experiments on the ferret were undertaken jointly by Elliott and Dale [312], with Dale winning the Nobel Prize for Physiology and Medicine along with Loewi in 1934 for studies on the mechanism of neurotransmission. Those readers familiar with this topic will recognize these observations as constituting two of “Dale's criteria” for the demonstration of neurotransmission. Thus, the unusual response of the ferret bladder played an important role in the elucidation of neurotransmission mechanisms. Interestingly, the pharmacologic basis of the dual effects of adrenalin and sympathetic stimulation on the bladder has not yet been established, although studies have demonstrated contractile responses to cholinergic and purinergic agonists in vitro [313].
Distribution of β-adrenoceptor subtypes in the detrusor of the ferret were investigated in functional experiments in vitro and in vivo using a variety of β-adrenoceptor agonists and antagonists. Relaxation of the ferret detrusor by β-adrenoceptor activation is mediated mainly via the β-adrenoceptor, as in the human detrusor [314,315].
Acknowledgments
The author gratefully acknowledges the work of Dr. Paul L.R. Andrews, who contributed to this chapter in the second edition of this book.
References
1. Onuma M, Kondo H, Ono S, Ueki M, Shibuya H, Sato T (2009) Radiographic measurement of cardiac size in 64 ferrets. J Vet Med Sci 71: 355–358.
2. Wagner RA (2009) Ferret cardiology. Vet Clin North Am Exot Anim Pract 12: 115–134, vii.
3. Stepien RL, Benson KG, Forrest LJ (1999) Radiographic measurement of cardiac size in normal ferrets. Vet Radiol Ultrasound 40: 606–610.
4. Scheer P, Svoboda P, Sepsi M, Janeckova K, Doubek J (2010) The electrocardiographic Holter monitoring in experimental veterinary practice. Physiol Res 59(Suppl. 1): S59–S64.
5. Truex RC, Belej R, Ginsberg LM, Hartman RL (1974) Anatomy of the ferret heart: an animal model for cardiac research. Anat Rec 179: 411–422.
6. Brahmajothi MV, Morales MJ, Campbell DL, Steenbergen C, Strauss HC (2010) Expression and distribution of voltage-gated ion channels in ferret sinoatrial node. Physiol Genomics 42A: 131–140.
7. Andrews PL, Bower AJ, Illman O (1979) Some aspects of the physiology and anatomy of the cardiovascular system of the ferret, Mustela putorius furo. Lab Anim 13: 215–220.
8. Erickson H, Swenson M, Reece W (1993) Exercise physiology. In: Swanson MJ, ed. Duke's physiology of domestic animals. Ames, IA: Iowa University Press, pp. 304–308.
9. Thornton PC, Wright PA, Sacra PJ, Goodier TE (1979) The ferret, Mustela putorius furo, as a new species in toxicology. Lab Anim 13: 119–124.
10. Kohler E, Bieniek H (1975) Pharmacological investigations on the cardiovascular system of the anesthetized mink (Mustela vison). Z Versuchstierkd 17: 145–154.
11. Dudas-Gyorki Z, Szabo Z, Manczur F, Voros K (2011) Echocardiographic and electrocardiographic examination of clinically healthy, conscious ferrets. J Small Anim Pract 52: 18–25.
12. Kempf JE, Chang HT (1949) The cardiac output and circulation time of ferrets. Proc Soc Exp Biol Med 72: 711–714.
13. Fish R, Kohn D, Wixson S (1997) Anesthesia and analgesia in laboratory animals. Boston: Academic Press.
14. Albrecht D, Davidowa H (1989) Action of urethane on dorsal lateral geniculate neurons. Brain Res Bull 22: 923–927.
15. Bell C, Gillespie JS (1981) Dopamine and noradrenaline levels in peripheral tissues of several mammalian species. J Neurochem 36: 703–706.
16. Liang C, Rounds NK, Dong E, Stevens SY, Shite J, Qin F (2000) Alterations by norepinephrine of cardiac sympathetic nerve terminal function and myocardial beta-adrenergic receptor sensitivity in the ferret: normalization by antioxidant vitamins. Circulation 102: 96–103.
17. Qin F, Rounds NK, Mao W, Kawai K, Liang CS (2001) Antioxidant vitamins prevent cardiomyocyte apoptosis produced by norepinephrine infusion in ferrets. Cardiovasc Res 51: 736–748.
18. Conlon K, Kidd C (1999) Neuronal nitric oxide facilitates vagal chronotropic and dromotropic actions on the heart. J Auton Nerv Syst 75: 136–146.
19. Cervero F (1982) Afferent activity evoked by natural stimulation of the biliary system in the ferret. Pain 13: 137–151.
20. Andrews P, Bhandari P, Gupta A, et al. (1992) The von Bezold-Jarisch reflex in the ferret: evidence for involvement of non-vagal 5HT3 receptors. Lorenzini Foundation Symposium Milan 223: A48.
21. Yamano M, Ito H, Kamato T, Miyata K (1995) Species difference in the 5-hydroxytryptamine 3 receptor associated with the von Bezold-Jarisch reflex. Arch Int Pharmacodyn Ther 330: 177–189.
22. Verberne AJ, Guyenet PG (1992) Medullary pathway of the Bezold-Jarisch reflex in the rat. Am J Physiol 263: R1195–R1202.
23. Watson JW, Gonsalves SF, Fossa AA, McLean S, Seeger T, Obach S, Andrews PL (1995) The anti-emetic effects of CP-99,994 in the ferret and the dog: role of the NK1 receptor. Br J Pharmacol 115: 84–94.
24. Makale MT, King GL (1992) Surgical and pharmacological dissociation of cardiovascular and emetic responses to intragastric CuSO4. Am J Physiol 263: R284–R291.
25. Andrews PL, Dutia MB, Harris PJ (1984) Angiotensin II does not inhibit vagally induced bradycardia or gastric contractions in the anaesthetized ferret. Br J Pharmacol 82: 833–837.
26. Evans CA, Waud DR (1973) A pressor effect of high doses of tubocurarine in the ferret. Pharmacology 10: 32–37.
27. Vastenburg MH, Boroffka SA, Schoemaker NJ (2004) Echocardiographic measurements in clinically healthy ferrets anesthetized with isoflurane. Vet Radiol Ultrasound 45: 228–232.
28. Maxwell MP, Hearse DJ, Yellon DM (1987) Species variation in the coronary collateral circulation during regional myocardial ischaemia: a critical determinant of the rate of evolution and extent of myocardial infarction. Cardiovasc Res 21: 737–746.
29. Gomoll AW, Lekich RF (1990) Use of the ferret for a myocardial ischemia/salvage model. J Pharmacol Methods 23: 213–223.
30. Gomoll AW (1996) Cardioprotection associated with preconditioning in the anesthetized ferret. Basic Res Cardiol 91: 433–443.
31. Flores NA, Davies RL, Penny WJ, Sheridan DJ (1984) Coronary microangiography in the guinea pig, rabbit and ferret. Int J Cardiol 6: 459–471.
32. Willis LS, Barrow MV (1971) The ferret (Mustela putorius furo L.) as a laboratory animal. Lab Anim Sci 21: 712–716.
33. Jallageas M, Boissin J, Mas N (1994) Differential photoperiodic control of seasonal variations in pulsatile luteinizing hormone release in long-day (ferret) and short-day (mink) mammals. J Biol Rhythms 9: 217–231.
34. Joffre J, Joffre M (1973) Seasonal changes in the testicular blood flow of seasonally breeding mammals: dormouse, Glis glis, ferret, Mustella furo, and fox, Vulpes vulpes. J Reprod Fertil 34: 227–233.
35. Andrews PL, Lawes IN (1985) Characteristics of the vagally driven non-adrenergic, non-cholinergic inhibitory innervation of ferret gastric corpus. J Physiol 363: 1–20.
36. Daniel P, Pritchard M (1975) Studies of the hypothalamus and the pituitary gland with special reference to the effects of transection of the pituitary stalk. Acta Endocrinol (Copenh) 201(Suppl.): 1–216.
37. Vinegar A, Sinnett EE, Kosch PC (1982) Respiratory mechanics of a small carnivore: the ferret. J Appl Physiol 52: 832–837.
38. Peake MD, Harabin AL, Brennan NJ, Sylvester JT (1981) Steady-state vascular responses to graded hypoxia in isolated lungs of five species. J Appl Physiol 51: 1214–1219.
39. Emery CJ, Sloan PJ, Mohammed FH, Barer GR (1977) The action of hypercapnia during hypoxia on pulmonary vessels. Bull Eur Physiopathol Respir 13: 763–776.
40. Suggett AJ, Mohammed FH, Barer GR (1980) Angiotensin, hypoxia, verapamil and pulmonary vessels. Clin Exp Pharmacol Physiol 7: 263–274.
41. Chammas JH, Rickaby DA, Guarin M, Linehan JH, Hanger CC, Dawson CA (1997) Flow-induced vasodilation in the ferret lung. J Appl Physiol 83: 495–502.
42. Wiener CM, Banta MR, Dowless MS, Flavahan NA, Sylvester JT (1995) Mechanisms of hypoxic vasodilation in ferret pulmonary arteries. Am J Physiol 269: L351–L357.
43. Yu AY, Frid MG, Shimoda LA, Wiener CM, Stenmark K, Semenza GL (1998) Temporal, spatial, and oxygen-regulated expression of hypoxia-inducible factor-1 in the lung. Am J Physiol 275: L818–L826.
44. Belser JA, Katz JM, Tumpey TM (2011) The ferret as a model organism to study influenza A virus infection. Dis Model Mech 4: 575–579.
45. Fisher JT, Zhang Y, Engelhardt JF (2011) Comparative biology of cystic fibrosis animal models. Methods Mol Biol 742: 311–334.
46. Johnson-Delaney CA, Orosz SE (2011) Ferret respiratory system: clinical anatomy, physiology, and disease. Vet Clin North Am Exot Anim Pract 14: 357–367, vii.
47. Vinegar A, Sinnett EE, Kosch PC, Miller ML (1985) Pulmonary physiology of the ferret and its potential as a model for inhalation toxicology. Lab Anim Sci 35: 246–250.
48. Robinson NP, Venning L, Kyle H, Widdicombe JG (1986) Quantitation of the secretory cells of the ferret tracheobronchial tree. J Anat 145: 173–188.
49. Boyd RL, Mangos JA (1981) Pulmonary mechanisms of the normal ferret. J Appl Physiol 50: 799–804.
50. Ben-Jebria A, Crozet Y, Eskew ML, Rudeen BL, Ultman JS (1995) Acrolein-induced smooth muscle hyperresponsiveness and eicosanoid release in excised ferret tracheae. Toxicol Appl Pharmacol 135: 35–44.
51. Oldham MJ, Phalen RF, Huxtable RF (1990) Growth of the ferret tracheobronchial tree. Lab Anim Sci 40: 186–191.
52. Phalen RF, Oldham MJ, Beaucage CB, Crocker TT, Mortensen JD (1985) Postnatal enlargement of human tracheobronchial airways and implications for particle deposition. Anat Rec 212: 368–380.
53. Wiener CM, Dunn A, Sylvester JT (1991) ATP-dependent K+ channels modulate vasoconstrictor responses to severe hypoxia in isolated ferret lungs. J Clin Invest 88: 500–504.
54. Zhang S, Garbutt V, McBride JT (1996) Strain-induced growth of the immature lung. J Appl Physiol 81: 1471–1476.
55. McBride JT (1985) Postpneumonectomy airway growth in the ferret. J Appl Physiol 58: 1010–1014.
56. Hyde DM, Samuelson DA, Blakeney WH, Kosch PC (1979) A correlative light microscopy, transmission and scanning electron microscopy study of the ferret lung. Scan Electron Microsc 3: 891–898.
57. Jacob S, Poddar S (1982) Mucous cells of the tracheobronchial tree in the ferret. Histochemistry 73: 599–605.
58. Boat T, Cheng P, Caterson B, Johnstone B, Leigh M (1992) Proteoglycan secretory components of surface epithelial cells in postnatal ferret trachea. Chest 101: 10S.
59. Barber HW, Small PA, Jr. (1977) Construction of an improved tracheal pouch in the ferret. Am Rev Respir Dis 115: 165–169.
60. Kim JS, Okamoto K, Arima S, Rubin BK (2006) Vasoactive intestinal peptide stimulates mucus secretion, but nitric oxide has no effect on mucus secretion in the ferret trachea. J Appl Physiol 101: 486–491.
61. Borson DB, Chin RA, Davis B, Nadel JA (1980) Adrenergic and cholinergic nerves mediate fluid secretion from tracheal glands of ferrets. J Appl Physiol 49: 1027–1031.
62. Cho HJ, Joo NS, Wine JJ (2010) Mucus secretion from individual submucosal glands of the ferret trachea. Am J Physiol Lung Cell Mol Physiol 299: L124–L136.
63. Loughlin GM, Gerencser GA, Crowder MA, Boyd RL, Mangos JA (1982) Fluid fluxes in the ferret trachea. Experientia 38: 1451–1452.
64. Corrales RJ, Nadel JA, Widdicombe JG (1986) Sources of fluid component of secretions from tracheal submucosal glands in cats. J Appl Physiol 56: 1076–1082.
65. Lazarus SC, Basbaum CB, Barnes PJ, Gold WM (1986) cAMP immunocytochemistry provides evidence for functional VIP receptors in trachea. Am J Physiol 251: C115–C119.
66. Kyle H, Robinson NP, Widdicombe JG (1986) Simultaneous measurement of airway mucus secretion and smooth muscle tone in ferret trachea in vitro. J Physiol 327: 38.
67. Coles SJ, Neill KH, Reid LM (1984) Potent stimulation of glycoprotein secretion in canine trachea by substance P. J Appl Physiol 57: 1323–1327.
68. Rogers DF, Aursudkij B, Barnes PJ (1989) Effects of tachykinins on mucus secretion in human bronchi in vitro. Eur J Pharmacol 174: 283–286.
69. Hahn HL, Hummer B, Freyer-Barski A, Bleicher S (1990) [Stimulation of mucous glands of the respiratory tract by neuropeptides: comparison of exogenous administration and endogenous liberation]. Pneumologie 44(Suppl. 1): 526–528.
70. Borson DB, Charlin M, Gold BD, Nadel JA (1984) Neural regulation of 35SO4-macromolecule secretion from tracheal glands of ferrets. J Appl Physiol 57: 457–466.
71. Peatfield AC, Barnes PJ, Bratcher C, Nadel JA, Davis B (1983) Vasoactive intestinal peptide stimulates tracheal submucosal gland secretion in ferret. Am Rev Respir Dis 128: 89–93.
72. Tom-Moy M, Basbaum CB, Nadel JA (1983) Localization and release of lysozyme from ferret trachea: effects of adrenergic and cholinergic drugs. Cell Tissue Res 228: 549–562.
73. Deffebach ME, Islami H, Price A, Webber SE, Widdicombe JG (1990) Prostaglandins alter methacholine-induced secretion in ferret in vitro trachea. Am J Physiol 258: L75–L80.
74. Barnes PJ, Basbaum CB, Nadel JA, Roberts JM (1982) Localization of beta-adrenoreceptors in mammalian lung by light microscopic autoradiography. Nature 299: 444–447.
75. Barnes PJ, Basbaum CB, Nadel JA, Roberts JM (1983) Pulmonary alpha-adrenoceptors: autoradiographic localization using [3H]prazosin. Eur J Pharmacol 88: 57–62.
76. Basbaum CB, Ueki I, Brezina L, Nadel JA (1981) Tracheal submucosal gland serous cells stimulated in vitro with adrenergic and cholinergic agonists: a morphometric study. Cell Tissue Res 220: 481–498.
77. Nadel JA (1990) Neutral endopeptidase modulation of neurogenic inflammation in airways. Eur Respir J Suppl 12: 645s–651s.
78. Adler KB, Schwarz JE, Anderson WH, Welton AF (1987) Platelet activating factor stimulates secretion of mucin by explants of rodent airways in organ culture. Exp Lung Res 13: 25–43.
79. McBride RK, Stone KK, Marin MG (1992) Arachidonic acid increases cholinergic secretory responsiveness of ferret tracheal glands. Am J Physiol 262: L694–L698.
80. Gao Y, Zhou H, Raj JU (1995) PAF induces relaxation of pulmonary arteries but contraction of pulmonary veins in the ferret. Am J Physiol 269: H704–H709.
81. Reo NV, Barnett JK, Neubecker TA, Alexander ME, Goecke CM (1992) A nuclear magnetic resonance investigation of the upper airways in ferrets: I. Effects of histamine and methacholine. Magn Reson Med 27: 21–33.
82. Baraniuk JN, Lundgren JD, Mizoguchi H, Peden D, Gawin A, Merida M, Shelhamer JH, Kaliner MA (1990) Bradykinin and respiratory mucous membranes: analysis of bradykinin binding site distribution and secretory responses in vitro and in vivo. Am Rev Respir Dis 141: 706–714.
83. Barnett JK, Cruse LW, Proud D (1990) Kinins are generated in nasal secretions during influenza A infections in ferrets. Am Rev Respir Dis 142: 162–166.
84. Hahn HL, Kleinschrot D, Hansen D (1992) Nicotine increases ciliary beat frequency by a direct effect on respiratory cilia. Clin Investig 70: 244–251.
85. Rasmussen RE, McClure TR (1992) Effect of chronic exposure to NO2 in the developing ferret lung. Toxicol Lett 63: 253–260.
86. McBride RK, Oberdoerster G, Marin MG (1991) Effects of ozone on the cholinergic secretory responsiveness of ferret tracheal glands. Environ Res 55: 79–90.
87. Coburn RF (1984) Neural coordination of excitation of ferret trachealis muscle. Am J Physiol 246: C459–C466.
88. Barnes PJ, Basbaum CB, Nadel JA (1983) Autoradiographic localization of autonomic receptors in airway smooth muscle: marked differences between large and small airways. Am Rev Respir Dis 127: 758–762.
89. McWilliam PN, Gray SJ (1990) The innervation of tracheal smooth muscle in the ferret. J Auton Nerv Syst 30: 233–238.
91. Holzer P (1988) Local effector functions of capsaicin-sensitive sensory nerve endings: involvement of tachykinins, calcitonin gene-related peptide and other neuropeptides. Neuroscience 24: 739–768.
92. Maggi CA, Meli A (1988) The sensory-efferent function of capsaicin-sensitive sensory neurons. Gen Pharmacol 19: 1–43.
94. Mizrahi J, D′Orleans-Juste P, Caranikas S, Regoli D (1982) Effects of peptides and amines on isolated guinea pig tracheae as influenced by inhibitors of the metabolism of arachidonic acid. Pharmacology 25: 320–326.
95. Vitkun SA, Foster WM, Bergofsky EH, Poppers PJ (1990) Large and small airway responses to bronchoconstrictors in the guinea pig and ferret. Lung 168: 249–257.
96. Franconi GM, Graf PD, Lazarus SC, Nadel JA, Caughey GH (1989) Mast cell tryptase and chymase reverse airway smooth muscle relaxation induced by vasoactive intestinal peptide in the ferret. J Pharmacol Exp Ther 248: 947–951.
97. Uddman R, Luts A, Arimura A, Sundler F (1991) Pituitary adenylate cyclase-activating peptide (PACAP), a new vasoactive intestinal peptide (VIP)-like peptide in the respiratory tract. Cell Tissue Res 265: 197–201.
98. Chimes MJ (1993) A technique for catheterization of ferrets for chronic intratracheal material administration. Lab Anim Sci 43: 346–349.
100. Young S (1981) The effect of vagotomy on the deflation reflex, tracheal occlusion and the pattern of breathing in the rat and ferret. J Physiol 310: 62.
101. Rarey KE, DeLacure MA, Sandridge SA, Small PA, Jr. (1987) Effect of upper respiratory infection on hearing in the ferret model. Am J Otolaryngol 8: 161–170.
102. Poddar S, Murgatroyd L (1977) Morphological and histological study of the gastrointestinal tract of the ferret. Acta Anat (Basel) 96: 321.
103. Elliott TR, Barclay-Smith E (1904) Antiperistalsis and other muscular activities of the colon. J Physiol 31: 272.
104. Poddar S, Jacob S (1977) Gross and microscopic anatomy of the major salivary glands of the ferret. Acta Anat (Basel) 98: 434–443.
105. Ekstrom J, Hakanson R, Mansson B, Tobin G (1988) Tachykinin involvement in parasympathetic nerve-evoked salivation of the ferret. Br J Pharmacol 94: 707–712.
106. Jacob S, Poddar S (1978) The histochemistry of mucosubstances in ferret salivary glands. Acta Histochem 61: 142–154.
107. Khosravani N, Ekman R, Ekstrom J (2007) Acetylcholine synthesis, muscarinic receptor subtypes, neuropeptides and secretion of ferret salivary glands with special reference to the zygomatic gland. Arch Oral Biol 52: 417–426.
108. Mangos JA, Boyd RL, Loughlin GM, Cockrell A, Fucci R (1981) Secretion of monovalent ions and water in ferret salivary glands: a micropuncture study. J Dent Res 60: 733–737.
109. Mangos JA, Boyd RL, Loughlin GM, Cockrell A, Fucci R (1981) Transductal fluxes of water and monovalent ions in ferret salivary glands. J Dent Res 60: 86–90.
110. Mangos JA, Boyd RL, Loughlin GM, Cockrell A, Fucci R (1981) Handling of calcium by the ferret submandibular gland. J Dent Res 60: 91–95.
111. Ekstrom J (1989) Autonomic control of salivary secretion. Proc Finn Dent Soc 85: 323–331; discussion 361–323.
112. Ekstrom J, Olgart L (1986) Substance P-evoked salivary secretion in the ferret. J Physiol 372: 41.
113. Tobin G, Mirfendereski S, Ahstrom T, Ekstrom J (1995) Fluid and protein secretion from ferret submandibular and parotid glands in response to sympathetic nerve stimulation or administration of sympathomimetics. Acta Physiol Scand 153: 231–241.
114. Ekstrom J, Mansson B, Olgart L, Tobin G (1988) Non-adrenergic, non-cholinergic salivary secretion in the ferret. Q J Exp Physiol 73: 163–173.
115. Skude G, Mardh PA (1976) Isoamylases in blood, urine, and tissue homogenates from some experimental mammals. Scand J Gastroenterol 11: 21–26.
116. Tobin G, Asztely A, Edwards AV, Ekstrom J, Hakanson R, Sundler F (1995) Presence and effects of pituitary adenylate cyclase activating peptide in the submandibular gland of the ferret. Neuroscience 66: 227–235.
117. Roudebush P, Jones BD, Vaughan RW (1986) Medical aspects of esophageal disease. In: Jones BD, Liska WD, eds. Canine and feline gastroenetrology. Philadelphia: W.B. Saunders, p. 57.
118. Blackshaw LA, Dent J (1997) Lower oesophageal sphincter responses to noxious oesophageal chemical stimuli in the ferret: involvement of tachykinin receptors. J Auton Nerv Syst 66: 189–200.
119. Blackshaw LA, Haupt JA, Omari T, Dent J (1997) Vagal and sympathetic influences on the ferret lower oesophageal sphincter. J Auton Nerv Syst 66: 179–188.
120. Young RL, Page AJ, Cooper NJ, Frisby CL, Blackshaw LA (2010) Sensory and motor innervation of the crural diaphragm by the vagus nerves. Gastroenterology 138: 1091–1101 e1091-1095.
121. Larson JE, Herring SW (1996) Movement of the epiglottis in mammals. Am J Phys Anthropol 100: 71–82.
122. Andrews PLR, Lang K (1983) Vagal afferent discharge from mechanoreceptors in the lower oesophagus of the ferret. J Physiol 332: 29.
123. Blackshaw LA, Nisyrios V, Dent J (1995) Responses of ferret lower esophageal sphincter to 5-hydroxytryptamine: pathways and receptor subtypes. Am J Physiol 268: G1004–G1011.
124. Blanco MC, Fox JG, Rosenthal K, Hillyer EV, Quesenberry KE, Murphy JC (1994) Megaesophagus in nine ferrets. J Am Vet Med Assoc 205: 444–447.
125. Fox JG, Correa P, Taylor NS, Lee A, Otto G, Murphy JC, Rose R (1990) Helicobacter mustelae-associated gastritis in ferrets: an animal model of Helicobacter pylori gastritis in humans. Gastroenterology 99: 352–361.
126. Whary MT, Palley LS, Batchelder M, Murphy JC, Yan L, Taylor NS, Fox JG (1997) Promotion of ulcerative duodenitis in young ferrets by oral immunization with Helicobacter mustelae and muramyl dipeptide. Helicobacter 2: 65–77.
127. Andrews PLR (1988) Data from the laboratory of PLR Andrews.
128. Pfeiffer CJ (1970) Surface topology of the stomach in man and the laboratory ferret. J Ultrastruct Res 33: 252.
129. Andrutis KA, Fox JG, Schauer DB, Marini RP, Li X, Yan L, Josenhans C, Suerbaum S (1997) Infection of the ferret stomach by isogenic flagellar mutant strains of Helicobacter mustelae. Infect Immun 65: 1962–1966.
130. Pfeiffer CJ, Roth JL (1970) Studies on the secretory and cytotoxic actions of caffeine on the ferret gastric mucosa. Exp Mol Pathol 13: 66–78.
131. Umeda N, Roth JL, Pfeiffer CJ (1971) Gastric lesions produced by cinchophen in the ferret. Toxicol Appl Pharmacol 18: 102–113.
132. Poddar S, Jacob S (1979) Mucosubstance histochemistry of Brunner's glands, pyloric glands and duodenal goblet cells in the ferret. Histochemistry 65: 67–81.
133. Pfeiffer CJ, Peters CM (1969) Gastric secretion in the chronic fistula ferret. Gastroenterology 57: 518–524.
134. Perciballi M, Conboy JJ, Hotchkiss JH (1989) Nitrite-cured meats as a source of endogenously and exogenously formed N-nitrosoproline in the ferret. Food Chem Toxicol 27: 111–116.
135. Roth SH, Schofield B, Yates JC (1979) Effects of atropine on secretion and motility in isolated gastric mucosa and attached muscularis externa from ferret and cat. J Physiol 292: 351–361.
136. Yates JC, Schofield B, Roth SH (1978) Acid secretion and motility of isolated mammalian gastric mucosa and attached muscularis externa. Am J Physiol 234: E319–E326.
138. Basso N, Passaro E, Jr. (1972) Effect of calcium on pentagastrin-, histamine-, bethanecol-, and insulin-stimulated gastric secretion in the ferret. J Surg Res 13: 32–38.
139. Desmond HP, Dockray GJ, Gregory RA, et al. (1984) Identification and characterization of a novel peptide generated during gastrin biosynthesis in the pig and ferret. J Physiol 354: 4.
140. Basso N, Umeda N, Roth JL, Passaro E, Jr., Pfeiffer CJ (1971) Effect of unilateral vagotomy on gastric secretion in the ferret. Gastroenterology 61: 207–212.
141. Jonsson AC, Dockray GJ (1984) Immunohistochemical localization to pyloric antral G cells of peptides derived from porcine preprogastrin. Regul Pept 8: 283–290.
142. Andrews PLR (1979) The vagal control of the gastrointestinal tract. PhD Thesis. Univ of Sheffield.
143. Basso N, Roth JL, Pfeiffer CJ (1970) Gastric secretion after pentagastrin and histamine in the basal secreting ferret. J Surg Res 10: 111–115.
144. Shillingford J, Lindup W, Parke D (1974) The effects of carbenoxolone on the biosynthesis of gastric glycoproteins in the rat and ferret. Biochem Soc Trans 2: 1104.
145. Bourke B, Sherman P, Drumm B (1994) Peptic ulcer disease: what is the role for Helicobacter pylori? Semin Gastrointest Dis 5: 24–31.
146. Fox JG, Wishnok JS, Murphy JC, Tannenbaum SR, Correa P (1993) MNNG-induced gastric carcinoma in ferrets infected with Helicobacter mustelae. Carcinogenesis 14: 1957–1961.
147. Fox JG, Otto G, Taylor NS, Rosenblad W, Murphy JC (1991) Helicobacter mustelae-induced gastritis and elevated gastric pH in the ferret (Mustela putorius furo). Infect Immun 59: 1875–1880.
148. Yu J, Russell RM, Salomon RN, Murphy JC, Palley LS, Fox JG (1995) Effect of Helicobacter mustelae infection on ferret gastric epithelial cell proliferation. Carcinogenesis 16: 1927–1931.
149. Huang JQ, Hunt RH (1997) Review: eradication of Helicobacter pylori: problems and recommendations. J Gastroenterol Hepatol 12: 590–598.
150. Grundy D, Scratcherd T (1982) Effect of stimulation of the vagus nerve in bursts on gastric acid secretion and motility in the anaesthetized ferret. J Physiol 333: 451–461.
151. Christensen J, Rick GA, Lowe LS (1992) Distributions of interstitial cells of Cajal in stomach and colon of cat, dog, ferret, opossum, rat, guinea pig and rabbit. J Auton Nerv Syst 37: 47–56.
152. Nagakura Y, Sohda K, Ito H, Miyata K (1997) Gastrointestinal motor activity in conscious ferrets. Eur J Pharmacol 321: 53–57.
153. Schwarz LA, Solano M, Manning A, Marini RP, Fox JG (2003) The normal upper gastrointestinal examination in the ferret. Vet Radiol Ultrasound 44: 165–172.
154. Mackay TW, Andrews PL (1983) A comparative study of the vagal innervation of the stomach in man and the ferret. J Anat 136: 449–481.
155. Andrews PL, Scratcherd T (1980) The gastric motility patterns induced by direct and reflex excitation of the vagus nerves in the anaesthetized ferret. J Physiol 302: 363–378.
156. Bingham S, Andrews PLR (1986) Preliminary studies on the mechansim of action of cisapride. Digestion 34: 138.
157. Andrews PLR, Wood KL (1984) The effects of chemical and mechanical stimuli applied to the gastric antral mucosa on corpus motility in the anesthetized ferret. J Physiol 348: 63.
158. Andrews PL, Grundy D, Scratcherd T (1980) Reflex excitation of antral motility induced by gastric distension in the ferret. J Physiol 298: 79–84.
159. Andrews PL, Grundy D, Scratcherd T (1980) Vagal afferent discharge from mechanoreceptors in different regions of the ferret stomach. J Physiol 298: 513–524.
160. Andrews PL, Grundy D, Lawes IN (1980) The role of the vagus and splanchnic nerves in the regulation of intragastric pressure in the ferret. J Physiol 307: 401–411.
161. Lingenfelser T, Blackshaw LA, Sun WM, Dent J (1997) Pyloric motor response to central and peripheral nitric oxide in the ferret. Neurogastroenterol Motil 9: 167–175.
162. Andrews PL, Bingham S (1990) Adaptation of the mechanisms controlling gastric motility following chronic vagotomy in the ferret. Exp Physiol 75: 811–825.
163. Andrews PL, Lawes IN (1984) Interactions between splanchnic and vagus nerves in the control of mean intragastric pressure in the ferret. J Physiol 351: 473–490.
164. Andrews PLR, Bingham S, Wood KI (1986) A comparison of the effects of vagotomy and GABA β receptor agonist on gastric corpus pressure in the ferret. J Physiol 378: 18.
165. Grundy D, Gharib-Naseri MK, Hutson D (1992) Effect of immunisation against vasoactive intestinal polypeptide on gastric corpus tone and motility in the ferret. Gut 33: 1473–1476.
166. Grundy D, Gharib-Naseri MK, Hutson D (1993) Plasticity in the gastric inhibitory innervation after immunization against VIP and vagotomy in the ferret. Am J Physiol 265: G432–G439.
167. Grundy D (1990) The effect of surgical anaesthesia on antral motility in the ferret. Exp Physiol 75: 701–708.
168. Olivier AK, Yi Y, Sun X, Sui H, Liang B, Hu S, Xie W, Fisher JT, Keiser NW, Lei D, Zhou W, Yan Z, Li G, Evans TI, Meyerholz DK, Wang K, Stewart ZA, Norris AW, Engelhardt JF (2012) Abnormal endocrine pancreas function at birth in cystic fibrosis ferrets. J Clin Invest 122: 3755–3768.
169. Beyerman HC, Grossman MI, Scratcherd T, Solomon TE, Voskamp D (1981) On the instability of secretin. Life Sci 29: 885–894.
170. Daniel PM, Henderson JR (1978) Circulation in the islets of Langerhans [proceedings]. J Physiol 275: 10P–11P.
171. Henderson JR (1969) Why are the islets of Langerhans? Lancet 2: 469–470.
172. Audy G, Kerly M (1952) The content of glycogenolytic factor in pancreas from different species. Biochem J 52: 77–78.
173. Sbarra V, Mas E, Henderson TR, Hamosh M, Lombardo D, Hamosh P (1996) Digestive lipases of the newborn ferret: compensatory role of milk bile salt-dependent lipase. Pediatr Res 40: 263–268.
174. Ellis LA, Hamosh M (1992) Bile salt stimulated lipase: comparative studies in ferret milk and lactating mammary gland. Lipids 27: 917–922.
175. Kang ES, Galloway MS, Ellis J, Solomon SS, Bean W, Reger JF, Cook GA, Olson G (1990) Hepatic steatosis during convalescence from influenza B infection in ferrets with postprandial hyperinsulinemia. J Lab Clin Med 116: 335–344.
176. Carpenter JW, Novilla MN (1977) Diabetes mellitus in a black-footed ferret. J Am Vet Med Assoc 171: 890–893.
177. Phair KA, Carpenter JW, Schermerhorn T, Ganta CK, DeBey BM (2011) Diabetic ketoacidosis with concurrent pancreatitis, pancreatic beta islet cell tumor, and adrenal disease in an obese ferret (Mustela putorius furo). J Am Assoc Lab Anim Sci 50: 531–535.
178. Boari A, Papa V, Di Silverio F, Aste G, Olivero D, Rocconi F (2010) Type 1 diabetes mellitus and hyperadrenocorticism in a ferret. Vet Res Commun 34(Suppl. 1): S107–S110.
179. Benoit-Biancamano MO, Morin M, Langlois I (2005) Histopathologic lesions of diabetes mellitus in a domestic ferret. Can Vet J 46: 895–897.
180. Poddar S (1977) Gross and microscopic anatomy of the biliary tract of the ferret. Acta Anat (Basel) 97: 121–131.
181. Mitan S, Greenwood B (1994) CCK-8 contracts the gallbladder and colon through different mechanisms in the ferret. Ann N Y Acad Sci 713: 401–403.
182. Ryden EB, Licht WR, Cabot EB, Fox JG (1990) Gastric nitrite processing in the surgically altered maximal and minimal bile reflux ferret model. Carcinogenesis 11: 405–409.
183. Scratcherd T, Syme G, Wynne R (1976) The release of cholecystokinin-pancreozymin from the isolated jejunum of the ferret. In: Case R, Goebell H, eds. Stimulus-secretion coupling in the gastrointestinal tract. Lancaster, UK: MTP Press, pp. 341.
184. Greenwood B, Doolittle T, See NA, Koch TR, Dodds WJ, Davison JS (1990) Effects of substance P and vasoactive intestinal polypeptide on contractile activity and epithelial transport in the ferret jejunum. Gastroenterology 98: 1509–1517.
185. Greenwood B, Davison JS (1985) Role of extrinsic and intrinsic nerves in the relationship between intestinal motility and transmural potential difference in the anesthetized ferret. Gastroenterology 89: 1286–1292.
186. Van de Perre P (2003) Transfer of antibody via mother's milk. Vaccine 21: 3374–3376.
187. Clarke RM, Hardy RN (1970) Structural changes in the small intestine associated with the uptake of polyvinyl pyrrolidone by the young ferret, rabbit, guinea-pig, cat and chicken. J Physiol 209: 669–687.
188. Carlile AE, Beck F (1983) Maturation in the ferret ileal epithelium and the effect of cortisone acetate. J Anat 136: 401–405.
189. Wang XD, Krinsky NI, Marini RP, Tang G, Yu J, Hurley R, Fox JG, Russell RM (1992) Intestinal uptake and lymphatic absorption of beta-carotene in ferrets: a model for human beta-carotene metabolism. Am J Physiol 263: G480–G486.
190. Hebuterne X, Wang XD, Smith DE, Tang G, Russell RM (1996) In vivo biosynthesis of retinoic acid from beta-carotene involves and excentric cleavage pathway in ferret intestine. J Lipid Res 37: 482–492.
191. Gugger ET, Bierer TL, Henze TM, White WS, Erdman JW, Jr. (1992) Beta-carotene uptake and tissue distribution in ferrets (Mustela putorius furo). J Nutr 122: 115–119.
192. Hore P, Messer M (1968) Studies on disaccharidase activities of the small intestine of the domestic cat and other carnivorous mammals. Comp Biochem Physiol 24: 717–725.
193. Oleinik VM (1995) Distribution of digestive enzyme activities along intestine in blue fox, mink, ferret and rat. Comp Biochem Physiol A Physiol 112: 55–58.
194. Blackshaw LA, Grundy D (1991) Locally and reflexly mediated effects of cholecystokinin-octapeptide on the ferret stomach. J Auton Nerv Syst 36: 129–137.
195. Collman PI, Grundy D, Scratcherd T (1983) Vagal influences on the jejunal “minute rhythm” in the anaesthetized ferret. J Physiol 345: 65–74.
196. Poddar S, Jacob S (1981) Mucosubstances in the colonic goblet cells of the ferret. Acta Histochem 68: 279–289.
197. Bueno L, Fioramonti J, More J (1981) Is there a functional large intestine in the ferret? Experientia 37: 275–277.
198. Collman PI, Grundy D, Scratcherd T (1984) Vagal control of colonic motility in the anaesthetized ferret: evidence for a non-cholinergic excitatory innervation. J Physiol 348: 35–42.
199. Scratcherd T, Grundy D, Collman PI (1986) Evidence for a non-cholinergic excitatory innervation in the control of colonic motility. Arch Int Pharmacodyn Ther 280: 164–175.
200. Bleavins MR, Aulerich RJ (1981) Feed consumption and food passage time in mink (Mustela vison) and European ferrets (Mustela putorius furo). Lab Anim Sci 31: 268–269.
201. Hein J, Spreyer F, Sauter-Louis C, Hartmann K (2012) Reference ranges for laboratory parameters in ferrets. Vet Rec 171: 218.
202. Ohwada K, Katahira K (1993) Reference values for organ weight, hematology and serum chemistry in the female ferret (Mustela putrius furo). Jikken Dobutsu 42: 135–142.
203. Marini RP, Jackson LR, Esteves MI, Andrutis KA, Goslant CM, Fox JG (1994) Effect of isoflurane on hematologic variables in ferrets. Am J Vet Res 55: 1479–1483.
204. Damgaard BM, Dalgaard TS, Larsen T, Hedemann MS, Hansen SW (2012) The effects of feed restriction on physical activity, body weight, physiology, haematology and immunology in female mink. Res Vet Sci 93: 936–942.
205. Manning DD, Bell JA (1990) Lack of detectable blood groups in domestic ferrets: implications for transfusion. J Am Vet Med Assoc 197: 84–86.
206. Hohenhaus AE (2004) Importance of blood groups and blood group antibodies in companion animals. Transfus Med Rev 18: 117–126.
207. Flatman PW, Andrews PL (1983) Cation and ATP content of ferret red cells. Comp Biochem Physiol A Comp Physiol 74: 939–943.
208. Flatman PW (1983) Sodium and potassium transport in ferret red cells. J Physiol 341: 545–557.
209. Hombrados I, Vidal Y, Rodewald K, Braunitzer G, Neuzil E (1989) Carnivora: the primary structure of the alpha-chains of ferret (Mustela putorius furo, Mustelidae) hemoglobins. Biol Chem Hoppe Seyler 370: 1133–1138.
210. Flatman PW (2005) Activation of ferret erythrocyte Na+-K+-2Cl- cotransport by deoxygenation. J Physiol 563: 421–431.
211. Schernthaner A, Lendl CE, Hartmann K, Pragst I, Preissel AK, Henke J (2011) Medetomidine/midazolam/ketamine anaesthesia in ferrets: effects on cardiorespiratory parameters and evaluation of plasma drug concentrations. Vet Anaesth Analg 38: 439–450.
212. Imai A, Steffey EP, Farver TB, Ilkiw JE (1999) Assessment of isoflurane-induced anesthesia in ferrets and rats. Am J Vet Res 60: 1577–1583.
213. Schumacher WA, Steinbacher TE, Megill JR, Durham SK (1996) A ferret model of electrical-induction of arterial thrombosis that is sensitive to aspirin. J Pharmacol Toxicol Methods 35: 3–10.
214. Takahashi S, Hirai N, Shirai M, Ito K, Asai F (2011) Comparison of the blood coagulation profiles of ferrets and rats. J Vet Med Sci 73: 953–956.
215. Kenyon AJ, Howard E, Buko L (1967) Hypergammaglobulinemia in ferrets with lymphoproliferative lesions (Aleutian disease). Am J Vet Res 28: 1167–1172.
216. Wallace MN, Roeda D, Harper MS (1997) Deoxyglucose uptake in the ferret auditory cortex. Exp Brain Res 117: 488–500.
217. Chen S (2010) Advanced diagnostic approaches and current medical management of insulinomas and adrenocortical disease in ferrets (Mustela putorius furo). Vet Clin North Am Exot Anim Pract 13: 439–452.
218. Weiss CA, Williams BH, Scott MV (1998) Insulinoma in the ferret: clinical findings and treatment comparison of 66 cases. J Am Anim Hosp Assoc 34: 471–475.
219. Cryer A, Sawyerr AM (1978) A comparison of the composition and apolipoprotein content of the lipoproteins isolated from human and ferret (Mustela putorius furo L.) serum. Comp Biochem Physiol B 61: 151–159.
220. Thomas PE, Deshmukh DR (1986) Effect of arginine-free diet on ammonia metabolism in young and adult ferrets. J Nutr 116: 545–551.
221. Deshmukh DR, Thomas PE (1985) Arginine deficiency, hyperammonemia and Reye's syndrome in ferrets. Lab Anim Sci 35: 242–245.
223. Lockard BI (1985) The forebrain of the ferret. Lab Anim Sci 35: 216–228.
224. Moore DR (1982) Late onset of hearing in the ferret. Brain Res 253: 309–311.
225. Morey AL, Carlile S (1990) Auditory brainstem of the ferret: maturation of the brainstem auditory evoked response. Brain Res Dev Brain Res 52: 279–288.
226. Henkel CK, Keiger CJ, Franklin SR, Brunso-Bechtold JK (2007) Development of banded afferent compartments in the inferior colliculus before onset of hearing in ferrets. Neuroscience 146: 225–235.
227. Sumner CJ, Palmer AR (2012) Auditory nerve fibre responses in the ferret. Eur J Neurosci 36: 2428–2439.
228. Yin P, Fritz JB, Shamma SA (2010) Do ferrets perceive relative pitch? J Acoust Soc Am 127: 1673–1680.
229. Guibal C, Baker GE (2009) Abnormal axons in the albino optic tract. Invest Ophthalmol Vis Sci 50: 5516–5521.
230. Stuart-Harris CH, Francis T (1938) Studies on the nasal histology of epidemic influenza virus infection in the ferret: II. The resistance of regenerating respiratory epithelium to reinfection and to physicochemical injury. J Exp Med 68: 803–812.
231. Reznik GK (1990) Comparative anatomy, physiology, and function of the upper respiratory tract. Environ Health Perspect 85: 171–176.
232. Funk KA, Frantz C, Dyke AS, Ryan PC, Jin H, Kemble G, Dixit R, Leininger JR (2010) Nasal turbinate enlargement due to cartilage and bone proliferation: a normal developmental finding in young ferrets. Toxicol Pathol 38: 1064–1069.
233. Dennis BJ, Kerr DI (1975) Olfactory bulb connections with basal rhinencephalon in the ferret: an evoked potential and neuroanatomical study. J Comp Neurol 159: 129–148.
234. Woodley SK, Baum MJ (2004) Differential activation of glomeruli in the ferret's main olfactory bulb by anal scent gland odours from males and females: an early step in mate identification. Eur J Neurosci 20: 1025–1032.
235. Katz LC, Crowley JC (2002) Development of cortical circuits: lessons from ocular dominance columns. Nat Rev Neurosci 3: 34–42.
236. Yates CA, Herbert J (1979) The effects of different photoperiods on circadian 5-HT rhythms in regional brain areas and their modulation by pinealectomy, melatonin and oestradiol. Brain Res 176: 311–326.
237. Carter DS (1981) Studies of pineal and hypothalamic 5-HT in the ferret. Brain Res 224: 95–104.
238. Fuller RW, Hemrick-Luecke SK (1983) Species differences in epinephrine concentration and norepinephrine N-methyltransferase activity in hypothalamus and brain stem. Comp Biochem Physiol C 74: 47–49.
239. Taylor A, Elias SA (1984) Interaction of periodontal and jaw elevator spindle afferents in the cerebellum: sensory calibration. Brain Behav Evol 25: 157–165.
240. Lou JS, Bloedel JR (1986) The responses of simultaneously recorded Purkinje cells to the perturbations of the step cycle in the walking ferret: a study using a new analytical method: the real-time postsynaptic response (RTPR). Brain Res 365: 340–344.
241. Eidelberg E, Staten E, Watkins JC, McGraw D, McFadden C (1976) A model of spinal cord injury. Surg Neurol 6: 35–38.
242. Kearney PA, Ridella SA, Viano DC, Anderson TE (1988) Interaction of contact velocity and cord compression in determining the severity of spinal cord injury. J Neurotrauma 5: 187–208.
243. Jirenhed DA, Hesslow G (2011) Time course of classically conditioned Purkinje cell response is determined by initial part of conditioned stimulus. J Neurosci 31: 9070–9074.
244. Odekunle A, Bower AJ (1985) Brainstem connections of vagal afferent nerves in the ferret: an autoradiographic study. J Anat 140(Pt 3): 461–469.
245. Ringkamp M, Johanek LM, Borzan J, Hartke TV, Wu G, Pogatzki-Zahn EM, Campbell JN, Shim B, Schepers RJ, Meyer RA (2010) Conduction properties distinguish unmyelinated sympathetic efferent fibers and unmyelinated primary afferent fibers in the monkey. PLoS ONE 5: e9076.
246. Andrews PL, Fussey IV, Scratcherd T (1980) The spontaneous discharge in abdominal vagal efferents in the dog and ferret. Pflugers Arch 387: 55–60.
247. Blackshaw L, Grundy D, Scratcherd T (1986) Responses of vagal effrent units modulated by gastrointestinal stimuli to central vagal stimulation. J Physiol 378: 15.
248. Andrews P, Davis C, Grahame-Smith D, et al. (1987) Increase in [3H]-2-deoxyglucose uptake in the ferret arae postrema produced by apopmorphine administration or electrical stimulation of the abdominal vagus. J Physiol 382: 187.
249. Northcutt RG, Kaas JH (1995) The emergence and evolution of mammalian neocortex. Trends Neurosci 18: 373–379.
250. Rowell JJ, Mallik AK, Dugas-Ford J, Ragsdale CW (2010) Molecular analysis of neocortical layer structure in the ferret. J Comp Neurol 518: 3272–3289.
251. Neal J, Takahashi M, Silva M, Tiao G, Walsh CA, Sheen VL (2007) Insights into the gyrification of developing ferret brain by magnetic resonance imaging. J Anat 210: 66–77.
252. Majkowski J, Lee MH, Kozlowski PB, Haddad R (1984) EEG and seizure threshold in normal and lissencephalic ferrets. Brain Res 307: 29–38.
253. Deshmukh DR, Shope TC (1983) Arginine requirement and ammonia toxicity in ferrets. J Nutr 113: 1664–1667.
254. Rarey KE, Rush NL, Davis JA, Deshmukh DR (1984) Altered auditory brainstem evoked responses in the ferret model for Reye's syndrome. Int J Pediatr Otorhinolaryngol 7: 221–228.
255. Brauer RW, Beaver RW, Lahser S, McCall RD, Venters R (1979) Comparative physiology of the high-pressure neurological syndrome: compression rate effects. J Appl Physiol 46: 128–135.
256. Cameron AR, Coburn RF (1984) Electrical and anatomic characteristics of cells of ferret paratracheal ganglion. Am J Physiol 246: C450–C458.
257. Baker DG, McDonald DM, Basbaum CB, Mitchell RA (1986) The architecture of nerves and ganglia of the ferret trachea as revealed by acetylcholinesterase histochemistry. J Comp Neurol 246: 513–526.
258. Conlon K, Collins T, Kidd C (1998) The role of nitric oxide in the control by the vagal nerves of the heart of the ferret. Exp Physiol 83: 469–480.
259. Young RL, Cooper NJ, Blackshaw LA (2008) Anatomy and function of group III metabotropic glutamate receptors in gastric vagal pathways. Neuropharmacology 54: 965–975.
260. Andrews P, Harris P, Mackay T (1984) Recordings of neural activity in the direct renal nerve of the anesthetized ferret. J Physiol 346: 11.
261. Elliott TR (1906) The innervation of the bladder and uretha. J Physiol 35: 367.
262. Sann H, Hoppe S, Baldwin L, Grundy D, Schemann M (1998) Presence of putative neurotransmitters in the myenteric plexus of the gastrointestinal tract and in the musculature of the urinary bladder of the ferret. Neurogastroenterol Motil 10: 35–47.
263. Jafri MS, Weinreich D (1996) Substance P hyperpolarizes vagal sensory neurones of the ferret. J Physiol 493(Pt 1): 157–166.
264. Baker DG, Basbaum CB, Herbert DA, Mitchell RA (1983) Transmission in airway ganglia of ferrets: inhibition by norepinephrine. Neurosci Lett 41: 139–143.
265. Skoogh BE, Holtzman MJ, Sheller JR, Nadel JA (1982) Barbiturates depress vagal motor pathway to ferret trachea at ganglia. J Appl Physiol 53: 253–257.
266. Dey RD, Altemus JB, Rodd A, Mayer B, Said SI, Coburn RF (1996) Neurochemical characterization of intrinsic neurons in ferret tracheal plexus. Am J Respir Cell Mol Biol 14: 207–216.
267. Haxhiu MA, Yamamoto B, Dreshaj IA, Bedol D, Ferguson DG (2000) Involvement of glutamate in transmission of afferent constrictive inputs from the airways to the nucleus tractus solitarius in ferrets. J Auton Nerv Syst 80: 22–30.
268. Ferguson DG, Haxhiu MA, To AJ, Erokwu B, Dreshaj IA (2000) The alpha3 subtype of the nicotinic acetylcholine receptor is expressed in airway-related neurons of the nucleus tractus solitarius, but is not essential for reflex bronchoconstriction in ferrets. Neurosci Lett 287: 141–145.
269. Blackshaw L, Grundy D, Scratcherd T (1986) Discharge characteristics of gastric mechanoreceptors during contraction and relaxation of the ferret corpus. J Physiol 378: 17.
270. Fitzakerley JL, Lucier GE (1988) Connections of a vagal communicating branch in the ferret: I. Pathways and cell body location. Brain Res Bull 20: 189–196.
271. Fitzakerley JL, Lucier GE (1988) Connections of a vagal communicating branch in the ferret: II. Central projections. Brain Res Bull 20: 479–486.
272. Knox AP, Strominger NL, Battles AH, Carpenter DO (1994) The central connections of the vagus nerve in the ferret. Brain Res Bull 33: 49–63.
273. Strominger NL, Knox AP, Carpenter DO (1994) The connectivity of the area postrema in the ferret. Brain Res Bull 33: 33–47.
274. Boissonade FM, Davison JS, Egizii R, Lucier GE, Sharkey KA (1996) The dorsal vagal complex of the ferret: anatomical and immunohistochemical studies. Neurogastroenterol Motil 8: 255–272.
275. Reynolds DJ, Barber NA, Grahame-Smith DG, Leslie RA (1991) Cisplatin-evoked induction of c-Fos protein in the brainstem of the ferret: the effect of cervical vagotomy and the anti-emetic 5-HT3 receptor antagonist granisetron (BRL 43694). Brain Res 565: 231–236.
276. Boissonade FM, Davison JS (1996) Effect of vagal and splanchnic nerve section on Fos expression in ferret brain stem after emetic stimuli. Am J Physiol 271: R228–R236.
277. Rieger D, Murphy BD (1977) Episodic fluctuation in plasma testosterone and dihydrotestosterone in male ferrets during the breeding season. J Reprod Fertil 51: 511–514.
278. Baum MJ, Goldfoot DA (1975) Effect of amygdaloid lesions on gonadal maturation in male and female ferrets. Am J Physiol 228: 1646–1651.
279. Neal J, Murphy BD, Moger WH, Oliphant LW (1977) Reproduction in the male ferret: gonadal activity during the annual cycle; recrudescence and maturation. Biol Reprod 17: 380–385.
280. Erskine MS, Baum MJ (1982) Plasma concentrations of testosterone and dihydrotestosterone during perinatal development in male and female ferrets. Endocrinology 111: 767–772.
281. Batista-Arteaga M, Suarez-Bonnet A, Santana M, Nino T, Reyes R, Alamo D (2011) Testicular neoplasms (interstitial and Sertoli cell tumours) in a domestic ferret (Mustela putorius furo). Reprod Domest Anim 46: 177–180.
282. Heap RB, Hammond J, Jr. (1974) Plasma progesterone levels in pregnant and pseudopregnant ferrets. J Reprod Fertil 39: 149–152.
283. Daniel JC, Jr. (1976) Plasma progesterone levels before and at the time of implantation in the ferret. J Reprod Fertil 48: 437–438.
284. Donovan BT, Matson C, Kilpatrick MJ (1983) Effect of exposure to long days on the secretion of oestradiol, oestrone, progesterone, testosterone, androstenedione, cortisol and follicle-stimulating hormone in intact and spayed ferrets. J Endocrinol 99: 361–368.
285. Prohaczik A, Kulcsar M, Glavits R, Driancourt MA, Huszenicza GY (2011) Reproductive features and faecal progesterone metabolite profile in female ferrets. Reprod Domest Anim 46: e54–e61.
286. Gaunt R, Remington J, Edelmann A (1939) Effect of progesterone and other hormones on liver glycogen. Proc Soc Exp Biol Med 41: 429.
287. Wagner RA, Dorn DP (1994) Evaluation of serum estradiol concentrations in alopecic ferrets with adrenal gland tumors. J Am Vet Med Assoc 205: 703–707.
288. Williams ES, Thorne ET, Kwiatkowski DR, Lutz K, Anderson SL (1992) Comparative vaginal cytology of the estrous cycle of black-footed ferrets (Mustela nigripes), Siberian polecats (M. eversmanni), and domestic ferrets (M. putorius furo). J Vet Diagn Invest 4: 38–44.
289. Donovan BT, ter Haar MB (1977) Effects of luteinizing hormone releasing hormone on plasma follicle-stimulating hormone and luteinizing hormone levels in the ferret. J Endocrinol 73: 37–52.
290. Ryan KD, Siegel SF, Robinson SL (1985) Influence of day length and endocrine status on luteinizing hormone secretion in intact and ovariectomized adult ferrets. Biol Reprod 33: 690–697.
291. Donovan BT, ter Haar MB (1977) Stimulation of the hypothalamus and FSH and LH secretion in the ferret. Neuroendocrinology 23: 268–278.
292. Donovan BT, Gledhill B (1984) Half-life of FSH and LH in the ferret. Acta Endocrinol (Copenh) 105: 14–18.
293. Berglund LA, Sisk CL (1990) Pituitary responsiveness to luteinizing hormone-releasing hormone in prepubertal and postpubertal male ferrets. Biol Reprod 43: 335–339.
294. Sisk CL (1990) Photoperiodic regulation of gonadal growth and pulsatile luteinizing hormone secretion in male ferrets. J Biol Rhythms 5: 177–186.
295. Holmes RL (1961) The adrenal glands of the ferret, Mustela putorius. J Anat 95: 325–336.
296. Heard DJ, Collins B, Chen DL, Coniglario J (1990) Thyroid and adrenal function tests in adult male ferrets. Am J Vet Res 51: 32–35.
297. Bielinska M, Kiiveri S, Parviainen H, Mannisto S, Heikinheimo M, Wilson DB (2006) Gonadectomy-induced adrenocortical neoplasia in the domestic ferret (Mustela putorius furo) and laboratory mouse. Vet Pathol 43: 97–117.
298. Simone-Freilicher E (2008) Adrenal gland disease in ferrets. Vet Clin North Am Exot Anim Pract 11: 125–137, vii.
299. Wagner S, Kiupel M, Peterson RA, 2nd, Heikinheimo M, Wilson DB (2008) Cytochrome b5 expression in gonadectomy-induced adrenocortical neoplasms of the domestic ferret (Mustela putorius furo). Vet Pathol 45: 439–442.
300. Rosenthal KL (1997) Adrenal gland disease in ferrets. Vet Clin North Am Small Anim Pract 27: 401–418.
301. Gould WJ, Reimers TJ, Bell JA, Lawrence HJ, Randolph JF, Rowland PH, Scarlett JM (1995) Evaluation of urinary cortisol: creatinine ratios for the diagnosis of hyperadrenocorticism associated with adrenal gland tumors in ferrets. J Am Vet Med Assoc 206: 42–46.
302. Garibaldi BA, Pecquet Goad ME, Fox JG, Sylvina TJ, Murray R (1988) Serum cortisol radioimmunoassay values in the normal ferret and response to ACTH stimulation and dexamethasone suppression tests. Lab Anim Sci 38: 452–454.
303. Ackermann J, Carpenter JW, Godshalk CP, Harms CA (1994) Ultrasonographic detection of adrenal gland tumors in two ferrets. J Am Vet Med Assoc 205: 1001–1003.
304. Hawthorn J, Andrews PL, Ang VT, Jenkins JS (1988) Differential release of vasopressin and oxytocin in response to abdominal vagal afferent stimulation or apomorphine in the ferret. Brain Res 438: 193–198.
305. Mohanty B, Takahara H, Tachibana T, Naik DR, Nogami H (1993) Light- and electron-microscopic immunocytochemistry of somatotropes in the anterior pituitary gland of European ferret, Mustela putorius furo. Cell Tissue Res 273: 427–434.
306. Esteves MI, Marini RP, Ryden EB, Murphy JC, Fox JG (1994) Estimation of glomerular filtration rate and evaluation of renal function in ferrets (Mustela putorius furo). Am J Vet Res 55: 166–172.
307. Moffat DB, Fourman J (1964) Ectopic glomeruli in the human and animal kidney. Anat Rec 149: 1–7.
308. Andrews P, Harris P, Mackay T (1982) Phsyiological and anatomical studies of the innervation of the ferret kidney. J Physiol 330: 90.
309. Szenasi G, Bencsath P, Szalay L, Takacs L (1985) Fasting induces denervation natriuresis in the conscious rat. Am J Physiol 249: F753–F758.
310. Eshar D, Wyre NR, Brown DC (2012) Urine specific gravity values in clinically healthy young pet ferrets (Mustela furo). J Small Anim Pract 53: 115–119.
311. Elliott T (1905) The action of adrenalin. J Physiol 32: 401.
312. Andrews P, Tansey E (1987) The reaction of the ferret bladder to adrenalin: a modern demonstration of an experiment by TR Elliott and HH Dale. J Physiol 382: 5.
313. Moss HE, Burnstock G (1985) A comparative study of electrical field stimulation of the guinea-pig, ferret and marmoset urinary bladder. Eur J Pharmacol 114: 311–316.
314. Takeda H, Igawa Y, Komatsu Y, Yamazaki Y, Akahane M, Nishizawa O, Ajisawa Y (2000) Characterization of beta-adrenoceptor subtypes in the ferret urinary bladder in vitro and in vivo. Eur J Pharmacol 403: 147–155.
315. Tanaka N, Tamai T, Mukaiyama H, Hirabayashi A, Muranaka H, Ishikawa T, Kobayashi J, Akahane S, Akahane M (2003) Relationship between stereochemistry and the beta3-adrenoceptor agonistic activity of 4′-hydroxynorephedrine derivative as an agent for treatment of frequent urination and urinary incontinence. J Med Chem 46: 105–112.