Chapter 16
The Evolution of Life
Evolutionary faith
The idea of evolution underlies much of modern political, economic, and social theory; it pervades geology and biology, and has become the basis of our entire cosmology. Indeed it shapes the way we think about practically everything. But it is more than a predominant way of thinking: it also has a deep intuitive appeal. One reason is that it is a theory of original unity: it explains the diversity of the world in terms of a common source. On the grandest scale, everything in the universe has its ultimate origin in the primordial explosion, the Big Bang. The Sun arose from the same galactic cloud as other stars in our galaxy; the Earth from the same spinning disc of matter as our brother and sister planets. All forms of life are thought to have come from a common ancestral form, perhaps even from a single primal cell. We are therefore related to all living things, and ultimately to everything in the universe. One of the great themes of traditional creation myths is the division of a primal unity into many parts, the emergence of the Many from the One. Clearly modern evolutionary theories fulfil this mythic role.
Another aspect of the idea of evolution that has a numinous appeal is its affirmation of a continuing creativity in the universe, in life, and in humanity. The creative process did not occur only in the distant past, in the mythic time of origins; it has been going on ever since, and is still going on today. Our fascination with innovations and human creativity is one way in which we experience evolution as a living idea, or more than an idea: a faith, which is not just a matter of belief, but also involves confidence and trust. Like other faiths, the evolutionary faith has a self-fulfilling quality, and enables innovations to be made at an ever-accelerating rate, at least in the scientific, technological, and economic realms.
Even those who reject a simple faith in material progress, or who fear worldwide disaster as a result of the advance of technology, do not generally reject the basic idea of evolutionary development. Rather, they stress that as well as material progress there is an urgent need for progress in the political, social, moral, or spiritual realms, or in all these areas together.
Whether or not the evolutionary faith is recognized as essentially religious or ideological in nature, it arouses what seem very like religious passions in its defenders;1 and like traditional religious faiths, evolution is interpreted very differently by different schools of thought.
Within biology, as in other fields, debates on the nature of evolution have usually been conducted in a sectarian spirit, and still are: neo-Darwinians versus neo-Lamarckians, gradualists versus saltationists, sociobiologists versus Marxists, and so on. The passions aroused can be intense.2 Truth itself seems to be at stake, and the opponents (whoever they are) seem to be propagating profoundly false theories, generally with dangerous political, social or religious implications.
The various interpretations of evolution are indeed closely linked to social, political and religious systems. Thus, for example, a school of biology in Japan emphasizes the importance of co-operation by groups of organisms in the evolutionary process, whereas neo-Darwinians emphasize competition between individuals. The two sides accuse one another of merely reflecting the social assumptions of their own culture.3 Likewise, sociobiologists emphasize the importance of selfish genes in competition with each other, while their left-wing opponents see in this a reflection of right-wing political doctrines.4 Materialists believe that the entire evolutionary process is purposeless and results from the interplay of blind chance and necessity;5 pantheists emphasize the spontaneity and creative power of nature;6 and theists believe that nature itself in some sense arises from the divine being and that the evolutionary process has a spiritual purpose.7
The various schools of thought commonly criticize each other on the grounds that they start from preconceived assumptions. And so they do. But who does not?
Darwin himself certainly based his ideas on assumptions that have been questioned again and again. I first turn to a re-examination of these assumptions, partly because this helps to clarify most of the subsequent evolutionary controversies and partly because this historical context makes the evolutionary implications of the hypothesis of formative causation easier to grasp. I then consider the evolution of morphic fields, and the possible role of morphic resonance in the phenomena of atavism and parallel evolution.
The ambiguity of Darwin
Underlying Darwin’s evolutionary vision was a strong sense of the autonomy, spontaneity and creativity of nature. He could not help thinking of nature as alive. But in order to affirm the creativity of nature on Earth, he had to deny its dependence on the transcendent God of nineteenth-century Protestant theology. In his denial of any creative role for God, conceived of as an external designing intelligence, he adopted the doctrine of materialism.8 He accordingly attempted to expel as much mystery as possible from the working of nature, reducing everything to the operation of blind laws and blind chance. He took these laws to include some of the social and economic principles that were influential in Victorian England, including Malthus’ theory of population and an emphasis on individual competition and self-advancement; he also took for granted a kind of common-sense utilitarianism.
Darwin’s attitude to nature was inevitably ambiguous. His sense that nature is alive seems to have been one of his primary intuitions; but then he continually denied it, or at least relegated it to unconsciousness. As a materialist, he assumed on theoretical grounds that nature is dead. He was well aware of this ambiguity:
The term ‘natural selection’ is in some respects a bad one, as it seems to imply conscious choice; but this will soon be disregarded after a little familiarity … For brevity’s sake I sometimes speak of natural selection as an intelligent power … I have, also, often personified the word Nature; for I have found it difficult to avoid this ambiguity; but I mean by nature only the aggregate action and product of many natural laws – and by laws only the ascertained sequence of events.9
Darwin’s primary model of the evolutionary process was the development through human activity of the many breeds and varieties of domesticated animals and cultivated plants. Drawing on the experience of animal and plant breeders, he concluded that three fundamental principles were involved in evolutionary transformation: the spontaneous variability of living organisms and the tendency for offspring to resemble their parents; the effects of the environment and of habit; and selection. We will discuss these in turn, and then consider how they can be re-evaluated in the light of formative causation.
Spontaneous variation
Darwin gave many examples of spontaneous variation: the loss or gain of entire structures such as vertebrae, petals, nipples, and even whole limbs; sudden and dramatic changes in patterns of growth and development; and many other kinds of variation, for example in colour, pattern, and behaviour. He showed how such variants or sports had on many occasions been selected by breeders and formed the basis of a new breed or variety: one example was the dwarf ancon sheep (Fig. 16.1); another was the spontaneous appearance of nectarines from peaches.10
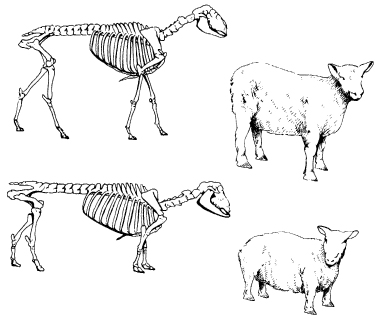
Figure 16.1 A normal sheep (above) compared with the dwarf ‘ancon’ or ‘otter’ breed, together with their corresponding skeletons. (After Stanley, 1979) This breed originated with a sudden mutation, rather than passing through gradual intermediate forms. Darwin described it as follows: ‘In 1791, a ram-lamb was born in Massachusetts, having short crooked legs and a long back, like a turnspit-dog. From this one lamb the otter or ancon semi-monstrous breed was raised; as these sheep could not leap over the fence it was thought they would be valuable … [T]he ancons have been observed to keep together, separating themselves from the rest of the flock when put into enclosures with other sheep’ (Darwin, 1875).
Biologists who continued this line of enquiry provided many more examples of spontaneous jumps or discontinuities; new types arose from the old directly, without passing through a series of intermediate forms.11 After the beginning of the twentieth century such changes were generally referred to as mutations, from the Latin mutare, to change. The study of the inheritance of variations has been the basis of the science of genetics from Mendel onwards. In the course of the twentieth century it became clear that at the genetic level, mutations could involve changes ranging from the loss or gain of entire chromosomes, through large-scale changes in chromosome structure, right down to changes in single base-pairs within DNA molecules.
The word mutation is now commonly used to refer to such genetic changes, especially to changes within DNA molecules; and this seems to imply that the spontaneous variation of organisms can be reduced to the spontaneous variation of genes. But a mutant organism is one in which such changes are expressed, and this expression takes place within the context of its co-ordinated development and behaviour. For example, genetic mutations in fruit flies that lead to the development of four wings instead of two (Fig. 5.6) alter the morphogenesis of an entire segment of the fly; but as we have seen, the mutant gene does not contain or program this development (see above). Recall the television analogy: the mutation of a condenser in the tuning circuit may cause it to tune in to a different channel; but the program on this channel is not encoded in the mutant condenser. Darwin himself was very aware of what he called the ‘co-ordinating power’ common to all organic beings, which according to the hypothesis of formative causation is due to morphic fields. In his own words:
We may infer that, when any part or organ is either greatly increased in size or wholly suppressed through variation and continued selection, the co-ordinating power of the organization will continually tend to bring again all the parts into harmony with each other.12
Ever since Darwin, a number of influential biologists have seen in such large-scale mutations the most probable way in which new types of organisms have arisen.13 The geneticist Richard Goldschmidt, for example, expressed a rather extreme form of this view:
Species and the higher categories originate in single macro-evolutionary steps as completely new genetic systems. The genetical process which is involved consists of a re-patterning of the chromosomes, which results in a new genetic system … This new genetic system … produces a change in development which is termed a systemic mutation … The facts of development, especially those furnished by experimental embryology, show that the potentialities, the mechanics of development, permit huge changes to take place in a single step.14
Goldschmidt called such radically mutant organisms ‘hopeful monsters.’ Such an emphasis on large-scale changes reduces the creative role of natural selection in the evolutionary process and is controversial precisely for this reason. It locates the main source of evolutionary creativity within living organization itself – new kinds of organisms appear spontaneously. Darwin knew that this was true of domesticated plants and animals and that it was the source of established breeds and varieties, such as the ancon sheep. But he denied that it played an important part in natural evolution, preferring to emphasize the creative power of natural selection instead. Although spontaneous variation remained an essential feature of his theory as the source of evolutionary novelty, he tried to minimize its role; and the simplest way to do this was to concentrate attention on small variations rather than large ones. The smaller they were, Darwin felt, the less mysterious they seemed, and the more scientific his theory became (see above).
The effects of habit
In Darwin’s day, it was widely assumed that acquired characteristics could be inherited. Darwin shared this belief and cited many examples to support it.15 In this respect he was a Lamarckian, not so much because he was following Lamarck but because he and Lamarck both accepted the inheritance of acquired characteristics as a matter of common sense. It was a view so widely accepted that it was taken for granted.16
Lamarck placed a strong emphasis on the role of behaviour in evolution: animals’ development of new habits in response to needs led to the use or disuse of organs, which were accordingly either strengthened or weakened. Over a period of generations, this process led to structural changes that became increasingly hereditary (Chapter 8). Lamarck’s most famous example was the giraffe:
It is interesting to observe the result of habit in the peculiar shape and size of the giraffe: the animal, the largest of the mammals, is known to live in the interior of Africa in places where the soil is nearly always arid and barren, so that it is obliged to browse on the leaves of trees and to make constant efforts to reach them. From this habit long maintained in all its race, it has resulted that the animal’s fore-legs have become longer than its hind legs, and that its neck is lengthened to such a degree that the giraffe, without standing up on its hind legs, attains a height of six metres.17
In this respect too Darwin agreed with Lamarck, and he provided various illustrations of the hereditary effects of the habits of life. For example, domestic fowls, ducks and geese have almost lost both the habit of flying and the power of flight. He carefully compared the skeletons of domesticated breeds with the wild parent species and showed that there had been a general decrease in weight and size of wing bones and an increase in leg bones relative to the rest of the skeleton, which he thought was probably the indirect result of the action of the muscles on the bones: ‘There can be no doubt that with our anciently domesticated animals, certain bones have increased in size and weight owing to increased or decreased use.’18 He supposed that similar principles had obtained under natural conditions: ostriches, for example, may have lost the power of flight through disuse and gained stronger legs through an increased use of them over successive generations.19
Darwin was very conscious of the power of habit, which was for him almost another name for nature. ‘Nature, by making habit omnipotent, and its effects hereditary, has fitted the Fuegian to the climate and productions of his miserable country’, he wrote succinctly.20 Francis Huxley summarized Darwin’s attitude as follows:
A structure to him meant a habit, and a habit implied not only an internal need but outer forces to which, for good or evil, the organism had to become habituated … In one sense, therefore, he might well have called his book The Origin of Habits rather than The Origin of Species. Like many others, he was never quite sure what a species was.21
Many biologists have followed Darwin in his emphasis on the cumulative effects of habit, and the ostrich remains a favourite example. These birds are born with horny calluses on their rumps, breast, and pubis, just where these will press upon the ground when they sit (Fig. 16.2). It is easy to suppose that their ancestors developed these calluses through their habit of sitting; over many generations the tendency to develop them became more and more pronounced, and eventually they developed even in embryos. Likewise, warthogs have hereditary calluses on their knees, corresponding with their habit of kneeling while they root in the ground.22 So do camels, again in agreement with their habit of kneeling (see above). We ourselves show such effects, as Darwin pointed out:
Everyone knows that hard work thickens the epidermis on the hands; and when we hear that with infants, long before birth, the epidermis is thicker on the palms and soles of the feet than on any other part of the body … we are naturally inclined to attribute this to the inherited effects of long-continued use or pressure.23
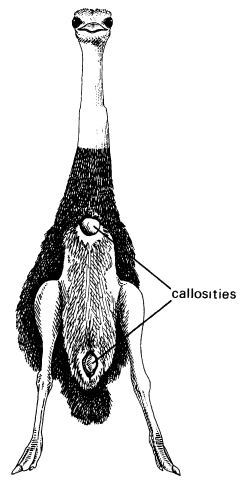
Figure 16.2 The underside of an ostrich’s body, showing the callosities on which it rests while sitting. (Cf. Duerden, 1929)
Such Lamarckian ideas have an immediate intuitive appeal. The problem was that no one could propose a plausible mechanism whereby acquired characteristics could be inherited materially. Darwin had a try: in his hypothesis of pangenesis he proposed that all the units of the body throw off tiny gemmules of ‘formative matter, which are dispersed throughout the body, multiply, and aggregate in the buds of plants and in germ cells, through which they are transmitted to the offspring’.24 This theory never attracted much support, and now seems highly implausible. More modern attempts to conceive of Lamarckian inheritance in terms of the transfer of specifically altered genetic material from various parts of the body to the germ cells have likewise met with no success.
Mendelian geneticists, following Weismann, denied axiomatically that such inheritance can occur. But then how can the inherited calluses of the ostrich, for example, be explained? The natural selection of chance mutations is the ready-made neo-Darwinian answer, but as C.H. Waddington observed:
Can we really be satisfied with a theory that suggests that, purely by chance, a hereditary change has turned up which produces callosities in just the right places, and that the sitting habit of the ostrich has nothing to do with it?25
Waddington, in his own experiments with fruit flies, showed that acquired characteristics could indeed be inherited. He explained this effect in terms of chreodes or pathways of development (Fig. 6.2), and called the process genetic assimilation (Fig. 8.3):
After a time we shall find that the path leading to the adapted condition is better defined than the main path, and also that it has become easier for development to choose that path. The threshold between the adapted alternative and the original main track will have been lowered. If this lowering goes far enough, the alternative will become the main track, and genetic assimilation will be complete.26
He attempted to account for this effect in terms of the selection and accumulation of mutant genes within the population, and thus provided a seemingly orthodox neo-Darwinian explanation for it, one that has been quite widely adopted by modern evolutionary theorists. But as we have seen, more recent reinvestigations have failed to support his explanation: the effect occurs even in the absence of genetic selection in favour of four-winged flies and seems much more Lamarckian than Waddington supposed (see above).
As we saw in Chapter 8, the inheritance of acquired characteristics used to be problematic from a mechanistic point of view. Both neo-Darwinians and neo-Lamarckians made the conventional assumption that heredity is explicable in terms of chemical genes, and hence inherited changes must be due to changes in the genes. However, since the beginning of the twenty-first century, the recognition of epigenetic inheritance has provided a new explanation: changes not in the genes themselves but in the way they are expressed can become hereditary, for example through modifications of the proteins in the chromosomes or through the methylation of DNA.
The idea of formative causation provides another way in which acquired characteristics can be inherited, through morphic fields, which are inherited non-genetically by morphic resonance. Through repetition, new patterns of development and behaviour become increasingly habitual. Organisms can indeed inherit habits of behaviour and of bodily development, as both Lamarck and Darwin supposed.
Natural selection
That natural selection plays some part in the evolutionary process seems beyond dispute: countless species and indeed whole ecosystems have become extinct, while others have not. Natural processes, including competition between organisms, climatic and ecological changes, and global catastrophes leading to mass extinctions, have had a selective effect. Natural selection weeds out organisms and species that are not adequately fitted to their environment, for whatever reason.
Darwin, however, gave natural selection a more positive and creative role:
It may metaphorically be said that natural selection is daily and hourly scrutinizing, throughout the world, the slightest variations; rejecting those that are bad, preserving those that are good; silently and insensibly working, whenever and wherever opportunity offers, at the improvement of each organic being in relation to its organic and inorganic conditions of life.27
This idea is plausible in so far as it relates to the development of locally adapted varieties, races, and subspecies, and it has never been widely disputed as a partial explanation of evolutionary adaptation. But Lamarckians, like Darwin himself, emphasized that the hereditary effects of the habits of life also played an important role. The main problem that Darwin and Darwinians have always faced is to account for the origin of species themselves, or of genera, families, and the higher orders of living organization. The idea that such large-scale evolutionary processes all took place gradually over very long periods of time has been challenged again and again. How could complex structures such as eyes, wings and feathers have evolved gradually before they became functional wholes? Why do plants and animals fall into distinct types, such as ferns, conifers, insects and birds, rather than lying on a continuous spectrum of living forms?
The fossil record has always seemed more consistent with the idea that new forms of life have arisen suddenly, or at least rapidly. But Darwin and neo-Darwinians have always argued that these discontinuities in the fossil record exist because of its patchiness and imperfections. This argument has been challenged from Darwin’s day onwards, and by the 1980s was under attack from a number of leading palaeontologists:
Today the fossil record – a rich store of information that was long untapped – is forcing us to revise this conventional view of evolution. As it turns out, myriads of species have inhabited the Earth for millions of years without evolving noticeably. On the other hand, major evolutionary transitions have been wrought during episodes of rapid change, when new species have quickly budded off from old ones. In short, evolution moves by fits and starts.28
But the firm adherence of Darwin and neo-Darwinians to the doctrine of gradual change has little to do with empirical evidence: behind this controversy lies a question of dogma. Darwin’s principal objective was to replace the idea of design by God with spontaneous natural processes. The conventional theology of his day sought to account for the intricate adaptations of plants and animals in terms of divine intelligence; Darwin postulated natural selection instead. The controversy still lives on, with advocates of ‘intelligent design’ putting forward molecular versions of the old ‘argument from design’.29
In order to prevent the reintroduction of God to account for sudden creative jumps, Darwin had to deny that such jumps were of any significance in the evolution of life, or he felt he had to minimize their importance. He equated this denial with science itself (see above). Most of his followers share this attitude, summarized by Richard Dawkins as follows:
In the context of the fight against creationism, gradualism is more or less synonymous with evolution itself. If you throw out gradualness you throw out the very thing that makes evolution more plausible than creation. Creation is a special case of saltation – the saltus is the large jump from nothing to fully formed modern life. When you think of what Darwin was fighting against, is it any wonder that he continually returned to the theme of slow, gradual, step-by-step change?30
The replacement of divine design by natural selection has led neo-Darwinians into an intellectual habit that, ironically, resembles that of old-fashioned theodicy, the attempt to justify the ways of God to men. According to this kind of theology, God as a perfect and omniscient being must have created the best of all possible worlds, hence everything that happens must have some providential reason, even if this is not at first apparent. Neo-Darwinians, confronted with any given feature of a species, generally assume that it must have some purpose or adaptive value, and then speculate about the selective pressures that must have given rise to it. Such speculations are usually untested and untestable; they are in fact rather like fables: how the rhino got its horn, how the peacock got its tail, and so on. Neo-Darwinism permits a limitless supply of ‘just so’ stories to be spun, all with the same moral of competitive success, and taking place in a relentlessly utilitarian world.
Darwin, with his customary and disarming honesty, towards the end of his life came to recognize that he had exaggerated the role of natural selection:
I was not, however, able to annul the influence of my former belief, then almost universal, that each species had been purposely created; and this led to my tacit assumption that every detail of structure, excluding rudiments, was of some special, though unrecognized, service. Anyone with this assumption in his mind would naturally extend too far the action of natural selection, either during past or present times.31
Some of those who accepted the idea of sudden jumps did indeed see in them the expression of creative power, if not of God himself.32 But others thought of them as a matter of chance, similar in nature but larger in scope than the small mutations on which neo-Darwinian theory is based. In this case, there is a difference only of degree between the evolutionary role of large and small random changes. Yet others stress that mutations occur in the context of developmental and behavioural patterns that constrain the possible changes that can occur. A mutant horse may have extra toes, for example, but there are no mutant horses with wings, feathers, or flowers.
One of Darwin’s staunchest supporters, T.H. Huxley, warned him at an early stage against his insistence on gradual change:
Mr. Darwin’s position might, we believe, have been even stronger than it is, if he had not embarrassed himself with the aphorism Natura non facit saltum which turns up so often in his pages. We believe that nature does make jumps now and then, and a recognition of this fact is of no small importance.33
But Darwin did not heed Huxley’s warning, and was plunged into continual controversy as a result. As he rather ruefully noted after years of dispute: ‘There are, however, some who still think that species have suddenly given birth, through quite unexplained means, to new and totally different forms.’34 This remains as true today as when he wrote it in The Origin of Species.
Darwin and his followers insisted on gradual changes in order to exclude as much mystery as possible from the evolutionary process, and above all to leave no openings for the creative activity of God. But the disadvantages of the dogma of gradualism are that it conflicts with the fossil record, which suggests that large evolutionary changes have in fact occurred quite suddenly, and with the well-established fact that sudden mutations with large-scale effects do indeed occur – for example in the ancon sheep that Darwin himself described (Fig. 16.1) and in homeotic mutations (Figs. 5.6 and 8.2).
The notion that biological evolution involves both sudden jumps and gradual changes seems to fit the facts far better than a dogmatic insistence on gradualism alone. Moreover, it is consistent with what we know about evolution in other spheres, including the evolution of science itself. New theories and paradigms come into being through sudden, intuitive leaps; but then within an established field of science, within the framework of a generally accepted paradigm that soon becomes habitual, research progresses in a relatively gradual and cumulative manner (Chapter 15).
The theory of evolution by natural selection itself is no exception. As we saw in the preceding chapter, it came to Wallace as a sudden illumination, during an attack of malarial fever in the tropics; and it came to Darwin himself in a similarly sudden manner. First, in 1837, he underwent a conversion to evolutionism, abandoning his former belief in the constancy of species: ‘suddenly everything appeared in a new light.’35 Then, on 28 September 1838, came the crucial moment of illumination. In Darwin’s own words:
Fifteen months after I had begun my systematic enquiry, I happened to read for amusement Malthus on population, and being well prepared to appreciate the struggle for existence which everywhere goes on from long-continued observation of the habits of animals and plants, it at once struck me that under these circumstances favourable variations would tend to be preserved and unfavourable ones to be destroyed. The result of this would be the formation of new species. Here, then, I had at last got a theory by which to work.36
After this essential insight, Darwin’s theory matured gradually over many years before he published The Origin of Species in 1859. Thus Darwin’s own intellectual evolution, like the evolutionary process in general, seems to have involved both sudden jumps and gradual adaptations and changes.
The evolution of morphic fields
Evolution occurs at all levels of organization, from atoms to galaxies. At each level of organization, the organized systems – insulin molecules, fruit flies, the instinctive patterns of nest-building behaviour in wasps, flocks of birds, tribal societies, scientific theories – are wholes, Gestalts, morphic units, holons. The evolutionary process is closely connected with the evolution of morphic fields. There are four major consequences of this view.
First, the appearance of new patterns of organization – of new kinds of crystals, for example, or new classes of organisms such as the mammals, or new scientific theories – is associated with the appearance of new morphic fields. Possible creative sources of new fields are discussed in the final chapter; for the purpose of the present discussion the important point is that the appearance of new fields inevitably involves a jump or discontinuity. These fields are wholes, and precisely because of their irreducible integrity they have to appear suddenly. Wholes at all levels of complexity, like the quanta of quantum physics, either exist or do not; by their very nature they cannot come into being gradually.
Of course in new morphic fields there are continuities with what went before, as well as discontinuities. All new fields embrace lower-level morphic units that existed prior to their appearance; these lower-level holons are the parts that are brought into relationship with each other in the new synthesis. For example, new kinds of molecules include atoms, such as carbon and oxygen, which evolved many billions of years ago; when cells with nuclei first arose, they incorporated pre-existing microbial cells within themselves in symbiotic associations;37 many elements of the ancestral reptile form were included in the body plan of the first birds; new instincts include behavioural elements that have already been practised for countless generations; new theories include already existing ideas within them, as for instance Darwin’s theory of evolution by natural selection was a new synthesis that incorporated the pre-existing ideas of evolutionary transformation and the struggle for existence. In general, new patterns include old ones within them. Nevertheless they come into being suddenly; they have a wholeness and integrity that do not admit of gradual appearance.
Second, morphic fields are subject to natural selection. The fields of new patterns of organization that are not viable will not be stabilized by morphic resonance. Only those patterns that are capable of surviving are able to occur again and again. The more frequently they occur, the stronger will their morphic fields become and the more probable the recurrence of the patterns, owing to the cumulative effects of morphic resonance. Natural selection favours some habits more than others; and the more successful a habit, the more it is stabilized by morphic resonance. In the realm of biology, this process results in the evolution of dominance and in the tendency for the most common patterns of form and behaviour to predominate. In the language of genetics this is the ‘dominance of the wild type’ (Chapter 8).
Third, this hypothesis allows for an inheritance of acquired characteristics that does not depend on selective modification of genes, as neo-Lamarckian theories do, or even on epigenetic inheritance through chemical changes in the chromosomes, but is based on heritable modifications of morphic fields in response to the habits of life. The inheritance of morphic fields by morphic resonance from previous similar organisms also allows new patterns of development or behaviour to spread more quickly than conventional genetic inheritance. One possible example is the spread of the habit of opening milk bottles by tits in Europe, or the adaptation of horses to barbed wire (Chapter 9).
Fourth, morphic fields undergo differentiation or specialization in the sense that some versions of the general patterns they organize become more probable than others. Indeed, much of the evolutionary process seems to involve the appearance of variations on basic morphic themes. Domesticated animals and plants provide clear illustrations: think, for example, of the variations on the dog theme represented by breeds such as Afghan hounds and dachshunds, or the varieties of cabbage, like broccoli, Brussels sprouts and kale, all members of the same species.
Many palaeontologists have deduced from the fossil record that when new evolutionary lines begin – when new basic body-plans appear – there is often an ‘intense radiation of types, an ‘explosive phase’ in the early part of their phylogeny, and that only a limited number of the branches continue to develop, and with decreasing speed.’38 One example is the adaptive radiation of the mammals after the sudden extinction of the dinosaurs over 60 million years ago. Most of the orders of mammals came into existence within about 12 million years: carnivores, whales, dolphins, rodents, marsupials, anteaters, horses, camels, elephants, bats, and many others. Indeed most of the basic mammalian forms that appeared then still exist.39
In such evolutionary branchings at whatever level – orders, families, genera, or species – it seems most likely that the various alternative versions of a common basic field came into being by saltation. Many of the new variants may have arisen as a result of chromosomal changes and genetic mutations; some may have been stabilized as a result of historical accidents, for example because they occurred in small, isolated populations; some may have evolved in adaptive response to the conditions of life; some may have evolved gradually in the classical Darwinian manner. But however they arose in the first place, if they proved viable and reproduced successfully, their characteristic versions of the basic morphic fields would have been reinforced by morphic resonance.
None of these considerations denies the role of natural selection at the genetic level; organisms whose genetic constitution is associated with more viable patterns of organization will be favoured by natural selection, and changes in gene frequencies within populations will consequently occur, as neo-Darwinians suppose. But evolution means more than a change in gene frequencies: it involves the natural selection and stabilization of patterns of organization brought about by morphic fields. These fields themselves evolve. Their expression is affected by the conditions and habits of life, as well as by genetic mutations.
This hypothesis is in surprisingly good agreement with much of Darwin’s thought, including his strong sense of the power of habit. It differs in that it allows for both sudden and gradual changes and leaves open the question of creativity.
Extinction and atavism
Countless species have become extinct for many reasons, including climatic changes and global catastrophes. Complex ecosystems have died out. Entire human languages and cultures have disappeared, and many skills and elements of culture have been lost. What has happened to their morphic fields?
According to the hypothesis of formative causation these fields in some sense still exist, although they cannot be expressed because there is nothing to tune in to them. Even the fields of the dinosaurs are potentially present here and now; but there are no appropriate tuning systems, like living dinosaur eggs, that can pick them up by morphic resonance.
If for any reason – for example a genetic mutation or an unusual environmental stress – a living system comes into resonance with the fields of an ancestral or extinct type, then these fields could be expressed again, and archaic structures could suddenly reappear.40 Such a phenomenon is well known, and is usually referred to as reversion, atavism or throwing back.
Darwin drew attention to many examples of throwing back known to plant breeders: ‘With most of our cultivated vegetables there is some tendency to reversion to what is known to be, or may be presumed to be, their aboriginal state.’41 A similar tendency is well known in domesticated animals, and is often observed when they have run wild. Feral pigs, for example, become more bristly, tend to redevelop tusks, and the coloured stripes reappear in their young, as in young wild pigs. As Darwin commented: ‘In this case, as in many others, we can only say that any change in the habits of life apparently favours a tendency, inherent or latent in the species, to return to their primitive state.’42
Such phenomena are closely related to the ‘dominance of the wild type’ (see above); the ancestral wild-type morphic fields have been around much longer and are more strongly stabilized by morphic resonance than those of the domesticated forms, and hence tend to predominate unless prevented from doing so by human activity and selection.
Darwin believed that atavism underlay many of the mysteries of spontaneous variation, and he concluded his discussion of the subject by reflecting that the germ ‘is crowded with invisible characters … separated by hundreds or even thousands of generations from the present time: and these characters, like those written on paper with invisible ink, lie ready to be evolved whenever the organization is disturbed by certain known or unknown conditions’.43
A striking human example of atavism is the occasional birth of babies with tails; others include the appearance of hind legs in whales and wings in flightless insects.44 One of the best-studied cases is the appearance of extra toes in horses. Modern horses represent the limit of an evolutionary trend for the reduction of toes: they have only one. The hoof is its toenail. Their putative ancestors over 20 million years ago had three or four, and yet further back their progenitors had the original mammalian complement of five. Modern horses develop only vestiges of the second and fourth toes as short splints of bone mounted high above the hoof.
Abnormal horses with extra digits appear from time to time and have been the object of much interest since at least the time of Julius Caesar. A close examination of such animals has shown that in most cases the additional toe is a duplicate of the functional third digit, but some hark back to their remote ancestors by developing one or both of their side splints into toes complete with hooves (Fig. 16.3).45
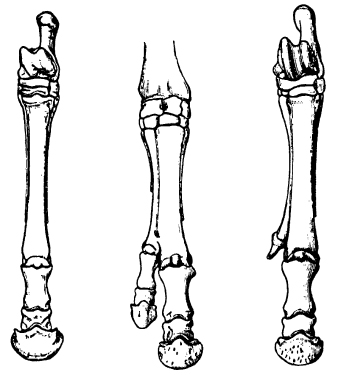
Figure 16.3 Left, the toe of a normal horse, with the vestiges of the second and fourth digits as small splints on either side (cf. Fig. 4.4). Centre, a mutant in which an extra toe has been formed by duplication of the third digit; the side splints are still present. Right, an atavistic mutant in which one of the side splints has developed into an extra toe. (After Marsh, 1892)
In the reptilian ancestors of the birds, the two leg bones between the knees and the ankles, the tibia and fibula, were equal in length, and the ankle region below included a series of small bones. In most modern birds, including hens, the fibula is reduced to a splint and the embryonic anklebones are engulfed by the growing tibia and fuse with it (Fig. 16.4). In some simple and ingenious experiments with chick embryos the archaic pattern reappeared. When a small mica plate was inserted between the tibia and fibula at an early stage of embryonic development, the fibula grew to its full extent, and the anklebones remained separate.46
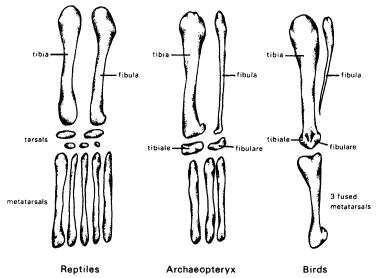
Figure 16.4 A comparison of the lower parts of the hind limbs of reptiles, Archaeopteryx, and modern birds. Note the reduction in size of the fibula and in the number of tarsal and metatarsal bones in Archaeopteryx, and their further reductions and fusions in birds. (After Hall, 1984)
Perhaps the most bizarre example of atavistic development is the formation of teeth by embryonic chick tissue. Archaeopteryx, which is commonly supposed to be the first fossil bird, possessed teeth, but no fossil birds for the last 60 million years have produced them. Epithelium taken from the embryonic gill arches of chicks (the structures that give rise to jaws in vertebrates) was cultured in the laboratory together with embryonic mouse tissue capable of forming the bone and dentin of teeth, but not enamel. The combined tissues gave rise to teeth with chick enamel, which did not look like those of mice.47 Stephen Jay Gould suggested that these may have resembled ‘the actual form of a latent bird’s tooth.’48
On the basis of the hypothesis of formative causation, comparable phenomena of atavism would also be expected in the social and cultural realms. Throughout the ages, those who have feared a breakdown of civilization and a reversion of the social order to a primitive or barbarous state have entertained this possibility. But there may be many less spectacular ways in which extinct ancestral patterns reappear in the course of social and cultural evolution, some as deliberate revivals, and others unconsciously.
Evolutionary convergences
The phenomena of atavism suggest that morphic fields can leap from past to present species across gaps of time and space. But there is no reason why fields should leap only from ancestors to their own descendants: they may also jump sideways, as it were, from one group or type of organism to another, even if these organisms live on different continents. Such a transmission could occur by morphic resonance if, through genetic mutation or through the influence of the environment, organisms of one species tune into the morphic fields of another. This process would permit a kind of morphic plagiarism, even in the absence of any material connection in space or time between the organisms that copy and those that are copied. Morphic fields are not subject to copyright.
In the human realm, there are many examples of parallel social and cultural patterns that seem to have originated independently in different parts of the world. Parallel inventions and discoveries in science and technology are relatively common: a classical case is the independent invention of the differential calculus by Newton and Leibniz.
In some cases, such parallels may be explicable in terms of ‘diffusion’, or in other words transfers by normal means of human communication. Others probably arose in circumstances when individuals or social groups were confronted with similar problems and hit on similar solutions. And no doubt a process of natural selection often similarly favoured solutions that worked.
The idea of morphic resonance is complementary to these well-known explanations. Diffusion would be aided by morphic resonance, which facilitates the learning process (Chapter 10). Confrontation of similar problems is likely to favour a tuning in to solutions already arrived at by others elsewhere. And, owing to morphic resonance, successful patterns of activity, through repetition, show an increasing tendency to reappear. Thus social and cultural morphic fields would be expected to move from one group to another by diffusion and also by leaps, by a kind of action at a distance.
Morphic plagiarism through the picking up of morphic fields by one group of organisms from another may have occurred frequently in the course of biological evolution. It may underlie parallel evolution, in which similar patterns appear in more or less closely related plant and animal species. In some cases, striking similarities are found among organisms that are only very distantly related, and these are generally referred to as evolutionary convergences.
Richard Dawkins has argued that on statistical grounds, following standard neo-Darwinian assumptions:
It is vanishingly improbable that the same evolutionary pathway should ever be followed twice. And it would seem similarly improbable, for the same statistical reasons, that two lines of evolution should converge on the same endpoint from different starting points. It is all the more striking … that numerous examples can be found in real nature, in which independent lines of evolution appear to have converged, from very different starting points, on what looks very like the same endpoint.49
In the plant kingdom, the most familiar examples of parallel evolution are in the forms of leaves, where very similar patterns have appeared again and again in separate genera and families. So striking are the resemblances that many species and varieties are actually named on the basis of these leaf forms, which are borrowed, as it were, from other kinds of plants: salicifolius means willow-leaved, ilicifolius, holly-leaved, and so on.
In some cases, parallel evolution occurs to a remarkable extent in particular geographical areas. One example, to which Frits Went drew attention, is of shrubs in New Zealand with divaricate forms, that is to say with wide-spreading interlaced branches and small orb-shaped leaves.
The frequency with which one encounters this divaricate habit in New Zealand (whereas it is hardly known anywhere else in the world) is indicated by the fact that in New Zealand there are a total of about 50 species of shrubs, divided over 21 families, which are divaricate with interlaced, tortuous branches and reduced leaves. Many of these shrubs look so much alike, without flowers, that one cannot guess even the family, since various species … have exactly the same habit.50
Strangely enough, in most cases this habit of growth appears only in young shrubs, as a juvenile phase, and then gives way to the growth habit typical of the genus; in other species it appears only in the mature phase, and not in the juvenile.
Went carefully considered the standard explanation that this habit is simply an adaptation to the environment favoured by natural selection, and found that it did not fit the facts. First, the idea the divaricate habit protects the plants against browsing animals is implausible because New Zealand was the only extensive geographical area without large native herbivores. Second, the habit appears only at certain stages of growth and has not completely replaced the more usual one. Third, only some species show this habit, and other closely related species do not, although they survive equally well in New Zealand. Finally, since this habit ‘occurs in so many shrubs from different habitats, it does not seem to be an adaptation to the environment.’51
This puzzling phenomenon of similar growth-forms in different species is not confined to New Zealand: there are other examples elsewhere. So striking are these parallelisms that Went considered that an explanation in terms of chance mutations was implausible and came to the conclusion that they indicate some kind of ‘non-sexual character transfer.’ He suggested that entire segments of chromosomes may somehow have been transferred bodily from one genus or family to another.52 However, this hypothetical mechanism could work only over short distances, and other examples of parallelism occur in widely separated places, in both the plant and animal kingdoms.
In butterflies, for example, many striking similarities are found in the patterns of wing colouration, both within and between families (Fig. 16.5). Some of these may be examples of mimicry, favoured by natural selection if a species of similar appearance living in the same environment is protected from its enemies by an unpleasant taste. Predators tend to avoid both the unpalatable insects and those that mimic them. But other cases of parallelism have no such explanation, especially when the species that look alike occur in different places.53

Figure 16.5 Three species of South American butterflies which closely mimic each other. They belong to quite distinct families. Their colours are the same: black, white, and brilliant orange (stippled areas). (After Hardy, 1965)
The two main branches of the mammals, the placentals and marsupials, are thought to have diverged from a common protomammalian ancestor over 60 million years ago and yet provide striking examples of convergence. The marsupials of Australia have given rise to a whole range of species that are similar to placental mammals elsewhere, like pouched versions of ant-eaters, moles, flying squirrels, cats and wolves (Figs. 16.6 and 16.7). The same phenomenon occurred in South America, where marsupials independently gave rise to a range of parallel forms.54 Among placental mammals themselves, there are many examples of parallel evolution, such as the porcupines of South America and the Old World. These are so alike that it has even been suggested that they crossed the Atlantic on rafts of vegetation.55

Figure 16.6 Examples of parallel evolution. A and B, a marsupial flying phalanger and a placental flying squirrel; C and D, marsupial and placental jerboas; E and F, marsupial and placental moles. (After Hardy, 1965)
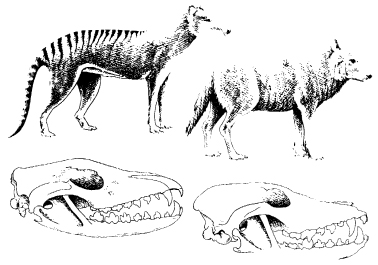
Figure 16.7 Another example of parallel evolution. Left, the Tasmanian wolf, a marsupial; right, the familiar placental wolf. The corresponding skulls are shown below. (After Hardy, 1965)
Even more mysterious is the convergent evolution of similar structures in organisms that are otherwise extremely different. The eyes of vertebrates, for example, have many features in common with the eyes of cephalopods, such as the octopus. When visiting an aquarium it is a strange experience to look an octopus in the eye; the extraordinary similarity in an animal so completely different is uncanny. Yet there are, of course, also differences; most notably, the retina of vertebrates is inverted – the nerves leading from the photocells point forward towards the light – whereas the cephalopod retina is not.
Less well known is the fact that camera-like eyes have evolved independently in several other groups, including the alciopids, a group of marine annelid worms, close relatives of earthworms. Similar eyes have also evolved independently in three different groups of snails. Even more surprisingly, camera-like eyes evolved independently in a group of jellyfish called cubozoans, which lack brains and yet exhibit obvious visual acuity and sophisticated behaviour.56
In his book Life’s Solution, the evolutionary biologist Simon Conway Morris has brought together an unprecedented collection of cases of convergent evolution in both plants and animals, at the level of molecules, organs, morphology and behaviour. He concluded that ‘convergence is ubiquitous and the constraints of life make the emergence of the various biological properties very probable, if not inevitable’.57
The standard neo-Darwinian explanation of such parallelisms and convergences is twofold: first, they have evolved on the basis of random mutations that survived because of similar selection pressures; second, that such convergences occur because of similar structural constraints: there may be only a very limited number of ways of designing an eye, for example. As Dawkins expressed it, such convergent resemblances:
provide most impressive demonstrations of the power of natural selection to put together good designs … The basic rationale is that, if a design is good enough to evolve once, the same design principle is good enough to evolve twice, from different starting points, in different parts of the animal kingdom.58
But what are these ‘good designs’ and ‘design principles’ – the ‘principles’ of porcupines, for instance? They remain unexplained in mechanistic terms (Chapters 5 and 6). From the present point of view, they are inherent in morphic fields. This hypothesis does not contradict the standard explanation but goes beyond it. Natural selection still plays an important role; but it is no longer the great creative power, the designer and sustainer of all forms of life, the ultimate explanatory principle that replaces the God of nineteenth-century natural theology. The designs of living organisms are not imposed upon them from without – by God or natural selection – but are inherent in the organisms themselves. They arise from morphic fields; and these fields are not coded in the genes but transmitted by morphic resonance. Usually subsequent organisms of the same species inherit them; but occasionally organisms of quite different species may pick them up as mutant forms. If these plagiaristic mutants are favoured by natural selection, their forms will tend to be repeated again and again and become habitual characteristics of the plagiarizing species.
The idea of formative causation sheds a new light on biological evolution and greatly extends Darwin’s conception of natural selection to include the natural selection of morphic fields. It emphasizes the role of habit, as Darwin himself did, and allows for a transfer of habits by morphic resonance, not only within a species but also to other species. It thus provides a new understanding of the phenomena of atavism and of parallel and convergent evolution.