To the question why we do not find rich fossiliferous deposits belonging to these assumed earliest periods prior to the Cambrian system, I can give no satisfactory answer…. Nevertheless, the difficulty of assigning any good reason for the absence of vast piles of strata rich in fossils beneath the Cambrian system is very great, and may be truly urged as a valid argument against the views here entertained.
—Charles Darwin, On the Origin of Species
Of all the distortions of the fossil record that the creationists promote, the worst is their version of the “Cambrian explosion.” The idea that most invertebrate fossils might first appear “suddenly” at the beginning of the Cambrian period with no fossils preceding them seems to suggest special creation. Creationists love to quote a variety of legitimate scientists about the “mystery” of the Cambrian explosion, although most of their quotes are grossly out of date, and many are out of context and say just the exact opposite when the full quote is read carefully. The quote by Darwin above is representative—but it was written over 150 years ago, when we knew very little about the Cambrian or Precambrian. This is true not only of the usual suspects, such as Gish, Morris, and Sarfati, but especially the intelligent design creationists, such as Meyer, Davis and Kenyon. Even the ID creationist biochemist Michael Behe (1996), who generally avoids talking about fossils because they are way outside his range of expertise, brings up the Cambrian explosion.
The problem with the creationists’ fascination with the Cambrian explosion is that it’s all wrong! The major groups of invertebrate fossils do not all appear suddenly at the base of the Cambrian but are spaced out over strata spanning 80 million years—hardly an instantaneous “explosion”! Some groups appear tens of millions of years earlier than others. And preceding the “Cambrian explosion” was a long slow buildup to the first appearance of typical Cambrian shelled invertebrates.
In this chapter, we’ll go step by step through many of the recent developments that turned the Cambrian explosion into the “Cambrian slow fuse.” Let us hope that even if creationists can’t keep up with the recent discoveries in science, their audiences will know when they hear such baloney.
For four-fifths of our history, our planet was populated by pond scum.
—J. W. Schopf, Cradle of Life
The solution to Darwin’s dilemma about the lack of apparent fossils in Precambrian rocks is that we were looking for them in the wrong way. The fossils were there all along, but they are nearly always microscopic. It wasn’t until the 1940s and 1950s that Stanley Tyler and Elso Barghoorn found cherts and flints like the 2-billion-year-old Gunflint Chert in Canada that preserve these delicate microfossils and made it possible for us to study them. So the answer to the first creationist misconception about the Precambrian fossil record is yes, there are many fossils before the trilobites—but you need a microscope to see them, and they’re only preserved in certain circumstances.
We have already discussed in chapter 6 the evidence for the earliest fossils (fig. 6.2) from rocks 3.5 billion years old in Australia and 3.4 billion years old in South Africa, as well as organic carbon and possible stromatolites from rocks 3.8 billion years in age. These fossils are all of simple prokaryotic bacteria and cyanobacteria (formerly but incorrectly known as “blue-green algae”). Their equivalents among the modern cyanobacteria are virtually indistinguishable from their fossil counterparts, showing that they have evolved very little (at least in the external anatomical sense) for the last 3.5 billion years. The earliest forms of life made simple microbial mats on the seafloor, and that way of living was so successful that they saw no reason to change it since.
As it was in the beginning (3.5 billion years ago), so it was for almost another 2 billion years. There are hundreds of microfossil localities around the world (see Schopf [1983] and Schopf and Klein [1992] for documentation of these fossils) in rocks dated between 3.5 and 1.75 billion years ago, and they yield plenty of good examples of prokaryotes (and occasionally their macroscopic sedimentary structures, the layered fossilized bacterial mats known as stromatolites). Schopf (1999) calls this extraordinarily slow rate of evolution hypobradytely, after George Gaylord Simpson’s (1944) term for slow rates of evolution (bradytely) with the additional prefix “hypo-” (meaning “below”) to indicate that cyanobacteria evolve slower than anything we know. Indeed, they show almost no visible change in 3.5 billion years. Everywhere we look in rocks between 3.5 billion years old and about 1.75 billion years old, we see nothing more complicated than prokaryotes and stromatolites. The first fossil cells that are large enough to have been eukaryotes do not appear until 1.75 billion years ago, and multicellular life does not appear until 600 million years ago. For almost 2 billion years, or about 60 percent of life’s history, there was nothing on the planet more complicated than a bacterium or a microbial mat, and for almost 3 billion years, or 85 percent of Earth’s history, there was nothing more complicated than single-celled organisms. It was truly the “planet of the scum.” If aliens existed and had visited the planet long ago, odds are they would have come at a time when there was nothing more interesting to see (fig. 7.1) than mats of cyanobacteria—and they would have probably blasted off immediately because this planet was so boring (unless they studied cyanobacteria, in which case it would be exciting).

FIGURE 7.1. A diorama of how life might have looked on this planet for almost the first 80 percent (3 billion years) of its history. The most complex form of life was the cyanobacterial mats forming domed stromatolites. If an alien had visited earth through most of its first 3.5 billion years, it would have seen nothing more complex than a mat of scum on the surf zone and probably left unimpressed with life on this planet. (Drawing by Carl Buell)
Humans like to think of themselves as special and the center of creation, but that anthropocentric view of the universe has been shocked again and again by the discovery (starting with Copernicus) that the earth is a minor planet in a small solar system on the fringe of an immense universe, and the discovery (starting with Hutton) that geologic time is immensely long and humans appeared in only the very last part of the age of the earth. Add to that the fact that most of life’s history is characterized by nothing more complicated than pond scum and that humans appeared in a tiny fraction of the final 1 percent of life’s history, and the blow to our cosmic arrogance is complete. Mark Twain said it best, “If the Eiffel Tower were now representing the world’s age, the skin of paint on the pinnacle-knob at its summit would represent man’s share of that age, and anybody would perceive that that skin was what the tower was built for. I reckon they would, I dunno.”
Aspiring paleontologists are typically attracted to the large, flashy specimens such as carnivorous dinosaurs and Pleistocene mammals. But to find the real monsters, the weird wonders of lost worlds, one must turn to invertebrate paleontology. Without question the strangest of all fossilized bodies are to be found among the Ediacarans.
—Mark McMenamin, The Garden of Ediacara
The next step in our progression from single-celled life to Cambrian trilobites is the appearance of fossils of multicellular life. Contrary to the creationists’ myths, we have abundant fossils in rocks older than the Early Cambrian (prior to 545 million years ago). Some of these date to 600 million years ago, and they are known as the Ediacara fauna (pronounced “Ee-dee-AK-ara”). This period of time from 600 million to the beginning of the Cambrian 545 million years ago is known as the Ediacaran Period of the Proterozoic Era. First discovered in the Rawnsley Quartzite in the Ediacara Hills of Australia by Reg Sprigg in 1946, the Ediacara fauna is now known from a wide variety of localities around the world, including many spectacular localities in China, Russia, Siberia, Namibia, England, Scandinavia, the Yukon, and Newfoundland. Most of these fossils (fig. 7.2A) are the impressions of soft-bodied organisms without skeletons, so there are no hard parts that make up the bulk of the later fossil record. Instead, these impressions have reminded some paleontologists (such as Martin Glaessner, who studied the classic Australian Ediacara fauna) of the impressions made by sea jellies, worms, soft corals, and other simple nonskeletonized organisms. Over 2,000 specimens are known, usually placed in about 30–40 genera and about 50–70 species, so they were relatively diverse.
Although the Ediacara fauna clearly represents fossils of multicellular organisms (some reach almost a meter in length), paleontologists have a wide spectrum of opinions about what made these impressions. The more conventional interpretation (fig. 7.2D) is that they are like fossils of groups we know today: sea jellies, sea pens, and worms of various sorts. Some do look like sea jellies, but if so, they have symmetry unlike any living sea jellies. Others vaguely resemble some of the known marine worms, although their symmetry and segmentation do not match any groups of worms alive in the ocean today. Nor do the “worms” have evidence of eyes, mouth, anus, locomotory appendages, or even a digestive tract.
For this reason, other paleontologists have suggested that the Ediacara fauna was composed of organisms unlike any that are alive today. They point to the lack of modern patterns of symmetry and the apparent large size of many of the fossils and argue that they are an early failed experiment in multicellularity. Adolf Seilacher (1989), for example, calls them the “Vendozoa” and suggests that they were constructed in a quilted or “water-filled air mattress” fashion that maximizes surface area. Instead of using internal digestive and circulatory systems to solve the problem of large multicellular bodies, Seilacher suggests that these simple organisms had no internal organs but instead received all their nutrients and oxygen and got rid of waste through the huge surface area of their outer membranes. Mark McMenamin (1998) suggested that they housed symbiotic algae (as do many living large invertebrates, such as reef corals and giant clams). In his “garden of Ediacara” hypothesis, McMenamin suggests that the large surface area of the Ediacarans maximizes the area of exposure of sunlight for these internal algae, which then help such large organisms metabolize. Still other hypotheses (such as the ideas that they are lichens, proposed by Greg Retallack) have been put forth. Unfortunately, because the Ediacarans are known entirely from the impressions on the soft sea bottom and not from any body fossils with internal organs or other important features, it is very difficult to resolve this controversy. Whatever the biological affinities of the Ediacara fauna, it is very clear that they are multicellular organisms, whether animals, plants, fungi, or some early experimental kingdom not in any living group.

FIGURE 7.2. The Ediacara fauna consists of the impressions of soft-bodied fossils without skeletons, whose biological affinities are still controversial. (A) The wormlike segmented creatures Dickinsonia, which reached almost a meter in length. (B) The elongated segmented creature Spriggina. (C) The odd form Parvancorina, which has been linked to arthropods. (D) A reconstruction of the Ediacaran community, with most of the fossils assumed to be related to jellyfish, sea pens, and worms. (Photos courtesy of the Smithsonian Institution)
Even more intriguing is the fact that some molecular clock estimates of the divergence times of the major invertebrate groups (fig. 5.7) place the branching points of the major invertebrates as old as 800 to 900 million years ago (Runnegar 1992; Wray et al. 1996; Ayala et al. 1998). We still do not have any fossils nor even any undisputed burrows or other evidence to support this prediction, but either way, it is clear that advanced multicellular life (but still soft-bodied without any trace of fossilizable skeletons) was on earth 600 million years ago (more than 50 million years before the Cambrian), and possibly as early as 900 million years ago.
The wave of discoveries that rewrote the story of the earliest Cambrian began when the former Soviet Union mustered sizable teams of scientists to explore geological resources in Siberia after the end of World War II. There, above thick sequences of Precambrian sedimentary rocks, lie thinner formations of early Cambrian sediments undisturbed by later mountain-building events (unlike the folded Cambrian of Wales). These rocks are beautifully exposed along the Lena and Aldan rivers, as well as in other parts of that vast and sparsely populated region. A team headed by Alexi Rozanov of the Paleontological Institute in Moscow discovered that the oldest limestones of Cambrian age contained a whole assortment of small and unfamiliar skeletons and skeletal components, few bigger than 1/2 in (1 cm) long. These fossils have been wrapped in strings of Latin syllables but have been more plainly baptized in English as the “small shelly fossils” (SSFs for short).
—Jack Sepkoski, The Book of Life
If the soft-bodied multicellular (but nonskeletonized) Ediacara fauna represents the next logical step up from single-celled life, then the next step beyond that would be the appearance of mineralized, fossilizable skeletons. But if life took almost 3 billion years to develop the ability to mineralize shells, we expect that it would be a difficult process and would not arise fully fledged. Sure enough, the earliest stages of the Cambrian (known as the Nemakit-Daldynian and the Tommotian stages, from 520 to 545 million years ago) are dominated by tiny (only a few millimeters) fossils nicknamed the “little shellies” or the “small shelly fossils (SSFs)” in the trade (fig. 7.3). For decades, these little fossils were overlooked as people hunted the beds above them for the more spectacular trilobite fossils. But as the quotation from Sepkoski points out, the Soviets were the first to study their long and detailed sequences of Proterozoic and Cambrian sediments in detail and named the stages of the Cambrian. And when they looked closer at the beds below the trilobites and took samples back to the lab to dissolve in acid or slice into thin sections, it became apparent that these long-neglected beds were chock-full of tiny fossils.

FIGURE 7.3. The earliest stages of the Cambrian (Nemakit-Daldynian and Tommotian) do not produce trilobites but are dominated by tiny phosphatic fossils nicknamed the “little shellies.” Some may have been mollusk shells (E, H, and I), while others are apparently sponge spicules or pieces of the “chain-mail armor” of larger creatures, such as worms. (A) Cloudina hartmannae, one of the earliest known skeletal fossils, from the same beds that produce Ediacaran fossils in China. (B) A spicule of a calcareous sponge. (C) A spicule of a possible coral. (D) Anabarites sexalox, a tube-dwelling animal with triradial body symmetry. (E) A spicule from a possible early mollusk. (F) Lapworthella, a cone-shaped fossil of unknown relationships. (G) A skeletal plate of Stoibostromus crenulatus, another creature of unknown relationships. (H) Skeletal plate of Mobergella, a possible mollusk. (I) Cap-shaped shell of Cyrtochites, another possible mollusk. All scale bars = 1 mm. (Photos courtesy S. Bengston)
Some of the little shellies look like simple cap-shaped or coiled mollusks; others look like primitive clams (fig. 7.3). Many others are simple tubular or conical fossils whose connections to any living group are hard to establish. Many of the fossils look like miniature jacks or little spiky Christmas ornaments. These appear to have been part of the “chain-mail armor” that studded the skins of larger organisms, such as the sponge Chancelloria, that were soft-bodied except for these little spiky objects (much like the tiny spikes in the skin of a shark or a sea cucumber). Most of the little shellies were made of calcium phosphate, the same mineral that makes up the bones of vertebrates. Today, most marine invertebrate shells are made of calcium carbonate (the minerals calcite and aragonite). To some scientists, this suggests that some sort of environmental condition (such as low atmospheric oxygen) made it hard to secrete calcite skeletons, but phosphate skeletons were easier to produce. They suggest that the appearance of large calcified trilobites and other fossils reflects the point where atmospheric oxygen passed a critical threshold and became abundant enough to allow this chemical mineralization.
Whatever the reason, for almost 25 million years, the Cambrian explosion was burning on a slow fuse. The little shellies were abundant, but larger fossils were not. The earliest sponges had already appeared back in the late Ediacaran, but this is not surprising, considering that all lines of evidence show that sponges are the most primitive animals alive today (fig. 5.6). By the Tommotian Stage (530 million years ago), a slow trickle of other groups of larger invertebrates began to appear, including the first “lamp shells” (brachiopods), and also members of an extinct spongelike group known as archaeocyathans. Diversity in the Tommotian reached only about 50 genera, about the same as in the Vendian. In addition, the sediments of the earliest Cambrian show abundant burrowing, proof that many other types of soft-bodied worms with a true internal fluid-filled cavity (a coelom) must have lived at that time. Thus the earliest Cambrian shows evidence of a gradual increase in diversity from the Vendian, but no “explosion.”
Was there really a Cambrian Explosion? Some have treated the issue as semantic—anything that plays out over tens of millions of years cannot be “explosive,” and if the Cambrian animals didn’t “explode,” perhaps they did nothing at all out of the ordinary. Cambrian evolution was certainly not cartoonishly fast…. Do we need to posit some unique but poorly understood evolutionary process to explain the emergence of modern animals? I don’t think so. The Cambrian Period contains plenty of time to accomplish what the Proterozoic didn’t without invoking processes unknown to population geneticists—20 million years is a long time for organisms that produce a new generation every year or two.
—Andrew Knoll, Life on a Young Planet
The third stage of the Early Cambrian is known as the Atdabanian Stage (515–520 million years ago), and with this stage, we finally see a great increase in diversity: more than 600 genera are recorded (fig. 7.4). However, this number is misleading and a bit inflated. Most of the genera are trilobites, which fossilize readily and so greatly increase the volume and diversity of large shelly fossils. Most of the other animal phyla had already appeared by this time (including mollusks, sponges, corals, echinoderms) or would appear later in the Cambrian (vertebrates) or even in the Ordovician Period that followed (e.g., the “moss animals” or bryozoans).
The second misleading aspect of this apparent diversity “explosion” is that during the Atdabanian Stage we get the first good fauna of soft-bodied fossils (the Chinese Chengjiang fauna), so we get the apparent (but not real) first appearance of phyla only known from soft tissues. Then in the Middle Cambrian we have the extraordinary soft-bodied preservation of fossils from places like the Burgess Shale in Canada (fig. 7.5). As pointed out by Stephen Jay Gould in his book Wonderful Life (1989), the soft-bodied animals preserved in these amazing deposits allow us to see what the normal fossil record is missing. We have many bizarre wormlike and odd fossils, many of which don’t fit into any living phylum. Some, like the five-eyed nozzle-nosed Opabinia (fig. 7.5, top left) or the soft flowerlike Dinomischus, are complete mysteries to zoologists. Others are apparently soft-shelled arthropods. One fossil, appropriately named Hallucigenia (fig. 8.17B), was a bizarre creature that seemed to have tentacles or spikes on a wormlike body, until recent better fossils from China showed it is related to the “velvet worms,” phylum Onychophora (discussed in chapter 8). The largest predator (about 2 feet long) was a soft-bodied swimmer known as Anomalocaris, which had a strange mouth that looked like a pineapple slice and was originally found and misinterpreted as a sea jelly.
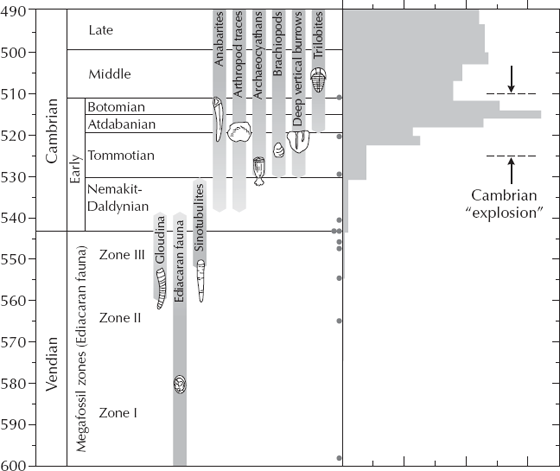
FIGURE 7.4. A detailed examination of the stratigraphic record of fossils through the late Precambrian and Cambrian shows that life did not “explode” in the Cambrian, but appeared in a number of steps spanning almost 100 million years. The large soft-bodied Ediacaran fossils (fig. 7.2) appeared first at 600 million years ago, in the late Precambrian (Vendian). Toward the end of their reign, we see the first tiny shelly fossils, including the simple conical Cloudina and Sinotubulites. The Nemakit-Daldynian and Tommotian stages are dominated by the “little shellies” (fig. 7.3) plus the earliest lamp-shells, or brachiopods, and the conical spongelike archaeocyathans, and many burrows showing that wormlike animals without hard skeletons were also common. Finally, in the third stage of the Cambrian (Atdabanian, around 520 million years ago), we see the radiation of trilobites and a huge diversification in total number of genera (histograms on the right side of the diagram). Thus the Cambrian explosion took over 80 million years to develop and was no “sudden” event, even by geological standards. (Modified from Dott and Prothero 2010: fig. 9.14, and from Kirschvink et al. 1997: fig. 1. Copyright © 1997 American Association for the Advancement of Science. Reprinted with permission.)
Thus we have seen that the “Cambrian explosion” is a myth. It is better described as the Cambrian slow fuse. It takes from 600 to 520 million years ago before the typical Cambrian fauna of large shelly organisms (especially trilobites) finally develops. Eighty million years is not explosive by any stretch of the imagination! Not only is the explosion a slow fuse, but it follows a series of logical stages from simple and small to larger and complex and mineralized. First, of course, we have microfossils of cyanobacteria and other eukaryotes going back to as far as 3.5 billion years ago and spanning the entire fossil record since that ancient time. Then, about 600 million years ago, we get the first good evidence of multicellular animals, the Ediacara fauna. They are larger and multicellular but did not have hard shells. The earliest stages of the Cambrian, the Nemakit-Daldynian and Tommotian stages, are dominated not by the little shellies, which were just beginning to develop small mineralized skeletons. Only after several more steps do we see the full Cambrian fauna. In short, the fossil record shows a gradual buildup from single-celled prokaryotes and then eukaryotes to multicellular soft-bodied animals to animals with tiny shells, and finally by the middle Cambrian, the full range of large shelled invertebrates. This gradual transformation by logical advances in body size and skeletonization bears no resemblance to an instantaneous Cambrian explosion that might be consistent with the Bible but instead clearly shows a series of evolutionary transformations.

FIGURE 7.5. Examples of the extraordinary soft-bodied middle Cambrian fauna from the Burgess Shale, near Field, British Columbia. Note the exquisite preservation of fine detail, including appendages and other soft tissues. (Photos courtesy of the Smithsonian Institution)
All of this information has been known for at least the past few decades, and the first Precambrian microfossils were discovered over 70 years ago. They are published in all the standard geology and paleontology textbooks and have been for decades. But the creationists either don’t want to know or cannot understand the implications of these discoveries. Their out-of-context quotations of real scientists puzzling about the Cambrian explosion are all from old sources that do not reflect what we have learned from recent discoveries. Even their most recent books, including the “intelligent design” texts, persist in perpetuating this out-of-date picture. A few years ago, I debated an ID creationist on the KPCC radio station in Los Angeles, and as soon as he mentioned the Cambrian explosion, it was clear he didn’t know anything about paleontology and hadn’t heard that the Cambrian explosion was a myth.
In 2013, ID creationist Stephen Meyer published an entire book on the topic entitled Darwin’s Doubt: The Explosive Origin of Animal Life and the Case for Intelligent Design. Most scientists ignored it, but the few who wasted their time reading it lambasted it (Cook 2013; Marshall 2013). As I wrote in my own review (Prothero 2013), the book is a piece of incompetent scholarship from one end to the other, with mistakes, misstatements, quote mining, cherry-picking of data, ignoring inconvenient facts, and outright lies about the fossil record in nearly every page. This is not surprising, since Meyer has no formal training in paleontology (his Ph.D. is in history of science), and no published research in paleontology, so everything he cites comes across as amateurish and filtered by his creationist biases. There is no space in this short chapter to list all the errors and lies in his book (for details, see Prothero 2013). The most crucial deception that Meyer pulls is that he completely ignores the first two stages of the Cambrian! Nowhere in the book are the “little shellies” or the Nemakit-Dalydinian or Tommotian stages even mentioned! Naturally, if you deliberately leave out the crucial evidence of the intermediate stage of life evolving from large soft-bodied Ediacarans (which he dismisses as irrelevant because we’re not sure they are members of modern phyla) to the large shelly trilobites of the third stage of the Cambrian (Atdabanian), it will look more explosive. I even debated him on this topic in Hollywood in 2009, and he dodged the issue entirely, so it cannot be a case of him not knowing any better. No, he’s fully aware that this evidence would invalidate his entire book, so he ignores it—and he counts on his readers to not know the difference.
Even if we grant the premise that a lot of phyla appear in the Atdabanian (solely because there are no soft-bodied faunas older than Chengjiang in the earliest Cambrian), Meyer claims the 5–6 million years of the Atdabanian are too fast for evolution to produce all the phyla of animals. Wrong again! Lieberman (2003) showed that rates of evolution during the “Cambrian explosion” are typical of any adaptive radiation in life’s history, whether you look at the Paleocene diversification of the mammals after the nonavian dinosaurs vanished, or even the diversification of humans from their common ancestor with apes 6 million years ago. As distinguished Harvard paleontologist Andrew Knoll put it in his book, Life on a Young Planet (also cited in the epigraph to this section), it wasn’t an “explosion,” nor was it “cartoonishly fast.”
Finally, one might wonder: what’s all the fuss about the “Cambrian explosion”? Why should it matter whether evolution was fast or slow during the third stage of the Cambrian? Some scientists might find this puzzling, but you must understand the minds of creationists. They operate by a god of the gaps argument: anything that is currently not easily explained by science is automatically attributed to supernatural causes. Even though ID creationists say that this supernatural designer could be any deity or even extraterrestrials, it is well documented that they are thinking of the Judeo-Christian god when they point to the complexity and “design” of life. They argue that if scientists haven’t completely explained every possible event of the early Cambrian, science has failed and we must consider supernatural causes.
Again and again, creationists persist in presenting a version of the Cambrian that is at least 70 years out of date either because they don’t know any better (the “clueless” hypothesis) or because they do know better (the “deceiver” hypothesis, of which Meyer is a good example). Either way, it is bad science.
For Further Reading
Ayala, F. J., and A. Rzhetsky. 1998. Origins of the metazoan phyla: molecular clocks confirm paleontological estimates. Proceedings of the National Academy of Sciences USA 95:606–611.
Briggs, D. E. G., and R. A. Fortey. 2005. Wonderful strife: systematics, stem groups, and the phylogenetic signal of the Cambrian radiation. Paleobiology 31(2):94–112.
Conway Morris, S. 1998. The Crucible of Creation. Oxford: Oxford University Press.
Conway Morris, S. 2000. The Cambrian “explosion”: slow-fuse or megatonnage? Proceedings of the National Academy of Sciences USA 97:4426–4429.
Cook, G. 2013. Doubting “Darwin’s doubt.” New Yorker, July 2, 2013.
Erwin, D., and J. W. Valentine. 2013. The Cambrian Explosion: The Construction of Biodiversity. New York: Roberts.
Glaessner, M. F. 1984. The Dawn of Animal Life. New York: Cambridge University Press.
Gould, S. J. 1989. Wonderful Life: The Burgess Shale and the Nature of History. New York: Norton.
Grotzinger, J. P., S. A. Bowring, B. Z. Saylor, and A. J. Kaufman. 1995. Biostratigraphic and geochronologic constraints on early animal evolution. Science 270:598–604.
Knoll, A. H. 2003. Life on a Young Planet: The First Three Billion Years of Evolution on Earth. Princeton, N.J.: Princeton University Press.
Knoll, A. H., and S. B. Carroll. 1999. Early animal evolution: emerging views from comparative biology and geology. Science 284:2129–2137.
Lieberman, B. S. 2003. Taking the pulse of the Cambrian radiation. Integrative and Comparative Biology 43:229–237.
Marshall, C. R. 2013. When prior beliefs trump scholarship. Science 341:1344.
McMenamin, M. A. S. 1998. The Garden of Ediacara. New York: Columbia University Press.
McMenamin, M. A. S., and D. L. S. McMenamin. 1990. The Emergence of Animals, the Cambrian Breakthrough. New York: Columbia University Press.
Narbonne, G. M. 1998. The Ediacara biota: a terminal Neoproterozoic experiment in the evolution of life. GSA Today 8(2):1–6.
Peterson, K., M. A. McPeek, and D. A. D. Evans. 2005. Tempo and mode of early animal evolution: inferences from rocks, Hox and molecular clocks. Paleobiology 31:36–55.
Prothero, D.R. 2013. Stephen Meyer’s fumbling bumbling Cambrian follies: a review of Darwin’s Doubt by Stephen Meyer. Skeptic 18(4):50–53.
Runnegar, B. 1992. Evolution of the earliest animals. In Major Events in the History of Life. ed. J. W. Schopf. New York: Jones and Bartlett, 65–94.
Schopf, J. W., ed. 1983. Earth’s Earliest Biosphere: Its Origin and Development. Princeton, N.J.: Princeton University Press.
Schopf, J. W. 1999. Cradle of Life: The Discovery of the Earth’s Earliest Fossils. Princeton, N.J.: Princeton University Press.
Schopf, J. W., and C. Klein, eds. 1992. The Proterozoic Biosphere, a Multidisciplinary Study. New York: Cambridge University Press.
Seilacher, A. 1989. Vendozoa: organismic construction in the Proterozoic biosphere. Lethaia 22:229–239.
Seilacher, A. 1992. Vendobionta and Psammocorallia. Journal of the Geological Society of London 149:607–613.
Valentine, J. W. 2004. On the Origin of Phyla. Chicago: University of Chicago Press.
Wray, G. A., J. S. Levinton, and L. H. Shapiro. 1996. Molecular evidence for deep Precambrian divergences among metazoan phyla. Science 274:568–573.