CHAPTER TEN
Evolution, Sex, and Gender
IN THE SPRING OF 2005, I WAS RECRUITED TO BE THE DEAN OF general education at NCATSU, now the nation’s largest public HBCU. I began my work there in July of that year. The story of how I came to this assignment is interesting in and of itself. In 2004 I was the director of the general education core curriculum at Fairleigh Dickinson University in New Jersey. I received an invitation to give a lecture at NCATSU. There was nothing special about receiving these invitations. By this point in my career, I was routinely lecturing all over the country as well as internationally. Amazingly, this was actually my first invitation to lecture at an HBCU campus.
When I arrived in Greensboro, North Carolina, and was given my itinerary for the day, it didn’t seem right for a lecture visit. My first meeting, at 8:00 a.m., was with the chancellor, James T. Renick. It was to be followed by meetings with the dean’s council and vice-chancellors and finally, in the late afternoon, by my campus-wide lecture. Chancellor Renick let the cat out of the bag during our meeting. In short, he wanted me to apply to be the dean of their new general education (GenEd) program. Their vision was to focus the GenEd curriculum around competencies rather than specific disciplines. The competencies were generated from high-level national discussions with leaders of industry and other constituencies concerned with reforming education to meet twenty-first-century priorities. In a break between the morning meetings, I called my wife to get a sense of her feelings about my taking on a deanship. She was not opposed, so by the end of the day I was convinced I should apply for the position.
When I arrived at NCATSU in the summer of 2005, I had no staff or space. I was tasked with building an entire program devoted solely to effectively teaching and assessing general education competencies. The program, called University Studies (UNST), offered interdisciplinary courses organized around building critical-thinking competencies in science, writing, history, social sciences, and African American studies. The coursework covered freshman and sophomore years. Within five years I brought together a program with fifty faculty members, with at least ten of them in tenure-track positions. Midway through the program, we were so successful in our efforts that I was asked by the American Association of Colleges and Universities (AAC&U) to be part of their general education training faculty. The AAC&U workshops engaged educators from top American universities (including top-tier research institutions) who were making reforms in their GenEd curricula.
In my five years as an academic dean, my primary responsibility was the implementation and assessment of the curriculum. Life in the HBCU environment was dramatically different from what I had experienced in HWIs. On the positive side, I did not have to deal with the death-by-a-million-cuts microaggression of the HWIs (or their overt macroaggression). I was also doing work to benefit a much larger percentage of African American students than I ever experienced at HWIs. NCATSU’s undergraduate enrollment is still well over 95 percent persons of African descent. On the negative side, HBCUs still suffer from the legacy of white supremacy and segregation. African American education in the former Confederate states is the victim of Jim Crow, Plessy v. Ferguson, the second Morrill Land-Grant Act (1890), and uneven and toxic implementation of Brown v. Board of Education.
The letter of Plessy v. Ferguson was that if facilities were to be separate, they had to be equal. During Jim Crow equal facilities never existed. Over this time period, southern states found ways to divert hundreds of millions of dollars away from Black schools to serve white schools instead.1 In 2018 a national study showed that southern states spend $5 billion less educating Black and Brown students than white students, at a rate of approximately $1,000 per year per student.2 The historical neglect of African American education and racism across the nation means that at NCATSU we admit students with a broader diversity of academic preparation than do our top-tier HWI neighbors. Many of our students are gifted and could have gone anywhere, but we also admit a large fraction of students who have holes in their preparation. My academic division was charged with repairing those holes via general education. We did so, with much less money to work with per student than the HWI state schools had. Thus, it was always more of a challenge for us than for the privileged HWIs to have enough faculty and infrastructure to conduct our work.
Despite the fact that the majority of my time was devoted to running the program, I wanted to stay intellectually active within my field. However, without a research laboratory, it simply wasn’t possible for me to contribute to new basic research. In this period I contributed primarily through my work on a variety of committees designed to provide guidance to important scholarly projects around the nation, including the advisory committee for Teaching Ethical, Social, and Legal Issues of the Human Genome Project (Dartmouth College, 2003–2005); the New Genetics and the Trans-Atlantic Slave Trade Working Group (W. E. B. DuBois Institute, Harvard University, cochaired by Henry Louis Gates Jr. and Evelynn Hammonds, fall 2006–2008); and, finally, my work on the senior advisory board of the National Evolutionary Synthesis Center (NESCent) at Duke University, which I eventually chaired from 2009 to 2012.
NESCent was funded by the NSF and was a collaboration between Duke University; the University of North Carolina, Chapel Hill; and North Carolina State University. Space for the NESCent center was provided by Duke University. Its goal was to bring together scholars to integrate concepts, methods, and data within the subdisciplines of evolutionary science for the purposes of synthesis. NESCent was specifically not allowed to fund projects that collected new data. The goal of the synthesis was to see new things in the data, things that might have escaped notice, and to thus stimulate new research. Given the proximity of Duke to NCATSU, it was an easy drive to attend programs at the center. During the ten years of the center’s operation, pretty much anyone who was anyone came through. My time at NESCent allowed me the opportunity to think about problems in evolution on which I had not done any prior work. One of them was a fundamental question that had begun to take on greater significance in my own life.

EVERYONE KNOWS OR HAS KNOWN SOMEONE WHO DOES NOT FIT the gender binary. I remember a student in my elementary school class who definitely did not behave like a “typical” boy. He was bullied for this. I did not participate in the bullying, but neither did I take (or know how to take) a stand as his ally. At that time I had a relative who grew up to be an openly gay male. In high school, when I was working at the lumber company in the summers, I worked with an older man who was always interested in hearing about my sexuality. At the time, there was nothing for me to tell, so my side of the conversations was short. However, though this man was married and had children, he would always joke about having sex with other men. In hindsight, I believe he was either bisexual or a closeted gay man. As a college student I came into contact with gay men and lesbian women more often, partly because it was easier for students to express their sexuality away from their parents and hometown. Many of the people I worked with when I was a revolutionary were gay, because gay people are among the most oppressed people on this planet. As this was in Detroit, most of them were African Americans, so they were dealing with racism, classism, and antigay bigotry. It was in these days that I learned to be a more effective ally for the LGBT struggle.
The LGBT struggle took on even more significance to me when I began to talk to parents I knew who had gay or lesbian children. These stories reminded me of how the person I knew in kindergarten was treated. Some talked about children who were more interested in dressing up in girls’ or boys’ clothes as opposed to the expected cisgender clothes. Others talked of children who preferred to play with more feminine or masculine toys than those of their gender stereotype. (It is important to understand that the toy gender binary results from the existence of gender stereotypes. Boys aren’t supposed to be interested in dolls, and girls aren’t supposed to be interested in commando action figures.)
He was bullied through elementary school, middle school, and high school. He was a talented swimmer, but he quit the high school swim team because of bullying (which his coaches did nothing to prevent). To this day, he is still bitter about the sport (he will not go to the pool or watch competitive swimming events on television). I was not surprised when he came out to his parents as gay in high school. When we heard from him that he was taking ownership of his sexual orientation, our first reaction was fear for his safety. He lives in a southern state with a strong history of biblical literalism and antigay bigotry. However, his family chose not to be paralyzed by fear but to act to guarantee his safety and rights in this society. Knowing of his struggles, when I began writing blogs for the Evolutionary Studies Consortium, one of my first pieces was on the myth of the gender binary.3 I wanted him to read the piece so he would be better equipped to understand himself and would not feel there was something wrong with his sexual orientation. As an evolutionary biologist I understood that biological sex is one of the most important components of evolution. By necessity it plays a key role in all sexual species, including humans. Gender, on the other hand, is an emergent property, not something one can understand solely by knowing an individual’s biological sex.
THE GENDER BINARY IS DEEPLY ENCODED IN WESTERN SOCIETY through the three Abrahamic religions. In Genesis, the J writer states in Chapter 1 that man and woman were created at the same time and the E writer states in Chapter 2 that man was created first and woman was created from Adam’s rib. Genesis 1:27 states, “Male and female he created them,” and the creation of two biological sexes is related to the command in Genesis 1:28 to reproduce: “Be fruitful and increase in number.” There is no discussion of hermaphrodites (both sexes in the same individual, human or otherwise) in Genesis—notable, in that the chapter discusses the creation of flowering plants, which contain an abundance of hermaphroditic species.
The Bible’s superficial treatment of gender and sexual orientation is further evidence that it was not designed to be a textbook of natural history. The discussion of homosexuality in the Bible in the main is as blasphemy or as abomination. The Hebrew Bible (Old Testament) has several passages that have been interpreted as condemnation of homosexuality (e.g., Genesis 9:20–27, on Ham, a son of Noah; Genesis 19:1–11, on Lot and his daughters; Leviticus 18:22 and 20:13).
However, attempting to use the story of Noah and Ham as a condemnation of homosexuality is a stretch. Saying that Ham looked upon the nakedness of his father (which was forbidden) does not mean that Ham had any sexual designs on his father. Indeed, the historical damage done by interpretations of this passage have more to do with its role in justifying slavery and racism.4 The story of Lot, the angels, his daughters, and the wicked men of Sodom need not be interpreted as a condemnation of homosexuality either. It could just as easily be read as a condemnation of sexual violence and the disrespect toward strangers. Indeed, biblical scholars disagree about the meaning of this text, especially in light of its great similarity to the story in Judges 19, on a Levite and his concubine.5 Some biblical scholars argue that the story in Judges is actually older than the one in Genesis (the books of the Bible, Hebrew or Christian, are not organized in the chronological order in which the events they purport to describe happened.)6 The supposed prohibitions in Leviticus are even harder to understand, let alone to use as evidence that God condemns homosexuality. In addition to the verses already noted, Leviticus 18 lists a number of taboos against having sex with relatives (incest taboos are common across all human cultures).7 Leviticus also frequently condemns the practices of foreign cults, including the sacrifice of infants to ensure the future fertility of the mother, a practice of the Canaanites as well as the Israelites.8 Leviticus 19 prohibits having sex with a woman during her menstrual period. Taking these passages as a whole, and considering issues of translation, it is hard to interpret them as God condemning homosexuality in general; rather, that interpretation seems more like another example of biblical literalists cherry-picking passages to support their vision of what human morality should be.
It is also clear that homosexuality was not uniformly condemned in the ancient world, as it was socially acceptable among both the Greeks and Romans. The apostle Paul’s condemnation of homosexuality in Romans 1:26–27 may have been related to his associating this with incidents in the Old Testament and in the Torah concerning the temptation to worship the gods and goddesses of the Canaanites. A requirement of Canaanite worship was to have sexual intercourse with a priest or goddess. Paul would have seen those acts of worship as an abomination to the God of Israel.9
WHETHER GREEK, ROMAN, OR HEBREW, THE ANCIENTS UNDERSTOOD very little about how sex operated in nature. In the main this understanding would begin to take shape in the twentieth century. The complexity of the phenomenon is well illustrated in an example from fiction.
In the novel Jurassic Park, InGen geneticist Henry Wu tells paleontologist Alan Grant and Ian Malcolm, “In fact, they’re all female. And, believe me, they can’t breed.”10 In all the Jurassic Park novels and movies, Wu is the villain I detest the most. One might have sympathy for John Hammond, despite his hubris. Quite frankly, he doesn’t know enough about biology to understand why his project is doomed to failure. Wu, on the other hand, epitomizes for me the often-dangerous arrogance of scientists; this arrogance is made even worse when combined with greed. Wu also illustrates another principle that must be avoided at all costs: he is misled by molecular reductionism, thinking that one understands biology best by working upward from molecules to organisms (a fallacy he shares with the real-world James Watson). For this reason, Wu is confident in his assertion that the genetically engineered monsters of Jurassic Park are incapable of breeding because they were originally all female. He is wrong, and because of his mistake (in the novels) the organisms escaped the park and found a reproducing population in Central America. Such a thing wouldn’t happen if Wu had understood Dobzhansky’s dictum “Nothing in biology makes sense except in the light of evolution.” Nowhere is this principle better illustrated than in the way it allows us to understand the biological significance of sex.
When evolutionary biologists discuss sex, they are referring to genetic recombination. In eukaryotes that means reproduction that includes meiosis, with crossing over and mating between two unrelated individuals. From the anthropomorphic lens utilized by most people (such as Wu), this description seems entirely reasonable. However, reproduction without sexual recombination is actually the rule among living things (especially if you count bacteria). In addition, reproduction by cloning (parthenogenesis) is common among plants, fungi, and animals.11 A few years ago, an example of parthenogenesis was observed in an aquarium when a zebra shark produced a “virgin birth.”12 During my master’s work at Lowell, I learned that some species of lake-dwelling Daphnia (called water fleas) generally reproduce only by parthenogenesis during the summer months; they produce males during the fall, undergo sexual reproduction, go into diapause (which is similar to hibernation) in the winter months, emerge in spring for another round of sexual recombination, and go back into parthenogenesis in the summer. In this case, the Daphnia are getting the most out of both modes of reproduction.13 In the summertime, when the lake environments are generally stable, the more rapid growth of parthenogenesis makes more sense than wasting reproductive effort on the production of males (which do not reproduce; see Figure 10.1). However, one of the well-supported advantages of sex is that it facilitates a greater ability to produce genotypes capable of adaptation to changing conditions (as expected in the fall and spring). Thus, again, it makes great sense to utilize sex to recombine genetic variants under these conditions.
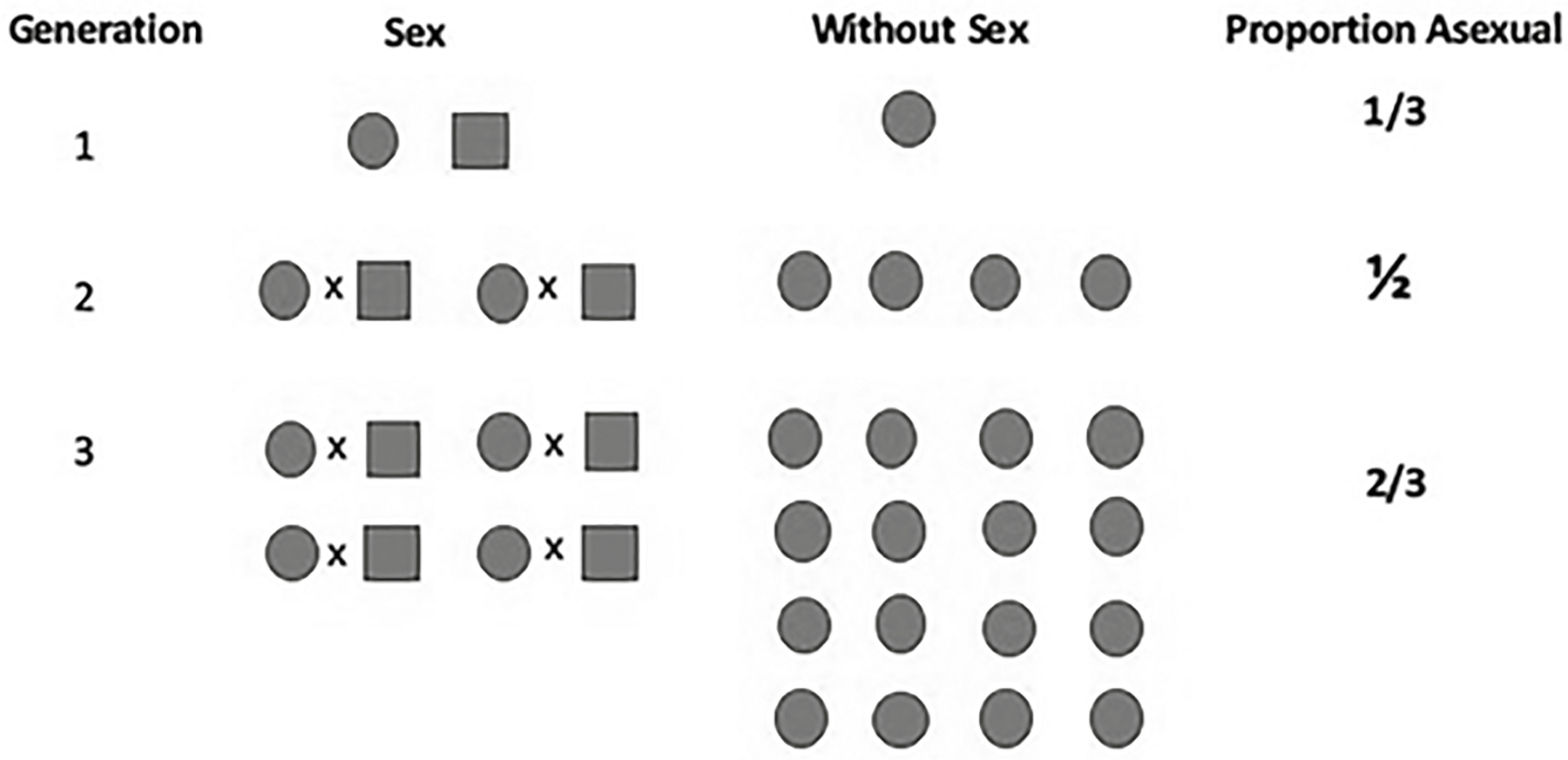
Figure 10.1. Comparative growth of genotypes with and without sex (circles are females; squares are males). In the first generation, the proportion of asexual to sexual individuals is 1 to 2. However, in the second generation it is 1 to 1, and by the third generation it has become 2 to 1. In the fourth generation (not shown), the ratio would be 3 asexual to 1 sexual. At this rate, sexual individuals would soon disappear from the population. This algorithm assumes that each female reproduces at the same rate (four offspring) and that the survival of offspring produced asexually and sexually is the same.
The question of the origin and maintenance of sex has been of great importance to evolutionary theory. The usefulness of recombination to bring together beneficial mutant alleles is illustrated in Figure 10.2. Put simply, sex speeds up the pace of evolution, and this capacity exists in a fundamental trade-off with the costs of producing males, searching for mates, expending energy to successfully mate, or risking additional exposure to pathogens as a result of mating. Indeed, because sex is often so beneficial, various organisms, from viruses through parasitic eukaryotes, specialize in infecting the reproductive tracts of other organisms. You have undoubtedly heard of the bevy of parasites transmitted this way in humans (causing venereal diseases—e.g., HIV, syphilis, gonorrhea, trichomoniasis, and others). A particularly interesting case of an organism once thought to be just the cause of a venereal disease of fruit flies is the bacterium Wolbachia. This bacterium inhabits the reproductive tract of Dipterans (flies, such as Drosophila) and plays a role in maintaining the reproductive barrier between closely related species. Recently it has been shown that Wolbachia alters patterns of gene expression that help enforce the reproductive barrier between six closely related semispecies of D. paulistorum.14 In addition, it has been shown that infection from Wolbachia provides protection against infection from RNA viruses—a fact that is now being deployed to help stop the spread of dangerous RNA viruses, such as Zika, to humans by mosquitoes (I discuss dangerous RNA viruses more in Chapter 11).15
However, the production of males is only beneficial if change is a consistent feature of the environment an organism occupies (as in the cyclical case of the lake Daphnia described above). Several elegant laboratory experiments have supported this thesis. A Caenorhabditis elegans roundworm can be hermaphroditic, or it can be a male that can mate with the female organs of the hermaphroditic worms. Biological sex determination in this species occurs by an X/0 system, where females are XX and males are X (lacking a second X chromosome, that is, being 0). Theoretically, in such a system males can only persist if the frequency of male mating × the differential fitness (measured by survival × reproduction) of their offspring exceeds the number 2.0.16 Experiments with C. elegans have demonstrated variation in male mating success and the fitness of their offspring.17 Once again, aspects of the environment, such as the presence of toxins or parasites or if individuals migrate consistently between environments, and predictability of environmental change play crucial roles in the maintenance of sexual reproduction.
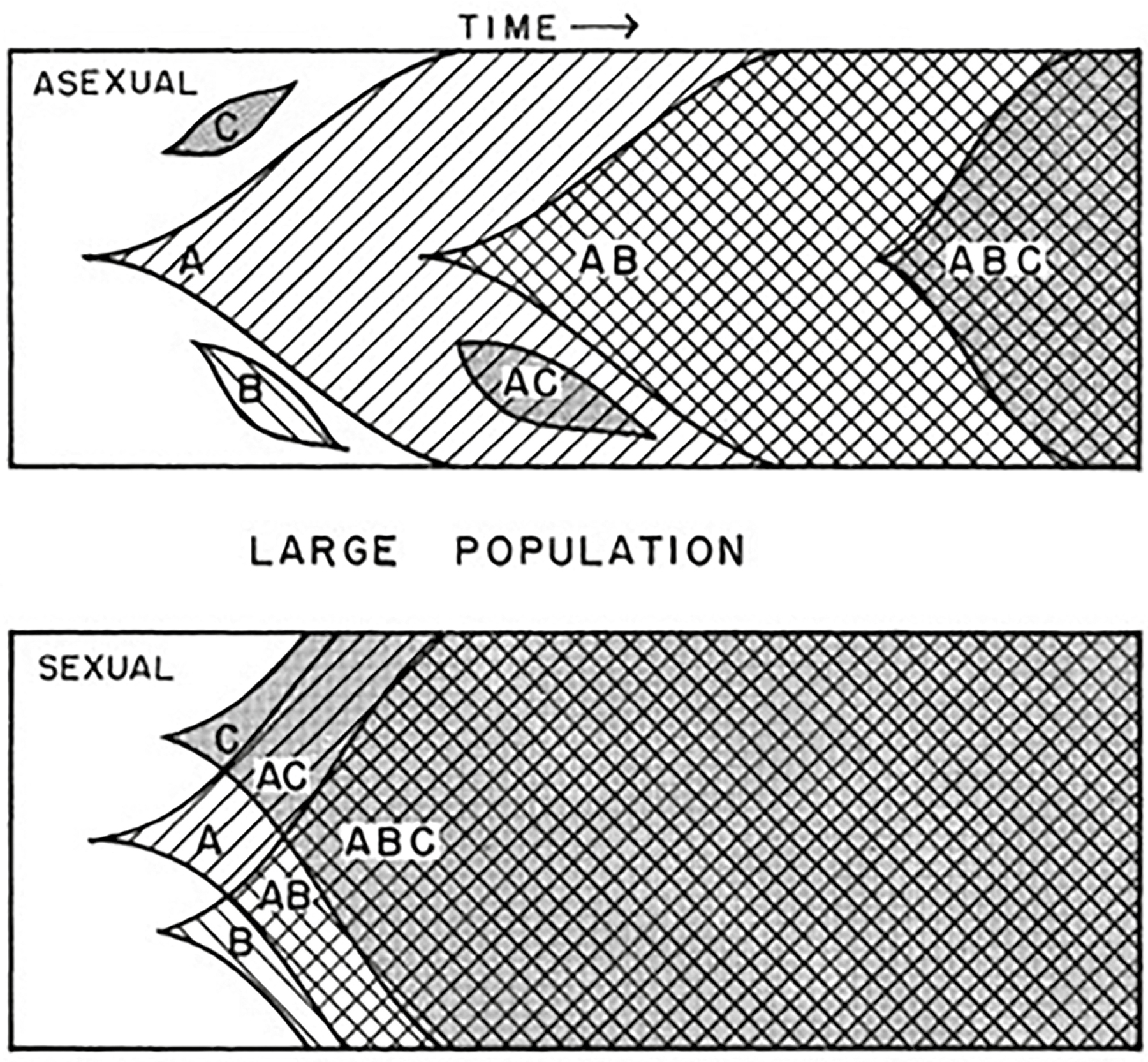
Figure 10.2. Genetic recombination by sex. This figure illustrates the way genetic recombination by sex can bring together useful genetic variants more quickly than is observed in asexual populations. In brief: Suppose that in an asexual population, a random beneficial mutation occurs in three different clones (A, B, C). For the A clone to acquire all three beneficial mutations would require subsequent mutations in the B and C genes. In the recombining case, if the mutation occurs in A, during meiosis during gamete formation, both B and C can end up on the same chromosome as A. Finally, a crossover event between chromosomes AC and AB produces a chromosome with ABC.
Source: Diagram from Crow JF and Kimura M, Evolution in sexual and asexual populations, American Naturalist 99, no. 909 (1965): 439–450; originated by Muller HJ, Some genetic aspects of sex, American Naturalist 66, no. 703 (1932): 118–138.
H. J. Muller’s concept of genetic load expanded on his original theoretical work on the significance of sex.18 I have a great fondness for this theory, as I remember fumbling it badly during my oral exam (see Chapter 3). Muller’s genetic load theory was premised on the idea that the frequency of a mutant allele in a population is the result of the mutation rate, the strength of selection against the allele, and, in finite populations, genetic drift. In asexual populations, individuals can be sorted into groups based on the number of bad mutations they contain in their genome (Figure 10.3). It is important to recognize that all organisms contain mutations in genes that make their function less optimal than the “best” gene within that species. The more bad mutations an individual has in its genome, the lower its evolutionary fitness. Muller proposed that asexual clones or species would continue to increase the mean number of their bad mutations over time because genetic drift would eventually cause the loss of those with the fewest bad genetic variants. This increasing number of bad mutations he dubbed “genetic load.” However, sexual individuals can reconstitute their “good” genotypes by recombination. Since sexual recombination is itself determined by genetic variation, association with the highest-fitness variants means that sexual recombination will sweep through populations, eventually replacing asexual variants.
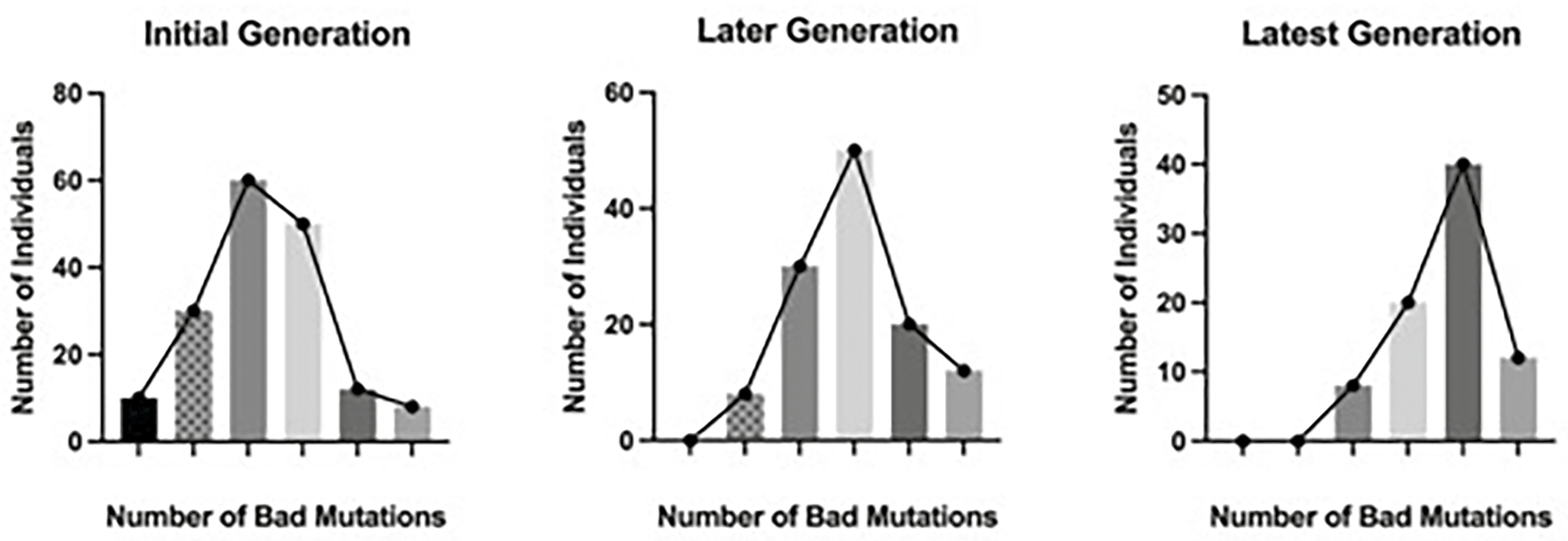
Figure 10.3. Increase in the frequency of bad mutations across time (Muller’s ratchet). This figure shows the frequency of bad mutations in the genome of a hypothetical asexual population across evolutionary time. In the initial population, the individuals from left to right contain 0, 1, 2, 3, 4, and 5 bad mutations in their genome. The relative evolutionary fitness of each class is determined by its number of bad mutations. In a later generation, individuals with 0 bad mutations have been lost through genetic drift. In the latest generation, individuals with only 1 bad mutation were lost through genetic drift. As this process continues, the mean number of bad mutations will continue to increase. This progression is called Muller’s ratchet.
Muller’s idea has been tested in a variety of asexual versus sexual organisms in the same phylogenetic group (plants, fungi, animals), and there is excellent experimental validation supporting it. In a test of six asexually reproducing versus sexually reproducing species of walking stick insects (genus Timema), the asexual species accumulated significantly more deleterious mutations than did the sexual species.19 Other tests of the idea have been conducted and validated in bacteria, plants, and fungi.20
Additional evidence in favor of the benefit of sexual recombination results from the constant pressure organisms are under from their parasites. Virtually all cellular life faces such pressure, but it is an even bigger problem for large-bodied organisms because of the disparity between the length of their generation times and that of parasites such as viruses, bacteria, and unicellular and small-bodied eukaryotes (e.g., sporozoans, trypanosomes, nematodes). The idea that continual evolution is necessary to prevent parasites from dismantling one’s defenses is called the Red Queen hypothesis, after the character in Lewis Carroll’s Through the Looking Glass. (This character said, “It takes all the running you can do, to keep in the same place.”)21 So, in summary, there are good theoretical reasons and experimental support for the origin and maintenance of sex. In the history of evolutionary biology, these reasons have fallen into four broad categories: selection favoring advantageous novel genetic combinations (an idea that both Fisher and Muller developed); the reconstitution of advantageous combinations to avoid accumulation of bad mutations (Muller’s ratchet); the evolutionary arms race between pathogens and host (the Red Queen hypothesis); and finally, the purging of synergistic bad mutations (which is sort of a combination of Fisher/Muller and Muller’s ratchet).
THE DETERMINATION OF BIOLOGICAL SEX IN DIFFERENT EVOLUTIONARY lineages is far from simple. What is clear is that genes determine biological sex (please note that I am discussing not gender or sexual orientation here but, rather, whether an organism is male or female). Genetic sex determination systems vary widely, even within the same phylogenetic lineages. Some organisms have “sex chromosomes,” in that genes that play an important role in determining biological sex are routinely found on them (such as the X, Y system in humans). However, even in this case, genes on autosomal (nonsex) chromosomes are also involved. Some organisms display individuals that are hermaphroditic; others have an obligate separation between male and female individuals. However, even in those groups with the obligate distinction, individuals may change their biological sex under specific conditions. For example, the bluehead wrasse (Thalassoma bifasciatum, a reef fish) undergoes socially determined sex change by a set of somewhat complicated changes in gene expression via epigenetic modification.22 An individual can change from female to male in response to the death of the males; usually the change occurs in the remaining most dominant female. Sex change also occurs widely in amphibians. Reptiles vary between those with genetic sex determination and others with no specific sex determination chromosomes. In the latter, the temperature at which an egg is incubated determines whether it will be male or female.23
The evolutionary argument for plasticity for biological sex determination follows from basic evolutionary theory concerning the stable sex ratio for populations. R. A. Fisher made a relatively simple argument for the stability of the 50:50 ratio in The Genetical Theory of Natural Selection.24 In brief, if an individual had a mutation that produced an all-female sex ratio, her offspring females would probably have lesser fitness because of competition with all the other females; similarly, if the mutation produced only male offspring, those males would have lower fitness because of competition with the other males. Thus, the stable system for determining sex ratio would be one that produced equal numbers of females and males. By this reasoning, some of Henry Wu’s monsters, if capable of altering their sex ratio, would produce males, as those males in an all-female environment would have greater fitness than the female offspring. In Jurassic Park, the reason Alan Grant provides for the dinosaurs’ ability to shift is that Wu had used “amphibian DNA.” However, in reality, we do not know whether dinosaurs lost the trait of environmental determination of sex that we know was within the stem reptiles they were derived from. It has been lost in birds, but not all groups of dinosaurs gave rise to the birds. So if Wu knew a little bit of evolutionary theory, he would not have clung to his molecular reductionist hubris, and maybe more human lives would have been saved.
BUT WHAT’S ALL THIS HERE TALKING ABOUT? THAT MAN OVER THERE SAYS THAT WOMEN NEED TO BE HELPED INTO CARRIAGES, AND LIFTED OVER DITCHES, AND TO HAVE THE BEST PLACE EVERYWHERE. NOBODY EVER HELPED ME INTO CARRIAGES, OR OVER MUD PUDDLES, OR GIVES ME ANY BEST PLACE. AIN’T I A WOMAN?
—SOJOURNER TRUTH, “AIN’T I A WOMAN?,” MAY 29, 1851
It is hard to know how many lives may have been saved if sexism and patriarchy had not triumphed in human societies. It is not my intention (nor do I think it is my place) to attempt a full discussion of the perfidy of the ideological justifications for the subordination of women and gender nonconforming persons. That has been achieved by far abler persons.25 However, what I can do here is debunk biological determinist arguments for the origin, maintenance, and legitimacy of these oppressions.
Discrediting such arguments begins by recognizing the crucial distinction between biological sex, gender, and sexual orientation. In the last section I discussed the variety of sex-determination systems in animals. In humans, sex determination is primarily conducted by the X and Y chromosomes, along with contributions from various autosomal genes. Specifically, the Y chromosome contains about 59 million nucleotides, or about 2 percent of the genome of males. The male-specific region differentiates the human sexes and is about 95 percent of the chromosome’s length. However, just as in the rest of the genome, most of the nucleotides on the Y chromosome are composed of noncoding sequences: Alu, retroviral, LINE1, and small repeat. There are only 156 transcriptional units, which encode about twenty-seven proteins.26 Of these, only eleven are primarily expressed in the testes. The sex-determining gene (SRY) is only 828 base pairs long. Yet the presence of the genes associated with the MSY portion (the male-specific region) of the Y chromosome do result in important differences between the biological sexes in our species.
Taken as a whole, human males and females are not that different biologically. I recognized that a long time ago as an athlete. Males tend to have more lean muscle mass and females tend to have more body fat for equivalent body mass indexes (BMIs). However, the means of the distributions are not so far apart that males and females of equivalent BMI can’t have similar values. With regard to athletic skill, there is even less difference. In the undergraduate co-ed recreational volleyball tournaments I played in, our varsity women players were generally better passers and setters than the men. The men were taller and better jumpers, but since volleyball depends so much on passing off the serve, the quality of the women on our team was a major factor in whether we won. In pickup basketball, women players were often passed over for men when teams were chosen. I never did that, especially since I knew several women whose skill sets (at guard) were just as good as those of a lot of the men at the court. I used to run a lot of games with one of my friends’ sisters, Natasha Parks, who was six foot, and we won a lot of games against all-male teams.
All mammalian (including human) embryos begin as females. Maleness happens relatively late in the developmental program, which explains why human males have nipples, even though their mammary glands are rudimentary and usually do not produce milk. This is one of my favorite arguments against special creationism.27 You have to jump through a lot of mental hoops to explain why an intelligent designer would require that design feature on males. On the other hand, there clearly are some sex-specific differences. Males generally lack ovaries, a uterus, and the other organs associated with reproduction. Reproduction also causes women to experience a different hormonal environment than do men, and this does have behavioral consequences. Male and female skeletal structures are also different: the human female pelvis differs from the male pelvis as an adaptation for birthing our large-headed offspring.28 These skeletal differences lead to sex-specific differences in posture. Childbirth has been and still is a significant cause of morbidity and mortality for women.29 So, although biologically males and females in our species do not display a great deal of sexual dimorphism, there is still enough difference to be worthy of study. Certainly the medical practice of ignoring female physiology and behavior and conducting biomedical research predominantly on male subjects must end.
Now, what of gender? Gender refers to social and/or cultural identities that do not necessarily correspond to biological sex. Not everyone agrees that there is a distinction between sex and gender. As with race concepts, lay individuals and professional scholars regularly conflate their meanings.30 Others, such as my old “friend” Charles R. Murray, reject the notion that gender has any meaning beyond that of biological sex. In his 2020 book Human Diversity Murray makes several strong claims. First, he claims that to make a distinction between sex and gender is meaningless, that there is no clear division between biological and socially constructed sex differences. He also predicts that that were will soon be a widely accepted comprehensive theory of sex differences, grounded in neuroscience, genetics, and evolutionary psychology.31 For Murray, the main neurological differences between males and females reside in the fact that men are “systematizers” and females are “empathizers.” He bases this distinction on the work of Simon Baron-Cohen on differences between male and female brains.32 Murray, in his section on sex differences, presents evidence from brain scans, claims of sexual dimorphism in human brains, and the activational effect of sex hormones. He also lists some phenotypic differences that he suggests support his thesis: females are better than males at social cognition; females are better than males at prosocial behavior; males are more likely to engage in impulsive behavior; females are more risk averse than males; and females display greater levels of emotional arousal than males. He also provides data on the differential prevalence of mental diseases between males and females. To summarize these data, he shows that there is a greater prevalence of mood disorders (such as major depressive disorder, bipolar II, generalized anxiety, posttraumatic stress, and bulimia) in females, whereas the autism/schizophrenia axis, conduct disorders, and alcohol/substance abuse are higher in males.
Modern neuroscience has convincingly demonstrated that humans display developmentally programmed sex differences associated with hormones and that there are some sexual dimorphisms in the structures of the human brain. However, the vast majority of these are subtle, involving patterns of connectivity and differences in specific regions of the brain. These claims are the subject of ongoing controversy.33 However, studies on the subject have also demonstrated that sex hormones act on the brain via a combination of genomic and nongenomic receptors. They have also shown the plasticity of human brain development, which means that, like every other phenotype, the brain is influenced by a combination of genetic, environmental, and epigenetic factors. Furthermore, as I demonstrated earlier in this chapter, sex has great evolutionary significance, so some of that genomic effect is the result of past natural and sexual selection. Sexual selection is exemplified by traits or behaviors that increase individuals’ reproductive success but that might not increase their survival; the showy plumage of the peacock is a well-known example.
However, recognizing that the brain is influenced by a combination of factors, including genetic factors that are the result of past natural and sexual selection, and claiming that evolutionary psychology is about to provide us with a comprehensive theory of sex differences in behavior are two very different things. At present the discipline of evolutionary psychology suffers from a distressing lack of testable hypotheses. For example, while there are solid reasons to believe that specific mental diseases may have resulted from past selection but are maladaptive in modern societies (evolutionary mismatch), providing the evidence to validate such a claim is particularly difficult. In part this is because we cannot re-create those past environments, and the reliability of testing evolutionary hypotheses on modern hunter-gatherers suffers from the amount of cultural contamination such groups routinely experience.
Thus, while one can make a solid case that the extremes of behavior of the biological sexes show strong differentiation, there is a solid region of overlap of personality and behavioral attributes between biological males and females. It is precisely here where the notion of gender resulting from socialization and culture finds its greatest utility. Furthermore, it is difficult to evaluate how much of these differences in behavior result from the social conditions themselves, conditions such as patriarchy. It makes a great deal of sense that women should be more risk averse in societies where women are far more likely to suffer sexual assault than males. It also makes a great deal of sense for women to be more prosocial than males in societies where the victims of war-related violence are overwhelmingly women and children. These attributes of female behavior could be products either of evolution or of simple common sense. In the end, does the origin of these behaviors really matter? What is important is that women are more likely to display them, and this is why I argued in the conclusion of my 2005 book The Race Myth that modern societies would be greatly advanced if all legislative bodies were required to be half women.34 Men did a great job of systematizing oppression; maybe women’s greater capacity to empathize with others would help us build more just societies.
THE NARRATIVE OF CREATION IN THE HEBREW BIBLE (OLD TESTAMENT) is unfortunate at many levels. It is obvious that the narrative does not explain the history of life on this planet, but one of its most harmful notions, which comes across clearly in Genesis 2, is that God created woman as an afterthought. It can easily be inferred from the language in Genesis 1:28 that God had no confusion concerning the need for there to be females: “Be fruitful and increase in number.” Increasing in number only happens by the reproductive effort of females. Furthermore, if we define “femaleness” as the capacity of an organism to reproduce, then the first organisms on this planet were asexual females. Sexual recombination did not always occur in the way we observe it now. Isogamy (gametes of equal size) was the condition of the universal ancestor of all eukaryotes.35 The evolution of anisogamy (small motile sperm cell, and large immobile egg cell) proceeded in all lineages of multicellular plants, fungi, and animals.
For example, at the base of the plant kingdom are single-celled organisms, such as the green algae Chlamydomonas. During unfavorable environmental conditions, sexual recombination is manifested by different mating strains. They are designated negative (−) and positive (+) strains, as their gametes are indistinguishable by observation (that is, they are isogametes). Negative strains cannot mate with negative strains, and positive strains cannot mate with positive strains. A closely related species within the volvocine green algae displays isogamy, but there are also those that display anisogamy (Figure 10.4).36 The organisms that produce the smaller motile cells are defined as “male.” We also know that hermaphrodites and organisms capable of switching sexes occurred in the ancestry of mammals. This would suggest that it was the presence of obligate “maleness” that was late to the ball game of life. Thus, from the perspective of evolutionary theory, males, not females, are the afterthought.
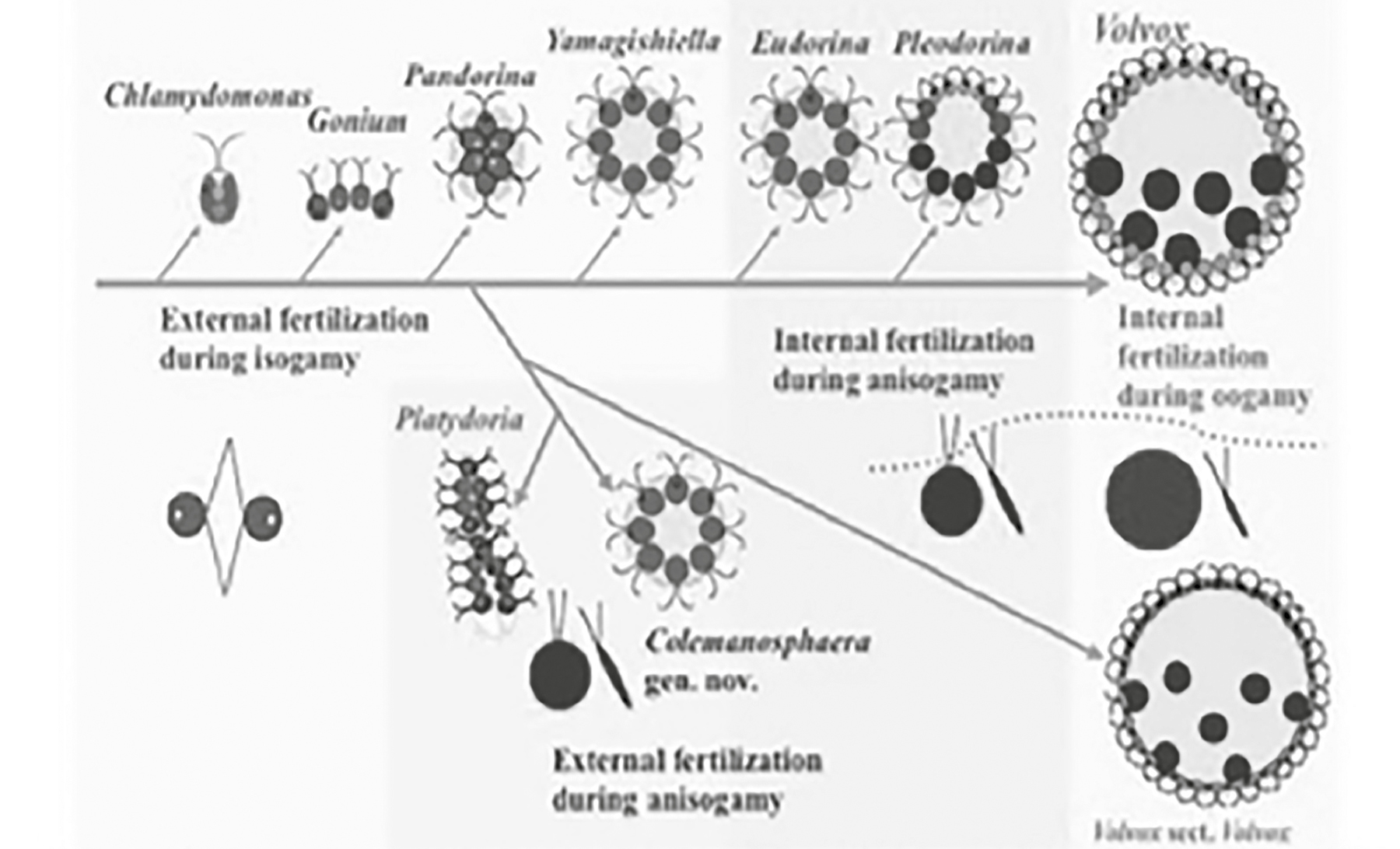
Figure 10.4. Evolution of isogamy in green algae. The single-cell species of genus Chlamydomonas show isogamy (+, −); the multicellular genera Eudorina and Pleodorina show internal fertilization with greater differentiation between gametes; and finally, the multicellular genus Volvox shows internal fertilization with oogamy (nonmotile egg cell). The branches in the diagram show how these genera are related to each other by descent with modification (evolution).
There are other examples of patriarchal thinking illustrated in Genesis. Biblical literalist interpretations of the human birth process claim that women are cursed to deliver their young in pain because of Eve’s sin (Genesis 3:16.) (The passage actually says God would greatly increase her pain in childbearing, possibly indicating that even without this event there might have been some pain.) Once again, evolutionary biology provides another explanation. We already know that the human female pelvis, which is relatively narrow in relation to the head size of our offspring, poses a problem in birth. Thus, human females experience more pain in childbirth than do other, closely related, primates (gorillas, chimpanzees, and orangutans).37 The increase in human head size and the relative narrowing of the human pelvis result from the mosaic history of human evolution. On the one hand is the increased head size of offspring (and delayed brain development); on the other is the upright posture and running concomitant with the reduced pelvis diameter. Daniel Lieberman argues that our capacity to run long distances, compared to other large-bodied animals, is a key adaptation in human evolution.38 However, these adaptations did not come without cost; human childbirth is risky, and until the age of modern medicine, childbirth was one of the largest causes of death among young women. That risk had effects on the social structure of our species. The capacity of women to form mutually supporting social networks may have played a crucial role in reducing the risk of death during childbirth.39
ALL THE PROHIBITIONS AGAINST GAY AND TRANSGENDER BEHAVIOR fall to pieces if the idea of a simple gender bifurcation into female and male is simply not true. As he was writing On the Origin of Species, Darwin was particularly impressed by the sexual characteristics and behavior of barnacles. The more he learned about them, the more he was convinced that his species theory was correct. He was astounded to find many species in which the males had become completely dependent upon the females (and were only found attached to the female body). In these species the entire male anatomy was reduced to simply being a “giant penis.”40 If this phenomenon were limited to barnacles, then the observation would not be particularly important; however, there are many species in which parasitic males are the norm. We have already established that hermaphroditic species are extremely common in nature, that separate sexes evolve under specific conditions, and that individuals can change their sex under specific conditions.41
Given that humans evolved from animal lineages with these features, why would anyone be surprised that human gender/sex determination is both complex and fluid? Instead of viewing human sex/gender as a dichotomy (male or female), it is more profitable to think of it as a continuum from individuals who look and behave in stereotypically female ways to those who look and behave in stereotypically male ways. To understand this, we must understand that sex/gender is influenced by events that occur at genetic, epigenetic, developmental, organ-differentiation (specifically of gonads and brains), and organ-system (bodily) levels. The expression of genes is influenced by chance and environmental effects, which can have important effects on development. The sex-determination system in humans is driven by the SRY region of the Y chromosome. Generally, individuals who are genetically XY develop testes and a penis. However, all human embryos start out with identical structures (that is, as females). Therefore, it is possible for individuals who are genetically XY to complete development in ways that produce external genitalia that are female. Anne Fausto-Sterling summarized the various common ways that embryos can undergo nondimorphic sexual development (Table 10.1). These data suggest that around 2 percent of live births are intersex individuals. These frequencies can differ by population as a result of genetic drift; for example, it has been shown that the frequency of hermaphrodites in the indigenous South African population is higher than the value reported in the table. However, some have argued that the number calculated by Fausto-Sterling for intersex births is too high; in particular, they argue, if individuals with Klinefelter syndrome or Turner syndrome (the two most common disorders in sex-chromosome numbers) are removed, the frequency of intersex births drops almost one hundredfold.42 (Individuals with Klinefelter syndrome have forty-seven chromosomes [XXY] and have genitalia and internal ducts that are usually male. Individuals with Turner syndrome have forty-five chromosomes [X0] and have rudimentary ovaries, short stature, and underdeveloped breast tissue.) The rationale for the latter calculation is the idea that the “intersex” category should be reserved for individuals whose chromosomal sex does not match the phenotype of their external bodies. However, given the common phenotypes of both Klinefelter and Turner syndromes, this definition seems overly restrictive. This issue can be extremely challenging for physicians and for the individuals who have intersex conditions. Prior to 2005 it was common for physicians to make decisions concerning the assignment of “biological” sex for individuals who had unambiguous genitalia at birth.43 However, in some cases, decisions as to which sex to best assign a child by surgical means had disastrous consequences, as in the well-publicized example of Brenda (originally named Bruce) Reimer. Brenda was born in 1965 with male genitalia and had the XY chromosome designation. However, due to a surgical accident during circumcision, his penis was destroyed. His family decided to accept the recommendations of John Money, an internationally respected surgeon from Johns Hopkins University. Money felt that he could surgically alter the baby to have female genitalia and that despite the absence of female reproductive organs, the child could be treated with hormones in such a way that “she” would grow up as a “normal” female. Surgical reassignment was conducted later that year. To make a long story short, it didn’t work. Brenda grew up feeling something was wrong. “She” was interested in stereotypically male pursuits and was consistently sexually attracted to women. Eventually Brenda had sex reassignment surgery as an adult and became David in 1980.44
Table 10.1 Percentage of intersex births
| Cause | Percent |
| Non-XX or non-XY (except Turner or Klinefelter syndromes) | 0.06390 |
| Turner syndrome | 0.03690 |
| Klinefelter syndrome | 0.09220 |
| Androgen insensitivity syndrome | 0.00760 |
| Partial androgen insensitivity syndrome | 0.00076 |
| Classic congenital adrenal hyperplasia | 0.00779 |
| Late-onset congenital adrenal hyperplasia | 1.50000 |
| Vaginal agenesis | 0.01690 |
| True hermaphrodites | 0.00120 |
| Idiopathic | 0.00009 |
| Total intersex births | 1.73 |
Source: Data from Fausto-Sterling A, Sexing the Body: Gender Politics and the Construction of Sexuality (New York: Basic Books, 2000).
WHEN IT COMES TO THE SEXUAL-BEHAVIOR ASPECT OF GENDER, particularly sexual orientation, knowing the frequency of intersex individuals is not particularly informative because what we think of as gender is influenced by a complicated interaction of genetic, cellular, physiological, and organic interactions. We know that there are a number of regions in the human brain that influence sexual orientation. For example, three areas in the hypothalamus are sexually dimorphic (SDN-POA, BSTc, and VIP-SCN). In nontransgendered heterosexual males, the BSTc is about 140 percent the size of the BSTc in nontransgendered heterosexual females (2.5 mm3 versus 1.75 mm3) and has about thirty-three thousand neurons compared to nineteen thousand neurons in nontransgendered heterosexual females’. The VIP-SCN size for homosexual men is bigger (1.7 times the size and 2.1 times the neurons) than that for heterosexual men and for heterosexual women.45 From this we conclude that gay male brains are different from straight male brains but are not equivalent to female brains in this region. Another region surrounding the hypothalamus, INAH3, measures 0.1 mm3 for heterosexual males and 0.05 mm3 for heterosexual women and gay males. One study found that the INAH3 subnucleus region of the anterior hypothalamus of male-to-female transgendered persons is similar in size and neuronal number to that of heterosexual females.46
If these were the only regions to control sexual orientation (which is highly unlikely), the number of combinations of brains resulting from these four regions would be 24 (= 16); and if we consider brain and body-type combinations (male/female organs and brain types), the number of combinations is 25 (= 32). Given that intersex individuals with ovo-testes exist, we could increase the number of combinations to 3 × 16 (= 48), suggesting that there are at least forty-eight different sexual orientation / gender possibilities in our species. In that case, it would be best to consider gender a continuous rather than a discrete attribute of individuals.
There are limitations to the utility of these studies, as virtually all of them were conducted with small sample sizes (usually less than fifty). Using positron emission tomography of sexual attraction, a study of twenty-five heterosexuals of each sex and twenty homosexuals of each sex found that cerebral and amygdule asymmetries of the homosexual group reflected those of the opposite sex in heterosexuals.47 This again suggests that gay individuals may inherit brain structures that more closely resemble those of the opposite sex, but how this might lead to attraction toward the same sex and whether such attraction is inevitable “from the womb” or is subject to social selection remain to be established. Thus, it is premature to conclude that we are born with a rigid sexual orientation. For example, identical twins, who share the same genes, display opposite sexual orientation around 50 percent of the time when one of the twins is gay.
SEXUAL ORIENTATION IS CLEARLY A COMPLEX TRAIT, WHICH means that genetics, epigenetics, environmental factors, and chance affect the way an individual’s sexual orientation is formed. Given the argument above, we expect that numerous genes would be involved in this trait, further supporting the notion of continuous variation in gender, from people who would be stereotypically female on one end of the continuum to those who would be stereotypically male on the other end. One of the earliest attempts to study the genetic basis of sexual orientation was published by Dean Hamer and colleagues in 1993.48 The lead author of this study, Dean Hamer, is an openly gay male who, when I met him, had an appointment as a genetics researcher at the NIH.
It is known that about 4 percent of men are gay across all human societies. The heritability of gay sexuality in men has been estimated at around h2 = 0.40. This is a very high value for the h2 of a complex trait. In addition, pedigree analysis has shown that gay men cluster in families.49 The Hamer study also found that gay men tended to cluster in families. It was based on genetic analysis of homosexual men (and their families) recruited from HIV clinics in the Washington, DC, area. The researchers found that the brother of a gay man had a 13 percent chance of being gay, whereas the brother of a heterosexual man had only about a 2 percent chance. This result suggests that genes are involved in determining gay male sexual orientation.
Subjects also rated their own sexuality on the Kinsey scale (from 0, for exclusive heterosexuality, to 6, for exclusive homosexuality). The Kinsey scale, published in 1948, resulted from the work of the pioneering sex researcher Alfred Kinsey. These ratings were revealing. For example, only 70 percent of the individuals who self-described as heterosexual claimed they had never experienced homosexual activity to orgasm, which means that 30 percent of the self-described heterosexual men admitted to having had some homosexual experience. Similarly, more than 40 percent of the men who described themselves as homosexual admitted to having had heterosexual activity to orgasm. This again underscores both the fluidity and the complexity of human gender and sexual orientation. Indeed, studies of how individuals rate their sexuality in online culture show a growing disconnection between the way people feel and the standard sexual categories of hetero- and homosexual.50
Modern genome-wide association (GWAS) studies of gay male sexual orientation perform as poorly as most of the rest of the GWAS studies of complex behavior in humans. GWAS is a method to show statistical associations between multiple genes across the genome to particular physical traits. They are usually structured in a case-and-control format. This means that the genomes of those displaying the trait (cases) are compared to those that do not display the trait (controls). One study published in 2017 found only one single nucleotide polymorphism (SNP) that reached the significance level of 10−8. An SNP refers to a nucleotide substitution at a particular point in the genome that is not shared by everyone. All living humans share 99.9 percent of their nucleotides in common. Thus, SNPs are what make us unique individuals. Because hundreds of thousands to millions of nucleotide positions are examined in GWAS, the statistical standard for a significant difference has to be much higher than usual studies that use probability values (p) of around 0.05 (or p = 0.05). In GWAS, the standard cutoff for statistical significance is p = 1 × 10−8, or one chance in one hundred million.
A follow-up study in 2019 that examined both female and male sexual orientation found only one SNP above the significance threshold (the level distinguishing significant from nonsignificant results) for both males and females, and none that were significant for either sex alone.51 So far, I can find no large-scale GWAS studies that have attempted to find genetic markers associated with female sexual orientation (hetero- or homosexual) in humans. It is pretty safe to say, based on the few reasonably powered GWAS studies that exist for humans, that sexual orientation is a multifactorial trait influenced by a large number of genes. The candidate gene approach, which is based on examining gene by gene for its impact on a trait, for this trait, which was powerfully hyped from the 1990s to the early 2000s, has simply failed to find anything significant.52
The Hamer study of the 1990s narrowed down the genetic contribution in the male homosexual population to five genetic loci on the tip of the X chromosome that were statistically significantly higher in gay males. If this is true, it raises an interesting evolutionary question: How could these genes circulate at such high frequency (about 4 percent) if homosexual males have a lower reproductive output than heterosexual males? No gene can increase in frequency by natural selection if its net impact is to lower the reproductive output of those that carry it. Two hypotheses were advanced to explain the existence of genes that increase the likelihood that their carrier will be gay.
Imagine that the role of these specific genetic markers is to signal that the carrier should be attracted to males. If the five markers are inherited together on the X chromosome, they can be symbolized as one locus, call it “s.” This results in five phenotypes: XX, “normal” woman; XsX, normal woman, possibly more highly attracted to men; XsXs, “super” woman, most highly attracted to men; XY, heterosexual man; and XsY, homosexual man. If we assume that only heterosexual men and heterosexual women are having children, then the predisposition to be a gay male can be produced by heterosexual males mating with any woman who carries the s allele. If the women carrying the s alleles are more likely to mate than women without it, that would explain the persistence of the “gay gene.” Obviously, this explanation only applies to gay maleness and doesn’t address any genetic contribution to lesbianism. There is no reason why a mechanism of this kind couldn’t be in operation. Such genes are labeled as “sexually antagonistic.”
A second hypothesis that has been offered to explain how gay behavior may evolve is the mechanism of kin-selection and multiple-gender families. Joan Roughgarden described this phenomenon in a number of nonhuman species. In short, in multiple-gender families individuals can contribute to the reproductive success of a relative even if they do not themselves reproduce. If multiple-gender families are more successful than dual-gender families, then genes contributing to the behavior would spread, even if they resulted in homosexual behavior. However, the simple fact is that we have no evidence supporting the claim that either the first or the second mechanism discussed here ever operates in humans. So it is best to see these as “possible” but not necessarily probable scenarios.
IN GENESIS 1:24 GOD SAYS, “LET THE LAND PRODUCE LIVING creatures according to their kinds: the livestock, the creatures that move along the ground, and the wild animals, each according to its kind.” Beyond this poetic language the passage does not describe specific mechanisms, and read literally, what it claims is wrong. Even if organisms reproduce after their kind, reproduction implies evolution, which means their kind will eventually give rise to a new kind. For this reason, anything in an organism’s reproductive biology is of great interest to evolutionary biologists. This is just as true for humans as for any other species. Applying evolutionary theories to sex and gender tells us a great number of things, including how someone could have a male-looking body, female genitalia, and female gender behavior (or any other combination). This means the fluidity of gender / sexual orientation in our species is “normal.” Taking this reality to its logical conclusion, we are long overdue in our efforts to overturn anti-LGBT bigotry. A case in point is North Carolina’s “crimes against nature” statute. As written, it applies mainly to oral and anal sexual acts, and in practice it is generally only enforced against homosexuals. The act is discriminatory and originated in the mythology of dichotomous gender and sex. Globally, gay marriage is only legal in twenty-nine countries (mostly in the global North). In most of the Islamic world, LGBT people can be imprisoned or put to death.53 The Christian world is locked in dispute over how to treat LGBT persons according to its interpretation of the scriptures. My denomination, the Episcopal Church USA, is the only Christian denomination that administers all the sacraments of the church to members without regard to their sexual/gender orientation.54 Some other denominations are also beginning to examine their treatment of LGBT individuals. One of my favorite ministers in our church is a lesbian woman, and I am good friends with her and her wife. Our position has caused trouble between the Episcopal Church USA and the Anglican Union.55 Several other denominations are friendly to LGBT people under the mantra “hate the sin, but love the sinner.” My answer to that is that if the biological processes that were put in motion by God result in a diversity of gender/sexual orientations, then it cannot be a sin to be LGBT and act on your fundamental human right to love whom you love.
Religious people are not the only ones who engage in anti-LGBT activity. Even those who claim not to be bigoted against LGBT people are allowing discrimination against them to continue by taking no action against the hegemony of heterosexual culture in American society. One of my most recent acts of political agitation was to demonstrate against anti-LGBT legislation in North Carolina. (There is a great picture of my wife and me in the Greensboro News and Record during the gathering against the bill.) Recently there has been a horrific uptick in violence against African American transwomen. In the summer of 2020, six African American transwomen were murdered.56 Despite these horrific crimes, there has been no national action. The violence and all remaining discrimination against LGBT people needs to be struck down immediately. It is time for a rational understanding of the human condition to determine our laws. Nations are judged by how their most vulnerable people are treated; if so America has woefully failed to pass muster.