Chapter 5
The Mystery of Morphogenesis
The unsolved problem of morphogenesis
As plants and animals develop from fertilized eggs, their form and organization become increasingly complex. How this happens is still a mystery.
From a materialistic point of view, the source of an organism’s form must already be present within the fertilized egg as a material structure. This theory was first elaborated in the seventeenth century in the doctrine of preformation. Preformationists maintained that the egg contained a tiny version of the adult organism, which then grew and unfolded. This theory turned out to be wrong. Preformationism was then resurrected in a subtle form in the late nineteenth-century doctrine of the germ-plasm (see below). In its modern form, the germ-plasm is identified with the chemical genes. The debate now centres on the question of whether or not the genes do in fact control and direct the processes of morphogenesis. Are genes alone sufficient? Or does development depend on non-material organizing principles as well? And if so, what are they, and how do they work?
In this chapter, we trace the history of this long debate and examine the forms in which it is still going on. It has been characterized again and again by mechanists’ denial of the existence of the purposive organizing principles of vitalism; and then by the reinvention of these principles in new guises such as the germ-plasm, selfish genes, genetic programs, patterns of information, and so on.
The discussion in this chapter enables the idea of morphogenetic fields to be seen in its biological context; in the following chapter we explore the nature of these fields.
Organisms are not preformed
According to the preformationists, fertilized eggs contained miniature organisms. Development was nothing but a growth and unfolding of these pre-existing material structures. This hypothetical process was called ‘evolution’, which literally means ‘unfolding’ or ‘unrolling’.
Preformationists in the seventeenth and eighteenth centuries differed among themselves as to whether these tiny organisms were contributed by the egg or the sperm; most favoured the sperm. Some thought they had proved it; they saw just what they were looking for. For example, one microscopist observed miniature horses in horse sperm, and similar tiny animals with big ears in the sperm of donkeys.1 Likewise, with the eye of faith, human sperm could be seen to contain tiny homunculi (Fig. 5.1).

Figure 5.1 A human sperm containing a homunculus, as seen by an early eighteenth-century microscopist. (From Cole, 1930)
Although this theory gave a satisfyingly simple explanation for the development of individual organisms, it ran into appalling theoretical difficulties when the succession of generations was considered. If a rabbit, for example, grows from a miniature rabbit in a fertilized egg, then it itself must contain miniature rabbits in its germ cells, and they must in turn contain nested within them an endless series of future generations. An eighteenth-century opponent of preformationism calculated that at least 10100,000 rabbits must have existed in the first rabbit, assuming that creation took place 6,000 years ago and that rabbits begin to breed at six months of age.2
In due course, the theory of preformation was completely refuted by empirical facts. When developing embryos were closely observed, new structures could be seen to appear that were not there before. For example, in 1768 Caspar Friedrich Wolff showed that in chick embryos ‘the intestine is formed by the folding back of a sheet of tissue which is detached from the ventral surface of the embryo, and that the folds produce a gutter which in the course of time transforms itself into a closed tube.’3 By the mid-nineteenth century, as the further study of embryology provided overwhelming evidence for it, development was generally agreed to be epigenetic, in other words to involve the appearance of material structures that were not present before (Fig. 5.2).

Figure 5.2 Stages in the embryonic development of a sea-squirt. (From Russell, 1916)
Epigenesis is just what would be expected from the point of view of Platonic and Aristotelian theories, which have never supposed that all of an organism’s form must be contained in the matter of the fertilized egg. By contrast, epigenesis is inevitably problematic from the mechanistic point of view. Somehow, more material form has to arise from less. Embryos have to pull themselves up by their own material bootstraps, as it were.
The same problem is presented by the phenomenon of regeneration, in which more complex forms can arise from less complex ones, as in embryonic development: a whole willow tree, for example, can regenerate from a small cutting.
The regeneration of wholeness
According to the preformationists, the organism developed by a kind of inflation of its original miniaturized form. But if that were so, how could it regenerate parts of itself that were lost? For a crude analogy, think of an inflatable rubber doll: how could it possibly regenerate an arm that was cut off?
If, on the other hand, various parts of organisms have the capacity to regenerate, then might not this same capacity account for their development in the first place? Nicolas Hartsoeker put it tersely in 1722: ‘An intelligence that can reproduce the lost claw of a crayfish can produce the entire animal.’4
The capacity to regenerate is in fact one of the most fundamental features of living organisms, and any theory of life has to try to explain it. All organisms have some regenerative powers, even if only when young or only in certain tissues. For example, we ourselves are continuously regenerating our blood, our intestinal lining, our skin; our wounds heal; broken bones knit themselves together; severed nerves grow out again; and if part of the liver is lost, new liver tissue develops to replace it.5 Many of the lower animals have such a strong regenerative ability that they can duplicate whole creatures from isolated parts. A flatworm, for example, can be cut into pieces, and each piece – a head, a tail, a side, or a mere slice – can grow into a complete worm (Fig. 5.3). And many plants can form new plants from separated parts: thousands of cuttings can be taken from a willow tree, each of which can grow into a new willow.
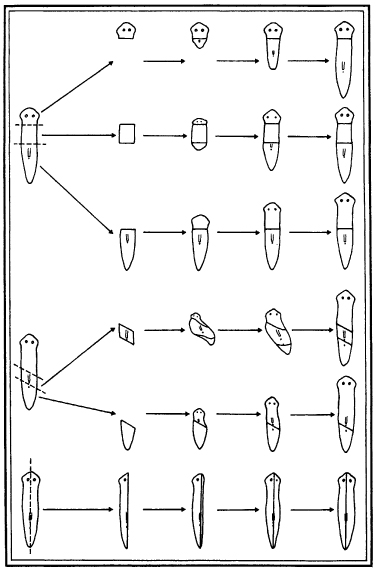
Figure 5.3 The regeneration of complete flatworms from pieces of worm (of the genus Planaria), cut as indicated on the left. (After Morgan, 1901)
Regeneration reveals that in some sense organisms have a wholeness that is more than the sum of their parts; parts can be removed, and yet wholeness can be restored. A part of a flatworm is more than just a part; it has an implicit wholeness that goes beyond its actual material structure: if it is isolated from the rest of the worm, it can turn into a whole worm. The capacity to regenerate is one of the most striking ways in which living organisms differ from machines. If a computer, for example, is cut into pieces, they do not grow into new computers. They remain pieces of a broken computer. The same goes for cars, telephones, and any other kind of machinery.
Nevertheless, there are some physical systems with holistic properties that survive the removal of parts. If an iron magnet, for example, is cut up, each part is a whole magnet with a complete magnetic field. Or if a part is removed from a hologram, which is a physical record of interference patterns in the electromagnetic field, this part can give rise to the entire original image.
Such physical analogies to the holistic properties of living organisms are examples of field phenomena. Fields are not material objects, but regions of influence. Do the epigenetic development and the regenerative capacities of living organisms depend on fields, or something like fields, with which they are associated? Or are they due to material objects that are present in the egg to start with?
We now follow this debate through another turn of the spiral in the theory of the germ-plasm and the vitalist reply to it in Hans Driesch’s theory of entelechy.
The germ-plasm
Preformationism in its original form had to be abandoned; the theory was proved to be false. It was resurrected in a subtle form in the 1880s by August Weismann, who proposed that fertilized eggs contain material structures that, although they do not have the actual form of the adult organism, somehow give rise to it. These material structures were in what Weismann called the germ-plasm.
Weismann made a theoretical division of organisms into two quite different parts, the body, or somatoplasm, and the germ-plasm. He described the germ-plasm as a ‘highly complex structure’ with the ‘power of developing into a complex organism.’6 This is the repository of all the specific causes of form observed in the adult organism: each particular part of the organism is caused by a particulate material unit, called a determinant.
By contrast, the somatoplasm is that part of the organism that is shaped and moulded by the germ-plasm. The germ-plasm is the active agent, and the somatoplasm responds passively to it. The germ-plasm affects the somatoplasm, but not vice versa. This process is represented in the familiar scheme in Figure 5.4, which emphasizes the potential immortality of the germ-plasm, and the mortality of the organisms to which it gives rise.
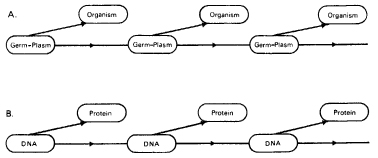
Figure 5.4 A: Weismann’s scheme, indicating the continuity of the germ-plasm from generation to generation, and the transient nature of the adult organisms. B: The ‘central dogma’ of molecular biology, in which Weismann’s scheme is interpreted in terms of DNA and proteins.
In animals, the embryonic germ cells are separated relatively early from the rest of the body, and Weismann supposed that there is no transfer of information from the body to the germ cells; he believed there could be no modification of the germ-plasm as a result of what happens in the body. Therefore organisms inherit only the inherited characteristics of their ancestors; they cannot inherit any characteristics that their ancestors acquired for themselves through adaptation to the environment or the development of new habits. We return to the question of the ‘Lamarckian’ inheritance of acquired characteristics in Chapter 8 and consider some of the evidence in its favour. But according to Weismann, such inheritance was impossible.
In plants, unlike animals, the germ cells are not set aside from the rest of the body at an early stage of development. Pollen and egg cells are formed in flower buds, which arise on the growing shoots. Nevertheless, Weismann’s theoretical principles were (and still are) assumed to apply to plants as well, in spite of their fundamental difference from animals.
Weismann believed that the determinants for each part of the body were parcelled out to various embryonic tissues in the course of development. Each received not a miniature version of the adult structure, as in old-style preformationism; instead the material structure of the determinants somehow ‘directed’ the formation of the adult structures.
This theory was taken up by the embryologist Wilhelm Roux, the founder of the school of ‘developmental mechanics.’ Roux postulated that the early embryo is like a mosaic, containing parts that develop independently, although in harmony with each other.
This theory soon turned out to be inadequate. In the 1890s, the embryologist Hans Driesch discovered that when half a young sea-urchin embryo was destroyed, the remaining half did not give rise to half a sea-urchin, as it would have done if the embryo were a mosaic of more or less independent parts; rather, it adjusted to the loss and went on to form a small but complete organism. Conversely, Driesch showed that if two young embryos were artificially fused together, they produced not a double sea-urchin, but a normal single one.7
This ability of embryos to adjust to damage is called regulation. It is closely related to regeneration. Many other examples of embryonic regulation have been discovered since the time of Driesch, not only in very young embryos, but in the developing organs of older embryos as well. For example, the developing wing buds of chick embryos have the ability to adjust themselves and produce normal wings in spite of quite severe damage.
Driesch pointed out that this ability of embryos to regulate shows that their different parts do not develop independently in a rigidly predetermined way. Rather, they respond and adjust to each other. They can change the way they develop if other parts are damaged; cells that would have given rise to one particular structure in a normal embryo can give rise to another one instead if need be. Thus Driesch refuted Weismann’s original theory of determinants parcelled out progressively into the developing embryonic tissues.
Towards the end of the nineteenth century, as a result of studies of chromosomes (threadlike structures within the cell nuclei), the cell nucleus was generally recognized to contain the material substance of heredity. Weismann consequently identified the germ-plasm with the chromosomes.8 With the rediscovery in 1900 of the findings of Gregor Mendel and the subsequent development of genetics, Weismann’s determinants were identified with the genes. Then with the discovery of the structure of the genetic material, DNA, and the way it ‘codes’ for the sequence of amino acids in proteins, the germ-plasm theory seemed to be fully confirmed. Well-defined biochemical substances could be written into Weismann’s scheme for the germ-plasm and somatoplasm: DNA and protein (Fig. 5.4). This scheme represents what is known as the central dogma of molecular biology: the genetic material acts as a template for the synthesis of proteins, but never the reverse. As in Weismann’s original scheme, this rules out on theoretical grounds the possibility of the inheritance of acquired characteristics. Neither the form, function, nor behaviour of the body can exert any specific influence on the genetic constitution, or genotype.
Darwin’s theory of evolution accepted the inheritance of acquired characteristics, and Darwin himself in his theory of panspermia proposed that the germ cells were modified through the incorporation of ‘gemmules’ from different parts of the body.9 The neo-Darwinian theory differs from Darwin’s in that it is firmly based on Weismannian principles. These are incorporated into the distinction between the genotype, or genetic constitution, on the one hand and the phenotype, or the organism as it actually appears, on the other. It is the genotype that evolves, and it is the genotype that determines the phenotype. Consequently, in principle, ‘a theory of development would effectively enable one to compute the adult organism from genetic information in the egg’ (L. Wolpert and J. Lewis).10
This statement was published in 1975, before it was possible to characterize in molecular detail the ‘genetic information in the egg’. How has this idea fared since then? By 2009, dozens of species’ genomes had been fully worked out, including the human genome. But it was still impossible to ‘compute the embryo,’ and Lewis Wolpert admitted in 2009 that ‘given the genome of the egg we cannot predict the way the embryo will develop’. But given vastly more information and enormous computing power he still believed, ‘We would, given a fertilized human egg, be able to have a picture of all the details of the newborn baby, including any abnormalities. We would also be able to program the egg to develop into any shape we desire. The time will come when this is possible.’11 This is a ringing statement of faith in the role of the genes and their explanatory power. However, in a public debate at Cambridge University in 2009 between Wolpert and myself, when I asked him how long it might take for this prediction to be fulfilled, he replied: ‘At least a century.’
I later challenged him to a wager about the predictive power of the genome in less complex organisms, with a twenty-year time span. He agreed. At stake is a case of fine port for which we paid half each, Quinta do Vesuvio 2005, being stored in the cellars of the Wine Society, near London. The wager was published in the New Scientist in July 2009, as follows:12
By 1 May 2029, given the genome of a fertilized egg of an animal or plant, we will be able to predict in at least one case all the details of the organism that develops from it, including any abnormalities.
Wolpert bets that this will happen. I bet that it will not. If the outcome is not obvious, the Royal Society will be asked to adjudicate.
Entelechy
In accounts written by contemporary biologists, vitalism is usually treated as if it were a kind of superstition that has been swept away by the advance of rational understanding.13 Its discrediting is usually said to have begun with the first artificial synthesis of an organic chemical, urea, by Friedrich Wöhler in 1828, and to have proceeded faster and faster ever since. Jacques Monod expressed the conventional view as follows:
Developments in molecular biology over the past two decades have singularly narrowed the domain of the mysterious, leaving little open to vitalist speculation but the field of subjectivity: that of consciousness itself. There is no great risk in predicting that also in this area, for the time being still ‘reserved,’ such speculation will prove as sterile as in all the others where it has been practised up to now.14
As history of science, this is naïve. And it misses the main point. Most nineteenth-century vitalists did not deny that living organisms contain organic chemicals that could be analysed and synthesized artificially; for example, the great chemist Justus von Liebig argued that in spite of the fact that many organic substances could be synthesized in the laboratory, and that many more would be made in the future, chemistry would never be in a position to create an eye or a leaf. These, he believed, were due to a kind of cause that organized the chemicals into ‘new forms so that they gain new qualities, forms and qualities which do not appear except in the organism.’15
Vague vitalist ideas such as this were common throughout the nineteenth century. It was only in the 1900s that Driesch worked out a vitalist theory in some detail. He started his career in the school of developmental mechanics, but he came to the conclusion that the embryonic regulation, regeneration, and reproduction show that something with an inherent wholeness acts on the living system, but is not a material part of it. Following Aristotle, he called this non-material causal factor entelechy, from en = in and telos = end or goal: that which has the end in itself, self-fulfillment. Driesch thought of entelechy as purposive or teleological, directing physical processes under its influence towards goals contained within itself.16
According to Driesch, entelechy guides the morphogenesis of the developing organism towards the characteristic form of its species. The genes are responsible for providing the material means for morphogenesis, the chemical substances to be ordered, but the ordering itself is due to entelechy. Similarly, the nervous system is the means for the behaviour of an animal, but entelechy organizes its activity, using it like an instrument, as a pianist plays a piano. Behaviour can be affected by damage to the brain, just as the music played by the pianist is affected by damage to the piano; but this only proves that the brain is a necessary means for behaviour, as the piano is a necessary means for the pianist.
Because entelechy ‘contains’ the end towards which a process under its control is directed, if a normal pattern of development is disturbed, the organism can reach the same goal in a different way: it can regulate or regenerate.
Driesch proposed that development and behaviour are under the control of a hierarchy of entelechies, all derived from and ultimately subordinate to the overall entelechy of the organism. He regarded these entelechies as natural causal factors, not ‘metaphysical’ or ‘mystical’ entities, that act on physical and chemical processes, imposing order and pattern on changes that would otherwise be indeterminate. But he developed his theory when science was still under the sway of classical physics; it was generally believed that all physical processes were fully deterministic and in principle completely predictable. In this case, there would be no scope for the action of entelechy, because everything would already be fully determined by the laws of physics.
Thus, Driesch suggested, for entelechy to be able to impose order on processes within living organisms, these processes must be physically indeterminate, at least on a microscopic scale. Since such indeterminism was not admitted by the physics of his day, Driesch was driven to suppose that entelechy itself introduced this indeterminism into organisms. He suggested that it did so by affecting the timing of physico-chemical processes. It suspended them, and then released them again as required for its purposes.17
This proposal seemed to be a fatal weakness in Driesch’s system. In the context of the reigning orthodoxy, any interference with physical determinism was unthinkable, and Driesch’s proposal seemed impossible in principle.
It is surely ironic that by the end of the 1920s, when vitalism seemed discredited, undreamt-of changes were occurring in physics. Heisenberg proposed the uncertainty principle in 1927, and as quantum theory developed it became clear that on the microscopic scale physical events are not fully determinate, but predictable only in terms of probabilities, statistically. It was no longer necessary for entelechy to introduce indeterminism into living organisms: indeterminism is inherent in their physical nature anyway.
Driesch agreed that some aspects of living organisms could be explained mechanistically; he was aware of the importance of enzymes and other proteins, and thought that genes would ultimately be understood in chemical terms. These opinions have been confirmed by subsequent discoveries. But he argued that development and behaviour would never be fully understood in mechanistic terms, but would only be comprehensible in terms of purposive organizing principles. At least so far, this prediction seems valid. Very little is actually understood about morphogenesis in physical and chemical terms; and the organizing principles of vitalism, which were denied by the mechanistic theory, have returned in such guises as selfish genes and genetic programs. The central paradigm of modern biology is genetic vitalism.
Selfish genes
Weismann’s germ-plasm was supposed to have a more or less immutable structure, which determined the form of the organism. His germ-plasm/somatoplasm duality, like its descendant, the genotype/phenotype dichotomy, recalls the Platonic distinction between the changeless Form or Idea and the phenomenon in which it is reflected. Just as the phenomenon has no effect on the Idea, so the phenotype has no effect on the genotype. It is as if Weismann incarnated the Idea of the organism into the germ-plasm, and also endowed it with the controlling and organizing properties of the psyche or entelechy; Weismann thought it was a ‘central directing agency.’18
At the same time his notion that each ‘determinant’ in the germ-plasm is responsible for a particular physical characteristic was atomistic in spirit. It is paralleled by the more recent notion that particular characteristics are determined by particular genes or sets of genes. In other words, inherited characteristics – for example the shape of a pigeon’s foot, or its inborn homing ability – are genetically determined: there are foot-shape genes and homing genes. Or again in other words, there are genes ‘for’ foot shape and genes ‘for’ homing.
This idea of genes ‘for’ plays a central role in neo-Darwinian evolutionary theory. Genes are in competition, and some genes are more successful than others in propagating copies of themselves. Genes ‘for’ characteristics that favour more copies of these genes surviving and reproducing will increase in frequency within the interbreeding population through natural selection. Genes ‘for’ unfavourable characteristics will be selected against; they will decrease in frequency. The rates of change of these frequencies can be calculated mathematically by theoretical population geneticists. But in order to formulate the appropriate equations, it is necessary to make some simplifying assumptions. One is the Weismannian view of genes as independent determinants that can be selected more or less independently of each other.
This assumption underlies neo-Darwinian thinking and is carried to its furthest extreme in the school of sociobiology and evolutionary psychology, which attempt to account for practically all aspects of animal behaviour and social life in terms of genetic determinants whose frequencies depend on natural selection. The leading proponent of sociobiology, Edward O. Wilson, an expert on ants, extended his analysis of animal behaviour to human society on the assumption that there are genes, subject to natural selection, ‘for’ traits such as homosexuality, xenophobia, and altruism.
Finally, the genes have come to life. They are intelligent, selfish, ruthless, and competitive, like ‘successful Chicago gangsters.’ This is the theory of selfish genes, propounded by Richard Dawkins. He traces their descent from primitive ‘replicator’ molecules in the primeval soup:
The replicators which survived were the ones that built survival machines for themselves to live in … Now they swarm in huge colonies, safe inside gigantic lumbering robots, sealed off from the outside world, communicating with it by tortuous indirect routes, manipulating it by remote control. They are in you and in me; they created us, body and mind; and their preservation is the ultimate rationale for our existence.19
Although organisms are regarded as ‘throwaway survival machines,’ there is nothing mechanistic about the selfish genes. They have powers to ‘create form’, to ‘mould matter’, to ‘choose’, to engage in ‘evolutionary arms races’ and even to ‘aspire to immortality’. As Dawkins says, ‘DNA works in mysterious ways.’20
The trouble with the selfish-gene theory is that it cannot possibly be true, as Dawkins acknowledges. DNA molecules cannot really be selfish or intelligent or mould matter or think. He also acknowledges that genes are not ‘for’ particular characteristics. Rather, alternative versions, or alleles, of genes are identified through differences between organisms. As he rightly says in his book The Selfish Gene,
Expressions like ‘gene for long legs’ or ‘gene for altruistic behaviour’ are convenient figures of speech, but it is important to understand what they mean. There is no gene which single-handedly builds a leg, long or short. Building a leg is a multi-gene cooperative enterprise. Influences from the external environment too are indispensible: after all, legs are actually made of food! But there may well be a single gene which, other things being equal, tends to make legs longer than they would have been under the influence of the gene’s allele.21
Nevertheless, despite this brief caveat, Dawkins promoted the idea of selfish genes as a ‘thought experiment’ and as a ‘powerful and illuminating’ metaphor. Indeed, he demonstrates through his popular appeal that it is more interesting to imagine that organisms are controlled by small living entities inside them, than that they are blind, unconscious mechanisms. Moreover, Dawkins is right in claiming that this way of looking at organisms is implicit within neo-Darwinism; he describes it as ‘a neo-Weismannist view of life.’22
Selfish genes may be like Weissman’s ‘determinants’, but they bear little resemblance to the chemical molecules of DNA. They have been endowed with the properties of life and mind.
DNA molecules have been given similar organizing and controlling powers by means of another powerful metaphor: the genetic program.23
Genetic programs
Whereas selfish genes are individualistic, like Weismann’s atomistic determinants, genetic programs are more holistic, recalling Weismann’s more holistic idea of the entire germ-plasm as a central directing agency. Genetic programs play much the same role as Driesch’s entelechies.
The notion of the genetic program has several attractions. First, it appears to account for the fact that most hereditary characteristics – for example the form of a cauliflower – have no obvious connection with DNA or protein molecules. If the genes somehow program the development of the cauliflower, then the vast gulf between this living structure and the DNA molecules is less obvious, even though nothing is actually known about the nature of the cauliflower program.
Secondly, the program is a subtler concept than the idea of genes ‘for’ particular characteristics. Genes are not atomistic determinants of separate features of the organism, but somehow many different genes co-operatively play a part. If they are like elements of a program, then their harmonious and co-operative activity is easier to understand. Thirdly, this notion suggests that development is purposive. Programs contain within themselves information about the end to which they are leading. Thus organisms can develop purposively towards these ends that are contained in their programs. Embryos can regulate and organisms can regenerate because of the purposive and holistic properties of their hereditary organizing principles. And ‘genetic programs’ seem to fit in well with the jargon of information theory and the linguistic metaphors in general use within modern biology. DNA ‘codes information,’ which can be ‘transcribed’ into RNA molecules, and then ‘translated’ into a sequence of amino acids as protein molecules are synthesized.
The metaphor of the genetic program inevitably suggests that development is organized by a pre-existing purposive principle that is either mind-like itself or designed by a mind. Computer programs are intelligently designed by human minds for particular purposes, and act upon and through the electronic machinery of a computer. The computer is a machine, but the program is not.
Perhaps morphogenesis is indeed organized by such a purposive directing principle. But if so, ‘genetic program’ is a misleading name for it: it is not genetic, in the sense that it is not in the genes, and morphogenesis is not in fact programmed in any meaningful sense of this word.
If the genetic program were carried in the genes, then all the cells of the body would be programmed identically, because in general they contain exactly the same genes. For example the cells of your arms and your legs are genetically identical and contain exactly the same kinds of protein molecules, and chemically identical bone, cartilage, and so forth. Yet they have different shapes. Clearly, the genes alone cannot explain these differences. They must depend on something else: on formative influences that act differently in different organs and tissues as they develop. These influences cannot be inside the genes; they extend over entire tissues and organs. At this stage the concept of the genetic program fades out, and is replaced by vague statements about ‘complex spatio-temporal patterns of physico-chemical activity not yet fully understood,’ or ‘mechanisms as yet obscure’.
The idea that development is programmed is misleading because for a phenomenon to be programmatic ‘it is a necessary condition that in addition to the phenomenon itself, there exists a second thing, the program, whose structure is isomorphic with, i.e., can be brought into a one-to-one correspondence with, the phenomenon.’24 This is indeed the case in the clear causal chain leading from the sequence of chemical bases in DNA molecules to the sequence of amino acids in peptides. But here the programming ends. The folding up of the peptides into the characteristic three-dimensional structure of proteins is not programmatic, for it has no isomorphic correspondence in the DNA. In relation to morphogenesis itself, the overall sequence of events is not isomorphic with the genes. For example:
Studies of the development of the nervous system have shown that the notion of genetic programming is not only defective at the conceptual level, but also represents a misinterpretation of the knowledge already available from developmental studies … We already know enough about its mode of establishment to make it most unlikely that the nervous system is pre-specified; rather, all indications point to stochastic [i.e., probabilistic] processes as underlying the apparent regularity of neural development.25
Modern mechanistic biology grew up in opposition to vitalism, the doctrine that living organisms are organized by purposive, mindlike principles (Fig. 5.5).26 But biology now has purposive mindlike organizing principles of its own: the genetic programs. Moreover, purpose is no longer denied but admitted. The old term teleology, with its Aristotelian associations, has been replaced by the new term teleonomy, the ‘science of adaptation.’ As Dawkins has pointed out, ‘in effect teleonomy is teleology made respectable by Darwin, but generations of biologists have been schooled to avoid ‘teleology’ as if it were an incorrect construction in Latin grammar, and many feel more comfortable with a euphemism.’27
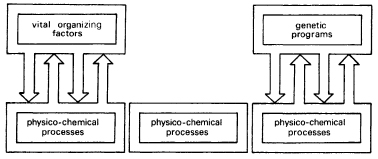
Figure 5.5 On the left, a diagrammatic representation of the vitalist theory: the physico-chemical processes within organisms interact with and are organized by vital organizing factors, such as entelechy. The mechanistic theory denies the existence of such vital factors, and asserts that life can be understood in terms of physico-chemical processes alone (centre). In the modern conception (right), these processes are organized by genetic programs or genetic information, which play much the same role as the organizing factors of vitalism.
Thus the paradigm of modern biology, although nominally mechanistic, has in effect become remarkably similar to vitalism, with genetic ‘programs’ or ‘information’ or ‘instructions’ or ‘messages’ playing the role formerly attributed to vital factors such as entelechies.
Mechanists have always accused vitalists of trying to explain the mysteries of life in terms of empty words, such as entelechy, which ‘explain everything and therefore nothing.’ But the vital factors in their mechanistic guises have exactly this characteristic. How does a marigold grow from a seed? Because it is genetically programmed to do so. How does a spider instinctively spin its web? Because of the information coded in its genes. And so on.
The duality of matter and information
All attempts to force the organizing principles of life into material objects such as genes have failed: they keep bursting out again. The concept of purposive organizing principles has been reinvented again and again.
In fact this duality of matter and organizing principles has been implicit in the mechanistic theory of life all along. It is an essential feature of the machine metaphor. All machines involve a duality between the materials of which they are made, like steel, and the purposive designs in the minds of their designers and makers. As the theoretical biologist Francisco Varela expressed it:
What defines a machine organization is relations, and hence … the organization of a machine has no connection with materiality, that is, with the properties of the components that define them as physical entities. In the organization of a machine, materiality is implied but does not enter per se.28
This duality of form and matter is inherent in all traditional philosophies of form, as we saw in the preceding chapter. In the modern context, it is usually conceived of in terms of the duality of matter and information. Information is what in-forms; it plays an in-formative role, as Norbert Weiner, the founder of cybernetics, emphasized in his concept of the primacy of information over matter and energy. He saw this distinction as essential for the doctrine of materialism: ‘No materialism which does not admit this can survive at the present day.’29 This may sound like a radical position, but in fact ever since the seventeenth century, the survival of materialism has depended on its combination with the Platonic notion of non-material organizing principles: the laws of nature (Chapter 2).
If biological ‘information’ cannot be understood in terms of the material structures of the genes alone, then what is it? Is the information Platonic, somehow transcending time and space? Or is it immanent within organisms?
In the following chapter we consider the possibility that such information is carried in morphogenetic fields, inherited by organisms from their predecessors. But before doing so, we look again at the assumption that this information is inherited materially in the genes.
Why genes are overrated
Genes are known to code information for the sequence of chemical building blocks in RNA and protein molecules. They help to provide a detailed understanding of the way in which organisms inherit biochemical potentialities. But they do not code for morphogenesis or for inherited patterns of behaviour. They are not ‘determinants’ of the characteristics of the organism.
Genetics deals with hereditary differences between organisms. For instance, if a certain gene is present in one form or another – the technical term is ‘allele’ which literally means ‘another’ – the structure of a fruit-fly may differ (Fig. 5.6). But the fact that different versions of genes lead to differences in form does not prove that genes determine the form.
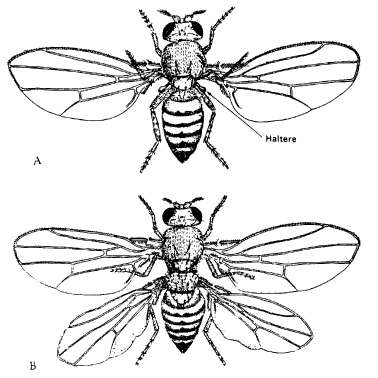
Figure 5.6 A: A normal specimen of the fruit-fly Drosophila. B: A mutant fly in which the third thoracic segment has been transformed in such a way that it duplicates the second thoracic segment. Such flies are known as bithorax mutants.
To see why, think of a radio set. A change or mutation in one of its transistors might cause the sounds that it is producing to become distorted; a mutation in one of the condensors in its tuning circuit might cause the set to pick up another radio station, with the result that an entirely different series of sounds comes out of the loudspeakers. But the fact that mutations in the set’s components can cause differences in sounds does not prove that the sounds are determined or programmed by the components of the set. They are in fact coming from radio stations and are transmitted through the electro-magnetic field. The mutant condenser is not a component ‘for’ a particular sequence of sounds.
Many biologists are well aware that it is misleading to speak of genes ‘for’ particular characteristics. Richard Dawkins, for instance, has made it clear that if a geneticist speaks of genes ‘for’ red eyes in the fruit-fly Drosophila, he implicitly means that ‘there is variation in eye colour in the population: other things being equal, a fly with this gene is more likely to have red eyes than a fly without the gene.’ However, he defends talking about genes ‘for’ particular characteristics as a ‘routine genetic practice.’30 And then on he goes with his selfish gene theory.
These habits of thought endure in spite of the fact that they are known to be misleading because they are almost unavoidable. They follow from the assumption that heredity must be material. Hence all the hereditary information for the shape of a pigeon’s foot and the web-spinning instincts of a spider must be in the genes: where else could they possibly be?
By contrast, the hypothesis of formative causation limits the role of genes to what they are known to do: namely to code information for the sequence of chemical building blocks in RNA and protein molecules. It does not suppose that the genes organize the whole plant or animal. Rather, it sees these hereditary organizing principles as fields that are inherited non-materially by morphic resonance.
But is there any need for such a concept? Why cannot embryonic development be understood in terms of chemical patterns that both arise from and control the activity of the genes?
Chemical theories of pattern formation
The genes an organism inherits underlie its capacity to make particular RNA and protein molecules. With the help of the ingenious techniques of genetic engineering, specific portions of DNA can now be transferred from one organism to another, enabling it to make proteins that it could not make before. For instance, the DNA that constitutes the human gene for insulin has been transferred to cells of the bacterium Escherichia coli, and this protein is made commercially by growing these modified bacteria and purifying the insulin they produce.
In the course of morphogenesis, cells differentiate, become different – as skin, muscle, and nerve cells, for example – and different kinds of cells make different proteins. Although they all contain the same genes, different genes are expressed. For example, in the development of a chrysanthemum flower, the enzymes responsible for making pigment molecules are made in the cells of the developing petals, and then through the activity of these enzymes, the coloured pigments appear in the petals. But to describe such chemical changes does not explain how morphogenesis is controlled, or why petals form instead of leaves.
Chemical changes accompany morphogenesis, and organisms could not develop without the production of the right molecules in the right places at the right times. But producing particular sets of molecules does not explain morphogenesis. The delivery of the right building materials and machinery to building sites does not result in the spontaneous growth of houses.
The main emphasis in research over the last few decades has not been on morphogenesis itself, but on the control of protein synthesis. How are the right proteins made in the right cells at the right times and in the right amounts? How is the expression of genes controlled as cells differentiate within the developing organism?
Clearly, patterning influences are at work within the developing tissues and organs. These are generally thought of as systems of ‘positional information,’ which ‘tell’ cells where they are and thus enable them to respond appropriately by making the right proteins. What is the nature of this so-called positional information?
The most popular idea is that it is chemical, and depends on concentration gradients of specific chemical substances called morphogens.31 The main progress has come from making mathematical models of the ways in which such chemical patterns could in theory be formed.
Many of these models depend on the principle that Ilya Prigogine has summed up in the phrase ‘order through fluctuations.’32 In a system that is unstable, far from thermodynamic equilibrium, random fluctuations can be amplified through various kinds of positive feedback, and under certain conditions they spontaneously give rise to patterns. For example, in certain kinds of chemical reaction, where at least two substances are reacting catalytically with each other and diffusion is taking place, concentration patterns can appear as the chemicals react (Fig. 5.7). Prigogine led the way in showing how such processes can be described mathematically in terms of non-equilibrium thermodynamics. He pointed out that order can arise from ‘chaos’ in comparable ways in many systems, ranging from patterns of convection in heated fluids to patterns of urban growth. For example, once towns and cities have started to develop in certain places, they tend to increase in size through the migration of population, leading in turn to an intensification of economic activity and more migration; but this growth is limited by a variety of other factors, including competition with other cities, especially those that are nearby.

Figure 5.7 The formation of spiral chemical waves when the Belousov-Zhabotinskii reagent is allowed to stand in a shallow dish. The waves appear spontaneously as the chemical reaction proceeds, or they can be initiated by touching the surface with a hot filament, as in the pictures shown here. The numbers indicate how many seconds had elapsed after the initial photograph was take. (Adapted with permission from Being to Becoming by Ilya Prigogine, copyright © 1980, W. H. Freeman and Co.)
Hans Meinhardt summarized the principles involved in the making of such models of pattern formation in developing organisms as follows:
Assuming that development is controlled by substances, any theory of development has to describe concentration changes of substances as a function of other substances involved and as a function of spatial co-ordinates and time. Two conditions have to be fulfilled before a stable pattern can be generated. (1) A local deviation from an average concentration should increase further, otherwise no pattern would be formed, and (2) the increase should not go to infinity. Instead, the emerging pattern should reach a stable steady state.33
On the basis of these assumptions, Meinhardt and his colleague Alfred Gierer and others have constructed a variety of mathematical models involving hypothetical ‘activator’ and ‘inhibitor’ substances. These have been used in computer simulations to show the kinds of patterns they can generate (Fig. 5.8). The interesting feature of some of these models is that they show self-regulatory properties such that the pattern can be restored after part of the model system is ‘removed.’ Meinhardt and Gierer have in fact suggested that they are models of morphogenetic fields. We will come back to them again in the next chapter in the context of other ideas about the nature of these fields.
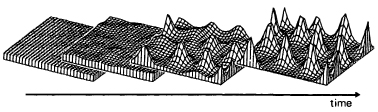
Figure 5.8 A computer model of pattern formation. A bristle-like pattern is formed in a non-growing field as a result of random fluctuations which lead to the local production of an ‘activator’ which enhances further ‘activator’ production, and at the same time to the production in these centres of an ‘inhibitor’, which diffuses outwards and inhibits further centres from forming nearby. (From Models of Biological Pattern Formation by H. Meinhardt; Academic Press, 1982. Reproduced by permission.)
If the hypothetical activators and inhibitors are actually identified in embryos, then they will help to explain how the synthesis of different proteins in different cells is controlled. But they will still leave unexplained what the cells do with the proteins, how they take up their shapes, how they behave, how some of them move around within embryos, how tissues and organs take up their forms, and how organisms respond to their environment. How can the gulf between these hypothetical chemical gradients and the actual organism be bridged? The answer suggested by the author of the concept of positional information, Lewis Wolpert, is that the cells ‘interpret that information according to their genetic program.’34
As we have seen, the concept of the genetic program is misleading, if only because development is not programmatic. A number of leading developmental biologists have suggested that this concept be abandoned.35 Sydney Brenner, for example, proposed that it be replaced by terms such as ‘internal representation’ or ‘internal description.’36 He has summarized thinking among developmental biologists as follows:
At the beginning it was said that the answer to the understanding of development was going to come from a knowledge of the molecular mechanisms of gene control. I doubt whether anyone believes that anymore. The molecular mechanisms look boringly simple, and they don’t tell us what we want to know. We have to try to discover the principles of organization.37
What might such principles of organization be? This is just the question that organismic philosophers and biologists have been wrestling with for decades.
Organic wholes
The organismic or holistic approach developed under the influence of Whitehead’s philosophy of organism (see above) has been influential within biology since the 1930s. It has enabled holistic properties of organisms to be recognized without the need to adopt a vitalist position; and indeed it has offered an attractive way of ‘transcending’ the vitalist-mechanist controversy.38
Vitalists stressed the holistic, organic qualities of plants and animals, but they did not challenge the mechanistic orthodoxy of physics in relation to non-living things; they made a sharp distinction between the inanimate realm and the realm of life.
By contrast, mechanists asserted that there is no difference in kind, only in degree, between the realm of biology and the realms of chemistry and physics. Organicists agree with the mechanists in this respect, and preserve their intuition of the fundamental unity of nature. But rather than regarding living organisms as inanimate machines, they regard physical and chemical systems, such as atoms, molecules, and crystals, as in some sense living; they are not mere inanimate material objects, but ‘structures of activity,’ or organisms.
The organismic approach is not reductionistic or atomistic in intent: it does not assume that atoms or subatomic particles have a privileged place in the scale of nature, and it does not attempt to explain all the properties of larger and more complex organisms in terms of the properties of their parts; at each hierarchical level of complexity, organisms behave as wholes, with an organic unity that is irreducible.
In general these hierarchies are ‘nested’ in such a way that higher-level wholes are made up of parts which are themselves organisms at a lower level. Thus sugar crystals, for example, are organisms whose parts are sugar molecules, which are wholes made up of carbon, hydrogen, and oxygen atoms, which are wholes composed of electrons in orbitals around nuclei, and the nuclei are wholes made up of yet smaller organisms, the nuclear particles, which are themselves composed of entities such as quarks. Living organisms show a similar hierarchical arrangement, with organs, containing tissues, containing cells, containing organelles such as nuclei and mitochondria, containing complex molecules, and so on (Fig. 5.9).
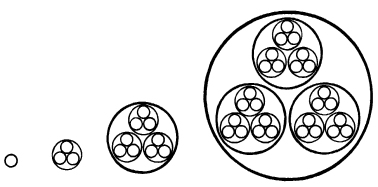
Figure 5.9 Successive levels in a nested hierarchy of morphic units, or holons. At each level, the holons are wholes containing parts, which are themselves wholes containing lower-level holons, and so on. The diagram could represent subatomic particles in atoms, in molecules, in crystals, for example; or cells in tissues, in organs, in organisms.
Arthur Koestler proposed the term holon for such organisms, which at the same time can be wholes made up of parts, and parts of higher-level wholes: ‘Every holon has a dual tendency to preserve and assert its individuality as a quasi-autonomous whole; and to function as an integrated part of an (existing or evolving) larger whole. This polarity between the Self-Assertive and Integrative tendencies is inherent in the concept of hierarchic order.’39 For such a nested hierarchy of holons, he proposed the term holarchy.
Another term with a meaning equivalent to holon is morphic unit.40 The word morphic emphasizes the aspect of form, and the word unit emphasizes the unity or wholeness. Such units are formed by what L. L. Whyte has called ‘morphic processes’ in which ‘well-formed terminal states can arise from less-formed initial ones.’41
The organismic approach has encouraged a search for general principles that apply to organisms or ‘systems’ at any level of complexity. This is the goal of general systems theory, which has been strongly influenced by cybernetics, the theory of communication and control, with its basic concepts of information-transfer and feedback. Many mathematical models have been constructed in the spirit of this systems approach, both within biology and in industry, commerce, and society in general.42 Related ‘systems approaches’ include game theory; and the metaphor of games, relying on the interplay of chance and rules, has been applied to biological evolution and to the development and behaviour of living organisms.43
In living systems theory, James G. Miller distinguished seven levels of living systems (cell, organ, organism, group, organization, society, and supra-national system) and identified nineteen ‘critical subsystems’ at each level: for example ‘reproducer’, ‘boundary,’ and ‘ingestor.’ At the cellular level, for instance, these particular subsystems are represented by, respectively, chromosomes, cell membranes, and gaps in cell membranes.44 Such a classification permits illuminating cross-level comparisons and insights. But the very generality of the systems approach has limited its usefulness in explaining the morphogenesis of any actual plants or animals.
Since the turn of the millennium, ‘systems biology’ has undergone a resurgence of popularity in response to the avalanche of data emerging from genome projects and from proteomics, the large-scale study of proteins. Modern systems biology views the organisms as ‘an integrated and interacting network of genes, proteins and biochemical reactions which give rise to life.’45 But although such computer models help in computing and predicting the rates of chemical processes, they shed very little light on morphogenesis.
Among organismically-minded biologists, the most fruitful idea has proved to be the concept of morphogenetic fields. Morphogenesis remains mysterious. Can morphogenetic fields help us to understand it?