Grape Species and Varieties
The distinctive structural and genetic nature of genus Vitis is covered, followed by a discussion of its geographic origin and distribution, notably V. vinifera – the wine grape. Subsequently, the domestication of V. vinifera is explored, as well as modern views on the likely origins of grape cultivars, their selection, and spread. This includes the development of rootstocks, demanded as a consequence of the European invasion by phylloxera. Standard as well as modern breeding techniques are noted, including clonal selection. The chapter ends with a discourse on the methods used in cultivar identification, origin studies, and the distinctive attributes of some of the more significant cultivars.
Keywords
Vitis; Vitis vinifera; wine grapes; grape cultivars; cultivar origin; rootstocks; grape breeding; cultivar identification
Introduction
Grapevines are classified in the genus Vitis, within the Vitaceae. Other well-known members of the family are the Boston Ivy (Parthenocissus tricuspidata) and Virginia Creeper (P. quinquefolia). Members of the Vitaceae are typically woody, show a climbing habit, have leaves that develop alternately on shoots (Fig. 2.1), and possess swollen or jointed nodes. These may generate tendrils or flower clusters opposite the leaves. The flowers are minute, uni- or bisexual, and occur in large clusters. Most flower parts appear in groups of fours or fives, with the stamens developing opposite the petals. The ovary consists of two carpels, partially enclosed by a receptacle that develops into a two-compartmented berry. The fruit contains up to four seeds.
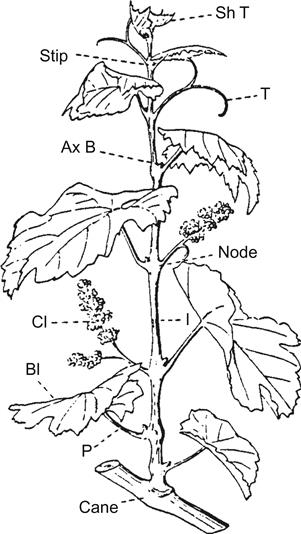
The Vitaceae is predominantly a tropical to subtropical family, containing about 900 species, divided among some 14 genera (Galet, 1988). In contrast, Vitis is primarily a temperate-zone genus, occurring indigenously only in the Northern Hemisphere. Related genera include Acareosperma, Ampelocissus, Ampelopsis, Cayratia, Cissus, Clematicissus, Cyphostemma, Nothocissus, Parthenocissus, Pterisanthes, Pterocissus, Rhoicissus, Tetrastigma, and Yua.
The Genus Vitis
Grapevines are distinguished from related genera primarily on floral characteristics. The flowers are typically functionally unisexual, being either male (possessing erect, functional anthers, and lacking a fully developed pistil) or female (containing a functional pistil, and either producing recurved stamens and sterile pollen, or lacking anthers) (Fig. 2.2). The petals are fused, forming a calyptra or cap. The petals remain connected at the apex, only splitting along the base at maturity, when the calyptra is shed (see Plate 3.6). Occasionally, though, the petals may separate at the top, while remaining attached at the base (Plate 2.1). These ‘star’ flowers possess an appearance resembling typical flowers. This situation characterizes some members of the Vitaceae, for example Cissus. In some cultivars, star flower production is induced by cool temperatures, whereas in others it is a constitutional property (Longbottom et al., 2008). The trait is not genetically transmissible because star flowers are sterile, generating seedless berries (Chardonnay), or no fruit (Shiraz).

Swollen nectaries occur at the base of the ovary (see Fig. 3.25C). Despite their name, they do not produce nectar, but they produce a mild fragrance that attracts pollinating insects. The sepals of the calyx form only as vestiges and degenerate early in flower development. The fruit is juicy and acidic.
The genus has typically been divided into two subgenera, Vitis1 and Muscadinia. Vitis (bunch grapes) is the larger of the two subgenera, containing all species except V. rotundifolia and V. popenoei. The latter are placed in the subgenus Muscadinia (muscadine grapes). The two subgenera are sufficiently distinct to have induced some taxonomists to separate the muscadine grapes into their own genus, Muscadinia.
Members of the subgenus Vitis are characterized by having shredding bark, nonprominent lenticels, a pith interrupted at nodes by woody tissue (the diaphragm), tangentially positioned phloem fibers, branched tendrils, elongated flower clusters, berries that adhere to the fruit stalk at maturity, and pear-shaped seed possessing a prominent beak and smooth chalaza. The chalaza is a pronounced, circular, depressed region on the dorsal (back) side of the seed (Fig. 2.3C). In contrast, species in the subgenus Muscadinia possess a tight, nonshredding bark, prominent lenticels, no diaphragm interrupting the pith at nodes, radially arranged phloem fibers, unbranched tendrils, small floral clusters, berries that separate individually from the cluster at maturity, and boat-shaped seed with a wrinkled chalaza. Some of these characteristics are diagrammatically illustrated in Fig. 2.3. Plate 2.2 illustrates the appearance of Muscadinia grapes and leaves.

The two subgenera also differ in their chromosomal composition. Vitis species contain 38 chromosomes (2n=6x=38), whereas Muscadinia species possess 40 chromosomes (2n=6x=40). The symbol n refers to the number of chromosome pairs formed during meiosis, and x refers to the number of chromosome complements (genomes that were involved in their evolution).
Successful crosses can be experimentally produced between species of the two subgenera, primarily when V. rotundifolia is used as the pollen source. When V. vinifera is used as the male plant, the pollen germinates, but does not effectively penetrate the style of the V. rotundifolia flower (Lu and Lamikanra, 1996). This may result from the synthesis of inhibitors, such as quercetin glycosides in the pistil (Okamoto et al., 1995). Although generally showing vigorous growth, the progeny frequently are infertile. This probably results from imprecise pairing of the unequal number of chromosomes (19+20) and the consequential imbalanced separation of the chromosomes during meiosis. The genetic instability so produced disrupts pollen growth, resulting in infertility.
The evolution of the Vitaceae appears to have involved hybridization and subsequent chromosome doubling, a feature common in many plants (Soltis and Soltis, 2009). In the Vitaceae, three separate whole-genome duplication events are suspected, based on DNA genomic analyses (Jaillon et al., 2007; Velasco et al., 2007). Their proposals differ basically only in the timing of the duplications – whether the last duplication occurred before or after the progenitors of the Vitaceae split from other rosid dicotyledons. Velasco et al. (2007) consider that the last duplication involved at least 10 chromosomes. These views differ from the proposals of Patel and Olmo (1955). They viewed the timing of the ploidy events occurring after the genus Vitis evolved from other Vitaceae. Patel and Olmo used cytogenetic evidence to envision two duplication events – the first involving hybridization between progenitors with six and seven chromosome pairs (6+7=13, doubling to 26). Later, a second set of hybridization events of such tetraploids, with diploids of 12 and 14 chromosomes respectively, followed by chromosome doubling, gave rise to current day hexaploids – the subgenera Vitis (13+6=19, doubling to 38) and Muscadinia (13+7=20, doubling to 40) (Fig. 2.4). Variations in the chromosome numbers of other genera in the Vitaceae could be viewed as later modifications, involving phenomena such as chromosome loss, fusion, translocation, and/or doubling. For example, other Vitaceae possess 22 chromosomes (Cyphostemma); 22, and occasionally 44 (Tetrastigma); 24, and occasionally 22 or 26 (Cissus); 32, 72, or 98 (Cayratia), and 40 (Ampelocissus and Ampelopsis).

Older cytogenetic evidence, consistent with polyploidy, involves the apparent association of four nucleoli in species possessing 24 and 26 chromosomes, and six nucleoli-related chromosomes in species possessing 38 or 40 chromosomes. Typically, diploid species possess two nucleoli-related chromosomes. Chromosome structure itself provides little information concerning evolutionary events in the Vitaceae, due to their minute size (0.8–2.0 μm) and morphological similarity (Fig. 2.5)

Although polyploidy was involved in the evolution of the Vitaceae, members also appear to have undergone diploidization, similar to other polyploid angiosperms (Soltis and Soltis, 1999; Cui et al., 2006). Diploidization typically involves a relatively rapid and significant genomic reorganization (usually involving transposons). This is expressed in a structural modification of duplicate homologous genes from the combined genomes. This may explain why only two nucleoli appear prominently in the cells of Vitis spp. (Haas et al., 1994), in contrast to the six that might otherwise be expected.
The increased potential for adaptation to environmental change provided by polyploidy (mutational modification of unnecessary, duplicate genetic traits) seems to have been the principal evolutionary feature favoring hybridization and chromosomal doubling (Fawcett et al., 2009). Alternatively, gene duplicates are silenced, becoming nonfunctional (null) pseudogenes. This modification converts polyploids into functional diploids. An important secondary consequence of these structural modifications is preventing, or regulating, multivalent crossovers between multiple sets of similar chromosomes. This avoids unbalanced chromosome separation during meiosis, and the development of aneuploidy. Unequal chromosome complements in ovules and pollen often lead to partial or complete seed sterility.
In contrast to the relative genetic isolation (infertility), imposed by the differing chromosome complements of the Vitis and Muscadinia subgenera, crossing among species within each subgenus produces fertile progeny. Although facilitating the incorporation of desirable traits from one species into another, the ease with which interspecies crossing occurs complicates the taxonomic task of delineating species. This is further compounded by many species possessing overlapping geographic distributions (Fig. 2.6), and the scarcity of distinctive morphological features. Quantitative differences that distinguish species, such as shoot and leaf hairiness, are often influenced by environmental conditions. This is often interpreted as evidence that speciation is still active within the genus, and evolution into distinct species remains incomplete – local populations often being viewed more as ecospecies or ecotypes than biological species. These complications may be partially resolved by molecular techniques (Tröndle et al., 2010).
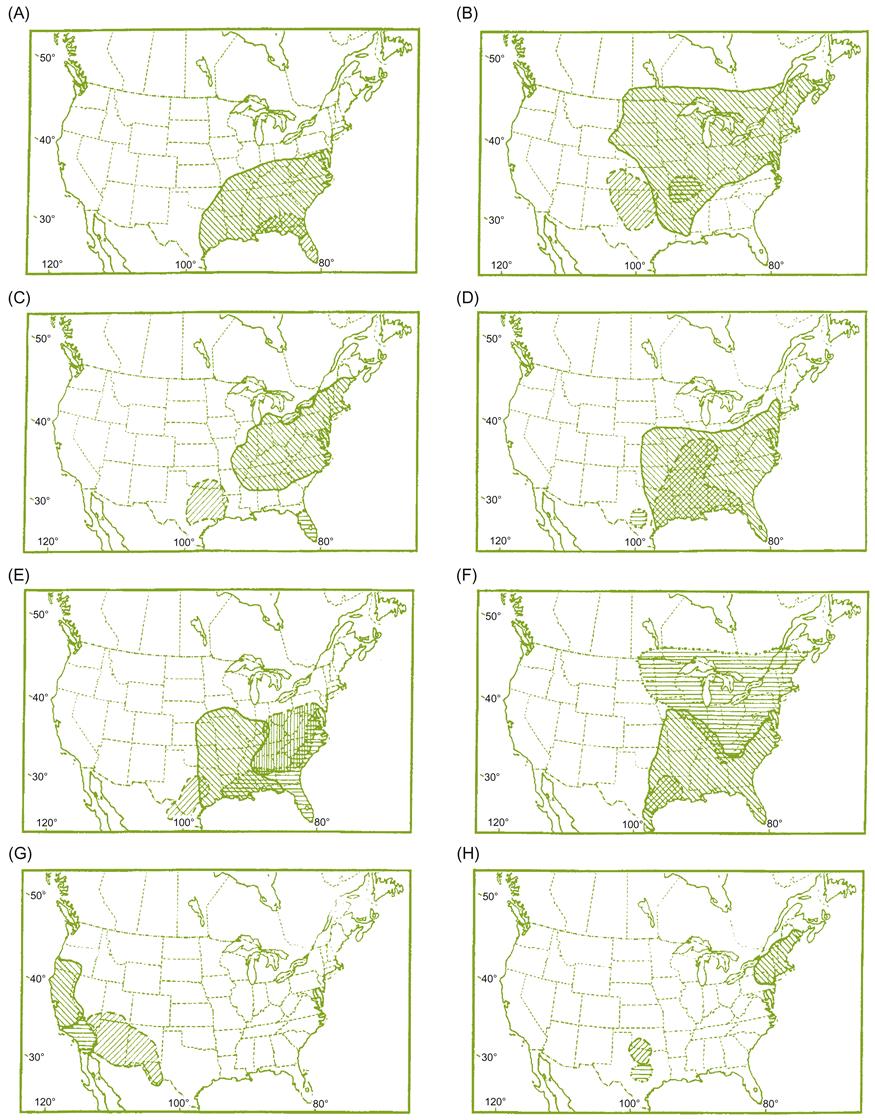
The most recent classification of eastern North American species of Vitis is given by Moore (1991). A summary of the classification of Chinese species is given in Wan et al. (2008). Data derived from molecular (DNA) markers generally support relationships based traditionally on morphologic features, but also highlight differences (Aradhya et al., 2008). Their data suggest considerable gene flow among species.
Geographic Origin and Distribution of Vitis and Vitis vinifera
When and where the genus Vitis evolved is unclear. The current distribution includes northern South America (the Andean highlands of Colombia and Venezuela), Central and North America, Asia, and Europe. This would correlate with the ancestry of at least the original polyploidy having occurred while North America and Europe were still contiguous parts of Pangea. In contrast, species in the subgenus Muscadinia are restricted to the southeastern United States and northeastern Mexico, possibly indicating that its origin occurred close to or after North America had split from Pangea (the Cretaceous). This was the time when flowering plants began to replace gymnosperms as the predominate land plants. The North American distribution of Vitis species is shown in Fig. 2.6.
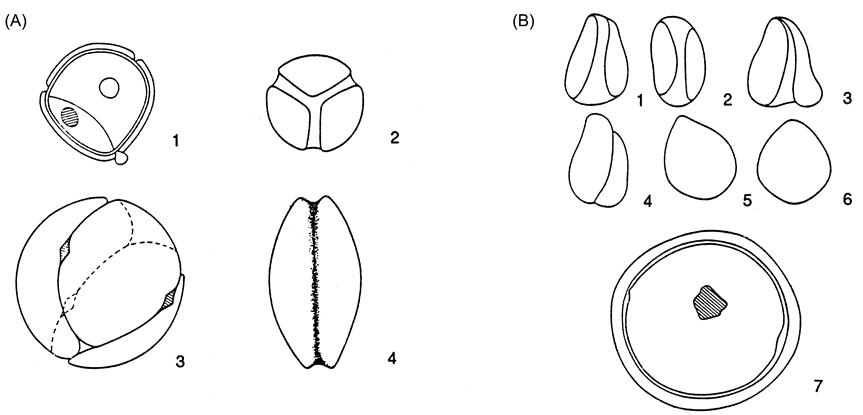
In the nineteenth century, many extinct species of Vitis were proposed, based on fossil leaf impressions (see Jongmans, 1939). The validity of many of these designations is now in question, due to the dubious nature of the evidence (Kirchheimer, 1938). Not only do several unrelated plants possess leaves similar in outline, but individual grapevines may show remarkable variation in leaf shape, lobbing, and dentation (Zapriagaeva, 1964). Of greater value in species delineation is seed morphology, even though significant interspecies variation also exists (Fig. 2.7). On the basis of seed morphology, two groups of fossilized grapes have been distinguished, Vitis ludwigii and V. teutonica. Seeds of the V. ludwigii type, resembling those of muscadine grapes, have been found in Europe from the Pliocene (2–10 million years b.p.). Those of the V. teutonica type, resembling those of bunch grapes, have been discovered as far back as the Eocene (34–55 million years b.p.). However, these identifications are based on comparatively few specimens, and, thus, any conclusions remain tenuous. In addition, related genera, such as Ampelocissus and Tetrastigma produce seed similar to those of Vitis. This would fit with the suspected much earlier evolution of these genera. In contrast, species radiation in the genus Vitis is estimated to have begun about 6–6.6 million years b.p. (Zecca et al., 2012). Although most grape fossils have been found in Europe, this may reflect more the distribution of appropriate sedimentary deposits (or paleobotanical interest and investigation) than Vitis distribution.

Baranov (in Zukovskij, 1950) suggests that the progenitors of Vitis were bushy and inhabited sunny locations. As forests expanded during the more humid Eocene, the development of a climbing habit would have permitted them to retain their preference for sunny conditions. This may have involved mutations modifying some floral clusters into tendrils, thus improving clinging ability. This hypothesis is not unreasonable in light of the differentiation of buds into flower clusters or tendrils, based on the relative balance of gibberellin versus cytokinin in the tissue (Srinivasan and Mullins, 1981; Martinez and Mantilla, 1993).
Regardless of the manner and geographic origin of Vitis, the genus established its present range by the end of the last major glacial period (~8000 b.c.). It is believed that periodic advances and retreats during the last glacial period markedly affected the evolution of Vitis, and its survival in Eurasia. The alignment of the major mountain ranges in the Americas, versus Eurasia, appears to have had an important bearing on the respective evolution of populations within the Northern Hemisphere. In the Americas and eastern China, the mountain ranges run predominantly north–south, whereas in Europe and western Asia they run principally east–west. This would have permitted North American and eastern Chinese species to relocate, south or north, relative to the movement of glacial ice sheets. In contrast, the southward movement of grapevine species in Europe and western Asia would have been largely restricted by the east–west mountain ranges (Pyrenees, Alps, Caucasus, and Himalayas). This may explain the much larger species presence in North and Central America (possessing some 34 endemic species – Rogers and Rogers, 1978) and China (possessing about 30 indigenous species – Fengqin et al., 1990). In contrast, Eurasia possesses but one, V. vinifera.
A few naturalized, foreign Vitis spp. are occasionally found in Europe. These are derived from ‘escaped’ rootstocks, imported for grafting, subsequent to the phylloxera epidemic in the late 1800 s (Arrigo and Arnold, 2007), or cultivated as a fruit crop (Cangi et al., 2006). Some of these have also yielded hybrids with indigenous wild Vitis vinifera (Bodor et al., 2010).
Although glaciation and cold destroyed most of the favorable habitats in the Northern Hemisphere during the last ice age, major southward displacement was not the only option open for survival. In certain areas, isolated but favorable sites (refuges) permitted continued existence throughout the glacial period. In Europe, refuges occurred around the Mediterranean basin, and south of the Black and Caspian Seas (Fig. 2.8). For example, grape seeds have been found associated with anthropogenic remains in caves in Southern Greece (Renfrew, 1995) and Southern France (Vaquer et al., 1985), both near the end of the last glacial advance. These refuges may have played a role in the evolution of the various varietal groups of Vitis vinifera (see below).
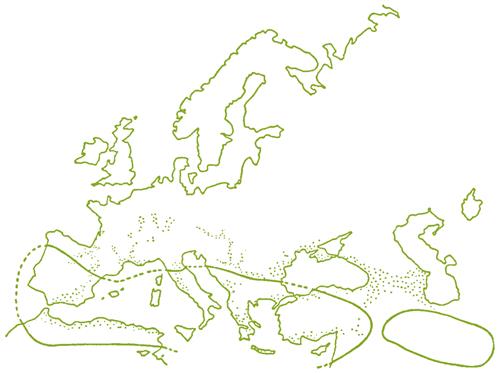
Although periodically displaced during the various Quaternary glacial periods (Fig. 2.9), V. vinifera was inhabiting southern regions of France and northern Greece some 10,000 years ago (Planchais and Vergara, 1984; Wijistra, 1969). For the next several thousand years, the climate slowly improved to an isotherm about 2–3 °C warmer than current (Dorf, 1960). Preferred habitats of wild V. vinifera were in the mild, humid forests south of the Caspian and Black Seas and adjacent Transcaucasia, along the fringes of the cooler mesic forests of the northern Mediterranean, and into the heartland of Europe, along the banks and hillsides of the Danube, Rhine, and Rhône rivers. The current situation of wild Vitis vinifera is discussed in Arnold et al. (1998).

Domestication of Vitis vinifera
Grapevine cultivars show few of the standard signs of plant domestication noted by Baker (1972) and de Wet and Harlan (1975). Their views can be summarized as follows: conversion from cross- to self-fertilization, release from the need for seed and bud vernalization (cold treatment), elimination of photoperiod phenologic regulation, inactivation of seed dehiscence or fruit separation upon maturation, development of parthenocissus (fruit production independent of seed development), increase in shoot to root ratio, enhanced fruit (or seed) size, augmentation in crop yield, reduction in phytotoxin production, and a shift from a bi- or perennial habit to an annual habit.
Of these changes, only conversion to self-fertility is clearly expressed in domesticated grapevines. Other modifications associated with domestication are less marked. There is a slight reduction in photoperiod sensitivity and need for seed vernalization; easier fruit separation from the cluster; increased fruit size (notably in table grapes); and seedlessness in some table and raisin cultivars. Other features that tend to differentiate wild from domesticated grapevines include a shift from small, round berries to larger, elongated fruit; more fruit per cluster; bark separating in wider, core-coherent strips (vs. bark separating in long thin strips); larger, elongated seeds (vs. smaller, rounded seeds); and large leaves with entire or shallow sinuses (vs. smaller, usually deep, three-lobed leaves) (Olmo, 1976). Some of these features are illustrated in Fig. 2.10. Plate 2.3 also illustrates the characteristic features of wild (sylvestris) grape clusters, including the dominant (wild-type) bluish fruit coloration.

The principal indicator of domestication, detectable in archaeological remains, is a shift in seed index – the ratio of seed width to length. Although of no known selective advantage, the shift appears to correlate with a change from cross- to self-fertilization. This may reflect some unknown and unsuspected pleiotrophic effects. Seed from wild (sylvestris) vines are rounder, possess a nonprominent beak, and show an average seed index of about 0.64 (ranging from 0.54 to 0.82). In contrast, seed from domesticated (sativa) vines are more elongated, possess a prominent beak, and have a seed index averaging about 0.55 (often ranging from 0.44 to 0.75) (Renfrew, 1973; Fig. 2.11). However, evidence based on carbonated seed remains may be of uncertain value. Charring appears to increase the relative seed length, decreasing the seed index (Margaritis and Jones, 2006). It does, though, enhance survival in sites. In addition, considerable variation in seed size and shape (Zapriagaeva, 1964) makes conclusions based on small sample sizes unreliable.

Seed index data have been used by Miller (1991) to suggest a slow domestication of grapevines in northeastern Iran and Anatolia. However, clear evidence of domestication begins only in the late fourth millennium b.c., in Jericho about 3200 b.c. (Hopf, 1983), and after 2800 b.c. in Macedonia and Greece (Renfrew, 1995). This is considerably later than the first archaeological evidence suggestive of wine production in northern Iran (5400–5000 b.c.) (McGovern et al., 1996). However, the time frame is consistent with the predicted spread of agriculture out of the Fertile Crescent into northern Iran and Anatolia (Ammerman and Cavalli-Sporza, 1971). Thus, the production of wine may have developed concurrently with agriculture, but predated preserved morphological evidence of grapevine domestication (Zohary, 1995).
In Europe, the earliest presumptive evidence of winemaking comes from the northern Aegean, dating from the latter half of the fifth millennium b.c. (Valamoti et al., 2007). In western Europe, evidence of wild grapevine use has been found in a Neolithic village near Paris (about 4000 b.c.) (Dietsch, 1996). Semidomesticated grape seed remains (2700 b.c.) have been discovered in England (Jones and Legge, 1987), and in several Neolithic and Bronze Age lake dwellings in northern Italy, Switzerland, and the former Yugoslavia (see Renfrew, 1973). In addition, grape seed and pollen remains have been identified from Neolithic sites in Southern Sweden and Denmark (see Rausing, 1990). At that time, the climate was warmer than today. Evidence consistent with grape culture in southwestern Spain has been found dating from about 2500 b.c. (Stevenson, 1985).
Differences in floral sexual expression and leaf shape occur between wild and domesticated grapevines (Levadoux, 1956). For example, male vines have larger inflorescences, flower earlier, and for a longer period than bisexual flowers (Negi and Olmo, 1971). Male flowers also show poorly developed nectaries. These features are, however, not preserved or recognizable in vine remains. Additional differences have been noted by Patel and Olmo (1955), but most of these features are readily influenced by cultivation.
Another potentially useful property, preserved in archaeological remains, involves changes in the fruit pedicel (see Fig. 3.23 for terminology). In domesticated Vitis vinifera, the pedicel frequently breaks at the main stem (rachis) when the berries are pulled. In contrast, this seldom occurs with wild vines, due to their stouter stems. Thus, the relative frequency of fruit remains attached to the pedicel may be an indicator of domestication in archaeological remains.
Nonetheless, of all fossil evidence, the greatest confidence can be placed in pollen data. Pollen found in sedimentary deposits is sufficiently distinct and well preserved to establish the presence of a species or subspecies at a prehistoric site (Fig. 2.9). The outer wall (exine) of pollen consists primarily of sporopollenin, a material particularly resistant to decay. Pollen frequency is often used as an indicator of the relative abundance of wind-pollinated species in a region. Correspondingly, grapevine pollen frequency has been used as an indicator of ancient viticulture (Turner and Brown, 2004). For example, a pronounced increase in pollen counts was associated with increased human activity and presumptive viticulture in northern Greece after 1500 a.d. (Bottema, 1982).
Of particular interest is the difference between fertile pollen (produced by male and bisexual flowers) and sterile pollen (produced by some female flowers). Fertile pollen in Vitis vinifera is tricolporate (containing three distinct ridges) and produces germ pores, whereas sterile pollen is generally acolporate (possessing no ridges) and produces no germ pores (Fig. 2.12). Thus far, differences in the fertile/infertile pollen ratio have not been used in assessing the relative frequency of dioecious (wild) and bisexual (cultivated) vines at prehistoric sites. This may result from the poorer release of pollen from bisexual flowers and corresponding limited collection in sedimentary deposits. Nevertheless, pollen differences have been used in distinguishing the presence, or absence, of domesticated vines near ancient lake and river sediments (Planchais, 1972–1973).
As noted, it is generally believed that domestication began in or around Transcaucasia, or neighboring Anatolia, about 4000 b.c. About 8000 years ago, the climate started to become warmer and generally more humid than currently. In particular, rainfall increased considerably in eastern Anatolia. This was associated with an advance of a Quercus-dominated steppe-forest into what had been a semiarid environment. The forest reached its maximum extension about 4200 b.c. (Wick et al., 2003). Thus, growth of V. vinifera was likely more extensive in regions associated with the earliest known archaeological evidence of wine production (northwestern Zargos mountains) than today. This is also the region in which domestication of wheat (Einkorn) is thought to have begun (Heun et al., 1997). Subsequent climatic drying, beginning about 2000 b.c., slowly shifted the vegetation to match the current semiarid state, resulting in a shrinkage of the Vitis habitat. Further shrinkage of feral V. vinifera populations in recent times has been due to human habitat destruction and the decimation caused by the phylloxera outbreak that ensued after its accidental introduction in the mid-1850s. Its likely distribution, just prior to the phylloxera epidemic, is illustrated in Fig. 2.8.
Conversely, Núñez and Walker (1989) have also found evidence strongly suggestive of grapevine domestication in Southern Spain. This is long before Phoenician and Phocaean (Canaanite and Ionian Greek, respectively) colonizations, beginning about 800 b.c. They established colonies along the western coast of North Africa and the eastern Iberian Peninsula. The existence of at least two independent centers of grapevine domestication has also received molecular support from Arroyo-García et al. (2006).
Cultivars were carried westward in association with colonization. Their transplantation in Italy and southern France by ancient Greek colonists, and into France, Spain, and Germany by Roman settlers, is suggested from ancient writings. Even more certain is the implantation of a grape-growing and winemaking culture; the transport of grapevines into areas such as Israel, Egypt, and ancient Babylonia is even more indisputable. The vine is indigenous to none of these regions.
Despite human migration and its consequences, changes in seed morphology and pollen shape indicate that local grape domestication was in progress in Europe long before the agricultural revolution reached southern and central Europe. Traits such as self-fertilization and large fruit size have often been attributed to varieties brought in by human activity, but this is not requisite. Fruit size often increases under cultivation, but berry size could have also resulted from mutation in the flb gene (or possibly related genes) (Fernandez et al., 2006). Bisexuality occurs in Vitis species, albeit rarely (Fengqin et al., 1990). Thus, the occurrence of a bisexual habit in indigenous wild (sylvestris) populations (Schumann, 1974; Failla et al., 1992; Anzani et al., 1990) need not presuppose crossing between wild and domesticated vines.
Diversification of fruit color is another feature often associated with domestication. From the wide color diversity in modern grape cultivars (e.g., Plate 2.4), this property seems to have been selected for centuries. Nonetheless, one of the genetic factors leading to a loss in anthocyanin synthesis appears to have occurred millennia before domestication (Mitani et al., 2009).
Locally derived cultivars would have had the advantage of being already adapted to prevailing soil and climatic conditions. In addition, data suggest that most cultivars fall into ecogeographic groups (Bourquin et al., 1993), consistent with local origin. The presence of numerous traits (such as rounder seeds), reminiscent of V. vinifera f. sylvestris, occur in cultivars such as Traminer, Pinot noir, and Riesling. This has been construed as evidence of indigenous origin. This, of course, does not preclude the possibility of crossing and trait introgression from imported cultivars. Comparison of the genetic traits of long-established Italian varieties with indigenous feral vines is consistent with some (Grassi et al., 2003), but not all, cultivars (Scienza et al., 1994) having been derived locally. This also applies to Iberian cultivars (Arroyo-García et al., 2006). Data from a range of studies suggest that European cultivars originated from a mix of importation, introgression, and de novo domestication.
Until devastated by foreign pests and disease in the middle of the nineteenth century, wild grapevines grew extensively from Spain to Turkmenistan. Although markedly diminished at present, wild vines still occur in significant numbers in some portions of its original range, notably the Caucasus. Efforts to discover and preserve these wild populations are in progress. They are considered a genetic reserve of diversity not existent in modern cultivars. This view is supported by the greater anthocyanin diversity in wild accessions than in domesticated cultivars (Revilla et al., 2010).
Domestication probably occurred progressively, with the slow accumulation of agronomically valuable mutations. The early association of wine with religious rites in the Near East (Stanislawski, 1975) could have provided the incentive for cultivar selection, and the beginnings of concerted viticulture. Initially, cultivation would have required the planting of both fruit-bearing (female) and non-fruit-bearing (male) vines. Propagating only fruit-bearing cuttings would have resulted in a reduction or cessation of productivity, by separating female vines from pollen-bearing vines (growing wild in potentially distant, open, moist woodlands or riparian habitats). So doing would have highlighted the productivity of any self-fertile revertants, and presumably led to their selection. Viticulture also would have favored the selection of vines showing increased fruit size, as well as improved visual, taste, and aroma attributes. An obvious example might be the augmented terpene production in Muscat cultivars.
Additional support for the connection between winemaking and grapevine domestication comes from the remarkable similarity between the words for wine and vine in European languages (Table 2.1). In contrast, little resemblance exists between words for grape. The persistence of local terms for grape in European languages suggests that grapes were used (and recognized verbally) long before the introduction of viticulture and winemaking. The spread of related terms for wine and the vine may have been associated with the demic diffusion of Neolithic farmers (Chicki et al., 2002; Bramanti et al., 2009), and the associated dispersion of Indo-European languages, into Europe (Renfrew, 1989). This tendency undoubtedly was accentuated by the dispersion of Roman culture, millennia later. This, in turn, was preceded by the implantation of Greek cultural traditions and colonies into Italy and southern France beginning in the eighth and seventh centuries b.c. The development of viticulture and winemaking spread (or developed independently) in Early Bronze Age Macedonia and Crete during the third millennium b.c. (Hamilakis, 1999; Valamoti et al., 2007). One perception of diffusion routes into Europe is given in Fig. 2.13.
Table 2.1
Comparison of synonyms for ‘wine,’ ‘vine,’ and ‘grape’ in the principal Indo-European languages

Source: Information from Buck (1949).

In contrast, in Near Eastern regions, where grapevines are not indigenous, the terms for grape, wine, and grapevines all have the same root derivation (Forbes, 1965). This similarity is consistent with the likelihood that wine preceded, or coincided with, the appearance of the grapevine and winemaking in Semitic cultures. Gamkrelidze and Ivanov (1990) believe their related terms are all derived from what may have been the archetypal word for wine, woi-no.
The advanced state of cultivar variation in the southern Caucasus is also consistent with this region being associated with early grapevine domestication. Local varieties possess many recessive mutants, such as smooth leaves, large branched grape clusters, and medium-size juicy fruit. The number of accumulated recessive traits is often considered an indicator of cultivar age. Varieties showing these traits were classified by Negrul (1938) as members of proles orientalis. Also included in this group are the genetically distinct V. vinifera cultivars of China and Japan (Goto-Yamamoto et al., 2006). Evidence from Jiang et al. (2009) suggests that the first tentative cultivation of V. vinifera began in China about 300 b.c. Cultivars in the northern Mediterranean and in central Europe were viewed by Negrul as being of relatively recent origin (middle to late first millennium b.c.). This view was based on their possessing few recessive traits, and their close resemblance to wild vines. They were placed in proles occidentalis. Varieties found in contiguous regions (e.g., Georgia and the Balkans) show properties intermediate between proles orientalis and proles occidentalis. These Negrul designated as proles pontica. Characteristics of the three groupings are given in Table 2.2.
Table 2.2
Classification of varieties of Vitis vinifera according to Negrul (1938)
| Proles orientalis | Proles pontica | Proles occidentalis |
| Regions | ||
| Central Asia, Afghanistan, Iran, Armenia, Azerbaijan | Georgia, Asia Minor, Greece, Bulgaria, Hungary, Romania | France, Germany, Spain, Portugal |
| Vine properties | ||
| Buds glabrous, shiny | Buds velvety, ash-gray to white | Buds weakly velvety |
| Lower leaf surfaces glabrous to setaceous pubescent | Lower leaf surface with mixed pubescence (webbed and setaceous) | Lower leaf surfaces with webbed pubescence |
| Leaf edges recurved toward the tip | Leaf edges variously recurved | Leaf edges recurved toward the base |
| Grape clusters large, loose, often branching | Grape clusters medium size, compact, rarely loose | Grape clusters generally very large, compact |
| Fruit generally oval, ovoid, or elongated, medium to large, pulpy | Fruit typically round, medium to small, juicy | Fruit often round, more rarely oval, small to medium, juicy |
| Varieties mostly white with about 30% rosés | About equal numbers of white, rosé, and red varieties | Varieties commonly white or red |
| Seeds medium to large with an elongated beak | Seeds small, medium, or large (table grapes) | Seeds small with a marked beak |
| Fruiting properties | ||
| Many varieties partially seedless, some seedless | Many varieties partially seedless, some completely so | Seedless varieties rare |
| Varieties produce few, low-yielding fruiting shoots | Varieties often produce several, highly productive fruiting shoots | Varieties typically produce several, highly productive fruiting shoots |
| Varieties short-day plants with long growing periods, not cold-hardy | Varieties relatively cold-hardy | Varieties long-day plants with short growing periods, cold-hardy |
| Most varieties are table grapes, few possess good winemaking properties | Many varieties are good winemaking cultivars, a few are table grapes | Most varieties possessing good winemaking properties |
| Grapes low in acidity (0.3–0.6%), sugar content commonly 18–20% | Grapes acidic (0.6–1.0%), sugar content commonly 18–20% | Grapes acidic (0.6–1%), sugar content commonly 18–20% |
| Self-crossed seedling of certain varieties possessing simple leaves | Self-crossed seedings of certain varieties with dwarfed shoots and rounded form | Self-crossed seedlings of certain varieties having mottled colored leaves |

Source: After Levadoux (1956), reproduced by permission.
Additional studies along these lines were conducted by Levadoux (1956). Extensions of Negrul’s classification system can be found in Tsertsvadze (1986), and Gramotenko and Troshin (1988). The division of cultivars, and the probability of distinct centers of origin, have received modern support from Aradhya et al. (2003) and Arroyo-García et al. (2006). In addition, table grapes (more common in the Near East) show greater genetic divergence from wild grapes than do wine grapes. Table grapes also possess distinctive traits not found in wine grapes.
Cultivar Origins
Except for cultivars of recent origin, for which there is a documented record, the origin of most cultivars is shrouded in mystery. Archaeological finds are insufficiently detailed, while ancient writings do not discuss the issue. The written record of intentional plant breeding begins only in the late 1600 s, with the development of hybrid hyacinths in Holland. However, selection of improved strains of food crops (probably associated with accidental crossings) clearly goes back to the origins of agriculture (Zohary and Hopf, 2000).
For annual crops, seed collection for the next year’s crop has functioned as the principal agent for cultivar propagation, and, indirectly, development. In contrast, for perennial crops such as grapevines, vegetative propagation has been the main means by which cultivars were reproduced and variation selected. In both instances, cultivar evolution undoubtedly occurred over long periods and occurred surreptitiously. For grapevines, it was both simpler and quicker to propagate by layering (and subsequently by cuttings). Had anyone purposely planted seeds, the new vines would have had properties considerably different from those of the parent vine. Astute growers would have quickly realized that layering (or cuttings) maintained and multiplied vines possessing desirable traits.
During Roman times, it is clear that the benefits of breeding were well known, at least for animals. However, if evidence from existing European grape cultivars is correct, introgression played a significant role in their development. Crossing probably occurred accidentally, due to pollen from proximate feral vines fertilizing cultivated vines. Alternatively, the random intermixture of cultivars, typical during the medieval period, would have favored intercultivar crossing. Cultivar admixture possibly had the advantage of buffering against the vagaries of climate and soil conditions. Although the fortuitous germination and growth of seeding to maturity in established vineyards seem unlikely, both events might have occurred in small peasant kitchen gardens or outbuildings. Because fruit as well as leaves often show marked phenotypic variability (the latter property being used as the basis for ampelography), recognizing differences among vines could have been easily detected by observant growers. If intentional breeding and selection were practiced, there appear to be no records of such activity. The most likely sites for such activities would have been abbeys and noble estates with established vineyards. Monks had the advantage of being able to read ancient texts, as well as the time and writing skills to observe and collect data on vine performance. That deliberate breeding was involved seems supported by molecular evidence that permits the construction of intertwined pedigree lines, linking the parentage for several Italian and French cultivars (Fig. 2.14).

Even before molecular means of assessing cultivar origins became available, many cultivars were considered ancient. However, the evidence was dubious. There were even attempts to associate some existant cultivars with those named by Pliny, for example Greco with vitis aminea gemina, Fiano with vitis apiano, and Sciascinoso with vitis oleagina (see Thomson, 2004). During the Middle Ages, varietal designation was largely abandoned. Wine itself was principally differentiated into two categories: vinum hunicum (poor quality) and vinum francicum (high quality). These terms subsequently were applied respectively to groups of cultivars (Heunisch and Frankisch) (Schumann, 1997). Precise cultivar designation began to reappear in the fourteenth and fifteenth centuries – examples being Traminer (1349), Ruländer (1375), and Riesling (1435) (Ambrosi et al., 1994).
Name derivation has occasionally been used to suggest local origin, for example Sémillon from semis (seed), and Sauvignon from sauvage (wild) (Levadoux, 1956). However, the existence of multiple and unrelated synonyms for many European cultivars does not lend credence to name derivation as an important line of argument. Much greater confidence can be placed in DNA studies. They have provided clear evidence of the local origin of many well-known European cultivars.
Until the development of DNA sequencing, the best evidence for varietal origin came from morphological (ampelographic) comparisons. Such data were particularly useful when cultivars had diverged from a common ancestor, via somatic mutation. Examples are the color mutants of Pinot noir—Pinot gris, and Pinot blanc. Vegetative propagation maintains the traits of the progenitor, except where modified by somatic mutations. In contrast, seed propagation in highly heterozygous plants, such as the grapevine, produces progeny that can possess markedly different characteristics than their parents (Bronner and Oliveira, 1990). This tends to blur morphologic traits, making leaf characteristics suspect as indicators of varietal origin.
Another line of evidence involved chemical indicators. Isozyme and phenolic distributions are examples. However, the growing number of DNA fingerprinting techniques, such as amplified fragment-polymorphism (AFLP) and microsatellite allele (simple sequence repeat) (SSR) analysis are far more powerful and universally applicable. They circumvent some of the difficulties associated with interpreting restriction fragment-length polymorphism (RFLP) patterns, and standardization with the random amplified polymorphic DNA (RAPD) procedure. In addition, because microsatellite markers are seldom located in functional genomic regions, mutations in them tend to be evolutionarily neutral. Thus, they are unaffected by selective pressure. This permits the rapid (in terms of centuries) accumulation of variations, permitting varietal (but usually not clonal) differentiation. Markers may contain one or multiple nucleotide repeats, for example as noted below in the accompanying illustration.

Microsatellite markers also have the advantage of high polymorphism. In addition, they are inherited co-dominantly (valuable in parentage studies). For example, crossing vines possessing VVMD5 genotypes with base-pair lengths 228/238 and 236/236 could yield offspring only with 228/236 or 238/236 genotypes. When comparing the genotypes of varieties for multiple SSR markers, it is possible to predict their likely relatedness (e.g., parents, offspring, or siblings) (see Table 2.3).
Table 2.3
Partial data indicating the presumptive parentage of Madeleine Royale as a cross between Pinot and Schiava Grossa. The relationship between Madeleine Royale and Chasselas is ruled out by discrepancies (bold)
| Marker | Pinot | Madeleine Royale | Schiava Grossa | Chasselas |
| VVMD5 | 238−228 | 236−228 | 238−236 | 236−228 |
| VVMD6 | 205−205 | 214−205 | 214−211 | 212−205 |
| VVMD7 | 243−239 | 247−243 | 247−247 | 247−239 |
| VVMD8 | 143−141 | 157−143 | 167−157 | 143−143 |
| VVMD17 | 221−212 | 222−212 | 222−222 | 212−212 |
| VVMD21 | 251−249 | 249−249 | 249−249 | 266−249 |
| VVMD24 | 218−216 | 216−214 | 214−210 | 214−210 |
| VVMD25 | 253−243 | 253−245 | 259−245 | 259−245 |
| VVMD26 | 255−249 | 255−249 | 251−249 | 251−249 |
| VVMD27 | 189−185 | 189−181 | 185−181 | 189−185 |
| VVMD32 | 273−241 | 273−253 | 273−253 | 241−241 |
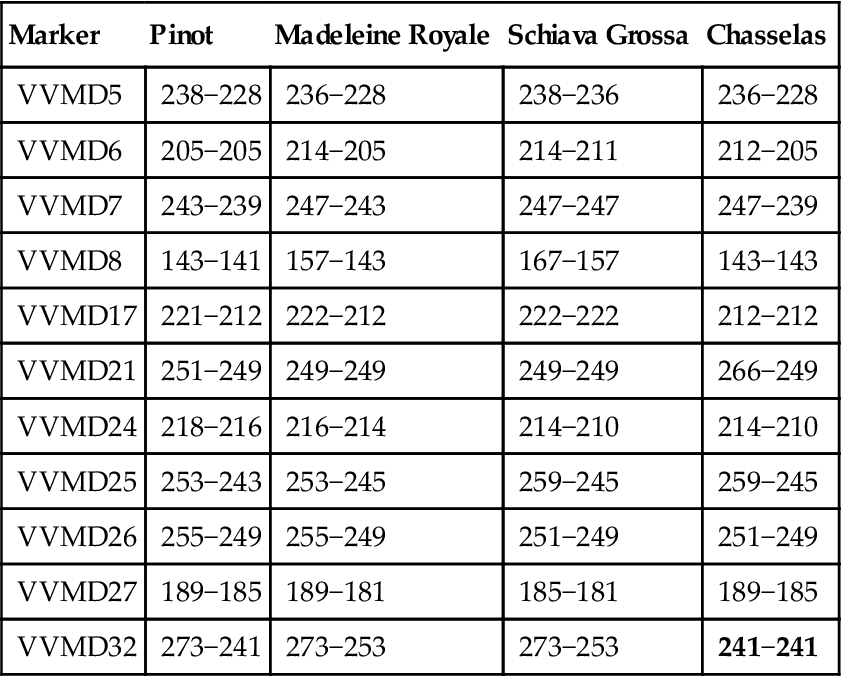
Source: Data from Vouillamoz and Arnold (2010).
Automation of microsatellite analysis, combined with the functional diploidy, heterozygosity, and the small genome of grapevines (475–500 Mb), have made SSR analysis the preferred tool in parentage and synonymy studies. The latter have been particularly useful in identifying the cultivars found in old, traditional vineyards (Jung and Maul, 2004). These have proven ‘treasure troves’ of variants of old and near-extinct varieties. Their preservation is sought as a reserve of the existant genetic diversity in cultivated Vitis vinifera. These studies are being assisted by the development of a map designating the relative position of large numbers of markers on 19 linkage groups (Vezzulli et al., 2008). Advances in this field are occurring at a dizzying pace.
Additional markers from chloroplast DNA are of particular interest in determining maternal inheritance – chloroplast DNA (as well as mitochondrial DNA) are derived only from the egg cell (Strefeler et al., 1992; Arroyo-García et al., 2002). This has permitted the identification of Gouais blanc as the maternal parent of cultivars such as Chardonnay, Gamay noir and Aligoté (Hunt et al., 2009), and Sauvignon blanc as the female parent of Cabernet Sauvignon. This determination is of more than just academic interest. Chloroplast DNA can influence properties such as chilling injury (Chung et al., 2007) and fungal toxin tolerance (Avni et al. 1992). In addition, it has attributes superior to nuclear DNA in discerning evolutionary trends, due to its low mutation rate and the absence of genetic recombination typical of nuclear DNA. These attributes have been used to identify two regions (central Italy and the Caucasus) as being ancient genomic refuges from the last European glacial period (Grassi et al., 2006); as demonstrating the extensive genetic diversity of Georgian cultivars (Schaal et al., 2010); and as denoting the likely center of diversity for many European cultivars (Imazio et al., 2006).
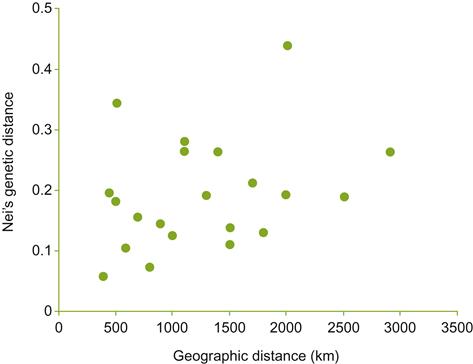
Molecular studies have also confirmed the involvement of local wild vines in the origin of many cultivars. For example, Grassi et al. (2003) demonstrated moderate genetic similarity between two Sardinian cultivars and local feral vines, suggestive of introgression. In contrast, mainland Italian varieties seem to be a mix of introduced cultivars and those introgressed with sympatric wild vines. The influence of indigenous vines on the origin of regional cultivars in western and central Europe has received support from studies by Sefc et al. (2003). Their data show that the genetic divergence among 164 cultivars was roughly correlated with their geographic separation (Fig. 2.15). Spanish and Portuguese cultivars showed the most marked similarity, followed by French/German/Austrian cultivars. Also, Vantini et al. (2003) have shown the close similarity of local Veronese cultivars. In Georgia, Ekhvaia et al. (2010) found extensive genetic similarity between local cultivars and sympatric feral vines; for example, Rkátsiteli showed 90% similarity with wild vines growing in the Lekjura Gorge. In addition, significant differentiation between Greek and Italian cultivars, as well as Italian and French ones, suggests that the importation of cultivars by Greek and Roman colonists into Italy has been exaggerated. Data from chloroplast DNA lend further support to the view that many western European cultivars were selected (bred) locally (Arroyo-García et al., 2002). Finally, Vouillamoz et al. (2006) have demonstrated the comparative genetic separation between Georgian, Armenian, and Turkish cultivars.
In contrast, several studies have provided data supportive of the importance of cultivar importation. Vouillamoz et al. (2006) had data implicative of the involvement of Georgian cultivars in the origin of several important Western European cultivars. These include Muscat, Chardonnay, Cabernet Sauvignon, Syrah, Pinot noir, Nebbiolo, and Chasselas. This view is further supported by an analysis of over 1000 French and German accessions (specifically designated and cultured clones). It implicates introgression with eastern cultivars (Myles et al., 2011). For table grapes, Dzhambazova et al. (2009) and Snoussi et al. (2004) also found data suggestive of introduction versus local domestication of Bulgarian and Tunisian table grapes, respectively. Importation also seem important in the origin of North African cultivars (Riahi et al., 2012).
Thus, not surprisingly, there appears to be no simple or common denominator to the origin of existent cultivars, some appearing to be selections from local wild vines, others being the offspring of imported and sympatric feral vines, and still others being transplanted from elsewhere, possessing no direct genetic relationship to indigenous vines.
For cultivars where presumptive parents are known, it is unknown whether the crosses occurred spontaneously or purposefully. Cultivars where parents are fairly certain include Chardonnay (Pinot noir×Gouais blanc) (Bowers et al., 1999); Cabernet Sauvignon (Cabernet franc×Sauvignon blanc) (Bowers and Meredith, 1997); and Merlot (Cabernet franc×Magdeleine Noire des Charentes) (Boursiquot et al. 2009). Other examples include Silvaner (likely the progeny of Traminer×Österreichisch weiß) (Sefc et al., 1998); Sangiovese (a crossing of the Calabrian cultivars, Ciliegiolo and Calabrese de Montenuovo) (Vouillamoz et al., 2007); and Syrah (Shiraz) (the offspring of two southern French cultivars, Dureza and Mondeuse blanche) (Bowers et al., 2000). Other suggestions, such as Pinot noir (Traminer×Meunier) (Regner et al., 2000) are possible, but have yet to be confirmed. Most researchers consider Meunier just a clonal variant of Pinot noir.
Many important cultivars appear to have involved a common parent (Fig. 2.16). Traminer appears to be one of the parents of cultivars such as Riesling, Sauvignon blanc, and Chenin Blanc. Pinot noir also appears to have parented multiple offspring. Surprisingly, a cultivar of little apparent enologic significance, Gouais blanc (syn. Weißer Heunisch), is the presumptive parent, along with Pinot, of multiple French cultivars, notably Chardonnay, Gamay noir, Aligoté, and Melon (Bowers et al., 1999).

The relative success of Gouais blanc×Pinot noir progeny is partially mirrored in the importance of Sangiovese and Garganega in the origins of many Italian cultivars (Fig. 2.17; Di Vecchi Staraz et al., 2007), and Listán Prieto in South America (Tapia et al., 2007). The latter, a little-known Spanish cultivar, crossed with Muscat of Alexandria, gave rise to two Torrontés varieties, and with Jaen B, and Negra Mole, to produce other local cultivars. Listán Prieto was important in the establishment of vineyards in the Americas, being variously called Mission, Pais, or Criolla chica in California, Chile, and Mexico, respectively.

Although the actual origins of many cultivars may never be known with certainly, due to actual parents no longer existing, DNA studies can suggest probable ancestry. The inclusion of multiple SSR markers greatly reduces the probability of false positives. Twenty-five markers are often considered a minimum. While certainty of parentage is difficult, DNA fingerprinting techniques can easily establish what is essentially impossible. For example, it has shown that Müller-Thurgau is neither the result of a Riesling self-cross, nor a Riesling×Silvaner cross (Büscher et al., 1994). Riesling and Madeleine Royale are most likely the actual progenitors of Müller-Thurgau (Dettweiler et al., 2000). Microsatellite analysis has also shown the presumed parentage of some recent Ukranian cultivars to be in error (Goryslavets et al., 2010). The technique has even revealed unsuspected aspects of the diversity and geographic distribution of Cabernet Sauvignon (Moncada et al., 2006) and Merlot (Herrera et al., 2002) clones. In the latter instance, the clone of Merlot grown in Chile is distinctly different than that typical in France.
One of the limitations of all such studies is that they must be based on existent cultivars. What potential lineages might be suggested were extinct cultivars or feral strains available for sampling will never be known. Although most cultivars thus far investigated appear to be monozygous (derived from a single seed), this does not necessarily imply genetic identity. Mutations, subsequent to origin, can give rise to a variable number of genetically identifiable clones, showing allelic polymorphy. Examples of clearly polyclonal varieties are Cabernet franc, Cabernet Sauvignon, Traminer, Riesling, Pinot noir, Chenin blanc, and Grolleau (Pelsy et al., 2010); Meunier (Franks et al., 2002; Stenkamp et al., 2009); Pinot gris, and Pinot blanc (Hocquigny et al., 2004); and Pinot noir and Chardonnay (Riaz et al., 2002). In addition, clonal divergence may also be derived polyzygotically (that is, where siblings are so morphologically similar as to be considered members of the same variety). A suspected example of a polyzygotic cultivar involves two lines of Fortana (Silvestroni et al., 1997). However, siblings are usually distinctly different, as indicated by the variety of cultivars derived from Gouais blanc×Pinot (Fig. 2.18). Additional sources of variation may arise if distinct clones are involved in multiple crosses (e.g., Pinot noir vs. Pinot blanc). Furthermore, differences can arise depending on which parent acted as the female – it being the sole origin of traits associated with chloroplast and mitochondrial2 genes.

The designation of a variant (accession), with a varietal or clonal designation, is partially subjective, depending on precedent (Boursiquot and This, 1999). If the variant is used in the sense of a subvariety, it is individually designated, for example the Musqué clone of Chardonnay. Alternatively, it may only be distinguished by a number, for example Riesling clones #239, #239-20, #239-12, and #110-1. However, if the variants make distinctly different wines, they usually go under separate but clearly related names, for example the color mutants of Pinot noir – Pinot gris and Pinot blanc. This is not a requirement, though, as some clones of Traminer used similarly possess distinct varietal designations (synonyms), e.g., Savagnin.
If the number of allelic variants can be used as a measure of cultivar age, then data from Pelsy et al. (2010) would suggest that Pinot noir and Traminer are ancient cultivars. This fits morphological attributes that give the impression that both are ancient and probably selections from wild or slightly domesticated vines. Their association as one of the parents of multiple other cultivars also supports their presumptive age. Nonetheless, based on the few accessions studied, the limited somatic diversity within Riesling clones (Pelsy et al., 2010) would be at variance with its presumptive ancient origin. However, the number of detected genotypes could easily vary, based on the number and appropriateness of the clones assessed. For example, in an investigation of 24 accessions of Traminer (Imazio et al., 2002), all were found to be genetically identical – a finding distinctly different from the observations of Pelsy et al. (2010). In addition, there is no evidence that the rate at which allelic SSR differences develop is constant, unlike point mutation rate which tends to be relatively constants over long periods of time (Drake, 1991). SSR differences appear to depend primarily on polymerase slippage, or the activity of transposons, not nucleotide substitutions induced by tautomeric shifts in the nitrogen bases that constitute primary structure of DNA.
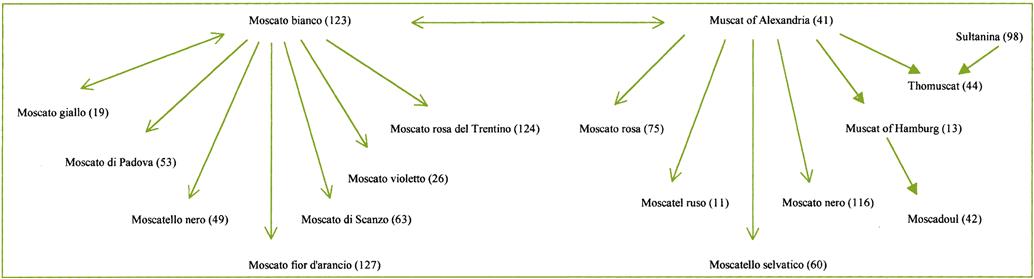
Another significant use of DNA fingerprinting has involved unraveling the often complex synonymy of cultivars. For example, Zinfandel in the United States is Primativo in Italy and Crljenak kaštelanski in Croatia (Maletiae et al., 2004). A related cultivar, Plavac mali, appears to be the progeny of a cross between another Croatian cultivar, Dobrièiae, and Crljenak kaštelanski (Zinfandel). Molecular analysis has also shown the synonymy between most Californian Petite Sirah and Durif (Meredith et al., 1999). Durif is itself the progeny of a crossing between Syrah and Peloursin. Sequence analysis has also permitted clarification of the interconnections among Muscat cultivars (Fig. 2.19).
Initially, sequencing techniques were thought to be independent of the tissues used – in contrast to isozymes and pigments, both of which are produced only in certain tissues or at particular developmental stages. However, tissue-specific DNA banding has now been identified (Donini et al., 1997). This is not overly surprising, due to the selective amplification or mutation of genes during tissue development. In addition, genetic differences found in chimeras may complicate patterns. Thus, some divergence in data interpretation among different research labs may originate from the tissues used for DNA extraction.
DNA fingerprinting has recently been applied to DNA isolated from seed (Manen et al., 2003) and other archaeological remains. Amplification has generated useful microsatellite markers from both waterlogged and charred seed (600 b.c. to 300 a.d.). DNA data are much more informative than seed morphology, which may suggest, but does not permit, the unambiguous differentiation between domesticated and wild vines. Although few useful markers have been obtained thus far, the data suggest that the specimens investigated are more closely related to varieties currently cultivated in regions where the seeds were isolated than to varieties cultivated elsewhere. These data are consistent with data noted earlier, suggesting that most European cultivars arose locally, rather than derived from elsewhere.
Regrettably, current DNA fingerprinting techniques involve markers of unknown genetic significance. For example, SSR markers appear to be located in sections of the genome other than the 4% that codes for functional (protein sequencing) genes (Lodhi and Reisch, 1995). The remaining DNA consists primarily of variously repeated segments of unknown function, transposon copies, gene regulator sections, and nonfunctional (null or pseudo-) genes. Thus, SSR relationships may have no phenotypically (selective) value. It is a moot point whether it would be preferable if relationships were based on evolutionary (ecologically) significant properties (structural genes), somewhat similar to taxonomic studies, or, as currently, on random changes in selectively neutral segments of the genome (see Jones and Brown, 2000).
Recorded Cultivar Development
Until little more than a century ago, deliberate attempts to develop new cultivars were limited (or at least unrecorded). Some of the earliest examples of suspected crosses occurred between indigenous Vitis spp. and V. vinifera cultivars planted in New England. Concord and Ives are thought to be chance crossings between V. labrusca and V. vinifera. Support for this view is provided by the presence of the Gret1 retrotransposon in several New England cultivars, including Concord (Mitani et al., 2009). This genetic insert is found in most V. vinifera cultivars, but not in North American and Asian Vitis spp. Another likely progeny of a chance crossing is Delaware – this time among three species (V. vinifera, V. labrusca, and V. aestivalis).
In contrast, Dutchess is thought to be the offspring of an intentional crossing between V. labrusca and V. vinifera. Other V. labrusca cultivars, such as Catawba and Isabella, are considered to be straight selections from local V. labrusca strains. Aside from V. labrusca, the involvement of native North American species in indigenous North American cultivars is limited. Exceptions include Noah and Clinton (V. labrusca×V. riparia), Herbemont and Lenoir (V. aestivalis×V. cinerea×V. vinifera), and Cynthiana and Norton (likely V. vinifera×V. aestivalis) (Stover et al., 2009). These early cultivars are termed American hybrids, to distinguish them from hybrids developed in France between V. vinifera and one or more of V. rupestris, V. riparia, and V. aestivalis var. lincecumii accessions. The latter are variously termed French-American hybrids, French hybrids, or direct producers. The last designation comes from their ability to grow ungrafted in phylloxera-infested soils. Some French-American hybrids are of highly complex parentage, based on subsequent backcrossing with one or more V. vinifera cultivars. Backcrossing was used to enhance the expression of vinifera-like winemaking qualities in the progeny. The relatedness of many of these cultivars has been studied by Pollefeys and Bousquet (2003).
The original aim of most French breeders was to develop cultivars containing the wine-producing attributes of V. vinifera and the phylloxera resistance of American species. It was hoped that this would avoid the expense and problems associated with grafting existing vinifera cultivars to American rootstocks. Grafting was the only effective control against the devastation being inflicted by phylloxera (Daktulosphaira vitifoliae). It also was hoped that breeding would incorporate resistance to other nonindigenous pathogens, notably powdery and downy mildew. More by accident than design, several of the hybrids proved more productive than pure vinifera cultivars. They became so popular with grape growers that by 1955 about one-third of French vineyards were planted with French-American hybrids. This success, and their different aromatic character, came to be viewed as a threat to the established reputation of viticultural regions, such as Bordeaux and Burgundy. The pressure for political action culminated in the enactment of laws intended to restrict and then eliminate their cultivation. Subsequently, similar legislation has been passed in other European countries.
Although largely rejected in the land of their origin, French-American hybrids have found broad acceptance in many northeastern regions of North America. They initially helped foster wine production in much of the continent, outside of California, and generated regionally distinctive wines. They are still thus used in regions where vinifera survival is tenuous. French-American hybrids are also grown extensively in some other parts of the world, notably Brazil and Japan.
In the coastal plains of the southeastern United States, commercial viticulture is based primarily on selections of Vitis rotundifolia var. rotundifolia. Scuppernong is the most widely known cultivar, but has the disadvantage of being unisexual. Newer cultivars such as Noble, Magnolia, and Carlos are bisexual, and so avoid the necessity of interplanting male vines to achieve adequate fruit set. Another complication of muscadine cultivars is the tendency of berries to separate (shatter) from the cluster as they ripen. Newer cultivars such as Fry and Pride show less tendency to shatter. In addition, ethephon (2-chloroethyl phosphonic acid) application can promote more uniform ripening. The resistance of muscadine grapes to most indigenous diseases and pests in the southern United States has permitted a local wine industry to develop, where the commercial cultivation of V. vinifera cultivars is difficult to impossible. Although Pierce’s disease has limited the cultivation of most nonmuscadine cultivars in the region, the varieties Herbemont, Lenoir, and Conquistador are exceptions. The first two are thought to be natural V. aestivalis×V. cinerea×V. vinifera hybrids, whereas Conquistador is a complex cross involving several local Vitis spp. and V. vinifera.
Although grafting prevented the demise of grape growing and winemaking in Europe during the late 1800s, early rootstock cultivars created their own problems. Most of the initial rootstock varieties were direct accessions of V. riparia or V. rupestris. As most were poorly adapted to the high-calcium soils found in many European regions, the incidence of lime-induced chlorosis increased markedly. Nevertheless, stock from both species rooted easily, grafted well, and were relatively phylloxera-tolerant. Vitis riparia rootstocks also provided some cold-hardiness, resistance to coulure (unusually poor fruit set), restricted vigor on deep rich soils, and favored early fruit maturity. In contrast, V. rupestris selections showed acceptable resistance to lime-induced chlorosis and tended to root deeply. Although the latter provided some drought tolerance on deep soils, these rootstocks were unsuitable on shallow soils.
The problems associated with early named rootstock selections, such as Gloire de Montpellier and St. George, led to the breeding of new hybrid rootstocks (Fig. 2.20), incorporating properties from several Vitis species. For example, V. cinerea var. helleri (V. berlandieri) can donate resistance to lime-induced chlorosis; V.×champinii can supply tolerance to root-knot nematodes; and V. vulpina (V. cordifolia) can provide drought tolerance under shallow soil conditions. Alone, however, these species have major drawbacks as rootstocks. Both V. cinerea var. helleri (V. berlandieri) and V. vulpina root with difficulty; V. mustangensis (V. candicans) has only moderate resistance to phylloxera; and both V. mustangensis and V. vulpina are susceptible to lime-induced chlorosis. Important cultural properties of some of the more widely planted rootstocks are given in Tables 4.6 and 4.7.

 =interspecies crossings. The Champin varieties, Dog Ridge and Salt Creek (Ramsey), are considered to be either natural V. mustangensis×V. rupestris hybrids, or strains of V.×champinii. Solonis is considered by some authorities to be a selection of V. acerifolia (V. longii), rather than a V. riparia×V. mustangensis hybrid. On this basis, some multiple-species hybrids noted above are bihybrids. (Modified from Pongrácz, 1983, reproduced by permission.)
=interspecies crossings. The Champin varieties, Dog Ridge and Salt Creek (Ramsey), are considered to be either natural V. mustangensis×V. rupestris hybrids, or strains of V.×champinii. Solonis is considered by some authorities to be a selection of V. acerifolia (V. longii), rather than a V. riparia×V. mustangensis hybrid. On this basis, some multiple-species hybrids noted above are bihybrids. (Modified from Pongrácz, 1983, reproduced by permission.)Grapevine Improvement
As with other perennial plants, and especially due to the small size and minor morphologic differences between the chromosomes (see Fig. 2.5), little data on the genetic properties of Vitis had accumulated. This gap is now being filled by genome sequence analysis (Jaillon et al., 2007; Velasco et al., 2007). Figure 2.21 represents the Vitis genome, noting the chromosomal location of many genes. It is estimated that the Vitis genome contains some 30,000 protein-coding genes. Preliminary data suggest that as many as 13% of the genes are heterozygous (Velasco et al., 2007). In addition, grapevines possess an abnormally high number of isogenes. For example, there are 43 stilbene synthase, 89 functional terpene synthase, and two geranyl diphosphate synthase genes (more than twice as many as most other plants) (Jaillon et al., 2007). These genes are particularly important to grapevine health and flavor characteristics. Stilbene synthases are involved in the production of resveratrol and other antimicrobial phytoalexins; terpene synthases are responsible for the production of aromatic terpenes; and geranyl diphosphate synthases are critical in the production of terpene progenitors. Knowledge of the location of genes should facilitate varietal improvement using marker-assisted breeding (Mackay et al., 2009). This could be particularly used in improving disease resistance.

Standard Breeding Techniques
The focus of most grape breeding has changed little since the early work of breeders such as F. Baco, A. Seibel, and B. Seyve (Neagu, 1968). Standard goals include improving the agronomic properties of rootstocks, and enhancing the viticultural and winemaking properties of fruit-bearing (scion) stock. The major changes involve the availability of modern analytical techniques and a better understanding of genetics. Such knowledge can dramatically increase a breeder’s effectiveness.
Of breeding programs, those developing new rootstocks are potentially the simplest. Improvement often involves enhancing properties, such as lime and drought tolerance or pest resistance. When one or a few genes are involved, incorporation may require only the crossing of a desirable variety with the source of the new trait, followed by selection of offspring possessing the trait(s) desired (Fig. 2.22A). Backcross breeding (Fig. 2.22B) can be particularly useful when integrating single dominant traits into an existing cultivar.

Developing new scion (fruit-bearing) varieties is far more complex. The repeated backcrossing, required to re-establish expression of desirable traits, may result in diminished seed viability and vine vigor, a phenomenon termed inbreeding depression. It occasionally can be countered if more than one cultivar can serve as a recurrent parent, or if embryo rescue is possible. Embryo rescue involves the isolation and cultivation of seedling embryos on culture media, which otherwise would abort during seed development (Spiegel-Roy et al., 1985; Agüero et al., 1995). Alternatively, inbreeding depression (Charlesworth and Willis, 2009) may be countered by treating germinating seeds with a demethylating agent, if recent findings on Scabiosa are confirmed in other plants (Pennisi, 2011). Gene inactivation is often associated with DNA methylation.
However, an even more intractable problem in breeding new grapevine scions is the highly heterozygous nature of most desirable traits (Plate 2.4). As a consequence, breeding tends to disrupt the complex balance that donates a variety’s desirable attributes. In addition, most of the properties associated with commercial success are quantitative, depending on the accumulative action of multiple genes, often located on separate chromosomes. Thus, the disruption of desirable combinations is particularly likely during recombination and the random assortment of chromosomes that occurs during meiosis. Gene reassortment also complicates the selection process, by blurring the distinction between environmentally and genetically induced variation.
The problem may be partially diminished if the genetic basis of the trait is known. For example, selection for cold-hardiness might be facilitated if the nature, location, and relative importance of the regulatory genetic loci were known. Individual aspects of this particular complex property involve diverse features, such as the control of cellular osmotic potential (timing and degree of starch/sugar interconversion), the unsaturated fatty-acid content of cellular membranes, and the population of ice-nucleating bacteria on leaf surfaces. This knowledge would permit each aspect to be individually assessed and selected for during breeding.
Chemical indicators and other genetically linked features are especially useful in the early selection of suitable offspring. The efficient and early elimination of undesirable progeny in a breeding program assures that precious time and resources are spent only on potentially useful offspring. For example, the presence of methyl anthranilate and other volatile esters was used in the early elimination of progeny possessing a labrusca fragrance (Fuleki, 1982). Improved knowledge of the factors associated with color stability in red wines would be helpful in the early selection of offspring possessing better color retention attributes. Selection for disease resistance would be both sped up and facilitated were this property assessable in seedling by a simple chemical test. Regrettably phytoalexin production and toxin resistance are often stage-specific and not necessarily adequately expressed in seedlings. In addition, laboratory trials frequently do not adequately simulate field conditions.
Another factor that can facilitate breeding is combining ability. This somewhat nebulous term refers to a strain’s relative success in breeding programs. Regrettably, combining ability is currently impossible to assess in advance. Its genetic basis is unknown. Nonetheless, it is suspected to be partially associated with the genetic difference between the parents. A possible example of combining ability may involve Gouais blanc and Pinot noir. Many important cultivars have them as their parents (see Fig. 2.18). Gouais blanc is suspected to be an ancient Croatian cultivar whereas Pinot noir is often viewed as a strain selected from wild vines, or introgressed strains retaining many feral characteristics.
Adding to the complexities of grape breeding is the time required for winemaking attributes to become assessable. Vines only begin to bear sufficient fruit for testing in their third year. Increasing the population of a potentially new cultivar to a point where features such as winemaking potential can be adequately evaluated can often take decades.
The length of a breeding program can, at least potentially, be shortened by inducing precocious flowering (Mullins and Rajasekaran, 1981). This especially applies to properties assessable with small fruit samples. Flowering can be induced within 4 weeks of seed germination, by the application of synthetic cytokinins such as benzyladenine or 6-(benzylamino)-9-(2-tetrahydropyranyl-9H). It induces tendrils to develop into flower clusters (Fig. 2.23). This shortens the interval between crosses. Generation cycling can be further condensed by exposing seed to peroxide and gibberellin, prior to chilling (Ellis et al., 1983). The treatment shortens the normally prolonged cold treatment required for seed germination. In theory, the combination of precocious flowering and shortened seed dormancy could reduce the generation time from 3–4 years to about 8 months. Inducing dormant cuttings to flower is another means of accelerating the breeding process. With a greenhouse, crossing is not limited by seasonal conditions, and could be conducted year-round.

Although clear indicators of gene expression are highly desirable, few agronomically important genetic traits are amenable to early detection. Assessing properties, such as winemaking quality, can easily extend beyond the career of an individual breeder. The long gestation period for generating new scion varieties is further complicated by consumer conservatism and regulatory intransigence. In Europe, most appellations rigidly limit cultivar use, effectively legislating against innovation. This automatically places new cultivars under legal and marketing constraints. Restrictions are even more severe when involving interspecies crosses, despite many hybrids possessing vinifera-like aromas (Becker, 1985). Correspondingly, incorporating new sources of disease and pest resistance into existing V. vinifera cultivars is essentially precluded using standard techniques. Although species other than V. vinifera are the primary sources of disease and pest resistance, untapped resistance may still exist within existing V. vinifera cultivars. For example, the offspring of a crossing between two Riesling clones (Arnsburger) showed resistance to Botrytis not apparent in either clonal progenitor (Becker and Konrad, 1990).
Despite North American Vitis species having been historically used as the primary source for new traits in grapevine breeding, Asian species, such as V. amurensis and V. armata, also possess desirable traits. For example, V. amurensis (and some strains of V. riparia) are potential sources of mildew and Botrytis resistance (along with V. armata in the latter case). In addition, V. amurensis and V. piasezkii (along with northern races of V. riparia) are valuable sources of cold-hardiness, early maturity, and resistance to coulure. For cultivation in subtropical and tropical regions, sources of disease and environmental stress resistance include V. aestivalis and V. shuttleworthii (from the southeastern United States and Mexico), V. caribaea (from Central America) (Jimenez and Ingalls, 1990), and V. davidii and V. pseudoreticulata (from southern China) (Fengqin et al., 1990).
Most interspecific hybridization has involved crossings within the subgenus Vitis. However, the production of partially fertile progeny between V. vinifera×V. rotundifolia is encouraging. Success is limited both by the inability of Vitis pollen to penetrate the style of muscadine grapes (Lu and Lamikanra, 1996), and the aneuploidy caused by unbalanced chromosome pairing during meiosis (Viljoen and Spies, 1995). Backcrossing to one of the parental species (usually the pollen source) has been used in several crops to restore fertility to interspecies hybrids. Although crossing bunch with muscadine grapes is far from simple, V. rotundifolia is the principal source of Pierce’s disease resistance, as well as resistance to downy and powdery mildew, anthracnose, black rot, and phylloxera; tolerance to root-knot and dagger nematodes; and enhanced heat insensitivity. Advances in vegetatively propagating V. vinifera×V. rotundifolia hybrids may enhance their commercial availability (Torregrosa and Bouquet, 1995) and adoption. Similar breeding programs could also introduce V. vinifera-like winemaking properties into muscadine cultivars.
Although most interspecies hybrids possess aromas distinct from those characteristic of V. vinifera cultivars, their aroma is not necessarily less enjoyable. Their distinctiveness could form the basis of regional wine styles. It was the regional distinctiveness of certain European wines that historically provided them with much of their appeal. There is no rational reason why this could or should not occur in North America or elsewhere. Regrettably, tradition-bound critics and many connoisseurs are inordinately opposed to change. Acceptable flavors seem to have become petrified with a few ‘premium’ cultivars. This situation limits the choice of wine flavors available to a new, and initially unbiased, generation of wine drinkers. It also restricts the spectrum of flavors available to all consumers. The associated boredom may be one of the root causes of stagnated wine sale worldwide.
Reticence to new sensory traits restricts the options in most grape-breeding programs. Traditional views demand that progeny (even of self-crosses) be given a new name. Thus, new cultivars lose the marketing advantage of their parents’ names. Most consumers are neophobic, generally viewing wines possessing unfamiliar names or places as inferior. This may also stem from new cultivars being relegated to regions where premium varieties do not grow profitably, or where increased yield or reduced production costs offset the loss of varietal-name recognition. The increasing popularity of ‘organically grown’ wines may enhance the acceptance and cultivation of new varieties. New cultivars often require less pesticide and fertilizer use than existing cultivars.
Modern Approaches to Vine Improvement
Although standard breeding techniques can be successful, they usually involve decades of work, much effort, and considerable expense. There is also, with wine grapes, the serious complication associated with disrupting the allelic combination that gives each cultivar its unique traits. In contrast, genetic engineering can introduce selected genes, without disrupting critical varietal traits. Consequently, the central commercial attributes of the cultivar remain unchanged.
Although many desirable traits, such as those regulating disease resistance and the synthesis of aromatics, may be associated with one or a few dominant traits, many others, such as yield/quality ratio, cold-hardiness, and overall flavor, are under multigenic control and/or recessive. Thus, they are not easily identified or localized, features requisite to facilitate transfer. Dominant traits, under monogenic control, are better candidates. Examples are Run1, Ren1 and PdR1. They donate resistance to powdery mildew in Vitis rotundifolia, V. vinifera, and Pierce’s disease in V. arizonica/candicans, respectively.
Advances in molecular genomics are providing genetic maps, showing the location of these genes, and adjacent SSR and SNP-based markers (Velasco et al., 2007; Di Gaspero and Cattonaro, 2010). The latter permit marker-assisted breeding (Mackay et al., 2009), where the adjacent sites facilitate recognition of the successful insertion of the desired gene(s). Embryos or seedlings not possessing the desirable gene(s) can be discarded early, saving time, effort, and space. The procedure also streamlines traditional breeding programs, by clearly identifying plants likely to possess the desired traits for subsequent crossing (Frisch and Melchinger, 2005). By directly assessing for the gene’s physical presence (without having to detect its phenotypic expression), marker-assisted selection avoids difficulties due to the masking of desirable, but recessive, traits by the presence of a dominant, but undesirable, allele. What may be missed, though, are epistatic or other allelic interactions. These may be crucial in gene expression, or present unsuspected and unsuitable pleiotrophic influences. These may be partially predicted with techniques such as metabolomics.
Metabolomics embraces advances in analytical chemistry that permit the simultaneous assessment of multiple metabolites (Fernie and Schauer, 2009). It associates phenotypic traits, notably those that affect berry chemistry, with the genetic characteristics of specific cultivars, the progeny of designed crosses, or changes induced by genetic transduction. The breeder can obtain a broad image of the effects of particular genes and their interaction within a specific genetic and environmental background. This is particularly valuable when attempting to understand the complexities of secondary metabolites, and the flavor chemistry that affects wine attributes. Metabolomics is also useful in enhancing our understanding of the metabolic pathways leading to varietal uniqueness and its genetic basis.
More restricted in application is chlorophyll fluorescence imaging. It can visually measure the photosynthetic efficiency of individual plants (Baker and Rosenqvist, 2004). This, in turn, can be used as an objective measure of vine health in assessing resistance to various biotic and abiotic stress factors.
Additional technical improvements have involved the isolation, amplification, and insertion of genes (Vivier and Pretorius, 2000; Pretorius and Høj, 2005). Insertion has typically used either the vector, Agrobacterium (Rhizobium), or biolistics (particle bombardment) (Colova-Tsolova et al., 2009). Viticultural examples have included incorporation of the GNA gene from Galanthus nivialis (snowdrop) and a protein-coat gene for the grapevine fanleaf virus (GFLV) (Mauro et al., 1995). They were inserted to enhance resistance to fanleaf degeneration, by either reducing feeding by viral-transmitting nematodes, or preventing infection directly, respectively. Alternatively, the addition of a superoxide dismutase gene from Arabidopsis thaliana has been incorporated into scion varieties to improve cold resistance.
Protocols for genetic transformation using Agrobacterium (Rhizobium) as the gene carrier are described in Agüero et al. (2006) and Bouquet et al. (2006). Despite some success, recent discoveries in genetics, relating to the widespread occurrence of pseudogenes and retrotransposons, suggest that progress may be fraught with unsuspected hurdles. In addition, as noted, most important attributes appear to be under accumulative (quantitative, multigene) control. If these are not closely linked on a single chromosome, successful transfer and expression is likely to be especially difficult.
The widespread application of transduction is even more dependent on solving practical problems associated with its use with grapevines (Tapia et al., 2009; Vidal et al., 2009). Currently, these include the low numbers of embryos that differentiate from cells in tissue culture and development into plantlets; integration of the gene into a durable state within host cells; and expression of the trait in whole plants. Several stages in plantlet generation are shown in Plate 2.5.
Another issue, often associated with propagating vines from tissue culture, is phenotypic variability. This is frequently no more than the expression of juvenility. Atypical phenotypic expression often decreases with vine age, or with repeated propagation (Mullins, 1990). It apparently can be minimized by using buds derived from the terminal ends of long canes (fortieth node onward), and grafting onto a desirable rootstock (Grenan, 1994). Desirably long shoots are most easily obtained from vines grown under greenhouse conditions.
Additional problems, not limited to grapevines, relate to the retention of only transformed cells in tissue culture. This often involves the simultaneous incorporation of resistance genes to traits that will inhibit the growth of all but the transformants. Due to public distrust of their presence, techniques are being developed to eliminate these traits after the initial selection process (Vidal et al., 2006; Dutt et al., 2008). This should eliminate one of the more common concerns expressed by those opposed to genetic engineering. Amazingly, those who would benefit most from genetic engineering are those most opposed to its use – producers and consumers of organic produce. The incorporation of disease and pest resistance would greatly reduce the need for control agents.
Even if GM vines are never cultivated commercially, data obtained by current attempts should add vital new information about grapevine genetics (Thomas et al., 2003). Basic information about issues such as the nature of disease resistance is essential if efforts toward reducing the environmental impact of disease control are to be successful. It should also facilitate the streamlining of traditional breeding procedures.
Clonal Selection
Although genetic engineering possesses the greatest potential for significant improvement, clonal selection remains the principal means by which grape growers can choose specific traits, without modifying the variety’s essential characteristics (and name). However, improvements are limited by the genetic variation that exists within the cultivar. This variation consists of gene mutations and epigenetic modifications that have accumulated since the cultivar’s origin. The most visually obvious mutations are those that affect grape pigmentation or vine morphology. Nevertheless, most clonal mutations have more subtle effects, such as modifying enzymes that affect aromatic character, or having influences on yield, disease resistance, or climatic sensitivity (Fig. 2.24).
Most mutations arise from random changes in the nucleotide sequence of genes. Because most genes encode the structure of enzymatic proteins, mutations often result in the production of isozymes (functional and/or structural variants of an enzyme). Other mutations can result from phenomena such as gene inversions, translocations, or the insertion of retrotransposons. Once a mutation has occurred, it will be reproduced essentially unmodified in cuttings. Because mutations tend to accumulate slowly, older cultivars have a greater tendency to possess mutations than those of more recent origin. Each mutated strain can be considered a distinct clone (a cultivar with a modified genotype). Old cultivars such as Pinot noir consist of a wide diversity of more or less readily differentiable clones.
Another source of clonal variation, usually undesirable, results from infection by endogenous pathogens. One of the principal initial functions of clonal selection is to eliminate these pathogenic agents. They typically limit the potential of the cultivar (Credi and Babini, 1997). In contrast, some endophytes may be beneficial. Some can donate desirable traits, such as increased salt tolerance (Rodriguez and Redman, 2008).
Clonal selection normally refers to a series of procedures designed to identify, isolate, propagate, and provide premium vine stock to grape growers. Selection usually goes through several steps (Fig. 2.25), where cuttings are multiplied and repeatedly assessed for their viti- and vini-cultural traits, and cleared of systemic pathogens.

Because clonal selection depends on genetic variability within a cultivar, it is particularly useful with older varieties. Depending on the age and genetic stability of the cultivar, it may possess few to many somatic mutations. This is especially true with monozygotic cultivars (derived from a single, fertilized, egg cell). Polyclonal varieties (derived from a collection of related individuals) are usually more variable, as they possess not only mutations that have accumulated in the individual lines, but also genetic differences inherent from the origin of the various lines.
As noted, there is no consistency in how clonal differences are designated. Color and aromatic differences have often been recognized at the varietal-name level, for example the gray and white variants of Pinot noir – Pinot gris and Pinot blanc, and the Musqué clone of Chardonnay. Other variants have been designated as biotypes, as with many Sangiovese clones (Calò et al., 1995). In other instances, clones have been grouped relative to their possession of similar traits. In Pinot noir, these may be grouped into the Pinot fin – trailing, low-yielding vines, with small tight clusters; Pinot droit – higher-yielding vines with upright shoots; Pinot fructifer – high-yielding strains; and Mariafeld-type strains – loose clustered, moderate-yielding vines (Wolpert, 1995). More recently, though, clones are designated only by accession numbers. These are usually supplied by the agency that isolated the clone, or national collection, such as those at the University of California, Davis, or the Institut National de la Recherche Agronomique in France. These preserve the clones for distribution to nurseries that propagate them for commercial release.
Because viruses and other systemic pathogenic agents invade most tissues, selection alone may be unable to eliminate these agents. Thermotherapy has been particularly valuable in this regard. One version involves exposing dormant cuttings to high temperatures (~38 °C) for several weeks. Alternatively, excision and micropropagation of vines from small portions of bud tissue can eliminate viruses that do not invade the apical, meristematic tissue. Nevertheless, isolating clones free of all known systemic agents (bacterial, phytoplasmal, viral, and viroidal) is difficult. In addition, reinfection often occurs where the agent is well established in a region. Another, more intractable, problem is that some systemic agents may not provoke readily recognizable disease symptoms. Thus, their presence may go unnoticed and the vine remains a reservoir of infection for sensitive cultivars. Detection may be possible only with immunological or nucleic acid analysis.
For rootstock varieties, clonal selection is often limited to certifying them as disease-free. Due to recent origin, insufficient time has elapsed for the accumulation of somatic mutations.
In addition to basic traits, genetic stability and phenotypic consistency are essential. Correspondingly, elimination of phenotypic variability within a clone is an important goal of clonal selection. Regrettably, detection of such instability is not necessarily simple, expression often depending on local environmental conditions. For example, the muscat character of some Chardonnay clones can vary from year to year, depending on fruit maturity (Versini et al., 1992). To compensate for this, clonal selection trials usually occur at a range of sites and over many years. Nonetheless, prudent vineyard owners conduct their own, on site, comparative assessments prior to making major planting commitments. Growers also need to be aware that clonal traits may require adjustment in viticultural practice to achieve the benefits desired. These may include modification to the rootstock or training system employed.
Most instances of clonal instability are due to chimeric mutations – genetic modifications that occur only in particular layer(s) of the apical meristem (Franks et al., 2002). These may involve point (single-nucleotide) mutations, duplications, substitutions, or deletions. As a result, chimeras possess outer (L1) and inner (L2) tissues that differ in one or more alleles (Fig. 2.26). Separate layers may be homozygous (aa) or heterozygous (Aa); possess polymorphic versions of an allele (a1a1 vs. a1a2); possess three copies (Aaa) vs. the regular two (Aa); or possess some other variant combination.
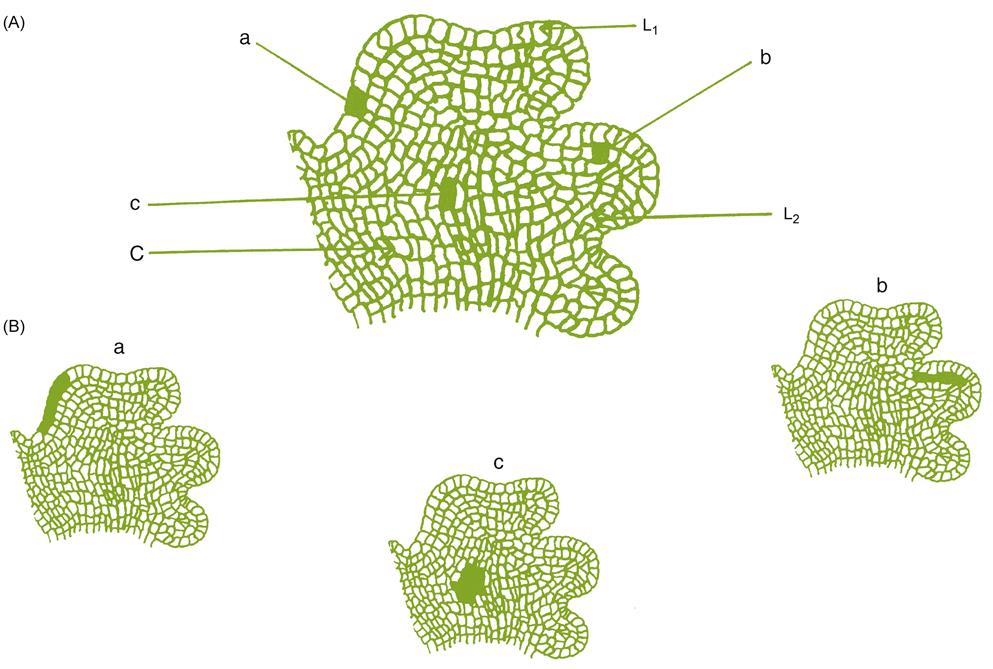
Apical meristematic cells in the outer, L1, layer (tunica) give rise to the epidermis, and divide only perpendicularly to the surface. The inner, L2, tissues give rise to a, more or less defined, inner tunica layer and corpus. Somatic mutation in either the L1 or L2 can produce a periclinal chimera (see Table 2.4 for examples). As the bud grows and differentiates into various structures of the vine, these genetic differences are propagated throughout the shoot.
Table 2.4
Microsatellite alleles detected in leaf, wood and root tissue
| Grape type | Clone | Marker | Alleles | Layer in which mutation occurs | ||
| Leaf | Wood | Root | ||||
| Pinot noir | CTPS 115 | VMCNg2fl2 | a, b, X | a, b | a, b | L1 |
| CTPS 386 | VMCNg2fl2 | a, b, Y | a, b | a, b | L1 | |
| VMClg3.2 | a, b, X | b, X | b, X | L2 | ||
| VMCle8 | a, b, X | b, X | b, X | L2 | ||
| Chardonnay | 16 | VMC2fl2 | a, b, X | a, b | a, b | L1 |
| S17 | VMCNglh7 | a, b, X | a, b | a, b | L1 | |
| CTPS 96 | VMC5g7 | a, b, X | a, b | a, b | L1 | |
| VMC6c 10 | b, X | b, X | b, X | L1, L2 | ||

a and b indicate the two standard alleles whereas X and Y are variant alleles. Source: From Riaz et al. (2002), reproduced by permission.
Because adventitious roots originate from the interfascicular cambium (Favre, 1973), epidermal chimeric traits are not found in the roots. In addition, epidermal chimeric traits are not transmitted sexually (via seed). Only internal cells are positioned such as to potentially generate gametes. This situation can change, though, if the normal L1 and L2 distinction is disrupted, as when tertiary buds develop, due to death of the primary and secondary buds, for example from herbivory (Marcotrigiano, 2000) or frost damage. Tissue layer disruption also results when cells are propagated in liquid culture. On embryogenesis, former epidermal cells may be located internally and potentially give rise to gametes.
Although chimeras can produce clonal instability, most are stable. Examples include the color variants of Pinot noir – Pinot blanc and Pinot gris (Hocquigny et al., 2004); the bronze and white variants of Cabernet Sauvignon – Malian and Shalistin, respectively (Plate 2.6); and the hairy leaf phenotype of Pinot Meunier (Boss and Thomas, 2002). Pale-colored chimeric clones tend to produce anthocyanins only in the epidermis (not in the hypodermis), whereas white chimeric clones produce anthocyanins in neither tissue. Plate 2.7 illustrates an example of an unstable epidermal chimera.
In addition to concerns about systemic pathogens and phenotypic stability, improved crop yield and grape quality are central to all clonal selection procedures. Improved yield, if associated with increased vegetative vigor, can result in reduced grape quality. This undesirable correlation, if found, can often be avoided by appropriate adjustment in viticultural practice, such as a more open canopy, basal leaf removal, or increased planting density. In addition, increased yield is not necessarily associated with reduced quality (e.g., sugar content) (Fig. 2.27). In a study of Riesling clones, the clone that showed both the highest yield and °Brix values showed the weakest growth (Schöfflinger and Stellmach, 1996). Such clones have the potential for combining the economic and environmental benefits of reduced fertilizer inputs with maximized yield and quality.
In clonal selection, quality is always paramount. Unfortunately, there are no universally accepted quality benchmarks. Brix and acidity values are standard indicators, but are by themselves insufficient. Measures of aroma potential are still insufficiently precise, and anthocyanin assessment complex and not a direct gauge of wine color intensity or stability. There is also no clear or simple correlation between these different factors. Typically, microvinification tests are conducted over several years from vines grown at different locations. The resultant wines need to be aged and assessed by a panel of experienced (and consistent) wine judges (McCarthy and Ewart, 1988). Regrettably, ‘quality’ as perceived by wine judges and winemakers is not necessary equivalent to that perceived by wine consumers. These demands make the establishment of significant differences long, complex, and costly. Nonetheless, it can be worth the effort as clones can differ in yield by as much as 60%.
Other features frequently integrated into clonal selection involve factors such as unique varietal aromas, for example, Chardonnay clones 77 and 809 with a muscat-like nuance (Boidron, 1995); differences in winemaking potential, for example, Gamay 222 vs. Gamay 509 used in the production of the nouveau vs. cru beaujolais wines (Boidron, 1995); selection of Pinot fin clones for red burgundy vs. Pinot droit clones for champagne production (Bernard and Leguay, 1985); choice of growth habit – erect for mechanical harvesting; preferences in berry shape and size that can affect flavor and color potential (Watson et al., 1988); and factors such as berry-cluster morphology or epidermal wax features that can affect resistance to pathogens or physiologic disorders.
One of the great successes in clonal selection, other than disease-freeness, has been providing growers with an increasing palate of subtle differences from which to choose. In addition, their physiological uniformity enhances a grower’s potential to produce a crop that is more consistent in ripeness and quality. Achieving this goal is also facilitated by precision viticulture, where field variables are minimized. These aims are based on the belief that wine quality is enhanced by crop uniformity. Vine homogeneity can also facilitate daily vineyard activities. If every vine is at the same stage, and bears its clusters identically, the timing and dosage of irrigation, fertilizers, and crop protectants should be more efficient and cost-effective.
Although clonal selection has many benefits, varietal variation must not be eroded in the process. Correspondingly, less desirable lines are often retained as a library of diversity, for potential future use.
Although uniformity is generally viewed as desirable, it could limit wine aromatic complexity, or exaggerate environmental sensitivity. This could be especially important where yearly climatic fluctuations are marked. Thus, the phenotypic plasticity, associated with the planting of several clones, can be a judicious form of crop insurance (Hubáčkova and Hubáček, 1984). Boidron (1995) recommends at least two clones in small plots, and up to five or six clones in large vineyards.
Another factor that may need consideration is vineyard fumigation. This is especially of concern where soil-borne viral diseases (transmitted by parasitic nematodes) is a problem. The alternative of leaving the soil fallow for 6 or more years is rarely a commercially viable option. The use of nematode-resistant rootstocks, where available and acceptable, is also advisable.
Somaclonal Selection and Mutation
The elimination of viral infection, in association with clonal selection, has made significant improvements in the planting material available to grape growers. However, further significant advances will depend on the generation or incorporation of new genetic variation (via mutation or genetic engineering), or other means by which existing variation can be enhanced (Kuksova et al., 1997). Somaclonal selection can enhance the expression and isolation of clonal variation, whereas exposing meristematic tissue or tissue culture to mutagenic chemicals or radiation can induce new variation (Fig. 2.28).

Somaclonal selection places tissue culture under conditions that can be adjusted to favor or permit only the growth of cell lines with desired attributes. For example, lines possessing tolerance for salinity or fungal toxins (Soulie et al., 1993) may be isolated by exposing cells to these conditions in culture media. However, the tolerance selected is occasionally not expressed in whole plants regenerated from the culture (Lebrun et al., 1985). This results when selection involves only cells that have temporarily adapted to, not genetically modified to, the culture conditions used. Genetic engineering also employs selective culture techniques to isolate transformed from nontransformed cell lines (Mauro et al., 1995; Kikkert et al., 1996).
Liquid tissue-culture conditions may also be used to disrupt the normal distinction between L1 and L2 cells (Skene and Barlass, 1983). As a result, callus tissue, derived from either cell line, can potentially generate embryos. This permits the isolation of plantlets that possess, or lack, somatic mutations throughout the regenerated vine. For example, strains have been developed from L1 cells of a chimeric strain of Chardonnay 96 (Bertsch et al., 2005), and a dwarf mutant strain of Meunier (Boss and Thomas, 2002). Another consequence is the opportunity to transmit traits, formerly isolated in the epidermis, via sexual reproduction (gametes being derived only from cells beneath the epidermis). Conversely, somaclonal selection could be used to eliminate undesirable chimeric traits.
Grapevine Cultivars
With the number of named grapevine cultivars approaching 15,000 (many of which are synonyms), a comprehensive system of cultivar classification would be useful. Regrettably no such system exists. In some countries, there have been attempts to rationalize local cultivars into related groups (Fig. 2.29). Most ecogeographic associations have been based on ampelographic attributes. Because of the localized distribution of each group, they may have been derived from one or a few related individuals. This view has received support from microsatellite DNA (SSR) analysis (see above). For example, most French cultivars appear to be distinct and partially related to wild accessions. As well, Armenian, Georgian and Turkish varieties appear to be isolated genetically (Vouillamoz et al., 2006). Nevertheless, the same study found evidence that some Western European cultivars (Chasselas, Nebbiolo, Pinot noir, and Syrah), or their ancestors, appear to possess traits derived from Georgian cultivars.

Cultivar nomenclature is further complicated by the absence of an international agreement to oversee grape cultivar designation. Thus, cultivars considered identical by ampelography may be nonuniform genomically. The situation could be put on a more solid founding if cultivars were associated with type specimens, as are most species names. For example, cultivar nomenclature could be associated with a specific genomic sequence. The same would be useful in clonal designation.
Former methods of assessing relatedness have involved comparing aroma profiles, for example Fig. 2.30. Numerical taxonomic procedures have also been used to reduce the subjectivity inherent in traditional classification techniques (Fanizza, 1980). Although DNA techniques supply more objective data than all older techniques combined, they still involve human interpretation and all its associated potential problems.
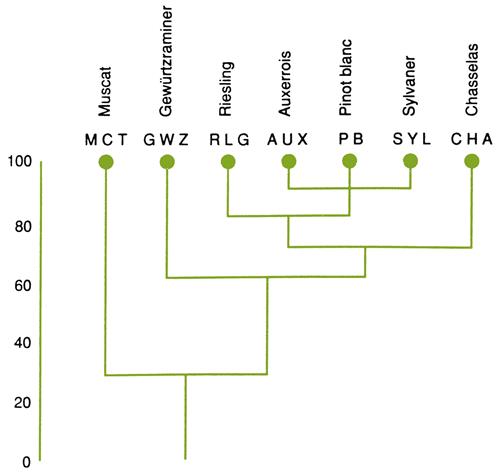
Nevertheless, for the foreseeable future, for pragmatic reasons, ampelographic procedures (Galet, 1979) will probably remain the primary means by which cultivars are identified, despite the plasticity of the traits employed. Ampelography uses vegetative characteristics, such as leaf shape, veinal branching patterns, pubescence (hairiness), and other features for identification. The development of a computer-aided digitizing system for determining ampelographic measurements (AmpeloCAD) will hopefully simplify grapevine identification for the nonampelographer (Alessandri et al., 1996).
On a broader scale, cultivars are grouped according to their specific or interspecific hybrid origin. Most commercial varieties are pure V. vinifera cultivars. French-American hybrids constitute the next largest group. They were derived from crosses between V. vinifera and one or more of the following: V. riparia, V. rupestris, and V. aestivalis. Early American cultivars are either selections from indigenous grapevines, or are hybrids between them and V. vinifera. Interspecific cultivars generally refer to modern (twentieth century and later) crosses between V. vinifera and species such as V. amurensis, V. riparia, V. armata, and V. rotundifolia. In most instances, their precise parentage is known, and where all due caution was taken to avoid selfing or unintended crossing.
Vitis vinifera Cultivars
Because of the huge number of named cultivars, only a few of the better-known varieties are discussed here. Although often world famous, these cultivars seldom constitute the principal varieties grown, even in the country of origin. Productivity is usually lower, and cultivation more demanding. Their reputation comes from their wines possessing distinctive varietal aromas, frequently combined with long-aging potential. In favorable locations, their excellent winemaking properties can command prices that more than compensate for reduced yield and increased production costs.
Many of the varieties listed below are French cultivars. This presumably is a geographic or historic accident, with many other worthy cultivars being little recognized outside their homelands. The cool climate of the best vineyards in France favored the retention of subtle fragrances, complete fermentation, and long aging potential. The former position of France as a major political and cultural power, and its proximity to rich connoisseur-conscious countries, encouraged the selection and development of the best local cultivars and wines. The long and frequent contacts between England and France, and the global expansion of British colonial power, fostered a preference for, and the dispersal of, French and a few German wines and cultivars throughout much of the English-speaking world. There is no doubt that these cultivars produce excellent wines. Regrettably, for the sensory diversity of wines, fine cultivars from southern and eastern Europe did not receive the same exposure and geographic dispersal.
Examples of several important regional, but less well-known, varieties with distinctive aromas are Arinto (white, w) and Ramisco (red, r) from Portugal; Corvina (r), Dolcetto (r), Negro Amaro (r), Fiano (w), Garganega (w), and Torbato (w) from Italy; Rhoditis (w or rosé) from Greece; Furmint (w) from Hungary; and Malvasia (w), Parellada (w), and Graciano (r) from Spain.
Red Cultivars
Barbera is the most widely cultivated variety in Piedmont. It is also important in the rest of Italy, being the third most cultivated variety, after Sangiovese and Trebbiano Toscano. Outside Italy, it is planted in California and Argentina. It is moderately high yielding, producing fruit with an intense color, high in acidity, but moderate tannin content. The clusters possess long green stalks, making mechanical and manual picking easy. Cultivation is also uncomplicated, due to its adaptability to different soils. Barbera can make a varietally distinctive, fruity wine, but is commonly blended with other cultivars to add acidity and fruit to wines of high pH. With regard to disease susceptibility, Barbera is sensitive to grapevine leafroll. Some clones may be severely affected by bunch rot.
Cabernet Sauvignon is likely the best-known red cultivar. This is due both to its association with one of Europe’s best-known red wines (bordeaux), and its production of equally fine wines in many parts of the world. Under optimal conditions, it produces a fragrant wine possessing a black-currant aroma (described as violet in France). Under less favorable conditions, it generates a bell-pepper aroma. The berries are small, acidic, seedy, and possess a darkly pigmented, tough skin. The cultivar is frequently cane-pruned to accentuate production and provide better sun exposure for its upright shoots. The easy separation of the smallish round berries from the cluster facilitates mechanical harvesting. The cultivar is highly susceptible to several fungal diseases, notably Eutypa and Esca wood decays, powdery mildew, and phomopsis. DNA fingerprinting techniques indicate that Cabernet Sauvignon is likely the progeny of a Cabernet franc and Sauvignon blanc cross (Bowers and Meredith, 1997). In Bordeaux, and increasingly in other regions, wines made from Cabernet Sauvignon are blended with wines produced from other, related cultivars, notably Cabernet franc and Merlot. The latter moderates the tannin content and accelerate maturation. Ruby Cabernet is a Davis cross between Carignan and Cabernet Sauvignon. It possesses a Cabernet aroma, but grows better in hot climates than Cabernet Sauvignon.
Merlot has the advantage of accepting cooler, more moist soils than Cabernet Sauvignon, but is more susceptible to coulure (a physiological disruption causing excessive and early flower and/or fruit dehiscence). It is related to Cabernet Sauvignon, also having Cabernet franc as one of its parents. The other parent is a (now obscure) variety, Magdeleine Noire des Charentes (Boursiquot et al., 2009). The tendency of Merlot to mature more quickly has made it a popular substitute for Cabernet Sauvignon.
Dolcetto is an Italian cultivar grown almost exclusively in Piedmont. Nevertheless, it still ranks eighth in overall hectarage in Italy. It has rarely been tried outside its homeland. In growth, Dolcetto possesses comparatively weak vigor, producing small to medium sized clusters, with small rounded berries. It produces a wine usually light, but bright in color, medium-bodied, and with a mild distinctive aroma.
Gamay noir à jus blanc is the primary (white-juiced) Gamay cultivar. Consumer awareness of the variety has increased in parallel with the increased popularity of Beaujolais wines. Half the hectarage of this cultivar is in Beaujolais. It is little cultivated outside of France. Crushed and fermented by standard procedures, Gamay produces a light red wine with few distinctive attributes. This may partially result from its generally high productivity. When processed by carbonic maceration, it yields a distinctly fruity wine. Most of these features come from the grape fermentative process that precedes alcoholic fermentation (see Chapter 9). Gamay produces medium-sized fruit, with a toughish skin. It is sensitive to most fungal grapevine diseases. The Gamay Beaujolais and Napa Gamay grown in California are not directly related to Gamay noir. They are considered to be clones of Pinot noir and Valdiguié, respectively.
Garnacha (Grenache, Cannonau) is a widely planted cultivar in Spain, southern France, southern Italy, Sardinia, Sicily, California and Australia. It exists in several phenotypic color variants (white and gray), in addition to the standard red clones. The vine has an upright growth habit, suitable for head training and spur pruning. It is well adapted to hot, dry conditions, but tends to be excessively productive with irrigation. Clusters are broad and compact, varying from pink to red, depending on the crop load. The cultivar is not well adapted to mechanical harvesting. By itself, it is often used to make rosé or fortified wines. It may be blended with other varieties to speed maturation. Although sensitive to powdery mildew, bunch rot and coulure, Garnacha is relatively resistant to downy mildew.
Graciano is principally cultivated in Rioja and Navarra, but hardly grown outside of northern Spain, possibly because of its relatively low yield. Nevertheless, it is comparatively resistant to most fungal diseases and is drought resistant. The fruit stalks are often woody, making the cultivar ill-suited to mechanical harvesting. The fruit is one of the most aromatic of Spanish varieties and possesses good acidity. It is an integral component of many of the best Rioja wines.
Nebbiolo, grown principally in northwestern Italy, is generally acknowledged as producing one of the most highly regarded red wines. With traditional vinification, it produces a wine high in tannin and acid content, requiring many years to mellow. Nonetheless, the color has a tendency to oxidize rapidly. Common varietal descriptors include tar, violets, and truffles. Nebbiolo yields well only when cane-pruned (due to low basal fertility), but adapts well to a wide range of soil pHs and types. Surprisingly, based on its reputation, it has not been cultivated extensively outside northern Italy. This may be due to the variety’s weak skin and juicy fruit that make it ill-suited to mechanical harvesting. It is particularly susceptible to powdery mildew, but is relatively resistant to bunch rot.
Pinot noir is the famous red grape of Burgundy. It is often considered to have arisen in southern Gaul about the time of the Romans. It possesses several distinctively feral characteristics. It appears to be one of the most environmentally sensitive varieties, and consists of a large number of distinctive clones. Fruit-color mutants have given rise to Pinot gris and Pinot blanc. These changes arose from color mutants in one or both L layers of the fruit. Another common Pinot variant, grown in Champagne, is Pinot Meunier. More prostrate, lower-yielding clones produce more flavorful wines, whereas upright, higher-yielding clones are more suited for the production of rosé and sparkling wines. Pinot noir produces an aromatically distinctive wine under optimal conditions; otherwise, it produces disappointingly nondistinctive wines. Various authors have proposed terms for its aroma, such as beets, peppermint, or cherries, but none seems sufficiently appropriate or adequate. The cultivar produces modest-size clusters of small- to medium-size fruit with large seeds. If the clusters are compact, it is particularly sensitive to bunch rot. Crossed with Cinsaut, it has produced one of the most distinctive of South African cultivars, Pinotage. The Californian cultivar Gamay Beaujolais is a clone of Pinot noir (Bowers et al., 1993).
Sangiovese is probably an ancient cultivar, composed of an extensive number of distinctive clones. It is grown extensively throughout central Italy. It is most well known for the light- to full-bodied wines from Chianti, and produces many of the finest red wines in Italy. Sangiovese is also grown under local synonyms, such as Brunello and Prugnolo. Sangiovese is relatively vigorous, but variable in yield. Its clusters are small- to medium-sized, possessing oval berries. The fruit is compatible with mechanical harvesting. Under optimal conditions, it yields a wine possessing an aroma reminiscent of cherries, violets, and licorice. Sangiovese has achieved little of the international recognition it deserves, the regrettable fate shared by most of its Italian brethren. Sangiovese is particularly sensitive to both bunch rot and powdery mildew. It appears to be the progeny of two ancient, southern Italian cultivars, Ciliegiolo and Calabrese di Montenuova (Vouillamoz et al., 2007).
Syrah is the most renowned French cultivar from the Rhône Valley. Lower-yielding strains produce a deep red, tannic wine, with long aging potential. Until recently, Syrah was hardly grown outside of France, except in Australia, where it is grown extensively under the synonym, Shiraz. Syrah is a vigorous grower with a spreading growth habit. The fruit clusters are elongated with small round to oval berries. It yields deep-colored, flavorful wines with aspects reminiscent of violets, raspberries, and currants, and possesses a peppery finish. Syrah is particularly prone to drought, bunch rot, and infestation by grape berry moths. Most vines labeled Petit Sirah in California are identical to the Rhône cultivar Durif (Meredith et al., 1999). The misidentification is not surprising as Petit Sirah has been used to refer to at least four different cultivars in France itself (Galet, 1990).
Tempranillo (Ull de Llebre) is possibly the finest, at least the most renowned, Spanish red grape variety. Under favorable conditions, it yields a fine, subtle wine that ages well. In addition, it can produce delicate nouveau-style wines. It is the most important red cultivar in Rioja (occupying about 33,000 ha), and is grown extensively throughout much of Spain. Outside Spain, it is primarily grown in Argentina. In California, it usually goes under the synonym, Valdepeñas. Tempranillo generates an aroma distinguished by a complex, berry-jam fragrance, with nuances of citrus and incense. Tempranillo produces mid-size, thick-skinned fruit that is subject to both powdery and downy mildews. The vine is comparatively vigorous and produces upright shoots. It is the progeny of Albillo Mayor, still well known in central Spain, and a (now obscure) cultivar Benedicto (Ibáñez et al., 2012).
Touriga National is one of the preeminent Portuguese grape varieties. It is grown predominantly in the Upper Douro, for the production of port, but is also cultivated in other regions to produce red table wines. The wine is deep in color and richly flavored. The vine is fairly vigorous, with a trailing growth habit, but low in yield. Its clusters are of small- to medium-size, containing small berries.
Zinfandel is extensively grown in California. It is synonymous with the Italian variety Primitivo (Bowers et al., 1993) and its Croatian equivalent, Crljenak kastelanski (Fanizza et al., 2005). Zinfandel is used to produce a wide range of wines, from ports to light blush wines. In rosé versions, it shows a raspberry fragrance, whereas full-bodied red wines possess rich berry flavors. Some of the difficulties with Zinfandel are the uneven manner with which the fruit ripens, and its tendency to produce a second crop later in the season. Both properties complicate harvesting fruit of uniform maturity.
White Cultivars
Chardonnay is undoubtedly the most widely grown white French cultivar. This stems from both its appealing fruit fragrance and its ability to do well in most wine-producing regions. In addition to producing fine table wines, it also yields one of the finest sparkling wines (champagne). The fruit is comparatively small, round, and forms compact clusters. Under optimal conditions, the wine develops aspects reminiscent of various fruits, including apple, peach, and melon. The vine and fruit are predisposed to powdery mildew and bunch rot.
Chenin blanc comes from the central Loire Valley of France. There it yields fine dry and sweet table wines, as well as sparkling wines. Chenin blanc is also grown extensively in Australia, California, and South Africa (in the latter, often under the synonym, Steen). Fine examples of its wine often exhibit a delicate fragrance loosely said to resemble guava fruit or camellia blossoms. The fruit is tough-skinned and of medium size. It is especially susceptible to both downy and powdery mildews, bunch rot, and grape berry moths.
Ehrenfelser is one of the best, newer German varieties, being derived from a Riesling×Silvaner cross. It has many of the characteristics of its Riesling parent, such as its flavor, acidity, disease resistance, and cold-hardiness. In addition, it has the advantage of ripening somewhat earlier. Ehrenfelser is still largely cultivated in Germany, but is being grown with considerable success in Canada.
Müller-Thurgau is possibly the most well-known modern V. vinifera cultivar, constituting nearly 30% of German hectarage. It was developed by H. Müller-Thurgau, supposedly from a crossing between Riesling and Silvaner in 1882. Nevertheless, microsatellite analysis has indicated that Müller-Thurgau is more likely a crossing between Riesling and Chasselas de Courtillier (Sefc et al., 1997) or Madeleine Royale (Dettweiler et al., 2000). The first commercial plantings of Müller-Thurgau began in 1903. It is now extensively grown in most cool regions of Europe and formerly in New Zealand. Its mild acidity and subtle fruity fragrance are ideal for producing light wines. Müller-Thurgau is a high-yielding cultivar that often produces lateral (side) clusters of mid-size fruit. It grows best in rich porous soils. The fruit is subject to both powdery and downy mildews and bunch rot.
Muscat blanc is one of many related Muscat varieties grown extensively throughout the world. Their aroma is so marked and distinctive that it is the only cultivar whose aroma is described in terms of itself, muscaty. Because of the intense flavor, slight bitterness (due to the high level of flavonoids extraction during maceration), and tendency to oxidize, Muscat grapes have most commonly been used in the production of sweet wines. Muscat grapes are also characterized by the presence of high levels of soluble proteins. Consequently, special precautions must be taken to avoid haze formation. The new Muscat cultivar Symphony is less bitter, and its lower susceptibility to oxidation gives it better aging ability. Muscat blanc (Moscato bianco in Italy) is the primary variety used in the flourishing sparkling wine industry in Asti. Other named Muscat varieties include the Orange Muscat (Muscato Fiori d’Arinico in Italy), Muscat of Alexandria, Muscat Ottonel, and the darkly pigmented Black Muscat (Muscat Hamburg).
Parellada is a wonderful variety distinctive to the Catalonian region of Spain. It produces an aroma that is apple- to citrus-like in character, occasionally showing hints of licorice or cinnamon.
Pinot gris and Pinot blanc (respectively Ruländer and Weissburgunder in Germany, and Pinot grigio and Pinot bianco in Italy) are color mutants of Pinot noir. Despite being Pinot noir clones, their vinification and wines are so different that they have been given their own varietal names. Both are cultivated throughout cool climatic regions in Europe for the production of dry, botrytized, and sparkling wines. Neither has gained much popularity outside Europe. Pinot gris can vary in color from bluish to white, depending on the microclimatic conditions of the cluster. It typically yields subtly fragrant wines with aspects of passion fruit. Pinot blanc yields more fruity wines, supposedly with aspects resembling hard cheese.
Riesling (White Riesling or Johannisberg Riesling) is without doubt Germany’s most highly esteemed grape variety. Outside Germany, its largest plantings are in California and Australia. It can produce fresh, aromatic, well-aged wines, which can vary from dry to sweet. Its floral aroma, commonly reminiscent of roses, has made it popular throughout central Europe and much of the world. This renown is reflected in the number of cultivars whose names have incorporated the word Riesling (i.e., Hunter Riesling, Goldriesling, Frankenriesling, and Wälschriesling), none of which bear genetic or aromatic relationship to Riesling. Riesling produces clusters of small- to medium-sized berries, that are particularly sensitive to powdery mildew and bunch rot. The fruit is cold-hardy, and matures relatively late. Yield tends to be moderate.
Sauvignon blanc is one of the primary white varieties in Bordeaux, and the main white cultivar in the upper Loire Valley. It has become popular in California and New Zealand in recent years. It also is grown in northern Italy and eastern Europe. Often, its aroma shows elements of green peppers, as well as a herbaceous aspect, especially in cooler climates. Better clones possess a subtly floral character. Its modest clusters produce small berries that are sensitive to powdery mildew and black rot, but possess partial resistance to bunch rot and downy mildew.
Sémillon is another white Bordeaux variety, and is most well known for its use in producing Sauternes wines. In Bordeaux, it is commonly blended with Sauvignon blanc. Significant plantings also occur in Australia and Argentina. Sémillon produces small clusters of medium-sized fruit, susceptible to bunch rot. The vine is also especially sensitive to fanleaf degeneration. When fully mature, and without the intervention of noble rot, Sémillon yields a dry wine, said to contain nuances of fig or melon. These aspects develop primarily during aging.
Traminer is an ancient cultivar, some clones of which possess a distinctive aromatic character. The cultivar is grown primarily in cooler climatic regions. Its clones can be grouped into two color classes: white, and pink to reddish. Pink strains appear to possess incomplete expression of the Gret1 retrotransposon (that inhibits anthocyanin synthesis), permitting partial berry coloration (Pelsy et al., 2010). Regardless of the grape’s color, all clones are processed as white grapes, producing dry to sweet wines, depending on regional preferences. The name Traminer is usually applied to white clones with a mild fragrance. White, aromatically neutral clones are often called Savagnin, notably in southeastern France. Intensely fragrant, pinkish, Gewürztraminer clones may produce wines possessing an intense aroma resembling that of lichi fruit. Nonetheless, depth of coloration and aromatic character may depend more on cultural and climatic conditions than on the clone’s color (Bourke, 2009; Duchêne et al., 2009). All forms produce modest clusters of small fruit with tough skins. The variety is prone to powdery mildew and bunch rot, and often expresses coulure. Albariño (Spain) and Alvarinho (Portugal) were once thought to be additional synonyms, but have been shown to be unrelated (Santiago et al., 2007).
Viognier has been a variety largely restricted to the Condrieu appellation in the Rhône Valley. Comparatively recently, it has caught the fancy of several North American and Australian producers. The variety has a tendency toward poor fertility and, correspondingly, low yield. Thus, it needs to be cane-pruned. The vine generally requires excellent drainage. Viognier produces small round berries possessing a muscaty fragrance. Correspondingly, the wine matures quickly. The variety is also reported to show peach or apricot resemblances.
Viura is the main white variety in Rioja. It produces few clusters, but they are of considerable size. In cool regions, it produces a fresh wine possessing a subtle floral aroma with aspects of lemon. After prolonged aging in wood, it develops a golden color and rich butterscotch or banana fragrance. These characterize the traditionally aged white wines of Rioja, now regrettably rare.
Interspecies Hybrids
American Cultivars and Their Hybrids
Although decreasing in significance throughout North America, early selections from native American grapevines or accidental interspecies hybrids are still extensively grown. American hybrids constitute major plantings in eastern North America, particularly Washington state, and they are grown commercially in South America and Asia.
Of American cultivars, the most important are based on V. labrusca. They possess a wide range of flavors. Some, such as Niagara, are characterized by the presence of a foxy aspect. Others are characterized more by a strawberry fragrance (Ives), the grapy aspect of methyl anthranilate (Concord), or a strong floral aroma (Catawba). Wines produced from V. riparia and V. cinerea do not possess detectable concentrations of methyl anthranilate, or 2-aminoacetophenone. The latter may be the principal source of the foxy aspect attributed to some V. labrusca grapes. In contrast, they are characterized by high concentrations of methoxypyrazines, the same compounds that characterize Cabernet Sauvignon and related cultivars. They are also characterized by herbaceous flavors, associated with eugenol, cis-3-henenol, and 1,8-cineole (Sun et al., 2011). The high acidity and low sugar content of American cultivars, and their hybrids, have made chaptalization (the addition of sugar to the juice) necessary for standard table-wine production.
Various methods have been used to diminish what is often viewed as the overabundant flavor of most American cultivars. Long aging results in the dissipation of labrusca flavors. This is generally non-feasible, as most V. labrusca wines are consumed young. The presence of high levels of carbon dioxide, as in sparkling wines, tends to mask most labrusca fragrances. Processing the grapes via carbonic maceration is another means of reducing (or masking) the intensity of labrusca flavors. However, the most generally accepted mechanism is early picking and cold fermentation. These limit the development and extraction, respectively, of labrusca flavors, while still producing a wine with fruitiness. Conventional wisdom suggests these reductions in labrusca flavors are necessary. Personally, this is another example of the dubious benefit of the lemming effect, so typical in assessing wine quality.
The varieties Norton and Cynthiana (possibly clonal variants; Reisch et al., 1993) are predominantly of Vitis aestivalis origin. These cultivars are primarily cultivated in the lower Midwest (Arkansas, Kentucky, Illinois, and Missouri) and mid-Atlantic states (Maryland and Virginia). Locally derived from wild vines, and possibly introgressed to V. vinifera, they possess resistance to the local indigenous diseases and pests that complicate cultivation of vinifera varieties in these states.
The other major group of American cultivars are those derived from V. rotundifolia. Although Scuppernong is the most well-recognized varietal name, it is rarely grown today. New self-fertile varieties, possessing different aromatic properties, are the principal muscadine cultivars grown in commercial vineyards (Morris and Brady, 2007). The excellent resistance of these cultivars to indigenous diseases, especially Pierce’s disease, has allowed them to flourish in the southeastern coastal United States. Similar to the V. labrusca cultivars, the low sugar content of the fruit usually requires chaptalization before vinification. The pulpy texture, tough skin, differential fruit maturation, and separation of fruit from the pedicel on maturation complicate their use in winemaking.
Most muscadine cultivars have a distinctive and marked fragrance, containing aspects of orange blossoms and roses. Some fertile crossings with V. vinifera show vinifera-like flavors, combined with the fruiting characteristics and disease resistance of their muscadine parentage.
French-American Hybrids (Direct Producers)
French-American hybrids were developed to avoid the necessity, complexity, and expense of grafting V. vinifera cultivars to phylloxera-resistant rootstocks. The easier cultivation, reduced sensitivity to several leaf pathogens, and higher yield made them popular with many grape growers in France. The tendency of base buds to grow and bear fruit lead to frequent overcropping. This exacerbated the increasingly serious problem of grape overproduction in France, a problem that still plagues much of Europe. This factor, combined with the nontraditional fragrance of these hybrids, led to a general ban on new plantings. Their use in appellation control (AC) wines was also prohibited. Restrictions against French-American hybrid use subsequently spread throughout the European Union (EU). The remaining, temporary, exception is the cultivation of Baco blanc for armagnac production, which is supposed to have expired.
In North America, with the exception of most of the southern, gulf, and western coastal states, French-American hybrids formed the basis of the expanding wine industry in the early 1960s. They are still grown extensively in some South American and Asian countries. In Europe, as well as other areas, French-American hybrids are often used in breeding programs as a source of resistance to several foliar, stem, and fruit pathogens. Many French-American hybrids suffer from sensitivity to soil-borne virus diseases, such as those of the tomato ringspot group (Alleweldt, 1993). Where this is a problem, grafting to resistant rootstocks is required.
Unlike American hybrids, few French-American hybrids possess Vitis labrusca parentage. Vitis rupestris, V. riparia, and V. aestivalis var. lincecumii were the principal species used. Although some French-American hybrids are simple crosses between an American Vitis species and a V. vinifera parent, most are derived from complex crosses between an American species and several V. vinifera cultivars. This is clearly evident from a study of their genetic diversity (Pollefeys and Bousquet, 2003).
A brief description of some of the better known American and French-American hybrids is provided below.
Baco noir is a Folle blanche×V. riparia hybrid. Its acidity, flavor, and pigmentation yield a wine with considerable aging potential. It develops a fruity aroma associated with aspects of herbs. It is sensitive to bunch rot and several soil-borne viruses. Poor cane maturity is often a problem in cold climatic regions due to vine vigor.
de Chaunac is a Seibel crossing of unknown parentage. Once widely planted in eastern North America, due to its cold-hardiness and high yield, its tendency to produce wine of neutral character and its susceptibility to several soil-borne viruses have resulted in a loss of favor.
Chambourcin is a Joannes Seyve hybrid of unknown parentage. Its popularity increased markedly in the Loire Valley during the 1960s and 1970s. It is considered one of the best of French-American hybrids. Chambourcin has also done remarkably well in Australia, possibly due to the long growing season. This permits full grape maturity and the development of a rich, wonderfully complex flavor. Chambourcin has also found a following in eastern North America. The variety possesses good resistance to both downy and powdery mildews.
Maréchal Foch is a Kuhlmann hybrid derived from crossing a V. riparia×V. rupestris selection with Goldriesling (Riesling×Courtiller musqué). It yields deeply colored, berry-scented, early-maturing wines. The variety’s characteristics of winter-hardiness, productiveness, early maturity, and resistance to downy mildew have also given it appeal in eastern North America.
Delaware is one of the finest, early ripening, light-red American hybrids. It is generally thought to be a V. labrusca×V. aestivalis×V. vinifera hybrid. Nevertheless, its susceptibility to phylloxera and various fungal pathogens, tendency to crack, and the need for well-drained soil has limited its widespread cultivation. It was once extensively used in sparkling-wine production.
Dutchess is another highly rated, older, late-ripening, white American hybrid. This Vitis labrusca×V. vinifera hybrid has a mild, fruity aroma with little labrusca character. As with Delaware, difficulty in growing the cultivar negates most of its enologic qualities.
Magnolia is one of the more popular, new, muscadine cultivars. It produces bisexual flowers and is self-fertile. It yields sweet, bronze-colored fruit.
Noble is a dark-red muscadine cultivar. Its deep-red color and bisexual habit have made it popular in the southeastern United States.
Seyval blanc is a Seyve-Villard hybrid of complex V. vinifera, V. rupestris, and V. aestivalis var. lincecumii parentage. The variety yields a mildly fruity white wine, with a pomade fragrance and bitterish finish. Although susceptible to bunch rot, it is relatively winter-hardy, tolerant of many soil types, and a consistent producer.
Vidal blanc is possibly the best of the white French-American hybrids. This Vidal hybrid of complex ancestry has both excellent winemaking and viticultural properties. Under optimal conditions Vidal blanc yields a wine of Riesling-like character. Its tough skin and late maturity assists in its being used in the production of icewines of excellent quality. The variety is relatively cold-hardy, but less so than Seyval blanc.
Although most interspecies hybridization ceased in France by the 1920s, it has continued unabated in Germany. Varieties such as Orion, Phoenix, and Regent show winemaking qualities equal or superior to several currently used V. vinifera cultivars. Breeding has also continued in eastern North America. Veeblanc is one of the newer white cultivars of complex parentage developed in Ontario. It generates a mildly fruity wine of good quality. New York also has an active breeding program. One of their most commercially successful introductions to date has been Cayuga White. It is well adapted to a range of wine styles; possesses a fruity fragrance resembling apples, citrus, and tropical fruit; and produces a wine without bitterness and a rich mouth-feel. When harvested early, it produces an excellent base for sparkling wines. Although very productive, Cayuga White seldom requires cluster thinning. It is also resistant to most common fungal diseases and, thus, needs little fungicidal protection. Another popular new introduction from Cornell is Traminette. It possess much of the aromatic character of its Gewürztraminer parent, but is more cold-hardy. In cold climatic regions, such as the midwestern and northeastern United States and Quebec, another interspecific hybrid, Frontenac, has proven both successful and popular. It is a University of Minnesota cross between Vitis riparia and the French-American hybrid, Landot 4511.