1. Overcoming the effects of habitat loss and fragmentation
2. The first principle is to preserve large habitat areas
3. Other principles reconcile ecological and economic trade-offs
4. Principles that increase the effective area of reserves
5. Identify conservation targets
8. Revisiting the Y2Y corridor
Perhaps the greatest challenge to biodiversity conservation is overcoming the devastating effects of habitat loss. The single best approach to preserve biodiversity is to conserve or restore large habitat areas. Yet, in landscapes dominated by farming, grazing, and development that support increasing human populations, there are limits to the areas that can be conserved. Given limited areas for conservation, can reserves of fixed area be designed to increase their value for biodiversity conservation? Ecological theory suggests that strategies that increase habitat connectivity and reduce negative edge effects will have higher conservation benefits.
connectivity. The degree to which the landscape facilitates movement
corridor. Habitat that connects two or more reserves, usually the same type as found in a reserve but long and thin relative to reserve size
ecological trap. The attraction of animals to habitats where they perform more poorly, even when higher-quality habitat is available
edge effects. Changes in population sizes, species richness, or other aspects of the ecology of individuals, populations, or communities at the interface between two habitat types
habitat fragmentation. The spatial isolation of small habitat areas that compounds the effects of habitat loss on populations and biodiversity
matrix. The habitat or land use, often urban, agricultural, or degraded habitat, surrounding native habitats in reserves
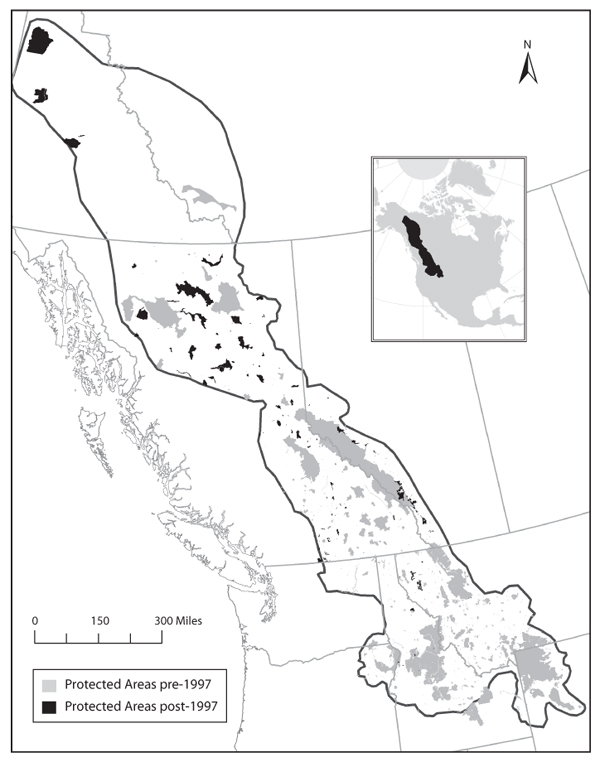
The Yellowstone-to-Yukon (Y2Y) Corridor is perhaps the grandest application of ecological theory to the design of nature reserves. When complete, the reserve network would extend 1800 miles northward from Yellowstone National Park in the northwestern United States into the Yukon Territory in Canada (figure 1). It is among the most expensive applications of ecological theory in history. Conservation organizations are actively investing tens of millions of dollars in this region to protect large and connected habitats. If successful, the corridor would conserve large predators such as grizzly bears and wolves and wide-ranging ungulates such as bison and caribou by providing safe passage between the Yellowstone and Yukon regions. Ideally, conserving these large vertebrate species would create a metaphorical umbrella and also cover smaller vertebrates as well as the invertebrates, fungi, and plants that make up the bulk of biodiversity.
Conservation efforts such as Y2Y have to answer the question: Can reserves be designed to enhance biodiversity protection by targeting key parcels of land? To answer this question, there are a number of more basic questions we must address first. The most obvious is: How do we design reserves? Stated simply, reserve designs seek to increase the effective (if not the actual) area of reserves. The answer to this first question leads to others, such as: Which design criteria are most effective? Is reserve design simply a matter of “more is better”? And questions about ecological effectiveness must be evaluated in the context of scarce conservation resources. When one must actually invest tens of millions of dollars, which strategies do we have enough confidence in to spend money on?
Figure 1. The Yellowstone to Yukon (Y2Y) Corridor. Within the Y2Y region, areas shaded in gray were protected before 1997, the year the Y2Y Conservation Initiative began. Areas shaded in black have received some level of protection since 1997.
Y2Y exemplifies a growing number of conservation plans that incorporate reserve design strategies and, we hope, maximize biodiversity protection. This chapter reviews the problem of habitat loss that necessitates reserve design. The easiest solution—restoring the habitat that was lost—quickly runs up against financial limitations, spurring other elements of design. The remainder of this chapter then focuses on aspects of design that seek to increase the effective area of reserves, specifically identifying conservation targets, reducing edge effects, and increasing habitat connectivity. At their core, the principles of reserve design draw on ecological theory and target conservation dollars to maximize biodiversity preservation. In this chapter, those theories—discussed in a number of other chapters—are put in their conservation context. It is here, in spending the big money, that the theoretical rubber hits the practical road.
What is our goal in designing reserves? It is an overly simple question but perhaps one worth considering. Principles of reserve design are applied to retard or reverse biodiversity loss caused by the destruction, fragmentation, and degradation of natural habitat (see chapter V.1). Over 40% of the world’s land area has been transformed or degraded by humans through agriculture, forestry, and urbanization. The severity of this threat has made it the leading cause of biodiversity loss. And the threat continues to increase as the human population grows.
At its core, reserve design seeks to minimize local and global extinctions of species. The immediate threat that habitat loss poses to biodiversity is that it reduces population sizes of plants and animals. This initial consequence of habitat loss is insidious, as smaller populations are then exposed to other threats that compound extinction risk. Small populations are at greater risk to random events, such as catastrophic fires or population fluctuations that skew population structures toward one gender and limit mating potential. Smaller populations suffer more from random genetic events such as inbreeding or genetic drift that tend to reduce population fitness (see chapter V.2). All of these consequences of living in small populations drive populations to smaller and smaller sizes, ultimately causing extinction. The principles of reserve design are intended to overcome these negative effects of habitat loss by increasing population size and increasing the viability of rare species.
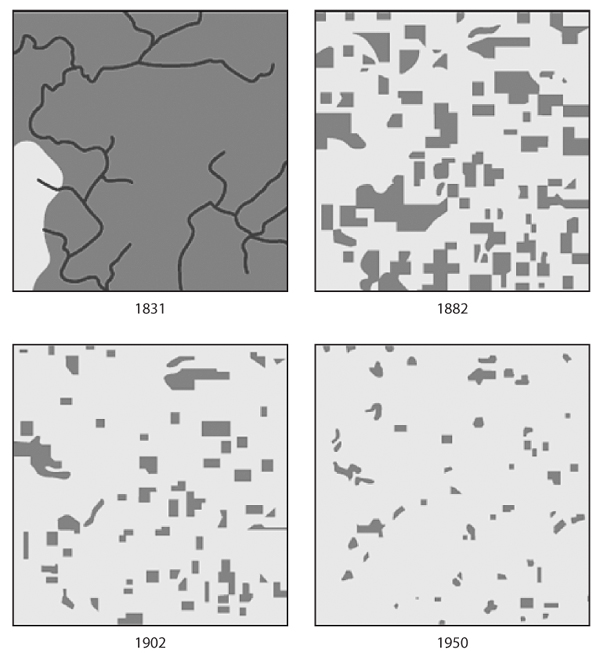
Figure 2. Habitat loss from 1831 to 1950. Cadiz Township, Wisconsin, 1831–1950. (After Curtis, J. T. 1956. The modification of midlatitude grasslands and forests by man. In W. L. Thomas Jr., ed. Man’s Role in Changing the Face of the Earth. Chicago: University of Chicago Press. Copyright Pearson Education, Inc., publishing as Benjamin Cummings)
The principles of reserve design are also intended to overcome the interrelated effects of habitat fragmentation. Figure 2 portrays a landscape in the midwestern United States where, over the course of the past 150 years, expanses of forest have been converted to agriculture. The forest left behind is not protected neatly in one reserve. Instead, the landscape is chopped into tiny fragments, woodlots that are spread among agricultural fields. Viewed in this way, populations are not just reduced by the amount of habitat lost; rather, single populations of plants and animals are now divided into larger numbers of much smaller populations. If those plants and animals are unable to move between woodlots, then habitat fragmentation further exacerbates the factors that threaten small populations and hastens extinction. The crux of reserve design principles is to account for the effects of both habitat fragmentation and habitat loss by conserving habitat where vulnerable species live and by maintaining big, interconnected reserves without barriers to dispersal of plants and animals.
In planning reserves, it is worth remembering the primary cause of the biodiversity crisis: habitat loss. It follows that the most important conservation measure is to increase the amount of habitat protected as wildlands. Protecting or restoring large areas of habitat counteracts the negative effects of habitat loss and fragmentation. Large reserves harbor larger populations of target species, averting decline into the extinction vortex (see chapter V.1). They also encompass more habitat types, each of which may harbor a different set of species that are protected in larger reserves. Different habitats protected in larger reserves also provide some animals with easier access to a diverse array of resources they need to complete their life cycle. Compared to the principle of protecting large reserves, all other principles make modest improvements to biodiversity conservation.
What area must be preserved to ensure preservation of all species? There is no set rule. At the minimum, to maintain population sizes at levels high enough to be out of reach of key extinction risk factors, populations should number in the thousands. And that is a minimum estimate; to accomplish more functional goals, such as preserving the functional role each species plays within its ecosystem, the numbers would need to be higher yet, and more evenly distributed across the range. For the largest and most wide-ranging species such as northern spotted owls and grizzly bears, conservation areas must number into the millions of hectares. Areas must then cover the ranges of the Earth’s millions of species. It takes only a few back-of-the-envelope calculations to show that the Earth’s biodiversity requires protection of a substantial portion of the biosphere.
Inevitably, the area needed to conserve all of earth’s biodiversity may outweigh the financial means of a large and growing human population. The ideal reserve network is limited, especially near cities or along coasts, where land values are high. (Conversely, vast areas are conserved on mountaintops, in deserts, and in tundra; see chapter V.4.) After attempting to conserve large areas, the other principles work within the limited areas available to conservation and with scarce conservation resources and are intended to maximize biodiversity by designing reserves in optimal configurations.
At the heart of reserve design is one key issue: funds to create reserves are limited, and land managers must make difficult choices about which areas to conserve. Should funds be invested to make reserves bigger or to restore lands that connect nearby reserves? The economic costs of reserve designs are easy to determine and involve the monetary costs of different land areas. The ecological benefits are more difficult to quantify because biodiversity is usually not monetized, and the relative benefits of alternative designs must be inferred from theory or limited observations.
The economic costs of reserves include land maintenance, purchase, and protection, and some reserve designs may carry higher costs than others. The cheapest strategy that has tended to be used most commonly is simply to conserve existing patches of vegetation. In some instances, it is more costly to restore areas that connect existing reserves than it is to expand reserves. Connecting reserves with corridors (discussed in detail below) may be more expensive because they are a specific shape and location, often passing through developed or agricultural lands that carry high values. Instead, funds could be spent to create larger reserves that are not connected. All other things being equal, managers will invest in the cheapest conservation alternative.
But the values of different reserve designs for biodiversity are likely not equal, and this is where more difficult decisions arise about ecological trade-offs. In the case of corridors versus larger reserves, ecological theories predict that connected reserves should maintain plant and animal populations and increase their diversity. An ongoing question in conservation biology is: To what extent do corridors increase population persistence and therefore preserve species? And what is that added value worth? Evaluating economic tradeoffs is made more difficult still because resources available to fund different designs may not be equal. Corridors may be more expensive, but the vision of connecting reserves with corridors may excite more donors and attract more funds. Corridors may also have other uses than biodiversity protection; for example, urban greenways may serve as recreational areas. These other factors create positive feedbacks that generate more resources for conservation. Finally, sometimes a trade-off does not exist: some ecosystems are so fragmented by human activities that creating large reserves is no longer possible, leaving corridors as the best available option.
Another intense debate that highlights the complexity of ecological trade-offs in reserve designs concerned whether to conserve a fixed habitat area in a single large reserve or several small reserves (called the “single large or several small”—SLOSS—debate). The original principle had been based solely on the role of larger habitat areas to reduce extinction risk. Yet, other ecologists rightly pointed out that one large, compact reserve may not encompass the diversity of habitats required by a large-variety species. In the end, there was no one right answer to the SLOSS debate, and decisions about how to partition fixed areas into reserves depend on the conservation context. But even in light of the complexity of trade-offs, there are still some principles of reserve design about which most ecologists would agree.
Many decisions about reserve design occur in landscapes that are already highly fragmented and heavily populated. In these landscapes, it is nearly impossible to create big reserves. The trade-offs that conservation biologists wrestle with are mainly those related to the biological benefits of alternative designs that are of the same area. In this context, an ecological watershed for reserve design principles came in the mid-1970s as an outgrowth of the theory of island biogeography (see chapter IV.2). Ecologists observed the parallel between islands in the ocean and habitat patches in a landscape of agriculture or development that is not used as habitat by most species. From these observations, scientists developed an initial set of principles to guide reserve design toward maximizing biodiversity conservation (plate 17). The principles were guided by two primary goals. The first was to create the largest reserves possible, reducing extinction risk. The second was to enhance dispersal, providing sources of new individuals to recolonize fragments in case of extinction and to promote genetic exchange. The initial principles were largely reinforced with the development of spatial theories in population ecology (see chapter II.4). The strong backing by theory and intuitive nature of the principles has inspired conservation planners, giving rise to the Y2Y and scores of other plans that create larger, well-connected reserves. The remainder of this chapter examines the key principles that attempt to increase the effective area of reserves.
In principle, reserves may be designed to preserve biodiversity in general. In practice, conservation targets are likely to be a handful of species of conservation interest—charismatic or rare species—or unique geographic or hydrologic features with which those species are associated. It is stating the obvious that areas should be protected that already contain populations that are targets of conservation. Such strategies must take into consideration that different areas may be used by species of conservation interest through the year. Some frogs and salamanders spend most of their life cycle in forests but breed in ponds; reserves must be designed to encompass both types of habitats and with pathways for individuals to move between them. One way to capture multiple habitat-specific rare species is to target geographic or hydrologic features where rare species abound (see chapter V.4). Once key areas are conserved, strategies for reserve designs vary in the degree to which degraded areas can be restored to enhance population viability.
Many reserves are designed with the assumptions that degraded habitat can be restored and that mobile animals can spread quickly to occupy restored areas. The Y2Y corridor will achieve conservation success when and if it protects expansive areas, crossing state and country borders, large enough to ensure the viability of grizzly bears, wolves, and caribou. Conservation targeted at species with large ranges may accomplish another goal of conserving biodiversity more generally. Protecting areas needed by large species may also protect the ranges of smaller ones, a strategy known as the umbrella species concept. Although applied widely, there is some debate about the utility of the umbrella species concept, especially because ranges of umbrella species do not always encompass the variety of habitats occupied by many smaller species. When conservation specifically targets smaller animals, the underpinnings of conservation remain the same as for large species: conserve an area large enough to encompass the home ranges of enough individuals to create viable populations that are protected from extinction. For mobile animals of any size, restoring corridors may promote dispersal and gene flow through the landscape.
When species have narrow habitat requirements, blocking off large and connected areas may not be the best strategy for conservation. Rare plants are often associated with very specific soil, moisture, or other environmental requirements. Where they occur, their populations can be quite dense, with thousands of individuals filling a hectare or less. These plants are less flexible in their use of nearby habitats that are conserved or restored, and distances between fragments may be too large for them to overcome via natural dispersal. It may be impossible to restore corridors with environmental characteristics needed for many plant species to spread. Thus, conservation for these kinds of plants often focuses specifically on protection of the highest-quality sites where they are found. Obviously, such species-specific plans target only the species we know well and miss all those species that have narrow requirements but are also poorly known.
Once reserve areas have been identified, their shape and setting may determine their effectiveness. Habitat loss and fragmentation introduce novel features into the landscape. Continuous natural habitats, once subdivided, run up against urban, agricultural, or other human-modified landscapes. These new landscape features are separated from conserved areas but can still influence protected areas from the outside. And outside influences are often strong, effectively reducing the area of a reserve for biodiversity conservation. Two interrelated landscape features are particularly important. First, and most immediate, is the boundary between conserved natural habitats and modified areas. These boundaries are often distinct and dramatic and give rise to changes in the abundances of individual plants or animals, the numbers of species, and other ecosystem attributes. Ecologists group all of these responses around habitat boundaries into the catchall phrase edge effects. The second feature depends on the type of modified habitat that is next to a conserved natural area, whether it be agriculture, buildings, or other degraded or natural habitats. Because this area is not the focus of conservation, it is referred to abstractly as the matrix. These concepts are strongly interrelated, as the type and extent of the matrix determine the types of species present at the edge and the degree to which edge effects penetrate natural areas. Later we will see how matrix habitats also influence landscape connectivity.
There has been a long history of considering positive and negative effects of edges in conservation and wildlife management. Aldo Leopold, a key figure in the early history of conservation biology, advocated the creation of edges to promote biodiversity. His views were not without merit. First, edges can provide new resources that are not available within either habitat forming them. This can happen when, for example, plants used as food by animals occur only at the edge. Second, they provide easy access to resources in the different habitats on either side of the edge; for example, adult butterflies may feed at flowers in fields, but their caterpillars may eat plants that grow in forest. For some wildlife species that are managed as game, such as white-tailed deer and bobwhite quail, high resource abundance near forest edges greatly increases their population sizes. Third, where landscapes were naturally fragmented, edges are essential in good reserve design. Many rare wetland flowering plants thrive at the boundary between bogs and upland forests. Taken together, these attributes make edges productive and diverse and help to explain why they might have some value in reserve design.
Yet we now know that early conservation biologists were mostly wrong about edges. Many imperiled species require large pristine areas of habitat. And just as there are some species that are attracted to edges, there are others that avoid them, and by large distances. Because of this avoidance, edge creation reduces the effective size of conserved habitats to sizes much smaller than the conserved area. The size can also be reduced by the presence of unwanted species that live on the edge or in the matrix. Edges create points of entry for unwanted species, including predator, invasive, or disease species, that are harmful to targets of conservation. A classic example of a harmful species that benefits from edges is the brown-headed cowbird. These birds live most of their lives in open fields but lay their eggs in the nests of other birds. Although not a forest species, brown-headed cowbirds will enter forests in search of host nests, especially when forest fragmentation increases the amount of edge and access to forest. When chicks hatch, unsuspecting mothers of forest birds feed cowbirds as their own, while brown-headed cowbird chicks work to push the mother’s chicks from her nest and outcompete remaining nestmates for food.
Edges create new environments that can disrupt native ecosystems. Where forests are exposed to wind or sun at edges, new and often weedy species thrive, and trees native to continuous forest can die. Because of the effects of edges on the presence of unwanted species and changes in climate conditions, many species of conservation concern settle far from the edge. In one tropical fragmentation experiment in Brazil, called the Biological Dynamics of Forest Fragments Project (part of which is seen in plate 17), researchers have found that most edge effects, such as on soil moisture, relative humidity, abundances of understory birds, and litterfall, occur within 100m of the edge. But some edge effects, such as on wind disturbance, tree mortality, and the composition of ant communities in the leaf litter, can extend nearly a half kilometer from the edge.
The severity of edge effects differs based on the types of matrix. Whether the habitat next to the edge is a regenerating forest, an agricultural field, or a subdivision can greatly affect what happens within conserved areas. The matrix determines the types and numbers of other species that may penetrate reserves and the degree to which matrix habitats buffer harsher environmental conditions. Often, the effect of matrix habitats depends on the degree of contrast between matrix and natural habitats. Urban areas that directly abut forest reserves create dramatic edges that can lead to more severe edge effects, whereas managed or regenerating forests cause less obvious effects of edges.
Many species of concern in conservation avoid edges, but others are attracted to edges. This can be problematic for species that are tricked into settling at edges created by humans that can mimic natural edges. In large forest areas, trees fall and create small and irregular edges that support abundant wildlife. Human-created edges may seem like these natural edges and attract animals. Yet, edges created by human landscape modification are usually quite different from natural edges, and environments there can be more risky. Indigo buntings are classic edge birds, and edges may provide excellent nesting habitats and abundant food resources for their chicks. One experiment showed that when forests are cleared by humans, buntings are attracted to areas with the highest amount of edge, but so are their predators. The end result is that chicks raised in edgier environments created by humans are less likely to fledge from nests. In this regard, human-induced habitat modification may create ecological traps that exacerbate population decline caused by habitat loss.
Conservation biologists generally recognize that edges reduce the conservation value of reserves, and the degree to which edges have negative effects is usually associated with the level of human impact in the matrix. But conservation biologists also recognize that edges can have positive effects in some circumstances when they are naturally part of the landscape, when they create new environments necessary for the maintenance of wildlife populations, or when they allow access to multiple resources that occur in each habitat. As a general rule, human-created edges should be minimized, but natural or seminatural edges between reserve and matrix habitat should be retained in reserves to improve habitat for native species.
A key goal of reserve design is to reconnect habitats that have been fragmented to allow the habitats to function as a whole rather than as a set of independent pieces. Reconnecting habitats increases genetic exchange, reducing the likelihood of inbreeding or genetic drift, and increases dispersal, aiding in the colonization of small sites where populations have become extinct. Connections also increase habitat area and may increase population sizes and biodiversity for that reason alone. However, if conservation aimed at increasing connectivity does not increase dispersal, then other strategies may add larger habitat areas for the same cost.
Corridors, or long and relatively thin strips of habitat, are the most direct way to increase landscape connectivity. By maintaining or restoring physical connections among patches, no edges or matrix must be crossed, and dispersal success should be highest. To be effective, corridors must include habitat that plants and animals would typically use during dispersal. Corridors often follow natural landscape features such as streams or mountain ridges. Connectivity can also be increased without physical connections simply by reducing the distance between reserves in the landscape. Reducing the distance between patches increases the likelihood that plants and animals can disperse between them. Lower travel distances decrease the amount of time plants or animals must spend in unsuitable, often risky matrix habitat. For species willing to leave their preferred habitat through matrix habitat, smaller reserves that form stepping-stones may provide a path connecting one larger patch to another.
For corridors and stepping-stones to be effective, increased connectivity must increase dispersal and gene flow. In once-continuous landscapes, it seems intuitive that reconnecting fragments should increase dispersal. This is often the case, and corridors have been shown to work for many species, including mountain lions, small mammals, birds, butterflies, and bird-dispersed plants. But physical connections will not always increase dispersal. At one extreme, increased connectivity may not benefit sedentary species if distances between reserves are too large or if corridors contain too much edge habitat. At the other, connectivity will not affect dispersal rates of highly mobile or generalist species, which fragmentation does not limit. To increase population persistence and biodiversity, connectivity must increase rare dispersal events to the point where they increase rates of colonization and reduce inbreeding and genetic drift. It is this high standard that justifies the strategy of increasing connectivity in reserve designs.
Deciding how to design landscapes that optimize connectivity can be tricky. The degree to which plants and animals will disperse through fragmented landscapes depend on characteristics such as the width, length, and degree of physical connection, which in turn depend on the quality of protected and matrix habitat. For example, some corridors (such as some urban greenways) may be so narrow that edge-avoiding species will not enter them. Alternatively, narrow corridors may enhance dispersal routes for species that move along edges. But edges may also provide habitat for predators to wait for unsuspecting dispersers. Usually, the optimal corridor width will depend on the distance at which edges affect movement behavior of animals or the distance to which negative edge effects penetrate corridors.
The optimal width of a corridor is related to its length. Animals and plants may be able to disperse through short corridors within hours or days, and easily within the lifespan of an individual. On the other hand, for gravitationally dispersed plants that disperse meters to tens of meters, it will take generations for them to pass through a kilometer-long corridor. Plants must disperse, establish, survive, and reproduce within these corridors. For these plants to succeed, habitat quality within the corridor must be very high. In landscapes where fragments are separated by long distances beyond the capacity of plants or animals to disperse through matrix, high-quality wide corridors are particularly important to increase connectivity.
The difference in quality between reserve and matrix habitat plays a key role in determining how landscape connectivity is designed. Some matrix habitats support species of conservation interest or serve as dispersal habitat. When the matrix is of moderate quality, then edge effects will be less severe, and corridors could be narrower. In such landscapes, stepping-stones, rather than corridors, can be used to promote connectivity. Other matrix habitats are extremely risky for plants and animals that exit reserves. Top predators such as grizzly bears and mountain lions often exit reserves, but when they do, they are likely to be killed by humans. Roads, which are increasingly fragmenting landscapes, can cause high levels of mortality. In cases where matrix habitat increases mortality risk, corridors are especially important to increase population viability.
Maintaining landscape connectivity is particularly important in a world with a changing climate. Species distributions are determined in large part by their physiological requirements, and for many species, the limiting environmental factor is temperature. However, species are also limited by their ability to disperse to those sites where their physiologies allow them to live. Higher global temperatures will shift suitable habitats to higher latitudes and higher elevations chapter V.6). Landscapes should be connected in anticipation of these future range shifts. To accommodate effects of a changing climate, landscapes will have to be connected at very large scales.
The Y2Y corridor exemplifies all of the key principles of reserve design. Its conservation targets are large predators and ungulates as well as a backbone infrastructure of established (and often prominent) reserves. It seeks to conserve the largest area possible within the region. Because of funding constraints, it targets conservation of lands that connect existing reserves, with wide corridors that reduce the effects of edges. And its scale makes it robust in the face of climate change. Y2Y has been successful in attracting investment to apply ecological theory. Time and research will tell if it is successful in conserving threatened populations and biodiversity within the ecoregion.
Beier, P., K. L. Penrod, C. Luke, W. D. Spencer, and C. Cabañero. 2006. South Coast missing linkages: Restoring connectivity to wildlands in the largest metropolitan area in the USA. In K. R. Crooks and M. A. Sanjayan, eds. Connectivity Conservation. Cambridge, UK: Cambridge University Press, 555–586.
Damschen, E. I., N. M. Haddad, J. L. Orrock, J. J. Tewksbury, and D. J. Levey. 2006. Corridors increase plant species richness at large scales. Science 313: 1284–1286.
Laurance, W. F., T. E. Lovejoy, H. L. Vaconcelos, E. M. Bruna, R. K. Didham, P. C. Stouffer, C. Gascon, R. O. Bierregaard, S. G. Laurance, and E. Sampaio. 2002. Ecosystem decay of Amazonian forest fragments: A 22-year investigation. Conservation Biology 16: 605–618.
Ricketts, T. 2001. The matrix matters: Effective isolation in fragmented landscapes. American Naturalist 158: 87–99.
Ries, L., R. J. Fletcher, Jr., J. Battin, and T. D. Sisk. 2004. Ecological responses to habitat edges: Mechanisms, models, and variability explained. Annual Review of Ecology and Systematics 35: 491–522.
Weldon, A. J., and N. M. Haddad. 2005. The effects of patch shape on indigo buntings: Evidence for an ecological trap. Ecology 86: 1422–1431.
Wilson, E. O., and E. O. Willis. 1975. Applied biogeography. In M. L. Cody and J. M. Diamond, eds. Ecology and Evolution of Communities. Cambridge, MA: The Belknap Press, 522–534.