Fig. 17-0: Image of the Manicouagan Reservoir, Canada, December 1983. The reservoir, a large annular lake, marks the site of an impact crater 60 miles (100 kilometers) wide. Formed almost 212 million years ago when a large meteorite hit Earth, the crater has been worn down by many advances and retreats of glaciers and other processes of erosion. The reservoir is drained at its south end by the Manicouagan River, which flows from the reservoir and empties into the Saint Lawrence River nearly 300 miles (483 km) south. (Image and information courtesy of the Image Science & Analysis Laboratory, NASA Johnson Space Center; http://eol.jsc.nasa.gov)
Oxidation of the planetary surface took place in the Archean and Proterozoic, where the limited life record does not allow high temporal resolution of events. Beginning in the Phanerozoic, the very high resolution of the fossil record reveals a variety of planetary processes that have a major effect on biological evolution. Catastrophes punctuate the Phanerozoic life record, and the causes appear to be diverse. The extinction of the dinosaurs that marks the Cretaceous/Tertiary boundary is unequivocally linked to the Chicxulub meteorite impact near the Yucatan peninsula, but it also occurs at the same time as the massive Deccan flood basalt province that marked the arrival of a mantle plume at the surface. The even larger extinction at the Permian/Triassic boundary has no known impact crater but occurred after two large flood basalt episodes, the largest of which intersected large coal beds, releasing vast amounts of gases to the atmosphere. Smaller extinction events are also closely associated with large volcanic outpourings from plumes. It may be that the largest extinctions are the result of coupled catastrophic events in a kind of “double whammy.” While catastrophes lead to mass extinctions on a planetary scale, they may be necessary for evolutionary innovations to have the ecospace to succeed and an important factor in promoting evolutionary change.
Plate tectonics is another planetary process that has a major influence on life, once continents become populated. As continents join and separate, and traverse different climate environments, isolation can exist for long periods of time, permitting evolution to proceed on different pathways, creating diversity and permitting multiple experiments at innovation. Having opportunities to explore multiple pathways, evolutionary innovation may be accelerated.
While all of evolution occurred by random mutations, there are nonetheless long-term progressive changes. With time, organisms have developed increasing internal and external relationships—from prokaryotic cells to larger and more complex eukaryotic cells, to multicellular organisms, to organisms with many differentiated organs—as food chains have also expanded. Energy transformation has also increased, with the advent of photosynthesis, the change from anaerobic to aerobic metabolism, and the development of organ specialization first in organelles and later in organs. Increased relationship, organ specialization, complexity, and enhanced energy utilization may be general characteristics of more successful organisms and provide directionality to evolution. The specific pathway of genetic changes is random, but changes that lead to increased energy availability and relationship may be favored. Evolution is also enhanced by a changing planetary surface and by periodic catastrophes that permit evolutionary innovations to be expressed. These catastrophes are delivered from the solar system through meteorite impacts, from the planetary interior through volcanism, and through climate change. Global catastrophes, a changing planetary surface, climate change, directionality in terms of energy and networks are all general phenomena likely to be common in other planetary environments. It is possible that there are general principles exemplified by Earth that apply to a process of co-evolution of planet and life on other habitable planets.
The evolution of life and planet during the Archean and most of the Proterozoic is a picture in low resolution, because the record of life itself in this period is obscure, and the absence of clearly defined and diverse fossils makes high temporal resolution very difficult. It is evident that life and planet co-evolved in the progressive oxidation of the planetary surface, but as we learned in previous chapters, the timing of the major events in the Archean and Proterozoic is only loosely constrained, and the record of unicellular life is vague and obscure. At the end of this period it is generally agreed that O2 in the atmosphere rose to near current levels, and this step in planetary evolution permitted the formation of multicellular animals and, ultimately, the Cambrian explosion of multicellular life that marks the dawn of the Phanerozoic era. Multicellular life is not possible without the energy available from aerobic respiration—no multicellular life, for example, occurs in anaerobic environments on Earth today. Coincident with this step in planetary evolution, our ability to interpret the geological record comes into much sharper focus. The fossil record allows paleontologists to track the numbers of species, how long they last, and when the species assemblage undergoes drastic change. The fossils also initiate an era of tight temporal resolution on a global scale. The increase in resolution permits far more specific questions about co-evolution of life and planet to be considered, as major events can be pinned to specific times and causes.
As we learned in Chapter 14, the geological record since the Cambrian does not show the steady change that might be expected from gradual and progressive mutation of DNA. Instead, there were long periods of gradual change, with individual species existing for an average of 5 to 10 million years—only 0.1–0.2% of Earth’s history—punctuated by times of more abrupt change, when mass extinctions wiped out large proportions of existing life and set the stage for new ecosystems with very different assemblages of organisms. These periods of abrupt change were used to delineate the geological timescale, giving rise to the major geological periods (Fig. 14-4).
Defining mass extinction events requires a quantitative measure of the abundance and diversity of life, and this is possible definitively only after the Cambrian when organisms developed hard parts and fossils became easily preserved. Careful work in slightly older rocks has revealed an earlier multicellular group of organisms called the Ediacaran, whose body parts are preserved in rare soft shales that never underwent significant metamorphism. These animals are sufficiently different from all later species that some paleontologists have inferred that a major extinction occurred near the Precambrian/Cambrian boundary, where the Ediacaran fauna disappeared to be replaced by early Paleozoic species. Earlier mass extinctions may also have occurred, but there is insufficient record to document them. “Snowball Earth” episodes, for example, may have led to mass extinctions.
The development of the fossil record in the Phanerozoic makes it possible to use criteria such as the overall numbers of different genera of fauna and flora to define mass extinctions. Marine organisms are the best index of the diversity of life because sediments progressively accumulate beneath the ocean and are destroyed through erosion on land. Figure 17-1 shows how the types of fauna vary along with total numbers of fauna from the careful compilation of marine genera through geological time (recall that genera are groups of related species). Examination of this figure reveals an overall growth in numbers of genera from the Cambrian to the present, but we see that this growth is not monotonic. There are periods of rapid growth, long plateaus, and sudden dips—the extinction events.
The style of life also has changed dramatically over time, starting with a typical Cambrian fauna from the early Phanerozoic that consisted of invertebrates, such as jellyfish and sponges, and the very recognizable trilobite (Fig. 17-2a). The remaining groups of invertebrate fauna all developed during an explosion of life in the Cambrian and early Ordovician. Between the Ordovician and Silurian there was a significant extinction event, reducing the number of genera by about 50%. Land plants first developed during the Silurian. Significant extinction events also separate the Devonian and Carboniferous. During the Carboniferous, land plants underwent major development, and reptiles first appeared. By the Permian most of the major fossil groups were present, with the exception of mammals. The Paleozoic era is thus marked by an explosive early growth in numbers of genera, and then a long period of relative stability where the number of genera remained relatively constant, subject to extinction events that preferentially eliminated much of the characteristically Cambrian life. Of course on the species level, extinctions and evolution continued apace through this period, but there is nonetheless a recognizable Paleozoic style to life. The difference in fossil assemblage of the Cambrian, the late Paleozoic, and modern times is apparent to the naked eye (Fig. 17-2b,c).
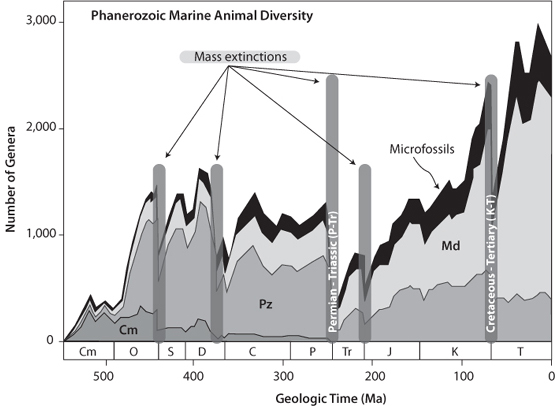
Fig. 17-1: Change in number of genera through time, showing the changes in types of life over time. Note that the Cambrian fauna, for example, peaked in the Ordovician and then went extinct at the Permo-Triassic mass extinction. Cm stands for Cambrian fauna, Pz for Paleozoic fauna, Md for modern fauna. (Modified after Sepkoski, Bulletins of American Paleontology 363 (2002)).
The late Paleozoic stability ended abruptly. At the end of the Permian there were two episodes of mass extinction, separated by about 10 million years. Combined, these two events extinguished 80% of genera on Earth. Trilobites and other families that originated in the Cambrian and had been present for 300 Ma disappeared forever. There were major extinctions on land as well, showing that the causes of this event were truly global in scale. This massive extinction is used as a time marker to separate the Paleozoic and Mesozoic eras. For the first tens of millions of years of the Triassic, the first period of the Mesozoic era, life barely recovered. In the mid-Triassic mammals first appeared. At the end of the Triassic there was a smaller extinction event, followed by a second rapid growth in the numbers of families, soon exceeding what had been present even under the most fecund periods of the Paleozoic. By the Cretaceous period, the giant reptile dinosaurs ruled the Earth at the top of food chains (Fig. 17-2c).

Fig. 17-2: Artist’s renditions of scenes from three different periods of Earth history, all distinct from one another and from modern ecosystems. (a) Cambrian marine scene, where only invertebrate life had developed (© The Field Museum, #GEO86500-052d, with permission). (b) The Devonian period, just before the Permo-Triassic extinction, where more complex plants, fish and other vertebrates were present (© The Field Museum, #GEO86500-125d, with permission). (c) The Cretaceous, the Age of Dinosaurs, just before the Cretaceous-Tertiary extinction (© Karen Carr, with permission (www.karencarr.com)).

The Mesozoic also ended with a mass extinction, used to separate the Mesozoic from the Cenozoic eras. The end Mesozoic extinction caused the demise of the dinosaurs, as well as extinctions of other major fossil groups on both land and sea. This set the stage for another renewal of diversity, with numbers of families recovering rapidly and leading to still greater diversity of life. This new and characteristic assemblage of living organisms had food chains dominated by the newly emergent mammals, which during the Mesozoic had been small, niche creatures scratching out a living in reptile-dominated ecosystems.
From the perspective of evolution, an important aspect of mass extinctions is that they lead to subsequent more rapid evolutionary change. Andrew Bambach and others looked at both genera elimination and genera formation over the last 450 million years. The extinction events led to large negative excursions in the numbers of species. And following these events was the most rapid growth in genera diversity (apparent in Fig. 17-1). The clearing out of the existing ecospace permitted new evolutionary adaptations to flourish. When there is ecological stability, dominant organisms control the ecosystems and there is little room for genetic innovations to take hold. With the wholesale destruction of life, ecological niches open and opportunities for new forms of life appear, permitting more expression of genetic variations. Improved adaptations that were not able to be fully expressed in a dominated ecosystem have room to flourish.
It is evident that evolution on Earth was punctuated and influenced by mass extinctions, catastrophes on a planetary scale that take place in very short periods of time. How short? As shown in Figure 17-3, extinction events occur over centimeters of sedimentary thickness, so the duration must be very short indeed. For the major extinction events that separate the Paleozoic, Mesozoic, and Cenozoic eras, which have received the most attention, the time is so short that its exact duration cannot be well resolved by radiometric dating. Very careful work with zircons has progressively narrowed the timeframes revealing that the Cretaceous/Tertiary extinction occurred in less than 1 million years and that the Permo-Triassic extinction was the result of two events that occurred about 10 million years apart at the end of the Permian. Each of the events was very short in duration from a geological perspective. Owing to the dramatic changes in life that mass extinctions represent, there has been intensive study of their cause, and much controversy. One reason for the controversy is that it now appears that there were multiple causes of mass extinctions, rather than one single theory that is able to explain all of them.

Fig. 17-3: The Cretaceous-Tertiary extinction from La Sierrita, showing the light gray claystone that marks the boundary. Notice the very brief stratigraphic interval over which the boundary occurs. (Photo courtesy of Gerta Keller, Princeton University)
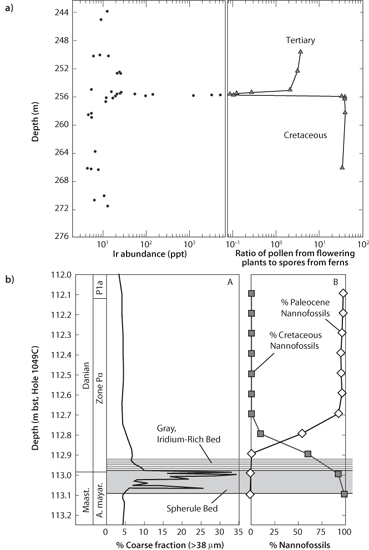
The most famous extinction event is the demise of the dinosaurs at the Cretaceous-Tertiary (K-T) boundary between the Mesozoic and Cenozoic eras. To try to determine the time duration of the extinction, Luis and Walter Alvarez decided to investigate the concentrations of the metal iridium (Ir) in sediments that traverse the time boundary. Iridium is highly siderophile, so most of Earth’s iridium is in the core, and there are very low abundances in the continental crust. Iridium in meteorites is 10,000 times greater than the crustal concentrations. Since a little bit of iridium is continually added to marine sediments, owing to cosmic dust entering the atmosphere and sinking through the ocean, the Alvarezes thought that by measuring the iridium they could constrain the time duration that the few centimeters of sediment represent.
To their surprise, Ir was exceptionally abundant exactly at the K-T boundary, so abundant that it could not be explained by the tiny amounts of cosmic dust entering the atmosphere (Fig. 17-4). This serendipitous discovery led them to propose that a large asteroid hit the Earth at that time, bringing in huge amounts of Ir that spread in a large dust cloud around the globe and accumulated exactly at the boundary. The meteorite impact would be such a catastrophe that it might extinguish much of life on Earth in a matter of years.
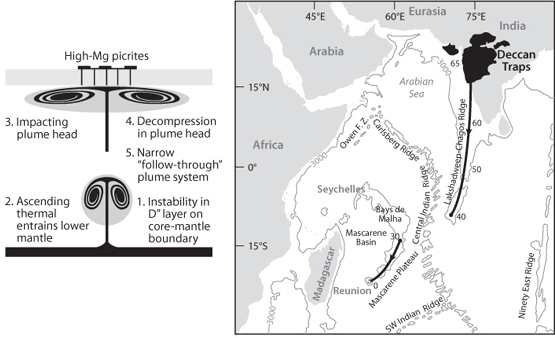
This idea was not universally accepted, and an alternative hypothesis was proposed that unusually abundant volcanism at the K-T boundary was the cause of the extinction. Over less than a few hundred thousand years that span the K-T boundary, a flood basalt province appeared on Earth’s surface. As we learned in Chapter 11, when a plume first arrives at the surface, the “plume head” has a massive volume that produces overwhelmingly abundant volcanism. Many flood basalt provinces represent these initial outpourings, after which a much reduced “plume tail” leads to a long ridge on the surface as the plate migrates over the plume. The first outpourings of the Deccan plume produced millions of km3 of basalts in northern India, called the Deccan Traps, and a hot spot track that is still active today at Reunion Island (Fig. 17-5). This event was large enough to more than double global subaerial volcanism for hundreds of thousands of years, potentially leading to very serious environmental effects that could impact global life. The volcanic hypothesis, however, could not easily explain the Ir evidence.
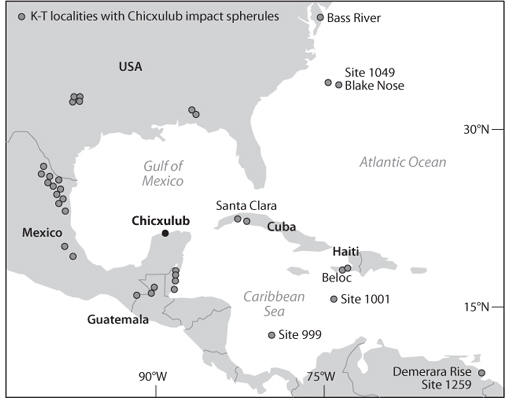
The meteorite impact hypothesis at the K-T boundary became fact by the discovery of the Chixculub impact crater (Fig. 17-6) near the Yucatan peninsula in Mexico that is the exact age of the boundary. Drilling of sediments off the coast of Florida (see Fig. 17-4b) also showed an abrupt change in ocean species coincident with the Ir anomaly, evidence for a huge tsunami, and the telltale signs of impact discussed in Chapter 8—shocked quartz and tektites.
Fig. 17-4: Examples of changes at the Cretaceous Boundary. (a) An example of the iridium spike immediately at the boundary, along with the data showing the dominance of ferns after the impact (creating a negative spike in the angiosperm pollen/fern ratio) (modified from Orth et al., Science 214 [1981], no. 4257: 1341–43). (b) Data from an ocean drilling core off the east coast of Florida showing the boundary and the abrupt change in species that occurs there (adapted from Norris et al., Geology 27 [1999], no. 5: 419–22).
Fig. 17-5: (Left) Illustration of the formation of a new plume from a lower boundary layer, probably the core-mantle boundary, lifting off and heading to the surface. There is a very large “plume head” followed by a narrow plume tail. When the plume head hits the surface it spreads out and generates massive volcanism over a wide range. Following this flood basalt event, which can increase global subaerial volcanism by a factor of 2 or more during its duration, a long and narrow hot-spot track is generated as the plume tail intersects the plate moving above it. (Figure derived from Griffiths and Campbell, Earth and Planetary Science Letters 99, 66–78 (1990)). (Right) The narrow hot spot track generated by the “tail” of the Deccan plume. The current location of the plume is the island of Reunion in the southern Indian Ocean. Numbers by the track indicate the age of volcanism when that portion of the plume track was generated. The track was generated as a continuous feature and later separated by the spreading of the Central Indian Ridge. (Modified from White and McKenzie, J. Geophys. Res. 94, 7685–7729 (1989)).
The fact that a meteorite impact occurred, however, does not necessarily mean that it was the sole causal event. Other large meteorite impacts for which craters have been found, for example, are not associated with mass extinctions in the fossil record. Furthermore, some paleontologists working on the K-T boundary argued that the extinction of life on continents was not as sudden as would be predicted from an impacting event. There is also evidence of radical environmental change prior to the Chixculub impact. This has led to the idea that the mass extinction was the combined result of two major events that overlapped in time—the flood basalt eruptions stressed the global biosphere, and in the midst of this crisis the impact delivered the critical blow.
Fig. 17-6: Map showing the location of the Chixculub impact crater off the coast of Yucatán and the various sites where there is evidence for ejecta, shocked quartz, impact spherules, massive tsunamis, etc., that vary quite regularly with distance from the impact site. (Modified from Gerta Keller, Princeton University; http://geoweb.princeton.edu/people/keller/Mass_Extinction/massex.html#7)
The Permo-Triassic mass extinction was even more catastrophic than the K-T extinction event. Whereas some 50% of genera disappeared at the K-T boundary, 80% disappeared at the end of the Permian. The Permo-Triassic boundary is very well exposed in sediments in China, and exhaustive investigations of these sedimentary sections have revealed that the extinction was a double event, with large extinctions some 10 million years apart (Fig. 17-7). An intensive global search for impact craters has been unsuccessful. In itself this is not definitive—the oldest seafloor is much younger than the P-T boundary, so any impact crater that occurred in the ocean would been subducted long ago, and the oceans are twice as likely to receive an impact as the continents. The impact should have left other traces, however, such as an iridium anomaly, tektites, and shocked quartz, and these have not been found. The events that do occur at the appropriate times for the two P-T events are large volcanic outpourings akin to the Deccan Traps. The older event corresponds with the Emeishan Traps in western China. The younger event that demarks the P-T boundary is simultaneous with the largest volcanic outpouring known in the Phanerozoic geological record. The Siberian Traps (Fig. 17-8) produced more than 2 * 106 km3 of lavas in a very brief period that is indistinguishable in time.
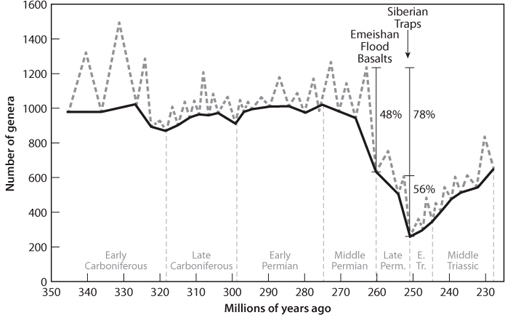
Fig. 17-7: Detailed plot of the change in numbers of genera across the Permo-Triassic boundary. Note the short timescale of the bottom boundary (in millions of years). The mass extinction is clearly seen as a double pulse, with each pulse associated with a continental flood basalt province. (Modified from Knoll et al., Earth Planet. Sci. Lett. 256 (2007):295–313)
The volcanic explanation is supported by one of the characteristics of the end Permian extinction, which is a marked change in the isotopic composition of carbon in the ocean. There is not enough carbon in a meteorite to influence the ocean’s carbon budget, and the meteorite carbon also does not have the necessary isotopic composition. Instead, the carbon isotopes suggest a huge addition of organic carbon to the atmosphere and ocean. Since organic carbon is produced by the biosphere, could volcanic outpourings somehow be associated with such an event?
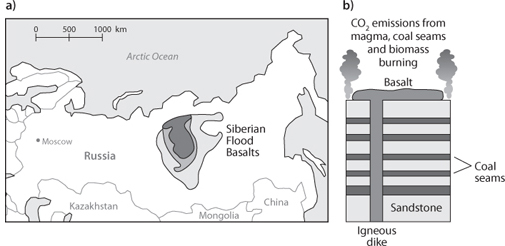
Fig. 17-8: (a) Map of the huge extent of the Siberian flood basalt province associated with the Permo-Triassic extinction. (b) Illustration of how flood basalts might interact with reduced carbon in crustal sediments to lead to huge outpourings of reduced carbon species to the atmosphere. The excess gas emissions would account both for the extinction and the large lowering of δ13C at the P-T boundary.
In the case of the Siberian Traps, they can. The Siberian Traps were erupted into and through sediments very rich in coal, and examples elsewhere of lava flows burning through huge coal seams have been observed. Coal was formed from plant matter and has the right carbon isotopic composition. Furthermore, the burning of coal would lead to massive emissions of CO2 that would have major climatic consequences, including a global warming episode and a change in the acidity of the ocean. The evidence from the Permo-Triassic supports the idea that massive and focused volcanic outpourings are a major driver of extinction events.
Vincent Courtillot, along with colleagues who specialize in precise radiometric dating, has argued that all the major extinction events in the Phanerozoic can be associated with massive flood basalt outpourings (Fig. 17-9a). This figure is not as convincing as it seems, however, because selection of volcanic events and extinctions can be used to force a good correlation (Fig. 17-9b). Impacts are more difficult to date precisely because of their limited aerial extent and lack of datable materials. Simon Kelley has shown, however, that even with the dating errors the correspondence is significantly better for volcanic events than for impacts (Fig. 17-10). Therefore, with the exception of the K-T boundary, which is unambiguously associated with a specific impact, massive volcanic outpourings seem to have a more lasting environmental and biological effect. Recently Courtillot and Olsen showed an intriguing relation between the largest extinction events and the behavior of Earth’s core, as apparent in the reversals of the magnetic field. The “killer plumes” that led to the largest mass extinctions occurred after long intervals where no magnetic reversals were taking place. Since large plumes are thought to originate at the core-mantle boundary, core-mantle interactions may be linked to the major evolutionary events of life at the surface.
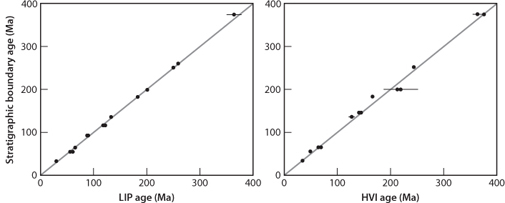
Fig. 17-9: (Right) Cross plots showing possible correlations between meteorite impacts (HVI = high velocity impacts) or continental flood basalt provinces (LIP = large igneous province). (Left) Times of massive environmental change, usually associated with mass extinctions. (Modified after Kelley, Geol. Soc. London 164 (2007):923–36)
It is probably significant that the largest extinctions, at the Permo-Triassic and Cretaceous-Tertiary boundaries, both appear to result from a double whammy, two flood basalts in close temporal proximity at the end of the Permian and a flood basalt and meteorite impact at the end of the Cretaceous. This would account for the observation that there are both meteorite impact and flood basalt events in the geological record that are not associated with mass extinctions. The biosphere appears to have been sufficiently robust that a “one-two punch” is necessary for the most catastrophic extinctions.
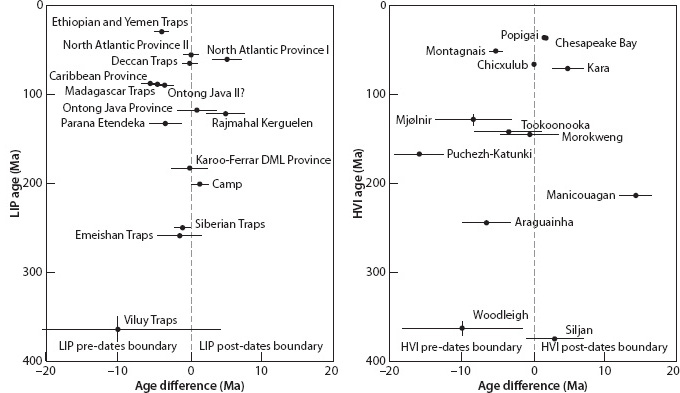
Fig. 17-10: A more detailed examination of the age offsets between dated impacts and mass extinctions. LIP stands for Large Igneous Province. Note the exact correspondence for the Chixculub impact at the K-T boundary, and the significant offsets for many other impacts, except for those with very large error bars. (Modified after Kelley, Geol. Soc. London 164 (2007): 923–36)
The Phanerozoic record thus shows the close linkages between biological evolution and events determined by the solar system and planetary interior. Meteorite impacts that are the remnants of planetary accretion have had a major influence on life, and core/mantle interactions that give rise to mantle plumes that are the active component of upwelling mantle convection have also greatly influenced life’s history. The association of volcanism with mass extinctions also suggests a finely tuned balance of biological and solid earth processes. Volcanism brings reduced rocks and gases to Earth’s surface. The vast outpourings of flood basalt provinces, doubling subaerial volcanic output for hundreds of thousands of years, may upset the normal balance of geochemical cycles and lead to massive short-term climate change. In the case of the Siberian Traps, the burning of coal would have represented an uncontrolled and catastrophic release of energy from Earth’s planetary fuel cell, as millions of years of accumulation of organic carbon were released and oxidized all at once, greatly perturbing the carbon cycle.
Plate tectonics has also had a substantial influence on biological evolution during the Phanerozoic. The process of evolution through natural selection leads to genetic differences as a result of time and separation. Because evolution follows different trajectories when ecosystems become isolated, the longer the time of isolation the greater the differences among organisms. Separation on Earth is largely caused by tectonic processes. On the largest scale, separation of continents on different plates can lead to tens of millions of years of independent evolution. We are aware of the differences among large mammals on the different continents, which are a consequence of this continental separation. The differences are most marked for Australia, which separated from Antarctica some 50 million years ago. Australian mammals can be traced to a common ancestor with other modern mammals, but after it became separated from other continents Australia gradually developed its own characteristic fauna, including “exotic and bizarre” species such as the emu, the duck-billed platypus, and the marsupial, kangaroo. In fact, owing to its tectonic isolation and unique climatic history, from 83% to more than 90% of its mammals, reptiles, insects, and amphibians are endemic to Australia (i.e., found on no other continent). A particular feature of Australian fauna is the relative absence of placental mammals and the abundance of marsupials. The marsupials likely flourished in Australia because of their low metabolic rate, which was suited to the hot and dry climate. Human import of other mammals, such as rabbits and domesticated animals, has led to extinction of many of the marsupials and other unique Australian fauna, as well as to significant changes in Australian ecosystems.
While continental isolation has the most pronounced influence on evolution, the displacement of continents and other tectonic events must also have been important. High mountain ranges produced by plate tectonics create diverse local environments and separate vast regions of very different climates. Often today there are distinct biota on westward- and eastward-facing slopes of major mountain ranges. The plate positions and the extent of their separation also influence ocean circulation and climate, including whether or not there are ice age cycles.
Glaciation can also have a significant impact on biodiversity. An ice sheet 2 km thick eliminates macroscopic species, leading to oscillating periods of radiation during interglacials and destruction during glacials. For this reason, glaciated regions have much lower diversity of species such as plants and birds as compared to regions that were unglaciated.
If all the continents were always together in a single supercontinent with a stable climate, with no large mountain ranges created by active plate convergence, then one set of species would dominate each ecosystem, and ecosystems could become stagnant, absent external catastrophes, with little pressure (or opportunity) for evolutionary change. Displacing the continents allows greater diversity to arise. When the continents rejoin, the most successful organisms will prevail. Having a dozen opportunities on separated continents to find the most successful evolutionary innovation would lead to greater evolutionary change over long timescales than on single continents where, though time may be vast, separation is limited.
We often consider life as somehow a separate phenomenon from Earth as a whole, and certainly isolated from planetary processes associated with Earth’s deep interior. The preceding chapters have shown some of the diverse linkages between life and planet. Can we extract principles from these interactions that might apply generally to a process of planetary evolution and would not necessarily be unique to Earth?
We discussed in Chapter 12 the differences between prokaryotic and eukaryotic cells, with the much larger eukaryotic cells being complex chemical factories compared to their prokaryotic cousins. Inside the eukaryotic cells are organelles, such as mitochondria and chloroplasts, that carry out energy production and photosynthesis. These organelles are believed to have formed by progressive symbiosis and incorporation of bacteria, called endosymbiosis, where symbiotic relationships of different species become specialized and permanent. This idea is supported by the fact that such organelles contain their own genetic material that is distinct from that of the eukaryotic cell nucleus and similar to bacteria that carry out similar functions. Chloroplasts, for example, contain DNA similar to cyanobacteria (photosynthetic prokaryotes). In addition, the structure and metabolism of the organelles are similar to prokaryotic cells, and organelles replicate by binary fission, similar to bacterial cell division. Notably, these endosymbiotic organelles are no longer able to survive independently outside the eukaryotic cell, and their survival depends on other aspects of the eukaryotic cell’s metabolism. This suggests a chain of events where early symbioses and incorporation lead to a partnership between initially separate organisms, and once the partnership is established, further evolution is based on interdependence.
This type of process is still evident today, as seen in the tubeworms and clams that flourish around hydrothermal vents and hydrocarbon seeps. The tubeworms are animals, but they have neither mouths nor stomachs. Instead, they have bacteria that live inside them—more than 250 billion bacteria per ounce of tubeworm tissue. The bacteria oxidize the sulfur using oxygen, and they employ this energy to reduce carbon dioxide to sugars, which feed the worm. The tubeworms have evolved to support the bacterial colonies on which they depend. For example, the red plumes of the tubeworms contain hemoglobin, which captures hydrogen sulfide as well as oxygen and transports these necessary raw materials to the bacteria living inside the worm. In this example we see a single organism whose metabolism has evolved to support an essential symbiosis. These cases demonstrate what may be a central aspect of evolutionary change, the tendency for living organisms to relate and develop symbioses that lead to more complex living machines with a more diverse set of processes and relationships.
The analogue to the development of eukaryotes from prokaryotic partnerships is mirrored in the development of multicellular from single-celled eukaryotic organisms, where individual cells become regulated and function together. Multicellular organisms in turn have gradually developed an increasing number of specialized cells. Human beings, for example, have some 75 trillion cells of 220 different types that carry out the various bodily functions, all working together to create a single functioning organism. The brain itself may be another example of such a developing network. There is little difference between the nerve cells that make up the brains of mice and human beings. The primary difference is the number of brain cells and the relationships among them. Increased relationship leads to increased capacity and ability to respond to diverse and changing environments. Relationships are also necessary to generate feedbacks, and increasing numbers of feedbacks lead to greater responsiveness and stability. And within species, it is clear that the relationship of working together often has advantages in terms of capability and survivability, such as the “superorganisms” of ant and bee colonies so well described by E. O. Wilson. Networks permit increased specialization of individual species, and complex feedbacks and networks lead to greater stability and survivability. Such trends are likely to be systemic and not restricted to Earth alone.
Energy is the driver of all systems, including living systems, and species that are able to harness more energy have a greater capacity to do work (e.g., chase prey or avoid predators), grow, and reproduce.
Increased access to O2 and hence aerobically derived energy is one of the characteristics of terrestrial evolution. All animals use oxygen to some degree (and many microbes do as well). As we saw in previous chapters, life progressively developed increasing access to aerobic energy through anaerobic photosynthesis, oxygenic photosynthesis, anaerobic metabolism, and ultimately aerobic metabolism. To take advantage of aerobic metabolism requires a steady supply of oxygen. For very small organisms, this can occur by diffusion and transport through the cell wall. Larger organisms need a means of active oxygen transport and efficiency of respiration, as well as active removal of waste products. The mitochondria organelles in eukaryotic cells carry out the aerobic metabolism and serve as specialized energy factories. The development of circulation systems and breathing eventually permits far more aerobic metabolism and a greater rate of energy production in multicellular life. Ultimately warm-blooded metabolism leads to an even higher metabolic rate. From this perspective the history of life involves a progressive increase in energy available for metabolic processes. Multicellular life cannot exist in anaerobic environments because there is insufficient energy production to support it. The triumph of mammals over reptiles would be a consequence of the higher mammalian metabolic rate and ability to generate energy independent of the external temperature environment. Ultimately this permits organs such as the brain to evolve. The brain is completely dependent on active oxygen transport. It neither makes nor stores food; its cells cannot survive on their own and require a vigorous blood supply to provide the active delivery of glucose and oxygen in order to function.
The change in energy metabolism during Earth’s history has parallels with the increase in relationship and complexity discussed in the previous section. Simple processes occur at the beginning, such as prokaryotic cells and anaerobic ATP production. Subsequent processes build upon the earlier developments. While some early innovations are discarded as they become superseded, others are integrated into the more evolved forms. The primitive forms also persist in their own ecological niches. Prokaryotes are everywhere, at the same time as the processes they invented have been co-opted by subsequent eukaryotic organisms. Multicellular life, for example, provides a vast ecospace for prokaryotic organisms—the skin, mouth, and digestive systems of human beings provide environments for trillions of prokaryotes of hundreds of species. There has thus been both expansion and inclusion. This principle applies to energetic processes as well. Within aerobic organisms, anaerobic metabolic processes have been integrated. For example, anaerobic metabolism processes ATP more rapidly than aerobic metabolism, so mammals use anaerobic metabolism for short bursts of high energy.
There is another important aspect to enhanced energy production. High energy production requires an interface between reservoirs that are out of equilibrium, since it is the passage from disequilibrium to equilibrium states that generates the energy for life processes. Access of organisms to increased energy requires the parallel development of energy potential through disequilibrium, and the formation and separation of oxidized and reduced planetary reservoirs—the planetary “fuel cell” discussed at length in Chapter 15. Enhanced energy utilization requires then a coupled change in biological mechanisms and planetary environment. Evolution is a planetary process that includes life, ocean, atmosphere, and rock.
A natural inference from this sort of discussion might be that life progressively evolves to organisms with increasing numbers of cells, an increased genome size, and more efficient energy processing, a progressionist view of evolution. This perspective is then often extended to human beings as the current apex of evolution. Very few biologists would subscribe to this view, however, because of the randomness inherent in the DNA mutation and the process of natural selection. A directional view is also difficult to justify on a quantitative biomolecular basis. Salamanders have an order of magnitude more DNA per cell than humans and other mammals. The rice genome encodes far more proteins than the human genome. The platypus cell has more chromosomes than the human cell. The elephant brain is four times the size of the human brain. On the broad scale of prokaryote to eukaryote to multicellularity to organ differentiation, there is a progression of increased relationship and complexity over the course of geological time, from which some progression might be argued, but the specifics of evolution and what makes an organism “advanced” are far richer and cannot be reduced to a simplistic directional framework.
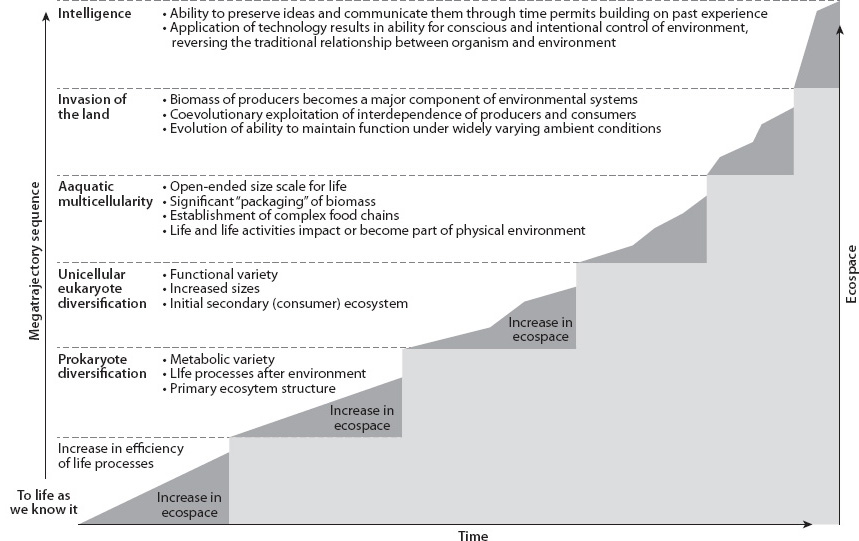
On the other hand, there is an apparent directionality to terrestrial life that Andy Knoll and Richard Bambach have called “an increase in utilization of ecospace” (Fig. 17-11). They point out five major stages of ecospace expansion within the biological realm after life began—prokaryotic diversification, unicellular eukaryote diversification, aquatic multicellularity, invasion of the land, and intelligent life. Each step expands from the previous ecospace by increasing the dimensionality of life through evolution of organisms of increasingly complex internal relationships. At the same time, each step can expand environments for life of the previous dimensions. While there may be no direction on the reductionist scale of individual DNA mutations, the observation is that there are directional attributes to biological evolution.
Fig. 17-11: Knoll and Bambach’s suggested “megatrajectories” of evolutionary stages through Earth history. (Knoll and Bambach, Paleobiology 26(2000)(sp4):1–14)
Directionality can occur from random change if there are external constraints that favor particular types of changes. Imagine, for example, a large collection of dice replicating in a box, and as they replicate there are periodic random changes in the numbers on the faces. Instead of having all the die have 1–6, the change could lead to a die with 1, 1, 3, 4, 5, 6 or 1, 2, 4, 5, 6, 6. These changes would be inherited for each succeeding generation. If there were no preference for any one of the numbers, and large numbers of dice, then despite the “mutations” there would be similar amounts of all the numbers in the box at any one time. But if we put in a “natural selection” such that some of the dice are less likely to be removed from the box depending on the number on their face, the distribution will change. The changes could all be random, but if for some reason the number 4 was favored through selection—dice with a 4 facing up were less often removed—eventually the dice would be almost all 4s. That result would be inevitable, despite the random process that led to it and the fact that the specific random steps in that direction would always be different in repeated experiments and could not be predicted.
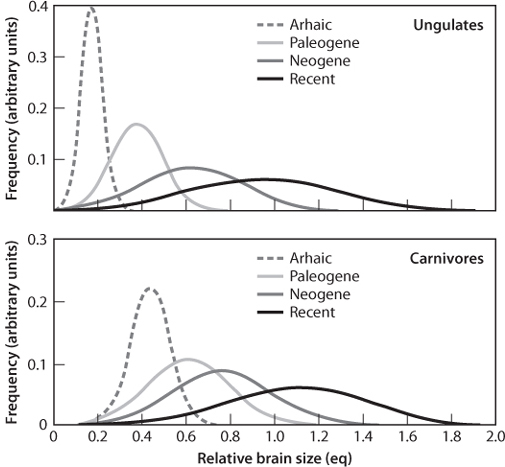
Can we imagine there might be such external constraints on evolution? One example might be the efficiency of energy processing, as discussed above. If organisms that are able to access more energy have a competitive advantage, this would inevitably lead to a directional change of increased energy usage. Changes that lead to increased system stability could also be favored, if increased stability is provided by the feedbacks that become possible with complex networks. And in the ongoing evolutionary battle between carnivores and their prey, the brain size of both has increased, suggesting there is an evolutionary advantage to larger brains (Fig. 17-12). Random genetic changes could lead to smaller or larger brains with no preference, but those changes leading to smaller brains would be selected against. Other examples might be bilateral symmetry or the development of increasing sensory systems to monitor one’s surroundings. Directionality then appears from selective advantage. Viewed in this context, directionality to evolution is possible within the context of random changes on the molecular level. The fact that random changes are the mechanism of evolution does not mean that larger-scale directionality could not emerge.
Fig. 17-12: Indications of directionality in evolution through the increase in brain size of both carnivores and their prey. (Modified from Radinsky, The American Naturalist 112 987 (1978): 815–83).
From such a perspective, a species capable of long-term forethought, enhanced energy use through tools and fuels, and the many advantages imparted by language would have great competitive advantages. These are all enhanced in modern civilization by global communications and transportation networks, genetic modifications for species advantage, and potential interplanetary travel. The emergence of intelligent life from the perspective of energy and networks could be a natural consequence of planetary evolution.
This discussion builds upon the factual observation that Earth has become increasingly habitable over time. Life has created enhanced environments for life. Life has participated in major changes of the planet that have led to a far greater diversity of organisms today than existed in the past, and as multicellularity and organ specialization came into being, to greater complexity. As concrete examples of increased habitability:
(1) no multicellular animal would be able to survive on Earth in the Precambrian, though microbes of that era might be extant in today’s ecosystems;
(2) it is likely that the mass of life on Earth today is far greater than it was during the Archean and Proterozoic;
(3) the amount of energy flowing through ecosystems is certainly greater.
For example, prior to oxygenic photosynthesis, the capability of the biosphere for transforming energy from the sun was quite limited. Hand in hand with the evolution of the species has been an evolution of habitability. Increased habitability is of course closely coupled to changes in the planetary environment. The changes in atmospheric composition, ocean chemistry, expansion of life from ocean to land, the development of soils—all of these create enhanced environments for life. Production of organic matter provides an energy source that supports more complex food webs and—eventually—more energy-intensive organisms such as mammals. Life provides and expands the environments for life.
We can ask why such developments have taken place. Why should random DNA mutations lead to an increasingly habitable planet? On many scales one can imagine that this is a natural process of planetary evolution. If there are two competing organisms, and one of them makes an environment more hospitable for its development, or for other organisms that are beneficial companions, it will preferentially survive. And if a network of organisms creates an environment more capable of sustaining life and processing energy—i.e., more habitable—this network will have an evolutionary advantage. Organisms that make the environment less habitable will ultimately fail. One of the natural consequences of a planet with life may be a progressive increase in planetary habitability, provided the external conditions for life can persist.
Evolution of habitability also can take place only if life is able to respond to inevitable planetary change. Many aspects of planetary evolution are long and slow, such as the rise of O2 and the development of cellular mechanisms to take advantage of it. This requires persistent life. And in the midst of this slow evolution, there are catastrophes from the cosmos and planetary interior and inevitable change of environmental conditions. For life to continue it must be adaptable enough to survive, and even better if it can make opportunity out of crisis. Absent such capabilities, life would become extinct and the planet uninhabited. This suggests another principle, that biological evolution—the capacity for change—is a prerequisite for planetary habitability.
If we consider life as a planetary process, rather than some separate phenomenon that occurs on a planetary surface, then planetary evolution includes all aspects of the planetary system—core, mantle, crust, ocean, atmosphere, and life. Life plays a critical role in planetary evolution, by capturing and storing solar energy in planetary materials, modifying the oxidation states of near-surface reservoirs. Life is in turn greatly influenced by the physical processes of planetary and solar system evolution. Its beginning was influenced by the late heavy bombardment, and meteorite impacts have continued to have an important influence during the Phanerozoic. The convective processes of Earth’s interior influenced the origin of life and climate stability, and surface expressions of active convection through plumes have likely caused many of the mass extinctions and periods of extreme climate change that punctuate the Phanerozoic record.
While biological evolution results from random changes in the microscopic DNA code, there is nonetheless progressive evolutionary change. Over the course of Earth’s history, organisms have developed increasing networks and complexity, both internally and externally, and have greatly increased their energetic productivity. Directionality can emerge from random change if there are underlying principles that supply a competitive advantage. From the Phanerozoic record, it appears that the progressive changes in life are in turn facilitated by the catastrophes coming from solar system and planetary interior. These processes destroy long-term stability and permit evolutionary innovations to be expressed in a newly level playing field. Plate tectonics and climate change have also enhanced evolutionary change, by permitting separate and simultaneous evolutionary experiments and creating variable environments. Through planetary evolution, Earth has become increasingly habitable.
While Earth’s history is inevitably specific as a story of one planet, principles that it embodies appear likely to apply on a universal scale. Evolution by natural selection is a general process clearly not restricted to a particular time or place. Increased stability through networks and increased access to and utilization of energy are also a kind of thermodynamic driver that would be guiding principles on any planetary body. Periodic instabilities from the solar system and planetary interior are inevitable consequences of solar system formation and planetary convection, creating the crises that destroy stasis and permit rapid evolutionary change. From this perspective, evolution toward increased habitability may be a common planetary process.
Andrew H. Knoll and Richard K. Bambach. 2000. Directionality in the history of life: Diffusion from the left wall or repeated scaling of the right? Paleobiology 26(4):1–14.
Bert Hölldobler and E. O. Wilson. 2009. The Superorganism: The Beauty, Elegance, and Strangeness of Insect Societies. New York: W. W. Norton & Co.
Walter Alvarez. 1997. T. Rex and the Crater of Doom. Princeton, NJ: Princeton University Press.
Douglas H. Erwin. 2006. Extinction: How Life on Earth Nearly Ended 250 Million Years Ago. Princeton, NJ: Princeton University Press