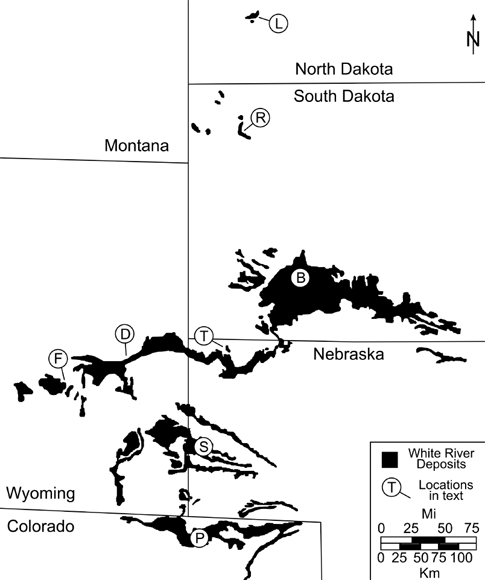
7.1. Map showing the distribution of the White River Group and correlative strata across the northern Great Plains. B = Badlands National Park, South Dakota; D = Douglas, Wyoming; F = Flagstaff Rim, southwest of Casper, Wyoming; L = Little Badlands of southwest North Dakota; P = Pawnee Buttes of northwest Colorado; R = Reva Gap, east of Buffalo, South Dakota; S = Scotts Bluff National Monument, Nebraska; T = Toadstool Geologic Park, north of Crawford, Nebraska.
The Eocene and Oligocene epochs are specific intervals of time recognized across the globe, but in order to understand their significance, it is important to understand how they are identified. The geologic history of our planet is stored within the archives of various types of rock bodies and strata, and the plant and animal remains preserved within them. Early scientists were aware that changes in plant and animal life had occurred, that the types of plants and animals preserved in a type of rock varied even if the rock type was the same, and that the existence of fossil remains in rock layers high on the tops of mountains suggested that the landscape had not always been as it appeared. Other than methods that allowed early geologists to establish the relative ages of various rock bodies, such as superposition, which states that in a sequence of horizontal strata the oldest layers of rock are on the bottom, these early scientists had no way to quantify the ages of strata and their associated fossils, when deposition started, how long it lasted, or when it stopped. Relative ages could be established across broad regions by tracing laterally continuous rock units or occurrences of particular fossils that allowed these early scientists to construct a relative geologic order of important paleontological and geological events, such as the appearance and extinction of the dinosaurs, but absolute ages for these events were unavailable. This changed with the development of radiometric dating of geologic materials and the discovery of magnetic signatures preserved in rock. The combination of radiometric dating plus paleomagnetism in sedimentary rock strata around the globe has allowed the establishment of a chronologic framework within which the patterns of evolution and geologic events portrayed in the geologic column can be placed.
The Eocene–Oligocene boundary represents the most dramatic change in global paleoclimatic conditions since the extinction of the dinosaurs. This change, also referred to as the Hothouse to Icehouse Transition (Prothero, 1994; Prothero, Ivany, and Nesbitt, 2003; Koeberl and Montanari, 2009), is associated with dramatic changes in global temperature, large drops in sea level, the establishment of permanent glacial ice on Antarctica, and extinctions in the nonmarine and marine fossil record. The causes of this dramatic climate change are still a matter of debate. Hypotheses to explain this climate change have included plate tectonic reorganization and opening of marine passageways between South America, Australia, and Antarctica; uplift of the Himalayan Mountain Range; and multiple extraterrestrial impact events during the late Eocene. Any attempt to understand the causes of this transition must take into account both terrestrial and marine records across the globe during this period of time.
Our particular interval of interest, the Eocene–Oligocene boundary, is one of several divisions of epochs within the Tertiary Period of the Cenozoic Era. For each of these divisions within the geologic column, one particular place on the globe is used as the global reference against which all other locations of the same age can be compared. These scientifically agreed-upon stratigraphic levels and locations are referred to as a Global Stratotype Section and Point (GSSP) and are formalized for many of the boundaries in the geologic column (Alvarez, Claeys, and Montanari, 2009). For the Eocene–Oligocene boundary, the global standard is a section of shallow marine limestones exposed along the Adriatic coast near Massignano, Italy, which contains several layers of impact ejecta, volcanic ashes, evidence of magnetic reversals, and abundant marine fossils (Premoli and Jenkins, 1993; Hyland et al., 2009). The Eocene–Oligocene boundary is currently dated at approximately 33.8 Ma and is associated with a particular set of normal and reversed paleomagnetic signatures referred to as 13N (normal) and 13R (reversed) polarity when compared to today (Hilgen and Kuiper, 2009). The numbers for polarity events increase with older rocks, counting backward from 1N/R for our modern record (Fig. 2.2).
Within the various epochs of the Tertiary such as the Eocene and Oligocene, smaller units of time can be identified. For the terrestrial record on a specific continent, these subdivisions are based on the mammalian fossil record. Mammals are used because they are abundantly preserved as fossils in most places, many lineages have a relatively rapid evolutionary rate, and their teeth are anatomically complex enough to allow recognition of different species. These are biostratigraphic units because they are based on living organisms and for North America are called the North American land mammal ages (NALMAs). As with any biostratigraphic unit, it is defined by a detailed knowledge of the stratigraphic ranges of the fossil taxa used to characterize the time interval. The boundaries of a NALMA may be recognized by the first appearance of a taxon in the fossil record, whether because it has dispersed into North America from another continent or evolved from an earlier ancestor in North America. It may also be defined by the absence of a genus or species that was previously present on the continent but becomes extinct. Not all taxa are equally useful as biochronologic markers. This is because some have a long geological record and so cannot be used to identify a small unit of time. Other taxa are not useful because they have a limited distribution, so the taxa cannot be identified over wide geographic regions. Taxa that have undergone relatively rapid evolutionary change, that have distinct morphologies in their dentition (the favored part of the body used by paleomammalogists) or the skeleton, and that have a widespread distribution are the most useful in terms of defining and refining land mammal ages.
Four North American land mammal ages are present in the Big Badlands: Chadronian, Orellan, Whitneyan, and the earliest Arikareean. They span the latest Eocene to early Oligocene, approximately from 37.1 to 27.7 Ma (Fig. 2.2). These land mammal ages were originally established by the Wood Committee (Wood et al., 1941), and each takes its name from the geographic locality from which a fauna with a distinctive combination of taxa was collected. The late Eocene Chadronian land mammal age is based on fauna recovered from the Chadron Formation near its type area in the vicinity of Chadron, in northwestern Nebraska and in southwestern South Dakota. These deposits are referred to as the Titanotherium beds in the older literature. This land mammal age was originally defined as the time when brontotheres and Mesohippus overlapped their geologic ranges (Wood et al., 1941). However, brontotheres and Mesohippus now are known to overlap in the older Duchesnean land mammal age (MacFadden, 1998), and the uppermost occurrence of brontotheres may extend into the earliest Orellan land mammal age (Prothero and Emry, 1996, 2004). The taxa that mark the start of the Chadronian land mammal age include the first occurrence of the oreodonts Bathygenys spp. and Limnenetes spp., the amphicyonid Brachyrhynchocyon dodgei, and the entelodont Archaeotherium coarctatum (Prothero and Emry, 2004). The end of the Chadronian is currently marked by the transition between the Chadronian Leptomeryx speciosus to the Orellan Leptomeryx evansi. The last occurrences of several mammal species also occur at the Chadronian–Orellan boundary, including the last occurrence of the entelodont Archaeotherium coarctatum, the horse Mesohippus westoni, the oreodont Miniochoerus chadronensis, and the rabbit Palaeolagus temnodon (for a complete listing, see Prothero and Emry, 2004). Brontotheres, the horse Miohippus grandis, and the camel Poebrotherium eximium have their highest occurrences just above the Chadronian–Orellan boundary (Prothero and Emry, 1996, 2004). Radiometric dates for sediments containing Chadronian faunas place its beginning at 37.1 Ma and ending at 33.4 Ma (Fig. 2.2).
The first NALMA in the Oligocene is the Orellan, originally based on the fauna from the Orella Member of the Brule Formation (Wood et al., 1941). This land mammal age takes its name from a type locality near the town of Orella in northwestern Nebraska but includes southwestern South Dakota and eastern Wyoming as well. The term replaced the older names “turtle–Oreodon beds” or “Oreodon beds” used for rocks of this age. Many of the taxa used to define the Orellan land mammal age by Wood et al. (1941) can no longer be used because of subsequent taxonomic revisions. While Eporeodon, Eumys, and Ischyromys were identified as first appearing in the Orellan, Eumys and Palaeolagus haydeni have been questionably identified in the upper Chadron Formation. Currently the base of the Orellan is marked by the first occurrence of Leptomeryx evansi (Prothero and Emry, 2004). Hypertragulus calcaratus first appears at the start of the Orellan within the White River Group, although outside the White River Group Hypertragulus has been reported from the Chadronian of Mexico (Ferrusquia-Villafranca, 1969). A large number of taxa have last occurrences at the end of the Orellan land mammal age. These include the hyaenodont Hyaenodon crucians, the rabbits Palaeolagus intermedius and all species of Megalagus, the entelodont Archaeotherium mortoni, the oreodont Miniochoerus gracilis, the rhinoceros Subhyracodon occidentalis, the small artiodactyl Hypertragulus calcaratus, and all the species of the horse Mesohippus. In addition to the large rodent Ischyromys typus, 12 genera of rodents may have become extinct at the end of the Orellan (for a complete listing, see Prothero and Emry, 2004). Radiometric dating places the beginning of the Orellan at 33.9 Ma and ending at 32.4 Ma (Fig. 2.2).
The Whitneyan land mammal age is based on the fauna from the Whitney Member of the Brule Formation (Wood et al., 1941). The type locality is near the town of Whitney in northwestern Nebraska, but this member extends into southwestern South Dakota and eastern Wyoming. The Whitneyan replaced the older term “Protoceras–Leptauchenia beds” for faunas from this time interval. While Protoceras is restricted to this time interval, its usefulness as a biostratigraphic indicator is limited because it is restricted to a limited depositional setting, the Protoceras channels. The traditional start of the Whitneyan land mammal age is the first occurrence of abundant Leptauchenia oreodonts. Leptauchenia major does have a first occurrence at the start of the Whitneyan, but Leptauchenia decora that is abundant in the basal Whitneyan rocks has its first occurrence in Orellan rocks. Other first occurrences at the start of the Whitneyan land mammal age include first species of the nimravids Nimravus and Eusmilus, the rhinoceros species Hyracodon leidyanus and Diceratherium tridactylum, the oreodont Miniochoerus starkensis, the camels Paratylopus primaevus and Paralabis cedrensis, the tapir Protapirus obliquidens, and the beaver Agnotocastor praetereadens. The end of the Whitneyan land mammal age is marked by the last occurrences of the tapir Protapirus obliquidens, the rabbit Palaeolagus burkei, all species of the camels Paratylopus and Paralabis, and the nimravids Dinictis and Hoplophoneus (for a complete list, see Prothero and Emry, 2004). Radiometric dates places the beginning of the Whitneyan at 32.4 Ma and ending at 30.0 Ma (Fig. 2.2).
Originally the Arikareean land mammal age was based on faunas found in the Arikaree Group of western Nebraska, and the fauna preserved at Agate Fossil Beds National Monument was considered to be the most typical of the time period (Wood et al., 1941). Recent research has greatly modified the definition of the Arikareean, and it is now the longest of the NALMAs, beginning at 30.0 Ma and ending at 18.6 Ma, and transgresses the Oligocene–Miocene boundary (Fig. 2.2). This land mammal age is defined by the first appearance of a number of Eurasian immigrants including the Talpinae (talpid moles) and the rodents Plesiosminthus, Leidymys, and Allomys. Other first appearances in this land mammal age include the oreodonts Desmatochoerus megalodon, Promerycochoerus spp., and Sespia nitida, the camels Miotylopus and Nanotragulus, and the dog Sunkahetanka geringensis. Taxa whose last occurrence is in the early early Arikareean include the primate (or dermopteran) Ekgmowechashala, the rodent Eumys, the creodont Hyaenodon, the nimravid cat Nimravus, and the rhinoceros genera Hyracodon and Subhyracodon.
The White River sequence across the northern Great Plains preserves the best terrestrial record of this Eocene–Oligocene climate transition anywhere in the world. The term “sequence” is used herein, instead of “group,” to recognize the different stratigraphic rankings of these rock units across the region. Understanding the spatiotemporal record of this paleoclimatic change within the terrestrial deposits of the White River sequence provides important information to establish linkages with the rate and timing of paleoclimatic changes in ancient marine environments. In turn, this can provide insights into ancient global climate dynamics, which can be used to understand our current episode of global climate change.
Although Badlands National Park is the best-known part of the White River sequence, these deposits stretch for hundreds of miles across the northern Great Plains (Fig. 7.1). From west to east and north to south, these deposits vary in their sedimentary composition and age. Some locations preserve an almost complete record of the Eocene–Oligocene transition (EOT); others are condensed or missing large spans of time. The reasons for this wide disparity in composition and preservation include proximity to sediment sources and tectonic influences on basin accommodation, or later erosion of parts of local sequences. In general, the White River sequence is more completely preserved in northwestern Nebraska and east-central Wyoming; it is less completely preserved toward the north in northwestern South Dakota and southwestern North Dakota. When put in a temporal framework, these widespread deposits provide a level of detail and insight into the geologic history of the White River sequence that would be unattainable by looking at any one location.
The best-known exposures of the White River sequence in northwest Nebraska can be found approximately 20 km north-northwest of the town of Crawford at Toadstool Geologic Park (Fig. 7.1). This exposure of badlands, while extremely small when compared to the Big Badlands of South Dakota, preserves a key interval of strata not found in Badlands National Park. In essence, Toadstool Geologic Park is a Rosetta stone for the White River sequence: it ties together the deposits of the White River sequence in southwest South Dakota to those in east Central Wyoming.
The stratigraphic nomenclature of the Nebraska Badlands has undergone significant revision over the recent past. Before the work of LaGarry (1998), Terry (1998), and Terry and LaGarry (1998), the hierarchy of Schultz and Stout (1955) was used to categorize the stratigraphy of the Toadstool Geologic Park region. This hierarchy was based on subdivisions of the White River sequence into distinct lettered intervals that corresponded to a combination of changes in rock type, fossil content, and marker beds (volcanic ashes and paleosol horizons). These lettered subdivisions were then interpreted as direct correlations to rock units in the Big Badlands of South Dakota. The stratigraphic revisions of LaGarry (1998), Terry (1998), and Terry and LaGarry (1998), which were based only on changes in lithology in the Toadstool Geologic Park region, resulted in dramatically different correlations to deposits in the Big Badlands of South Dakota. These revisions and correlations are followed herein, with references made to the hierarchy of Schultz and Stout (1955) as needed for clarification.
The base of the badlands stratigraphic sequence around Toadstool Geologic Park is similar to that already established in Badlands National Park. After the retreat of the Western Interior Seaway at the end of the Cretaceous, exposure, weathering, and ancient soil formation modified the marine Pierre Shale into bright hues of red, yellow, and purple (Plates 2, 14). This episode of weathering and soil formation persisted for approximately 30 million years to produce paleosols similar to those of the Yellow Mounds Paleosol Series in South Dakota before the first phases of fluvial deposition began. Just as in South Dakota, these first pulses of late Eocene fluvial sedimentation were also pedogenically modified into bright red paleosols similar to those described in the Chamberlain Pass Formation (Terry and Evans, 1994) that laterally correlate to bright white bodies of channel sandstone up to 7 m thick (Chadron A of Schultz and Stout, 1955). Pedogenic modification of the Chamberlain Pass Formation in northwest Nebraska was not as intense as that in South Dakota, which resulted in the preservation of rare vertebrate fossils of brontotheres and tortoises (Vondra, 1956; LaGarry, LaGarry, and Terry, 1996) in channel sandstones.
The Chamberlain Pass Formation in northwest Nebraska is overlain by the Peanut Peak Member of the Chadron Formation (Chadron B of Schultz and Stout, 1955), which is exposed as the same greenish-gray, smectite-rich, haystack-shaped outcrops seen in the Big Badlands. The Peanut Peak Member is in turn overlain by the Big Cottonwood Creek Member (Chadron C and Orella A of Schultz and Stout, 1955), a unit of the Chadron Formation not seen in Badlands National Park. The Big Cottonwood Creek Member has a greater silt content than the underlying Peanut Peak Member; it is recognized by a change in erosional relief to more cliff-forming exposures (Plate 14).
Just as in South Dakota, the Chadron Formation is overlain by the Brule Formation in northwest Nebraska, but the names of individual members differ, as do lithologies. The Orella Member of the Brule Formation (Orella B–D of Schultz and Stout, 1955) directly overlies the Chadron Formation and is recognized by a change to more silty sediments, multiple thin sheets of sandstone, and an overall browner coloration (Plate 14). The Orella Member is also noted for downcutting of ancient river channels into underlying strata by up to 10 m. This downcutting, referred to as the Toadstool Park Channel Complex (Terry and LaGarry, 1998), is easily seen at Toadstool Geologic Park as thick units of cross-bedded sandstone that incised the Chadron Formation (Plate 14) to form a prominent paleovalley. This paleovalley was subsequently backfilled with additional sediments of the Orella Member. The lower part of the Orella Member is most similar to the Scenic Member of the Brule Formation in the Big Badlands, whereas the upper part is similar to the Poleslide Member.
The Orella Member is overlain by the Whitney Member in northwest Nebraska (Whitney A–C of Schultz and Stout, 1955). It is recognized by a change from brownish-beige fluvial sheet sandstones and overbank mudstones to pinkish, volcaniclastic eolian siltstone with abundant potato-ball concretions of calcium carbonate (Plate 14). The Whitney Member has no direct lithologic counterpart in the Big Badlands of South Dakota. The Whitney Member is in turn capped by the informally designated, beige Brown Siltstone Member of the Brule Formation (LaGarry, 1998) and is recognized by a change to a greater concentration of volcanic ash. The Brown Siltstone Member, which is capped by coarse cobbles of the Arikaree Group, are considered to be equivalent to the lower Sharps Formation of southwest South Dakota (J. Swinehart, pers. comm. to E. Evanoff, 2010).
With a lithostratigraphic framework in place between Toadstool Geologic Park of northwest Nebraska and the Big Badlands of South Dakota, geologic dating of these deposits, either through correlation of paleomagnetic zonations, radioisotopic dating of discrete volcanic ash beds, and to a lesser extent biochronology, provides a spatiotemporal model of deposition for the White River sequence across this region (Fig. 7.1). On the basis of thickness alone, the White River sequence is more complete at Toadstool Geologic Park compared to the Big Badlands of South Dakota, which in turn allows for a more detailed temporal model using the exposures at Toadstool Geologic Park as a reference.
The Chamberlain Pass and Chadron formations of Nebraska were deposited during the late Eocene through the early Oligocene based on magnetostratigraphy and radioisotopic dating of several volcanic ashes within the Big Cottonwood Creek Member of the Chadron Formation (Prothero and Swisher, 1992; Prothero, 1996; Sahy et al., 2012). The time represented by the deposition of the Big Cottonwood Creek Member in Nebraska is not represented by deposition in the Big Badlands but is instead manifested as an erosional unconformity of up to 1.05 million years, which created paleotopography on top of the Peanut Peak Member of the Chadron Formation before burial by the Scenic Member of the Brule Formation (Evanoff in Benton et al., 2007).
The Eocene–Oligocene boundary occurs within the upper 10 m of the Big Cottonwood Creek Member, approximately 2 to 4 m above the Upper Purplish White Layer, a volcanic ash dated at 33.907 ± 0.032 Ma (Sahy et al., 2010). In the Big Badlands of South Dakota, the Eocene–Oligocene boundary is lost within the 1.05 my unconformity at the Chadron–Brule contact. Biochronological data suggest that the Chadron Formation is primarily Chadronian, with the exception of the upper 6 to 8 m of the Big Cottonwood Creek Member, which is Orellan.
The Brule Formation of northwest Nebraska was deposited during the early Oligocene based on magnetostratigraphy and radioisotopic dating of several volcanic ashes within the Whitney Member (Prothero and Swisher, 1992; Prothero, 1996; Sahy et al., 2012). Although the Orella Member contains two distinct ash beds (the Horus and Serendipity ash beds) within the Toadstool Park channel complex paleovalley backfill sequence (Kennedy, 2011), analysis of these ashes has yet to provide a reliable radiometric date. When compared to the Badlands of South Dakota, the timing of deposition for the Orella Member is similar to the Scenic Member of the Brule Formation, but precise tie points are elusive. Evanoff (in Benton et al., 2007) reports the presence of a monazite-bearing volcanic ash within the lower Scenic Member, but no radiometric dates have been produced. Terry (unpubl. data) has noted distinctive pockets of bentonite (altered volcanic ash) within the Scenic Member along BIA 2 just west of the White River Visitor Center, and southwest of Sheep Mountain Table within the Indian Creek drainage (Fig. P.1). Whether or not these bentonites correlate with previously recognized ash beds is unknown. Biochronological data for the Orella and Scenic members are identical (both are Orellan); they provide no additional refinement to ages.
The Whitney Member of the Brule Formation in Nebraska is temporally equivalent, on the basis of magnetostratigraphy, to the Poleslide Member in the Big Badlands of South Dakota, but not lithologically. The upper part of the Orella Member in Nebraska is a better lithologic match to the Poleslide Member of South Dakota. In Nebraska, the Orella and Whitney members are transitional across their boundary, suggesting a gradual change from one depositional regime to another. In the Big Badlands, the Scenic and Poleslide members are separated by a recently recognized thin, ashy layer, the Cactus Flat marker bed (Stinchcomb, 2007; Stinchcomb, Terry, and Mintz, 2007). Although no attempts have been made to obtain radiometric dates, its stratigraphic position and magnetostratigraphic signature suggest that it matches the Lower Whitney Ash of Nebraska at 31.85 ± 0.02 Ma. Biochronologic data for these units suggest a Whitneyan age but provide no additional refinement.
The uppermost member of the Brule Formation in Nebraska is the informally designated Brown Siltstone Member. The Brown Siltstone has up to three recognized ash beds in its lower part, the Nonpareil Ash Zone of Swinehart et al. (1985), which collectively yield a date of 30.05 ± 0.19 Ma. The most likely ash equivalent to the Nonpareil Ash Zone is the Rockyford Ash in the Big Badlands. The White River sequence in northwest Nebraska is uniformly overlain by coarse conglomerates of the Arikaree Group.
The disparity in direct lithologic correlations for some units between Toadstool Geologic Park and the Big Badlands, in addition to variations in timing of deposition across the region (such as the Peanut Peak Member), can be attributed to several processes. Simply put, deposition is not instantaneous across an entire region. Climates and environments are diachronous – that is, they migrate through space and time. For example, modern-day reductions in Arctic sea ice volume are attributed to increases in global temperature, a trend that has been active since the last major continental glaciers in North America began their retreat approximately 20,000 thousand years ago. Areas once dominated by periglacial features 20,000 years ago are now dominated by prairie and forest ecosystems through the Midwest United States. Deserts do not deposit sand across an entire region at once. Eolian environments migrate and can become repetitively stabilized and reactivated depending on changes in climate conditions and the activity of stabilizing vegetation, like the Sandhills of Nebraska. Likewise, the fluvial and eolian environments that were responsible for deposition of the Chadron and Brule formations of Nebraska and South Dakota generated sedimentary deposits that accumulated across large areas over time, but deposition across the region was diachronous.
In some instances, deposition of a particular lithologic unit did not extend into the Big Badlands. All rock units must taper and end laterally because the particular environment responsible for their deposition eventually gives way to a new environmental setting. This seems to be the case with the Big Cottonwood Creek, Whitney, and Brown Siltstone members of the Chadron and Brule formations in Nebraska. Direct lithologic equivalents of these deposits have yet to be formally recognized in the North Unit of Badlands National Park, but units similar to these are present within the South Unit of the park and can be seen on the southwest corner of Cuny Table near the intersection of BIA Highways 2 and 41. At this location, a 3 to 5 m interval of strata similar to the Big Cottonwood Creek Member rests on the Peanut Peak Member of the Chadron Formation and is capped in turn by an interval of strata tens of meters thick that is similar to the Orella Member, followed by a thicker interval of strata similar to the Whitney Member in color, erosional relief, and concentration of potato-ball concretions. This suggests that at least some of these lithologic units are wedge shaped and taper to the north and east from Nebraska into South Dakota, although detailed measurements are needed for better characterization of these units. The most critical observation from measurement of these units is that the physical rock record of the EOT from hothouse to icehouse conditions is better preserved in northwest Nebraska and east central Wyoming than in the Big Badlands of South Dakota as a result of 1.05 million years of missing rock record between the Chadron and Brule formations in Badlands National Park.
When taken as a whole, the paleoclimatic data from the White River sequence suggest an overall trend of cooling and drying. This trend was not a sharp event at the EOT proper but was the result of gradual changes that began in the late Eocene and carried over across the Eocene–Oligocene boundary to culminate in a dramatic change in paleoclimates and paleoenvironments of the early Oligocene. The clues to this overall change from warmer and wetter conditions of the late Eocene to cooler and drier conditions of the early Oligocene is based on the analysis of the types of sediment that were deposited and the ancient environments that they represent, a change in the types of fossil soils, and changes in the types of animals that survived into the Oligocene (Plates 15, 16).
Sediments in Badlands National Park change from clay-dominated, greenish-blue haystack mounds of the late Eocene Chadron Formation to the progressively more cliff-forming mudstone units of the early Oligocene Brule Formation (Plate 4). The clays of the Chadron Formation represent floodplain muds that were deposited as rivers overflowed their banks, burying former landscapes and preserving bones of animals that were either caught up in the flooding or that had accumulated on the ancient landscape as a result of predation. The rivers that deposited these muds were similar to the meandering rivers seen today in the eastern half of the United States. Rivers such as these are characterized by an abundance of water, with finer sediments transported in suspension and coarser sand and gravel transported as bed load at the bottom of the river channel. The type of clay that was deposited on the floodplains (smectite) is derived from the chemical weathering of volcanic ash, which we know existed in this region in great abundance because we see occasional layers of volcanic ash from large eruptions, as well as shards of ash in rock samples. The particular type of weathering that creates smectite from volcanic ash is termed hydrolysis, which represents the alteration of ash by water. As hydrolysis proceeds, additional atoms are stripped from the smectite to produce kaolinite clay. Kaolinite is abundant in the Chamberlain Pass Formation at the base of the Badlands (Plate 2). Hydrolysis is favored under humid climate conditions.
As we move closer in time to the Eocene–Oligocene boundary, the proportion of smectite clay to volcanic glass changes in favor of the glass shards. Although smectite and kaolinite are still present in the rocks of the Badlands, they are becoming progressively less abundant. As this change to progressively greater concentrations of silt-size ash occurs, the rivers of the Badlands change from deeper gravel- and sand-rich channels of the Chamberlain Pass and Chadron formations to channels with finer grain size, sedimentary structures, and bedding suggestive of drier, and possibly ephemeral, conditions. The amount of available water appears to be decreasing across the Eocene–Oligocene boundary, which results in a decrease in the amounts of hydrolysis, and in turn less smectite and kaolinite. By the time we reach the Poleslide Member of the Brule Formation in the Big Badlands, we see a strong influence of wind-dominated deposition. Windblown volcanic dust deposits steadily accumulate but are interrupted by periods of landscape stability represented by greater concentrations of fossil remains and well-expressed paleosols (Figs. 3.13, 3.14).
The paleosols dramatically change leading up to and across the Eocene–Oligocene boundary in Badlands National Park, and at Toadstool Geologic Park in northwest Nebraska. Paleosols of the Chamberlain Pass and Chadron formations are similar to modern soils that would be found in forests today (Plates 6, 7, 15). Fossil roots are suggestive of treelike vegetation, and occasional petrified logs and wood fragments have been found in the Chamberlain Pass Formation (Fig. 5.2). Fossil roots in paleosols of the early Oligocene Brule Formation are dramatically smaller and suggest an open rangeland with occasional trees (Plate 16). Higher up in the Brule Formation, roots are commonly no larger in diameter than a pencil, with most much smaller. Paleosols in the upper part of the Brule Formation are similar to those found on modern prairies (Retallack, 1983b; Mintz, 2007; Stinchcomb, 2007).
As for fossil vertebrates, the most dramatic change is the extinction of the large brontotheres at the end of the Eocene. Brontotheres were browsers, according to their tooth structure, and they probably inhabited a forest environment and likely could not adapt to the progressively open landscapes of the Oligocene. An extinction of many types of reptiles is also associated with the EOT and suggests a change to progressively cooler and drier conditions (Hutchison, 1992). This is also supported by a change in fossil land snails from the more humid-adapted varieties of the Eocene to forms indicative of the drier and cooler environments of the Oligocene (Evanoff, 1990b; Evanoff, Prothero, and Lander, 1992).
Although all of these indicators suggest a change toward progressively cooler and drier conditions from the Eocene to Oligocene, not all data sets agree. Isotopic data from fossil bones at Toadstool Geologic Park, Nebraska, suggest that temperatures dropped by as much as 8°C across the Eocene–Oligocene boundary, but that the amount of available moisture stayed relatively constant (Zanazzi, Kohn, and Terry, 2009). In contrast, Boardman and Secord (2013) analyzed isotopes from fossil tooth enamel that record diet and water intake and concluded that precipitation decreased slightly across the EOT in Nebraska but that temperatures remained relatively unchanged. It is also possible that the changes we see in the rock record are nothing more than an environmental response to an increase in the rate at which sediment was being introduced into the region. The Oligocene experienced sedimentation rates almost double that of the late Eocene in this region (Zanazzi et al., 2007), suggesting a greater amount of sediment input. This pattern is also seen in Badlands National Park (Terry, 2010). Increased sedimentation could produce the changes seen in the change from river- to wind-dominated environments and associated paleosols of the Brule Formation.
Further to the east in the Wyoming badlands, strata become thicker, but some parts of the rock record that were preserved in Nebraska are absent. As in Nebraska, the White River sequence rests on Cretaceous strata. The zone of intense pedogenic alteration at the base of the sequence similar to the Yellow Mounds Paleosol Series is present in the far eastern part of the state between Lusk, Wyoming, and the Nebraska–Wyoming border, but is not visible toward Douglas, Wyoming (Terry and LaGarry, 1998). Fluvial units similar to the Chamberlain Pass Formation are also present between Lusk and the state line, but also are not visible further to the west (Terry and LaGarry, 1998). Units similar to the Peanut Peak and Big Cottonwood Creek members of the Chadron Formation are present, as is the Orella Member of the Brule Formation.
One of the most striking differences between the Nebraska and Wyoming exposures is the dramatic increase in the number of distinct volcanic ash beds that are preserved in Wyoming, which is the result of the western volcanic sources responsible for the voluminous amounts of volcanic ash in the White River sequence (Larson and Evanoff, 1998). These numerous ashes have provided key radiometric dates that have enhanced our ability to correlate the deposits, magnetozones, and biochrons of the White River sequence across various exposures in Wyoming and to the east in Nebraska (Evanoff, Prothero, and Lander, 1992; Prothero and Swisher, 1992; Prothero, 1996; Larson and Evanoff, 1998).
The most detailed studies of the White River sequence in Wyoming have been carried out on deposits preserved in paleovalleys (Evanoff, 1990a), such as in and around the town of Douglas (Fig. 7.1). Similar to the deposits of northwestern Nebraska, the exposures at Douglas reflect a change upward from fluvial-dominated deposits during the Eocene within units similar to the Peanut Peak and Big Cottonwood Creek members to progressively more eolian conditions during the Oligocene within strata similar to the Orella Member of the Brule Formation. The White River sequence at Douglas is unconformably capped by conglomeratic strata similar to the Arikaree Group. Vertebrate fossil assemblages at Douglas mirror those in Nebraska, with Chadronian fossils in units similar to the Peanut Peak and Big Cottonwood Creek members of the Chadron Formation and fossils of the Orellan within strata similar to the Orella Member of the Brule Formation. The Eocene–Oligocene boundary in the Douglas area falls within the uppermost part of strata similar to the Big Cottonwood Creek Member at approximately the same level as a volcanic ash, referred to as the 5 tuff, that has been dated to 33.59 ± 0.02 Ma to 33.91 ± 0.06 Ma (Evanoff, Prothero, and Lander, 1992; Prothero and Swisher, 1992).
Further to the west and south of Douglas, the White River sequence is preserved within additional paleovalleys that were cut into underlying Mesozoic, Paleozoic, and Archean bedrock (Evanoff, 1990a). The most detailed studies have been carried out at Flagstaff Rim, approximately 30 km southwest of Casper, which has been developed for extraction of petroleum resources from strata underlying the White River sequence. The base of the White River sequence at Flagstaff Rim is marked by large boulders and cobbles that are overlain by strata similar to the Peanut Peak Member of the Chadron Formation farther to the east (Emry, 1973), but at Flagstaff Rim, this interval is intermixed with vibrant red mudstone and white channel sandstone (Plate 14). This is overlain by cliff-forming greenish-gray strata similar to the Big Cottonwood Creek Member, but enriched in volcanic ash compared to strata further to the east. Numerous volcanic ash beds are also preserved and help to date the rock strata and associated vertebrate fossils (Emry, 1973; Prothero and Swisher, 1992; Sahy et al., 2012). Although this is a thick section of strata, the amount of time preserved is limited to only the late Eocene. These strata are unconformably overlain by conglomeratic units of the Split Rock Formation of late Oligocene and possibly early Miocene age (Lillegraven, 1993).
From a sedimentological point of view, the strata at Flagstaff Rim follow the same pattern seen in Badlands National Park: clay-rich units at the base of the section become increasingly dominated by silt-size volcanic ash up section. The big difference is that this change occurs totally within the late Eocene. Just as in the Badlands of South Dakota and Nebraska, preliminary data from paleosols at Flagstaff Rim suggest that climatic conditions were becoming progressively drier and that temperature was decreasing (Griffis, 2010; Griffis and Terry, 2010). The vertebrate fossil record from Flagstaff Rim is extensive (Emry, 1973) but is of limited utility in deciphering paleoclimatic conditions.
To the south of Toadstool Geologic Park, the White River sequence is exposed at Scotts Bluff National Monument in west central Nebraska (Fig. 7.1). The exposures are similar to the Brule Formation, and dated volcanic ash beds place the majority of the strata and vertebrate fossils in the Oligocene (Prothero and Swisher, 1992). The Eocene–Oligocene boundary is not exposed at Scotts Bluff National Monument. In northeast Colorado, strata of the White River sequence are preserved in isolated outcrops and buttes, the most notable of which are the Pawnee Buttes. These buttes rest on the same zone of intense weathering seen at the base of the Badlands in South Dakota, but the strata that comprise the buttes are relatively similar to those seen at Scotts Bluff National Monument.
North of Badlands National Park, the strata of the White River sequence thin and disappear just beyond the town of Wall before appearing again in isolated patches and buttes in the northwest corner of South Dakota near Buffalo (Fig. 7.1). Outcrops are thin compared to the Big Badlands, and they rest on a variety of Paleocene and Eocene strata. The base of the White River sequence is marked in places by white channel sandstones similar to the Chamberlain Pass Formation, referred to as the Chalky Buttes Member of the Chadron Formation in North Dakota (Hoganson, Murphy, and Forsman, 1998). The Chalky Buttes Member is overlain by the South Heart Member of the Chadron Formation, which is identical to the Peanut Peak Member seen in the Big Badlands (Plate 14). The Chadron Formation is in turn overlain by the undifferentiated Brule Formation. The exact position of the Eocene–Oligocene boundary is uncertain, but it likely falls near the contact between the Chadron and Brule formations, according to magnetostratigraphy and biochronology (Prothero, 1996; Hoganson, Murphy, and Forsman, 1998). A volcanic ash referred to as the Ash Creek tuff is described from the Brule Formation in North Dakota (Hoganson, Murphy, and Forsman, 1998), but it has yet to be correlated with any of the ashes found elsewhere in the White River sequence. As in the Big Badlands of South Dakota, rock units across the Eocene–Oligocene boundary in North Dakota change from claystone-dominated units in the Eocene to silt-dominated units in the Oligocene. The paleosols of this region have yet to be investigated.
Although the strata of the White River sequence across the northern Great Plains represent the best-preserved nonmarine archive of the EOT, other sites across the globe, both marine and nonmarine, provide important regional-scale data that can be used to derive an overall synthesis of this critical time in Earth’s history. Our comparisons of ancient paleoclimatic conditions are, by default, referenced to our modern-day climate conditions across the globe. Just like today, climatic and environmental conditions during the Eocene and Oligocene varied across the globe.
In North America, the other notable records important to our understanding of the of the terrestrial EOT are preserved in isolated pockets and buttes of strata and associated fossil assemblages extending from Saskatchewan to New Mexico, and Oregon to California (Prothero, 1994; Prothero and Emry, 1996). Although none of these areas has the spatiotemporal resolution of the White River sequence, each of these sites preserves important floral and faunal evidence that documents local paleoenvironmental and paleoclimatic conditions associated with the EOT. Only a few are discussed here. Prothero and Emry (1996) provide detailed descriptions of these other areas.
The strata of John Day Fossil Beds National Monument are several hundred meters thick and are noted for their preservation of a rich vertebrate fossil record, as well as unique assemblages of fossilized plant material. Just as in Badlands National Park, the John Day beds also contain numerous paleosols, some of which are brightly colored (Retallack, Bestland, and Fremd, 2000). Numerous volcanic ashes are also preserved at John Day Fossil Beds National Monument, which allows the record of paleoclimatic change to be tightly constrained in time and compared to other parts of the world. Just as in the Big Badlands of South Dakota, the EOT in Oregon is interpreted as a change from warm and humid conditions to cooler temperate conditions, based on a combination of data from paleosols and paleobotany (Sheldon and Retallack, 2004; Retallack, 2007).
The paleobotanical record of the Pacific Northwest indicates several fluctuations in temperature and precipitation during the last 5 million years of the late Eocene, culminating with a major episode of cooling of 3 to 6°C, increased seasonality, and a decrease in precipitation (Myers, 2003). According to Wolfe (1992, 1994), a pronounced changeover in terrestrial flora occurred over approximately 1 million years just after the Eocene–Oligocene boundary. Warm, subtropical vegetation was replaced by deciduous, temperate forms, especially at high and midlatitudes.
At Florissant Fossil Beds National Monument, volcanic activity to the west during the late Eocene blocked a stream and formed lakes, similar to the catastrophic events surrounding the eruption of Mount Saint Helens in 1980. Within the lakes, numerous insects, plants, and occasional small animals were preserved in the layers of ash and sediments that washed into the lake. Surrounding forests were also buried, preserving the stumps of redwood and other trees as large petrified edifices. Although the deposits of Florissant Fossil Beds National Monument do not preserve the Eocene–Oligocene boundary proper, the fossil record of plants and insects provides evidence of forested conditions during the late Eocene (Meyer, 2003) and can be used to better understand regional variations in paleoclimatic conditions during the EOT.
In Montana, numerous small patches of terrestrial strata are preserved in lowlands between mountain blocks (Sheldon and Retallack, 2004; Retallack, 2007). As in the Big Badlands of South Dakota, these fluvial strata preserve vertebrate fossils and paleosols, the analysis of which suggests that paleoclimatic change associated with the Eocene–Oligocene boundary was minimal, if any. On the basis of chemical data from paleosols, overall conditions during the Eocene and Oligocene were wetter in Oregon and drier in Nebraska (Sheldon and Retallack, 2004).
The paleoclimatic record of the EOT in South America, which is based on stable isotopic records from fossil mammalian tooth enamel from the Patagonia region, also shows no discernible change in climate (Kohn et al., 2004; Kohn, Zanazzi, and Josef, 2010). In comparison to other regions across the globe, the typically observed change from forested to open conditions across the EOT had already occurred in Argentina by the late Eocene.
Records of the nonmarine EOT in Europe are variable in both their degree of preservation and the interpretations that have been drawn from them. Paleobotanical records from Germany suggest cooling (Roth-Nebelsick et al., 2004), whereas isotopic records from mammalian tooth enamel, mollusks, and fish fossils suggest fluctuations in climate but no clear trend in either temperature or humidity (Grimes et al., 2005). According to Hren et al. (2013), the EOT in southern England does indeed record a transition to cooler and likely drier conditions. On the Isle of Wight in southern England, paleoclimatic conditions were static, with consistent temperatures and an overall increase in precipitation, although there may have been an increase in seasonality (Sheldon et al., 2009). Records of paleoclimatic conditions preserved in paleosols from the Ebro basin of Spain also suggest that conditions were unchanged in this region across the EOT.
As in North America, the most dramatic evidence of paleoenvironmental disturbance associated with the EOT in Europe is the extinction of numerous lineages of fossil vertebrates. This event, referred to as the Grande Coupure, marks the extinction of endemic European fossil communities and their replacement with new species from Asia, and is coincident with the first major pulse of glaciation on Antarctica (Costa et al., 2011).
In Africa, the Eocene–Oligocene boundary is difficult to define because of the lack of datable material, but estimates of the position of the boundary based on mammalian fauna assemblages suggest that climate change, extinctions, and turnovers were minimal, and that they were dominated by wet, tropical conditions (Rasmussen, Bown, and Simons, 1992).
The record of the terrestrial EOT in China is sparsely preserved (Xiao et al., 2010). On the northeast corner of the Tibetan plateau, the EOT is preserved as a thick, stacked sequence of ancient lakes, with limited data from ancient soils. The lake record across the EOT in China is one of aridification associated with a reduction in temperature based on evidence provided by the sedimentological record and paleobotanical evidence (Xiao et al., 2010). In the late Eocene these lakes are dominated by couplets of mudstone and gypsum, with the gypsum representing evaporitic phases of lake hydrology. The gypsum beds disappear as deposition continues into the Oligocene, suggesting a reduction in available moisture that would have mobilized the gypsum into these former lake basins. This reduction in moisture and temperature may have been due in part to the uplift of the Himalayan Mountain system, initiated by the collision of India with Southeast Asia earlier in the Paleogene, which altered weather patterns and helped to reduce the amount of CO2 in the atmosphere as a result of increased rates of rock weathering (Ruddiman, Kutzbach, and Prentice, 1997). On the basis of the fossil record of pollen and calculation of paleoaltimetry for this region (Dupont-Nivet, Hoorn, and Konert, 2008), the uplift of the Himalayan Range began at least 4 million years before the EOT and would have been a significant contributor to climate change. More recent research by Licht et al. (2014) suggests that monsoonal intensity during the late Eocene in southeast Asia was as strong as today, and that elevated levels of CO2 in the late Eocene compensated for the decreased elevation of the Himalayan Range at that time. Their data also suggest a weakening of the Asian monsoon during the EOT.
The majority of paleoclimatic and paleoenvironmental data that is used to interpret the dynamics of the EOT has been recovered from marine settings, in particular from analysis of various fossil assemblages and stable isotopic data of carbon and oxygen from these assemblages and associated sediments (Pagani et al., 2005; Pearson et al., 2008). In brief, the EOT in the marine realm was a time of major reorganization. The magnitude of reorganization and response to the EOT varied across the globe as a function of latitude and water depth. Data from marine records show dramatic shifts in oxygen isotope signatures of paleotemperatures, atmospheric shifts in greenhouse gases, shifts in ocean currents, dramatic drops in sea level, and the extinction or proliferation of particular fossil lineages.
The oxygen-stable isotopic record of ocean water conditions is preserved within the calcareous shells (tests) of single-celled planktonic and benthic foraminifera. As individual foraminifera grow, they extract calcium, carbon, and oxygen from seawater to create their tests of calcium carbonate. Although foraminifera generally incorporate slightly greater amounts of O18 into their shell structure, cooler ocean conditions favor even greater amounts of O18. With greater cooling there is a greater discrepancy of O16 versus O18 uptake by the foraminifera. Because the ratio of O16 to O18 present in water molecules is related to temperature, measurements of this ratio in fossil foraminifera over time can show relative trends. For the EOT, the oxygen isotope record shows an overall cooling trend during the late Eocene, with the first pronounced step of cooling at approximately 34.2 Ma and the second, most pronounced step within the earliest Oligocene at 33.6 Ma, which is coincident with a pronounced increase in Antarctic ice volume (Coxall et al., 2005; Miller et al., 2009; Xiao et al., 2010).
Records of carbon isotopes can be extracted from organic sediments. In particular, certain types of organic molecules preserve ratios of carbon isotopes that can be used to calculate atmospheric levels of CO2, which is a greenhouse gas. The carbon isotopes within organic marine sediments over the last 2 million years of the late Eocene show several punctuated drops in the amount of atmospheric CO2 within a trend of gradual declining values leading up to the Eocene–Oligocene boundary (Pagani et al., 2005). By the early Oligocene, levels of CO2 reached modern levels. Such a decline in atmospheric CO2 would promote global cooling.
These stepwise shifts in the isotopic record of stable isotopes in marine environments culminate with a dramatic shift at the EOT. Individual smaller shifts correspond with periods of marked cooling with subsequent periods of gradual and stable warming (Coccioni, Frontalini, and Spezzaferri, 2009). With respect to the EOT, there is a dramatic increase in the abundance of O18 within foraminifera tests, indicating a substantial drop in seawater temperatures of 5°C on average (Liu et al., 2009). This isotopic shift is coincident with both a dramatic drop in global sea levels and the appearance of the first permanent glaciers on Antarctica. Leading up to the EOT, the record of glaciation on Antarctica is sporadic (Miller et al., 2009).
It was also at this time that oceanic circulation patterns were significantly changed by reorganization of tectonic plates. The collision of Arabia with Eurasia, combined with a reduction in global sea level, resulted in the eventual blockage of the subtropical Eocene Neotethys current from the Indian Ocean into the Atlantic (Jovane et al., 2009). The EOT also saw the opening of Drake Passage between Antarctica and South America and between Antarctica and Tasmania, thus allowing the establishment of the Antarctic Circumpolar Current and progressive cooling of ocean waters (Diester-Haass and Zahn, 1996; Exon et al., 2002). According to Miller et al. (2005), the rate of oceanic crustal spreading slowed during the EOT, which would have resulted in a reduction in the size of midocean ridges and would have also allowed ocean levels to fall. This combination of falling sea level and reorganization of oceanic circulation patterns led to numerous chemical changes in the ocean, including greater amounts of calcium carbonate precipitation, increased amounts of nutrient-rich upwelling bottom waters, increased abundances of diatoms (photosynthetic microorganisms with siliceous tests), and a reduction in overall ocean water acidity, especially in deeper ocean settings (Miller et al., 2009).
The overall paleobiological response in the marine realm to the EOT shows a complex combination of appearances, extinctions, and changes in size and diversity of particular lineages depending on latitude and water depth. This is not surprising, given the effects of falling sea levels on habitats due to glaciation and associated drops in global temperature. Prothero, Ivany, and Nesbitt (2003) and Koeberl and Montanari (2009) provide an in-depth treatment of individual marine fossil groups and their response to the Eocene–Oligocene boundary.
The underlying cause of the EOT is still debated. More than likely its causes are similar to the proposed scenarios of climate change and extinction at the Permian–Triassic and Cretaceous–Tertiary boundaries, which were the result of a multitude of factors acting together and influencing each other. In contrast to the Permian–Triassic and Cretaceous–Tertiary boundaries, the EOT in North America did not see a major extinction in the terrestrial realm. Certain lineages went extinct, such as the brontotheres, but most lineages survived. In the marine realm, the extinction of certain microfossil lineages was offset by the origination of others. Although the EOT itself may not have been a period of extreme paleobiological change, the preceding 2 million years witnessed at least three separate asteroid or comet impacts (Poag, Mankinen, and Norris, 2003), as well as the separation of Antarctica from South America and subsequent establishment of the Antarctic Circumpolar Current (Diester-Haass and Zahn, 1996; Exon et al., 2002), the uplift of the Himalayas (Ruddiman, Kutzbach, and Prentice, 1997), and an overall drop in the amount of CO2 in the atmosphere (Pagani et al., 2005).
Evidence of comet and asteroid impacts are preserved throughout the geologic record. The degree to which an impact event can possibly influence environmental conditions is variable and depends on the size, composition, and speed of the impactor, as well as the chemistry of the target rock (land versus ocean). Comets are essentially dirty snowballs with variable proportions of frozen materials, such as methane and water, mixed with rock. Asteroids and meteors are more variable in composition; they include stony bodies and almost pure metal.
During an impact, the velocity of the impactor is transferred to the target rock as kinetic energy in the form of heat and pressure. This results in the vaporization of the target rock for a defined distance away from the site of impact, as well as the destruction of the impactor. Vaporization is accompanied by melting of target materials and excavation of a crater. The material created by the impact (ejecta) will vary as a function of distance away from the target site. Areas proximal to the impact site will preserve coarser material as an ejecta blanket from the excavation of the crater and the size of the ejecta decreases with distance. Materials that were vaporized or melted eventually cool and condense into glassy spherules. These layers of impact debris (impactites) can sometimes traverse the globe, depending on the magnitude of the event (e.g., the end-Cretaceous impact at Chicxulub). In addition to glassy spherules, evidence of impacts in ejecta layers includes enrichments in certain elements that are not found in great abundance in the Earth’s crust (such as iridium), as well as shocked minerals (most commonly quartz) that show microcrystalline lines of deformation that result from the pressure of impact at the target site.
At least three separate impact events are reported over 2 million years of the late Eocene based on records from Italy (Poag, Mankinen, and Norris, 2003), and at least three separate impact craters have been reported. The two largest craters are Popigai in northern Siberia (100 km wide) and the Chesapeake Bay crater (85 km wide), located at the tip of the Delmarva Peninsula along the Atlantic coast of the United States (Coccioni, Frontalini, and Spezzaferri, 2009). A third, smaller crater (20 km) is found on the continental shelf east of Toms River, New Jersey. The relationship of these impacts to particular ejecta layers is unknown, although links to distinct proximal ejecta deposits have been established, such as the correlation between the North American strewn field and the Chesapeake Bay impact (Poag, Mankinen, and Norris, 2003). Some issues of correlation are related to ambiguity of geologic dating of individual ejecta layers, as well as dating between sections.
Ejecta layers have not yet been reported from terrestrial EOT deposits, even though several locations preserve strata of the correct age (such as the Flagstaff Rim section of Wyoming). Numerous EOT marine core sections with ejecta have been reported (Poag, Mankinen, and Norris, 2003), which is likely due to the enhanced preservation potential of marine settings. When placed within a temporal framework of impacts versus extinctions and paleoclimatic perturbations at the GSSP in Massagnano, Italy, impacts appear to have had no direct effects on planktonic foraminifera, although changes in the proportions of particular types of warm-water versus cool-water foraminifera are seen (Coccioni, Frontalini, and Spezzaferri, 2009). The sizes of the Popigai and Chesapeake Bay impacts suggest that some form of environmental perturbation would be expected. For comparison, fragments of comet Shoemaker-Levy 9, which impacted Jupiter in July 1994, were 1 km or less in diameter, but the atmospheric blemishes that were created were larger than the planet Earth. Although these individual late Eocene impacts did not generate punctuated extinction events, the last 2 million years of the late Eocene may have been subjected to an increased rate of overall cosmic bombardment based on elevated amounts of extraterrestrial helium found in sediments (Koeberl, 2009).
All of these events likely contributed to the transition from a hothouse to icehouse paleoclimate, but what was the tipping point? Was it a gradual trend that eventually reached a threshold? What were the drivers? In order to answer these questions, the rate and timing of paleoenvironmental and paleoclimatic changes in terrestrial and marine realms need to be correlated and compared. The paleoclimatic records of the EOT do not always agree, which is not unexpected, given the current global variation in localized climates. The key is to understand the global variation and regional trends of paleoclimatic conditions during the EOT. With a terrestrial–marine chronologic framework of paleoclimatic data, global climate models can be developed in order to decipher the EOT. Such models are important for critical periods of paleoclimatic transition because they can be used to better understand our current patterns of climate change and to predict future climate responses. Once the drivers have been identified, it may be possible to link these to the patterns of extinction of some lineages and changes in the North American fauna across the EOT.