The History of Evolutionary Thought
Garland E. Allen
OUTLINE
1. Species and the origins of diversity in the ancient world
2. The eighteenth century and ideas of the transmutation of species
3. The rise of natural history
4. The development of geology in the late eighteenth and nineteenth centuries
5. Ideas of transmutation of species before Darwin
6. Charles Darwin and On the Origin of Species (1809–1859)
7. Post-Darwinian controversies and the “eclipse of Darwinism,” 1890–1920
8. Heredity and evolution: Mendelism, Darwinism, and the “evolutionary synthesis”
9. Evolutionary theory in the era of molecular biology
GLOSSARY
Catastrophism. Geological theory that the earth’s surface was transformed by a series of great catastrophic events in the past, such as floods and massive volcanic eruptions, on a much greater scale than at present.
Evolutionary Synthesis. A term usually applied to developments in evolutionary theory between roughly 1930 and 1950 or 1960, and characterized by the union of Darwinian evolutionary theory with Mendelian genetics (as population genetics), taxonomy, and paleontology.
Population Genetics. The study of populations in terms of gene frequencies and their changes over time, based on mathematical and statistical treatments of interbreeding groups of organisms involving field studies, model building, and experimentation on laboratory populations.
Transformation (Transformism or Development Hypothesis). Older terms for what later (post-1860) became known as evolution, or descent with modification.
Uniformitarianism. Geological term associated with James Hutton and later Charles Lyell to account for the formation of features of the earth’s crust by slow, everyday forces such as erosion, and pressure of the oceans against continental boundaries.
1. SPECIES AND THE ORIGINS OF DIVERSITY IN THE ANCIENT WORLD
While philosophers and naturalists in the ancient world did not have a concept of “evolution” in the modern sense, certain traditions or schools of thought in Greece and Rome developed ideas about the origins of biological diversity by natural, as opposed to supernatural, processes. The basic idea that living organisms of one kind can become transformed into living organisms of another kind has its roots in the works of the Greek philosopher Epicurus (341–270 BCE) and his school (the “Epicureans”) in Athens. Epicureans were philosophical materialists who believed the world was composed of small particles, or atoms, that were continually in motion in otherwise empty space. By bumping into each other randomly, atoms produced all the physical and chemical processes we observe in the world. One consequence of this general view was that the world was seen to be continually in flux and change; nothing was static or permanent. One of the Epicureans most influential on the development of later European philosophy and natural history was the Roman poet Titus Lucretius Carus (99–54 BCE). In his long poem “De Rerum Naturae” (On the Nature of Things) Lucretius argued forcefully against supernatural explanations of nature and human life, in either the creation of the world or its daily operations. According to Lucretius, nature is purposeless, occurrences are traceable to the random action of atoms, and death is the end of being. Lucretius, along with other Epicureans, was opposed to all forms of established religion. The legacy of the Epicureans is thus a belief in the inevitability of change, a rejection of supernatural causes, and a search for naturalistic explanations. This view would eventually be revived in various forms during the scientific revolution of the seventeenth and eighteenth centuries, and would play an important role in thinking about evolutionary transformation from that period into the nineteenth century.
The materialism of Epicurean philosophy was not the dominant view, however, in either ancient Greece or Rome. Far more pervasive was the idealist (or nonmaterialist) philosophy of Plato (and later Aristotle), which saw the universe and beings within it as creations in the mind of God and had no place for large-scale developmental change. The Platonic universe was based on the distinction between ideal forms, or categories existing in the mind of the Creator, and real, material forms that existed on earth. These categories represented the essence of the forms, which were universal, permanent (immutable), and perfect. For example, the abstract category of “catness” would include all the essential characteristics of what made an animal a cat rather than a dog or horse. While recognizing the existence of variation among real-life cats, Plato’s conception focused attention on an idealized world of stable forms or categories (of which “catness” was just one example). The Platonic philosophy is referred to as “idealist” because it was centered on the abstract, nonmaterial world of forms and categories rather than on attention to detailed observations.
Aristotle espoused many of Plato’s basic philosophical views, but as a naturalist he paid much closer attention to empirical observations, as his remarkable studies on chick development clearly show. Yet, two aspects of Aristotelian thought provided challenges to later evolutionary thinkers. One was teleological (goal-oriented) thinking: the view that changes in the world are always directed, as in embryonic development, toward a fixed or final goal. Another was the idea of purposefulness, in which the goal also has a purpose, a function, or an adaptation, as in wings as organs of flight. Aristotle also made a distinction between final and efficient causes. Efficient causes are the immediate factors that lead to some particular outcome (in embryonic development, the fertilization of an egg leads to the development of an adult). Final causes are the answers to the long-range questions, why or for what end? The final cause leading to embryonic development is the teleological purpose of forming a completely new individual. By extension, all nature was seen as directed toward purposeful ends. The notions of fixed categories of beings, teleological processes, and purposeful organization of the natural world were thus important legacies of Greece and Rome with which later naturalists had to grapple.
There was little that could be thought of as evolutionary thinking in the period from the fall of Rome in the fifth century through the medieval and early modern (sixteenth to seventeenth) centuries, owing partly to the rise to political power of the Roman Catholic Church and other sects of Christianity that held to an interpretation of the origin of species conforming with the biblical story of creation. In this view, species had been specially created by God and were fixed in their original forms in compliance with the generally pervasive Platonic concept of essences. There matters stood until the consequences of the scientific revolution of the seventeenth century began to be played out in natural history in the eighteenth century.
2. THE EIGHTEENTH CENTURY AND IDEAS OF TRANSMUTATION OF SPECIES
What has been called the scientific revolution (roughly 1500–1700) centered on problems of astronomy and dynamics (the science of motion) and grew out of a series of commercial (navigation and calendar reform) and intellectual concerns about the structure and function of the universe, especially the solar system. Building on the work of Nicolaus Copernicus (1473–1543), which centered the universe on the sun rather than the earth, his successors in the seventeenth century, Johannes Kepler (1571–1630), Galileo Galilei (1564–1642), and Isaac Newton (1642–1727), among others, showed that the universe, including events on earth, could be understood as natural “laws” that led to regular and predictable outcomes. God may have started the clockwork of the universe, but it functioned according to its own laws from then on. Organic life, some argued, could also be understood as subject to laws.
A second development, during the middle and later eighteenth century, especially in conjunction with economic developments such as mining, road building, and the growth of industry, was the rapid development of geology. Mining and quarrying for stone (for building), for coal and metal (for a burgeoning weapons industry), and for road and canal construction exposed new layers of the earth’s crust, greatly extending the estimates of the earth’s age. The discovery of numerous fossils, some similar to and some very different from organisms living on earth today, suggested that life also had a long and varied history. A third development was a result of the economic, social, and political changes surrounding the French Revolution of 1789. This immense upheaval suggested that no established practices, including the supposedly divinely given power of monarchy and the authority of the church, were immutable. These developments sparked a new materialism and antireligious sentiment, emphasizing that the world in all its aspects was subject to change and flux; nothing was fixed and permanent. These views gained considerable exposure in the works of French philosophes such as Paul-Henri Thiry, Baron d’Holbach (1723–1789), and the encyclopedist Denis Diderot (1713–1784).
One of the English followers of the philosophes was the physician and savant Erasmus Darwin (1731–1802), the grandfather of Charles, whose poetical works such as Zoonomia; or the Laws of Organic Life (1794–1796) contained many protoevolutionary ideas: for instance, a constantly changing, dynamic nature, a “world without end.” Although none of these developments led to a fully explicit view of evolution in the later nineteenth-century form, they emphasized change and transformation as natural and eternal and thus provided a context for thinking about how species might become transformed by natural causes.
3. THE RISE OF NATURAL HISTORY
Natural history, especially compared with physics and the exact sciences, had been held in low esteem in the sixteenth and seventeenth centuries. But with the expansion of colonial trade, natural history as a source for new crops and commodities began to assume greater importance. As vast numbers of new forms were brought back by commercial voyages, the diversity of the biological world became a matter of great interest and study. New forms from exotic geographic areas were placed in a kind of linear scala naturae (“ladder of nature,” or “chain of beings,” as it was called), ranging from simplest microorganisms to humans. The scala naturae was based on the idea that every organism fit into a place in the natural order, with forms slightly simpler below it and more complex above it. Theoretically, there should be no major gaps in the scala naturae because, following an old principle, nature is a continuity. The scala naturae presented one view of order in the natural world, but it was fixed and static: organisms did not move up or down the ladder, and they did not fall into natural groupings.
A far more useful framework was developed by Swedish botanist Karl von Linné (in Latin, Carolus Linnaeus) (1707–1778). Linnaeus believed that all plants and animals had been created by God and that they could therefore be placed into natural groups that revealed God’s divine plan. Whereas previous groupings or classifications had been based on utilitarian principles, Linnaeus sought a system that would group organisms by their natural or structural characteristics, such as numbers and kinds of flower parts, or reproductive organs. Linnaeus’s large-scale classification system, including both plants and animals, appeared in the 10 editions of his treatise, Systema Naturae (1735–1758). His system involved a hierarchical series of categories—with kingdom as the most inclusive, through phylum, class, order, family, genus, to species as the most specific—that included all organisms of the same kind. He also devised a binomial system for naming organisms by their genus and species names, such as Canis familiaris (common dog) or Acer rubrum (red maple). The important feature of Linnaeus’s system was that organisms grouped by one set of fundamental characteristics would also share other common structural and functional features, at whatever level of the hierarchy one focused. Whether this order originated in God’s divine plan or somehow emerged by natural laws (causes) was debatable, but the existence of the Linnaean order was well established by the end of the eighteenth century.
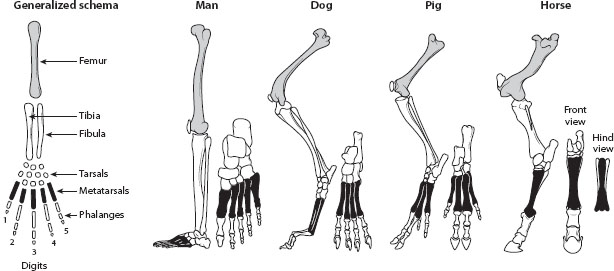
One of the most important concurrent developments in natural history was the rise of comparative anatomy, particularly as espoused by the French anatomist and paleontologist Georges Cuvier (1769–1832). He became familiar with hosts of fossil organisms unearthed during much of the excavation and building projects associated with the Napoleonic era in France and elsewhere. Most of these organisms somewhat resembled modern forms, but many seemed unique. Cuvier studied them, comparing various structures, such as leg bones or skulls. In this manner, Cuvier and others noticed that most organisms in the same Linnaean category showed structures that were modifications of the same basic plan—as in the front limbs of vertebrates—and that were later referred to as homologies by the British anatomist Richard Owen (1804–1892) (figure 1). Cuvier also emphasized the close correlation of parts in the anatomy of any organism, arguing that all the parts existed in complete harmony; no part could be changed without affecting all others. Therefore, it seemed impossible to Cuvier that any species could be modified and transformed into another. Despite founding the field of comparative anatomy, Cuvier viewed it as only a reflection of the divine (and immutable) order of the universe.
Figure 1. Comparison of the hind appendages of four vertebrates, showing the homologous relationships among the bones. All the femurs are homologous to each other, as are the tibia, fibula, and tarsals; metatarsals; and phalanges. It is evident the same bones have been modified for quite different functions over time. Such comparative homologies provided strong evidence for evolution, as descent with modification from a common ancestor. Special creationists would have to argue that the Creator repeatedly used the same archetypal form, as in the generalized appendage on the far left, for all vertebrate appendages. (From Jeffrey J. W. Baker and Garland E. Allen. 1982. The Study of Biology. Reading, MA: Addison-Wesley, 671; used with permission of the authors.)
4. THE DEVELOPMENT OF GEOLOGY IN THE LATE EIGHTEENTH AND NINETEENTH CENTURIES
With the rapid expansion of building, mining, and various forms of excavation between 1790 and 1830, much was being learned about the structure of the earth and (especially) its fossil contents. The succession of strata told a story of continual changes in the earth’s surface: submersion, elevation, erosion, further submersion, and so on. Some layers were clearly produced by volcanic action in what appeared to have been massive upheavals; others were produced by flooding and underwater deposition of mineral contents. One consequence of these observations was the growing view that the earth was considerably older than the 4004 BCE creation date claimed by Archbishop James Usher. It now appeared that the earth was hundreds of thousands or even possibly millions of years old. One school of thought, catastrophism, explained the various upheavals observed in the geological record by a large-scale series of ancient worldwide floods or volcanic eruptions. That these forces were no longer operating to the same extent as in the past fit well with the prevalent theory that the earth had been formed as a spin-off from the sun and had thus originally been very hot but by then had cooled considerably (the “cooling earth theory”).
Another school of geology, developed in Britain by James Hutton (1726–1797), emphasized a process called uniformitarianism. Hutton claimed that changes in the face of the earth came about in the past by the same sorts of gradual processes observed on the earth at his time and did not require the supposition of vast cataclysms in the past. Hutton’s ideas were taken up by a younger geologist in the early nineteenth century, Charles Lyell (1797–1875), who published a three-volume treatise, The Principles of Geology (1830–1831), which provided the most influential argument for uniformitarianism. Lyell showed that it was the constant, daily action of wind and water that eroded the land, and the pressure of the sea against the continents that formed and elevated mountains. These processes operated very gradually, almost imperceptibly, yet over long periods of time could have dramatic effects. Uniformitarianism fit well with the positivist philosophy of science, which argued that the best theories required the simplest assumptions and sought the most general and constant laws. Some historians have also argued that uniformitarianism gained its popularity in the wake of the French Revolution, which was claimed to have demonstrated that cataclysmic change was unnatural.
Uniformitarianism posed one problem, however: for vast changes in the earth’s surface to have occurred so slowly the age of the earth must be far greater than calculated by earlier estimates. This incongruity raised problems not only with geologists’ methods for estimating the age of the earth but also with biblical orthodoxy. The question of the earth’s age was to continue to be highly controversial throughout the nineteenth and much of the twentieth centuries.
5. IDEAS OF TRANSMUTATION OF SPECIES BEFORE DARWIN
The idea that species can change in various ways was not original with Darwin but had a variety of incarnations between 1750 and 1850. Erasmus Darwin’s idea that species were not fixed was but one of many prevailing theories of transformation in the late eighteenth and early nineteenth centuries. An author more influential with naturalists was Georges Louis Leclerq, Comte de Buffon (1707–1788), whose massive Histoire naturelle (Natural History) began to appear in 1749. A committed Newtonian and staunch materialist, Buffon sought natural explanations for all phenomena. Opposed to Linnaeus’s view of an abstract, formalistic view of nature as categories, Buffon argued that organisms in nature existed as an array of species and that some individuals could be intermediate in form and function between two species. He developed a theory of degeneration to account for at least the process by which transformation of species could occur within a broad group, such as the family Felidae (cats). The original ancestor could have had the mane of a lion, the stripes of a tiger, and the size of the current large cats. By degeneration, various descendant groups lost one or more of these characters, becoming modern lions (no stripes), tigers (no mane), leopards (no mane, broken stripes), and house cats (small size). Although highly speculative, Buffon’s system did promote in a limited way the idea of species transformation by natural processes. Histoire naturelle was one of the most influential natural history works of the eighteenth century.
One of the most prolific figures in the period before Darwin was the French naturalist Jean-Baptiste Pierre Antoine de Monet, Chevalier de Lamarck (1744–1829). Lamarck was one of the first naturalists to develop a full-fledged theory of transformation that also provided a mechanism for how organisms could become adapted to their environment. Although Lamarck used many examples in his writings, principally his Philosophie zoologique (Zoological Philosophy) of 1809, the development of the giraffe’s long neck has been the one most often repeated. Lamarck postulated that as ancestral giraffes ate leaves from lower branches of trees they continually stretched their necks to reach the leaves higher up. The increase in a giraffe’s neck during its own lifetime was passed on to its offspring, through the inheritance of acquired characters. Although many people doubted Lamarck’s example, the underlying mechanism of the inheritance of acquired characters remained a well-accepted concept throughout the nineteenth (and well into the twentieth) century.
Lamarck’s theory of transformation was not what we would recognize as evolution in the modern sense. Although he came to believe in the mutability of species, Lamarck did not maintain that various organisms are related to one another through branching lineages. Rather, he saw simple organisms as continually being generated spontaneously and then gradually becoming transformed by their environment and their own internal drives into more complex forms. For Lamarck, the organism “willed” itself to change and adapt to new conditions and thus was an active participant in its own transformation. More often than not it was this aspect of Lamarck’s theory, and not the inheritance of acquired characters, that brought his claims about species transformation into disfavor.
That many naturalists and writers about natural history were beginning to think that species transformation was a reality is evidenced by the immense popularity of a book written in 1844 by the Scottish publisher Robert Chambers (1802–1871) and published anonymously. The Vestiges of the Natural History of Creation was a sensational account of the continuous development of life on earth, based on a notion of inevitable progression toward higher, better-adapted forms, including humans. In support of his idea of progressive, goal-directed transformation of species Chambers paid particular attention to the fossil record and what he saw as its parallel in embryonic development. Although he left a place in his process for the Creator (who not only started the whole process but also periodically stepped in to formulate new “laws” of development), Chambers was universally labeled a “materialist,” one of the most damning epithets in Victorian culture. Perhaps in part owing to the controversial elements, Vestiges was immensely popular and went through numerous editions, to each of which Chambers added new arguments to offset criticisms from both naturalists and theologians. While Chambers in no way presented the idea of evolution in the same vein and with the same attention to detail as Darwin, Vestiges did in fact provide an imaginative and fascinating synthesis of geology, paleontology, and natural history that supported the idea of transmutation of species.
6. CHARLES DARWIN AND ON THE ORIGIN OF SPECIES (1809–1859)
Early Education and Influences
Charles Robert Darwin (1809–1882) was born into a wealthy upper-middle-class family in Shrewsbury: his father was a successful physician and his mother a daughter of Josiah Wedgwood, of pottery fame. Young Darwin originally attended Edinburgh University to study medicine like his father but was nauseated at dissections and the sight of blood, so he abandoned these plans. Intending to prepare for the ministry, he matriculated at Christ’s College, Cambridge, in 1827 but soon realized he did not have the requisite religious convictions for this occupation. Although directionless in the conventional sense, Darwin did show an early interest in natural history. At Edinburgh he met and became friends with the comparative anatomist Robert Grant (1793–1874), who introduced him to current trends in continental comparative anatomy, having studied with Cuvier and Étienne Geoffroy Saint-Hilaire (1772–1844). Grant openly espoused a notion of transmutation of species, which he had gleaned from both Geoffroy and Erasmus Darwin’s Zoonomia.
At Cambridge, Darwin was strongly influenced first by the writings of William Paley (1743–1805) on natural theology, the view that the study of nature in all its manifestations was a way to understand the glory of the Creator. Paley argued in his book Natural Theology (1802) that if one found a watch lying on a heath, it would be clear that it must have had a Creator, and that by studying the watch it would be possible to learn something of the Creator’s mind. Similarly, the marvelous adaptations of animals and plants in nature indicated to Paley the existence of a Creator, who produced the most perfect adaptations. Darwin was also strongly influenced by personal contact with two Cambridge professors, the botanist John Stevens Henslow (1796–1861) and the geologist Adam Sedgwick (1785–1873). Both introduced him to organized fieldwork and encouraged him in his natural history inclinations. Darwin considered an extended field trip with Sedgwick to Wales in the summer of 1831 seminal in teaching him how to think scientifically. In these early years Darwin was as much a geologist, in interest and experience, as he was a naturalist. Through his contact with both Henslow and Sedgwick, Darwin began to learn about the vital importance in science of balancing detailed observations with broad-based causal reasoning.
Henslow was to play an even more prominent part in Darwin’s life than introducing him to methods of investigation in natural history. Shortly after Darwin graduated in the spring of 1831, Henslow received a request from the British Admiralty to recommend someone to serve as a naturalist and companion to Captain Robert FitzRoy (1805–1865) on the HMS Beagle, a hydrographic survey vessel that was to embark on an around-the-world trip later that year. For family reasons Henslow had to decline, and eventually the invitation was extended to Charles. Robert Darwin was at first opposed, considering it dangerous, and thinking it would not lead to any sort of useful career beyond the voyage itself. After intercession by his uncle Josiah Wedgwood, however, Charles gained his father’s permission and set off with FitzRoy on the Beagle on December 27, 1831. The ship was instructed to collect data about water depths, currents, and climatological and meteorological information as well as to make commercial contacts that would promote British trade throughout South America and the Pacific.
The Beagle Voyage
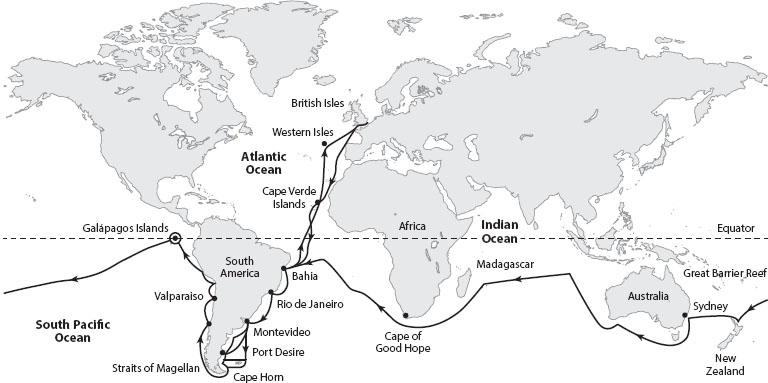
Figure 2. Path of the Beagle voyage between December 1831 and October 1836. The voyage completed an extensive around-the-world voyage, exposing Darwin to environments as different as the Canary Islands, the east and west coasts of South America, the Galápagos archipelago, the South Seas, and Australia. (From Jeffrey J. W. Baker and Garland E. Allen. 1982. The Study of Biology. Reading, MA: Addison-Wesley, 676; used with permission of the authors.)
Originally planned to last two years, the voyage extended to five, traversing the Atlantic and Pacific Oceans, and returned to England on October 2, 1836 (figure 2). Darwin took only two books with him initially: the first volume of Lyell’s Principles of Geology (he had the others sent as they were published) and a new edition of John Milton’s Paradise Lost, with its highly romanticized engravings by John Martin (1789–1854). Historian David Kohn has noted the significance of this choice in foreshadowing Darwin’s own loss of innocence during the voyage about the fixity of species (a metaphoric journey out of Eden) as a result of his experiences during the voyage.
A number of observations of organisms in a variety of geographic settings got Darwin to thinking about the nature of species and their origin. First, reading Lyell in conjunction with his own personal observations of geological formations in South America made Darwin a convinced uniformitarian. Second, in South America he found the fossil of a large, extinct ground sloth (Megatherium) in a chalk cliff while in the forests above he observed present-day living tree sloths; Darwin wondered if the latter could have originated by modification of the former. Third, as the Beagle traversed many geographic regions, and as Darwin had frequent opportunities to collect organisms on land, he noted that there were certain patterns in the geographic distribution of species. For example, the fauna and flora of Tristan da Cunha, an island halfway across the southern Atlantic between Africa and South America, were a mixture of those found on the two continents. Darwin also noted that the forms inhabiting islands off the coast of larger land areas were similar to, but modified from, the forms on the mainland. He concluded that migrations had played a significant part in shaping the geographic distribution of life on earth. Fourth, a seminal two weeks spent on the Galápagos Islands, some 700 miles off the coast of Ecuador, provided Darwin with crucial examples of the differences between related species on adjacent islands. Alerted to the fact that the populations of giant tortoises were recognizably different from island to island, Darwin noticed that this phenomenon also held true for other species, such as the finches and mockingbirds. It had become clear to Darwin by this point in the voyage (1835) that the only way to explain these various observations was descent with modification from common ancestors. At the time, however, Darwin had no mechanism for such modifications to occur.
Genesis of a Theory and Publication of On the Origin of Species
Returning home in 1836, Darwin began writing up his notes and observations from the Beagle and working up his specimens. He became immediately acquainted with a number of leading British naturalists in London, including Lyell and the comparative anatomist Richard Owen (1804–1892), and began to frequent major scientific circles. He was elected a Fellow of the Royal Society in 1839. In 1837 he opened a series of notebooks, two of which (the B and C Notebooks) he labeled “Notebooks on the Transmutation of Species.” In these he jotted down his ideas, reading notes, and observations about the possibility of species change. Several lines of evidence were important to Darwin’s thinking at this time. One was the ubiquitous nature of variation among organisms of the same species. These variations were often minute, “almost imperceptible,” as Darwin called them. He saw these differences as more important than the supposed “essential” nature of a species. The biologist and historian Ernst Mayr (1904–2005) argued that Darwin was at this time beginning to move away from a typological to a population view of species, in which variation, rather than the fixed type, became the focus of the naturalist’s attention. A second line of evidence came from animal and plant breeding. For centuries plant and animal breeders had been able to produce widely divergent forms, such as fancy breeds of pigeons (Darwin himself bred pigeons), dogs, cattle, or plant crops by selecting small variations over many generations. Breeders had shown species to be malleable, and to Darwin this suggested that given enough time, full transmutation was possible. It was a gradual process, but as with Lyell’s uniformitarianism, great changes could accrue from the buildup of many small events. At this time, however, Darwin knew that without a basic mechanism in nature, analogous to the actions of the animal and plant breeders, his theory lacked plausibility.
Then, in September 1838, Darwin read “for pleasure” Thomas Robert Malthus’s (1766–1834) An Essay on the Principle of Population, originally published in 1798 and by 1838 in its sixth edition. In this work Malthus, a clergyman and professor of history and political economy at East India Company College, Hertfordshire, put forward his “law” of population: whereas populations grow exponentially, their food supply grows arithmetically, creating constant shortages and thus competition for resources. Written in the immediate wake of the French Revolution, Malthus’s essay attempted to show that shortage of resources was not a function of economic and social policies but was inherent in population dynamics. As a naturalist, Darwin immediately recognized that in any species, far more offspring are born than survive to reproductive maturity, thus generating a Malthusian “struggle for existence.” The slight variations among members of the same species might give one individual an advantage over another in gaining food or a mate, or avoiding a predator. Those individuals with slight advantages would be expected, on average, to reproduce a little more successfully than others and thus leave more offspring (and vice versa for those individuals with less favorable variations). Assuming, as Darwin did, that many of these variations are inherited, the characteristics of the population would gradually change over time, leading eventually to formation of a new species. Darwin prepared two privately circulated essays, in 1842 and 1844, outlining his overall theory. His hesitancy to publish may have resulted from fear of negative reactions from both scientific colleagues and the public, especially his wife and the church. He also may have wanted to establish his reputation as a solid naturalist by publishing additional technical works, including a theory of coral reef formation and a detailed taxonomic reorganization of the family Cirripedia, the barnacles.
Everything changed when, in the spring of 1858, Darwin received a paper from a young naturalist working in the Malayan archipelago, Alfred Russel Wallace (1823–1913), outlining virtually the same theory of natural selection that Darwin had proposed. It was perhaps no accident that Wallace came up with the same ideas as Darwin, since Wallace, too, had read both Lyell and Malthus and recognized the dramatic role that competition and selection could play in species transformation. Darwin immediately consulted his colleagues Lyell, Thomas Henry Huxley (1825–1895), and Joseph Dalton Hooker (1817–1911), who suggested that one of Darwin’s earlier essays and Wallace’s paper should be read jointly at a meeting of the Linnaean Society of London that summer. Published in the society’s bulletin, these papers established the dual priority of both men, but it also spurred Darwin to publish, on November 24, 1859, his main work, On the Origin of Species by Means of Natural Selection, or the Preservation of Favoured Races in the Struggle for Life.
On the Origin of Species: Components of the Darwinian Paradigm
Ernst Mayr argued that Darwin’s formulation of the theory of evolution is not a single theory but, rather, a composite of five different theories (thus it functions as a paradigm in Thomas Kuhn’s sense). (1) Evolution as Such (descent with modification): As already discussed, by 1859, transmutation was more widely accepted among naturalists, and although controversial, it was not seen as implausible. (2) Common Descent: The idea that current forms have descended from common ancestors by a process of divergence, as in the various finches or mockingbirds of the Galápagos. Common descent as a general principle, found nowhere else in the writings of earlier transmutationists, and one of Darwin’s most original ideas, was readily accepted by his contemporaries for its power to explain phenomena such as geographic distribution and homologous structures (those that result from modification of the same basic structures—as in the bones of the hand of the human, the foot of the dog, and the wings of the bat or bird—leading to varied functions). (3) Gradualism: The view that evolution occurs in very slow, small steps and not by sudden, large-scale changes (called “sports” or “monstrosities” in Darwin’s day). Gradualism was also one of Darwin’s more original concepts. (4) Multiplication of Species: Common descent and divergence lead to an endless multiplication of species over time as new forms replace old ones that eventually become extinct (figure 3). (5) Natural Selection: Factors of the environment select for favorable and against unfavorable variations, in analogy with the work of the practical breeder (see chapter VIII.5). The difference is, of course, that breeders select for variations desirable for their own purposes (such as higher productivity), while natural selection has no ulterior purpose, no goal for improvement. It provides only for how variations are selected for or against in a particular environment at a particular time. Although natural selection was clearly one of Darwin’s most important contributions to the transmutation paradigm, the idea had been put forward earlier by several writers in the 1830s, including William Charles Wells (1757–1817), Patrick Matthew (1790–1874), and Edward Blyth (1810–1873), However, none of these writers made a case for the operation of selection as a general process in the way that Darwin did. Natural selection, as the mechanism for how transmutation occurs, was the most controversial component of Darwin’s paradigm.
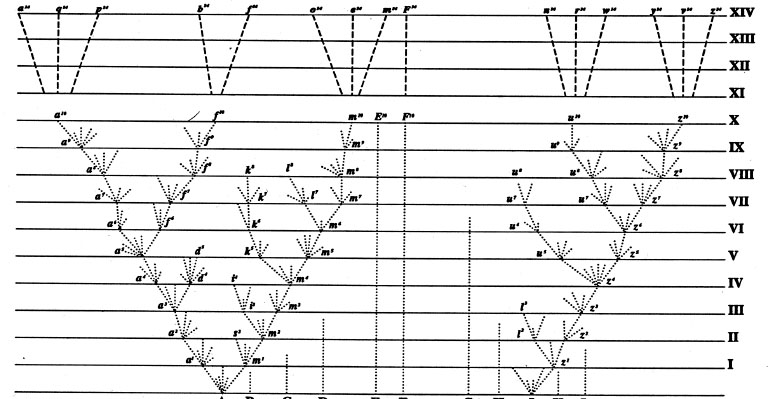
Figure 3. Darwin’s diagram in The Origin, showing divergence from common ancestors. Horizontal lines represent time periods, from most remote (I) to most recent (XIV). Darwin was aware that most lineages go extinct, as evidenced by the lines that end at a certain point in time. (From Charles Darwin. 1859. On the Origin of Species by Means of Natural Selection, or the Preservation of Favoured Races in the Struggle for Existence. London: John Murray.)
One of the most important problems that Darwin faced, and never properly resolved in his lifetime, centered on the nature of heredity, particularly the origin of variations. Like many of his contemporaries, Darwin believed in blending inheritance, the idea that the contributions of each parent for most traits were blended as an intermediate form in the offspring. This meant that new variations would tend to be diluted and less likely to be maintained in the population unless they were highly selected. Darwin did recognize that some traits were inherited in an all-or-nothing way, and these could both skip generations and revert to the original ancestral trait in future generations. On the origin of variations, Darwin held two views. Many variations, he thought, were induced by the environment—for example, food, moisture, temperature—or resulted from the effects of use and disuse, and these could be passed on to the offspring (a kind of Lamarckian inheritance). He also held that many variations occurred by chance, or spontaneously, and were not directly attributable to a specific factor in the environment. Although Gregor Mendel (1822–1884) published his studies on hybridization in 1866, neither Darwin nor most of his contemporaries were aware of this work, which did not come to general scientific attention until after 1900.
The Reception of On the Origin of Species
When The Origin was released in 1859, all 1250 copies were immediately sold out, either to subscribers or through booksellers. It became the center of much discussion in both scientific and lay circles. Public reaction was largely negative, repelled by Darwin’s seemingly materialistic worldview. Theologians were appalled, with one referring to Darwin as “the most dangerous man in England.” While some younger scientists, such as Huxley and Hooker, became ardent supporters, the majority of established scientists found the paradigm seriously flawed. (1) A major objection was that as breeding work showed, no new species had ever been produced by selection: although selection had been practiced with dogs for hundreds if not thousands of years, terriers, collies, and Great Danes were all still dogs and could freely interbreed. (2) Blending inheritance meant that new variations, even favorable ones, would hardly ever have a chance to become established except under extremely high levels of selection. (3) While Darwin’s theory of slow, incremental change should produce a fossil record with many intermediate forms, the actual strata showed large gaps and discontinuities from one level to another. In most cases there were no intermediates. (4) Estimates of the age of the earth by geologists and physicists, such as William Thompson, Lord Kelvin (1824–1907), claimed that the earth was too young for Darwin’s slow, gradual process to have produced all the diversity that currently existed. (5) Although incorrectly attributed to Darwin, the view that humans “came from monkeys” caused considerable controversy by giving humans an animal ancestry that was unacceptable from a secular as well as theological perspective. In the subsequent five editions of The Origin Darwin tried to answer some of these suggestions, but it was only well into the twentieth century and the so-called evolutionary synthesis of the 1930s–1950s that many of these original problems with Darwin’s paradigm were eventually resolved.
Darwin did have his supporters, however. In England, T. H. Huxley acted as his “bulldog,” championing his basic paradigm in both scientific and lay circles. In Germany, August Weismann (1834–1914) and morphologist Ernst Haeckel (1834–1919) both became ardent Darwinians. Weismann’s doctrine of the separation of the germ- and somatoplasm contradicted any notion of Lamarckian inheritance. Haeckel more closely followed Darwin in advocating the inheritance of acquired characters and in emphasizing the importance of comparative embryology as evidence for evolution. His “biogenetic law” (that ontogeny, or development of the individual, recapitulates phylogeny, or development of the evolutionary lineage) was based on the remarkable similarity of the early embryonic stages among related organisms, such as the vertebrates. Although he introduced Darwinism to a wide audience, Haeckel’s grandiose ideas gave evolutionary theory a reputation among many biologists for excessive speculation. In the United States, Harvard botanist Asa Gray (1810–1888) championed the basic idea of evolution against his colleague Louis Agassiz (1807–1873) while attempting to reconcile the apparent cruelty and wastefulness of natural selection with Christian theology.
Darwin, Evolution and Society: The Religious Reaction and “Social Darwinism”
The lay reaction to Darwin occurred on both religious and secular grounds. The main religious objection was that evolution as such contradicted the biblical story of creation. Also, the consequence of the view that humans were derived from lower animals was that they lost their place as the special and final creation by God. Darwin had effectively banished God from the everyday universe and, in so doing, had introduced a thoroughly materialistic view of the universe and life, one that lacked purpose (was nonteleological) and thus seemed to lack meaning (see chapters VIII.13 and VIII.14).
On secular grounds, the extension of Darwin’s natural history to explain the workings of society flourished in the late nineteenth century. One aspect, known as “social Darwinism” (by its critics), promoted by the English philosopher and sociologist Herbert Spencer (1820–1903), the American political economist William Graham Sumner (1840–1910), and the German Ernst Haeckel, argued that “survival of the fittest” (Spencer’s term) was a law for human society as well as nature and thus dictated a laissez-faire economic and social policy. Hard-nosed “Social Darwinists” spoke out against public charity or support for the poor on the grounds that it would encourage the unfit to propagate. Some applied Darwinian models to explain racial differences and the superiority of European races over those in Africa and South America (see chapter VIII.11). A very different extension was promoted by Prince Peter Kropotkin (1842–1921) in Russia, emphasizing cooperation (symbiosis, mutualism) rather than competition as the major characteristic of evolution. The problem with all such extensions of Darwin’s (or any other) scientific theory to society is that the theory can be stretched to support almost any a priori political or philosophical views. Supporters from the political left as well as the right claimed the authority of Darwin, partly by drawing on only certain aspects of evolutionary theory (competition and cooperation both exist in the natural world). And since Darwin’s metaphor of competition, overproduction, and selection were all processes he initially drew from the social world (i.e., from Malthus), extending these processes to explain society becomes tautological (i.e., circular reasoning).
7. POST-DARWINIAN CONTROVERSIES AND THE “ECLIPSE OF DARWINISM,” 1890–1920
Evolutionary debates continued unabated from Darwin’s day through the interwar period of the 1920s and 1930s. Of Darwin’s five theories, the most generally and quickly accepted (by the 1870s) was descent with modification. The most controversial was the theory of natural selection, which by 1900 most naturalists viewed with considerable skepticism. In addition to voicing earlier objections, some scientists now argued that while selection might occur, it was at best only a negative process: it weeded out the unfit but could not account for the origin of the fit. And of course, Darwin’s lack of consistent ideas about heredity was recognized as a major problem. Thus, if the core of Darwinism is evolution by natural selection, by 1900 Darwin’s paradigm was, according to one German commentator, “on its deathbed.”
In response to the perceived inadequacies of Darwinism, biologists introduced a variety of alternative processes, or mechanisms, by which evolution might occur, among the most important of which were neo-Lamarckism, orthogenesis, and mutationism.
Taking their lead from Darwin, a number of naturalists adopted a neo-Lamarckian view to explain the origin of variation from various environmental effects, as well as the use and disuse of parts. Among the strongest supporters were the American paleontologists Alpheus Hyatt (1838–1902), Edward Drinker Cope (1840–1897), Henry Fairfield Osborn (1857–1935), and the Austrian zoologist Paul Kammerer (1880–1926). Neo-Lamarckism solved the problems of adaptation, since variations arose in response to specific environmental demands. It also shortened the time required for evolution to occur, since it did not depend on chance variations. Even in light of Weismann’s experiments on mice (see previous discussion) many biologists maintained a neo-Lamarckian position well into the mid-twentieth century, especially in France and the Soviet Union. In the latter, neo-Lamarckism became the basis for agricultural policy after 1948 under the direction of agronomist Trofim D. Lysenko (1898–1976), who claimed that physiological traits such as flowering time could be altered by exposing seeds to varying conditions such as low temperature or artificial light cycles. These effects, Lysenko argued, could be transmitted to future generations.
Orthogenesis
Orthogenesis was the view that evolution inexorably proceeds along certain lines, or directions, and often leads to extinction. The two best-known examples were increase in size of the antlers of the now-extinct Irish elk, and the enormous canine teeth of saber-toothed tigers. A key feature of orthogenesis was the claim that once evolutionary trends were established, even though adaptive initially, they gained momentum and eventually progressed to nonadaptive extremes. Orthogenesis, often accompanied by neo-Lamarckism, was particularly popular among paleontologists because it seemed to explain the various trends in the fossil record such as increase in size or complexity of a trait prior to extinction of the lineage. Orthogenesis also revived a form of Aristotelian teleology in which large-scale trends suggested a direction and purpose in nature, an idea many naturalists were reluctant to give up completely.
Mutationism
Still another alternative to Darwinian evolution was mutationism, the claim that large-scale changes could lead to formation of a new species in one generation. Most effectively promoted by Dutch plant physiologist Hugo de Vries (1848–1934) in his two-volume Die Mutationstheorie (The mutation theory) in 1904, mutationism gained a significant following because it seemed to be empirically supported and, most of all, was “experimental,” meaning that evolution could be investigated in the laboratory. De Vries had observed the evening primrose (Oenothera lamarckiana) producing offspring that were not only strikingly different from their parents but also infertile. De Vries thought he had discovered a mechanism by which evolution occurred in short periods of time and could thus be studied experimentally under controlled conditions. As it turned out, Oenothera was an unusual plant with very atypical chromosome structures and meiotic processes that accounted for the unusual forms of the offspring. The theory was ultimately abandoned by the 1920s, when other examples failed to materialize and when laboratory genetics revealed a variety of mechanisms for the origin of variations on which selection could act (point mutations, chromosomal rearrangements, recombination).
Isolation and the Role of Geographic Barriers
One of the main problems Darwin had not fully resolved in The Origin was that of speciation itself: how one ancestral species diverged into two or more descendant species. While he recognized that geographic separation certainly played a role, he thought it was only one of many factors (self-fertilizing organisms might diverge without being geographically separated). In the decades after Darwin’s death this ambiguity led to a controversy between two models of speciation: what would later be called allopatric (populations diverge into new species only when separated by a barrier) versus sympatric (populations diverge in the same region, not physically isolated from one another). One of the first evolutionists to emphasize the necessity of isolation by means of geographic barriers for speciation to occur was the American missionary-naturalist John Thomas Gulick (1832–1923) in the 1870s and 1880s. Raised by missionary parents in Hawaii, Gulick collected land snails of the family Achatinellidae from adjacent valleys on Oahu. He noted that considerable differences in the shell color and banding patterns existed among specimens from different valleys, which suggested that the populations had diverged as a result of being isolated from one another by the steep valley walls. The same point was made by German naturalist Moritz Wagner (1813–1887) from studies of insects in the Caucasus and Andes Mountains and by American ichthyologist David Starr Jordan (1851–1931) in studies of fish populations in lakes separated by land barriers. The central argument in all these claims was that geographic barriers prevent interbreeding and thus allow each population to accumulate its own unique set of variations, and through natural selection, its adaptation to its particular environment. Isolation came to be regarded as necessary for speciation.
Those who argued for the possibility of some form of sympatric speciation pointed to de Vries’s mutation theory and other kinds of “sports” as examples of speciation without isolation. Later, when polyploidy (duplication of chromosomes without cell division) was discovered, it, too, was cited as a mechanism for speciation without the necessity for geographic barriers. However, the eventual discrediting of de Vries’s mutation theory as a general process and the recognition that polyploidy is restricted to a few species of plants led to increasing acceptance of the role of geographic isolation as a major and necessary component to most speciation in the natural world.
Evolution and the Evangelical Opposition
The somewhat-confused state of evolutionary theory in the period 1910–1930 was not helped by the revival of religious objections, especially in the United States. For a variety of reasons, including suspicion of science as an aspect of “modernism” (the erosion of traditional values due to industrialization and urbanization), the rise of populism, and the association of “social Darwinism” with German aggression in World War I (1914–1918), evolution came under attack by Evangelical Christians and was dramatically highlighted by the trial of teacher John T. Scopes (1900–1970) in Dayton, Tennessee, in 1925. A test case of a bill introduced into the Tennessee legislature earlier that year prohibiting the teaching of evolution in public schools, the trial pitted two highly public figures, as attorneys, against each other: Chicago lawyer Clarence Darrow (1857–1938), who defended Scopes, and two-time presidential candidate and former Secretary of State William Jennings Bryan (1860–1925), who prosecuted the state’s position. By the unorthodox strategy of cross-examining his opponent, Darrow was able to turn the trial into a strong plea for the primacy of science over religious ideology, and for rationality over superstition and folk belief. Although Scopes was found guilty and fined a nominal $100, the trial generated worldwide controversy and began a trend to attack evolution in the United States that has persisted, largely in religious circles, to the present (see chapters VIII.13 and VIII.14).
8. HEREDITY AND EVOLUTION: MENDELISM, DARWINISM, AND THE “EVOLUTIONARY SYNTHESIS”
The Problem of Heredity and the Rediscovery of Mendel (1900–1925)
Given Darwin’s lack of a hereditary mechanism, it would seem that the rediscovery in 1900 of Gregor Mendel’s experiments with hybridization in peas would have immediately resolved many of the problems confronting Darwinian theory (for example, blending inheritance or reversion). This was not the case, however. Many biologists were skeptical of the general applicability of Mendel’s work, and a group known as the “biometricians” in England (and their followers elsewhere) claimed that Mendel’s work had no bearing on evolution. Led by Darwin’s cousin Francis Galton (1822–1911) and his protégé Karl Pearson (1857–1936), biometricians stressed the importance of investigating matters of heredity and variation quantitatively while developing statistical procedures for analyzing data (they introduced methods such as correlation and regression). Committed to Darwin’s emphasis on the importance of continuous variations as the raw material for evolution, they rejected Mendelism as a theory of discontinuous variation, allied in many people’s minds with de Vries’s mutationism. A major battle emerged in England between the biometricians, under Pearson, and the Mendelians, championed by William Bateson (1861–1926), one of the most forceful early promoters of Mendel’s work. This controversy, which raged between 1901 and 1908, left the distinct impression that Mendelism, whatever its value might be for plant or animal breeders, had little to contribute to understanding the evolutionary process. Although some Mendelians came to see that small, but discrete, Mendelian variations could serve as the raw material on which selection could act, it took almost two decades for a new generation of investigators, more thoroughly trained in mathematics and statistics than their predecessors, and freed from the earlier hostility to Mendelism, to see the ways in which Mendel’s theory could be directly applied to Darwinism. Beginning in the late 1920s and early 1930s, and continuing through the 1940s and 1950s, this period has been labeled the “evolutionary synthesis.” It was one of the most important developments in twentieth-century evolutionary biology.
The “Evolutionary Synthesis,” 1930–1940
The period of the evolutionary synthesis brought together a number of divergent views and fashioned a comprehensive understanding of the evolutionary process that included Mendelian and quantitative genetics, biometry, classical Darwinian selection theory, taxonomy, biogeography, and paleontology. It was also noted for its role in excluding from evolutionary theory once and for all the older alternative views of neo-Lamarckism, orthogenesis, saltation (macro)mutation, and sympatric speciation (embryology, so central to Darwin’s views, was notably absent, leading to its neglect by evolutionary biologists for most of the remainder of the century). By the 1930s, geology and paleontology were beginning to provide a considerably longer time frame for the history of the earth, thus supporting the gradualist views so central to orthodox Darwinism. Incorporating Mendelian genetics into evolutionary theory made it possible to demonstrate that inheritance was not blending but discrete and thus that new variations, even if they were slight and recessive, were fully recoverable in future generations. Moreover, because they were discrete, Mendelian genes could be treated as mathematical entities whose change in frequency from one generation to another provided a quantitative measure of evolution. Combined with Darwin’s inherent population thinking, the immediate effect of the synthesis was the emergence of the new field of mathematical population genetics.
For simplicity, the synthesis may conveniently be seen as occurring in three phases. The first (roughly 1918–1940) was dominated by the theoretical population genetics of Ronald A. Fisher (1890–1962), Sewall Wright (1889–1988), and John Burdon Sanderson Haldane (1892–1964) and signified the combining of Mendelism with Darwinian selection theory. This first phase involved the application of the then rather novel methods of mathematical model building to populations of organisms, in which various parameters such as population size, random versus selective mating, alterations in initial gene frequencies, relative fitness, and the effects of selection could be manipulated within the model, and the various outcomes predicted. It was through developments in this phase that the conflict between Mendelians and biometricians was finally resolved. The second phase (roughly 1940–1970) involved the unification of mathematical population genetics with other aspects of evolutionary theory, including paleontology, taxonomy (including the ongoing debate about species definitions), biogeography, field studies of natural variation, population structure, and early studies on the origin of life (abiogenesis). The third phase (roughly 1970–1990s) involved expansion of topics such as extinction (including mass extinctions), evolutionary ecology, experimental evolution (especially using microbial systems), and molecular evolution. All three phases drew on the more general introduction into biology of statistical thinking that had emerged in the physical sciences in the later years of the nineteenth century.
The First Phase: Mathematical Population Genetics
The seminal contribution in the first phase of the synthesis was made by the British mathematician Ronald A. Fisher, who sought to promote a synthesis between orthodox Darwinism and Mendelism and to apply these principles to both agricultural breeding and eugenics, or the genetic improvement of the human species. All these interests were brought together in his major book, The Genetical Theory of Natural Selection (1930), which aimed to demonstrate the efficacy of natural and artificial selection acting on discrete Mendelian genes. Fisher showed theoretically that, among other things, small selective forces applied over a sufficiently long period could lead to significant genetic change within a population. These changes could be measured quantitatively as shifts in the frequency of certain genes over successive generations: the new, mathematical definition of evolution. Particularly important, Fisher’s “fundamental theorem,” put forward in the Genetical Theory, claimed that the effectiveness of selection was directly related to the total amount of variation in a population: natural or artificial selection could produce change only if there was sufficient variation on which to act. This became an important principle for both the practical breeder and the student of evolutionary mechanisms, as it resolved the old problem of whether selection was a “creative” or only a “negative” force. It was neither: selection required a source of genetic variation to produce inherited change within a population, and both weeded out the unfit and preserved the fit.
Fisher’s model was based on populations that were large, panmictic (freely interbreeding) groups akin to the physicist’s gases in a finite chamber. Like gas molecules, individuals within such a population interacted (mated) randomly, and all combinations were equally possible. A population was defined by its gene frequencies, by what came to be called its gene pool. Fisher aimed to treat evolution as lawlike, subject to mathematical formulation like the kinetic theory of gases, according to which organized and predictable outcomes could result from myriads of random events. Despite this oversimplification, there is no doubt that Fisher’s approach showed that evolutionary processes could be represented mathematically and that model building was a useful approach to understanding how selection, dominance, recessiveness, and other factors could quantitatively alter the course of evolution in a population.
A mathematical contribution of a different sort was made by American biologist Sewall Wright. A decade of work at the United States Department of Agriculture (USDA) between 1916 and 1925 brought Wright face to face with issues of breeding and the agricultural problems of how to increase the effectiveness of selection at the population level. Wright concluded that the most effective approach was to subdivide a large population (a herd of cattle, for example) into small breeding groups. These would be inbred for several generations and then outcrossed periodically with other subgroups. Inbreeding within the small groups would help fix desirable genotypes, while outbreeding among groups would bring new combinations together that would provide additional raw material for selection. This early work led Wright to recognize that the breeding structure of a population was critical to understanding how evolutionary change most likely occurs in nature. He came to oppose the model of a large panmictic population, promulgated by Fisher, as having little reality in nature. In contrast, Wright claimed that large populations in nature were usually subdivided into smaller subpopulations, which he called demes, each occupying its own ecological space (“microniche”) within the population’s overall geographic range. Two aspects of Wright’s model became important innovations in the evolutionary synthesis. One was the shifting balance theory of evolution, and the other, as a consequence, was genetic drift.
Employing the metaphor of a topographical map, Wright’s shifting balance theory depicted each deme on its own “adaptive peak,” separated from other demes by valleys of ecologically less adaptive “terrain.” Each deme would thus have its own characteristic distribution of gene frequencies, although through migration, genes could be exchanged between demes within the population at large. The adaptive landscape was not static, however, since climatic or other external conditions, as well as the activities of the organisms themselves, gradually “eroded” the peaks, altering the overall topography. As conditions changed, demes would be forced to move off their adaptive peaks, either “migrating” to another peak where their particular combination of genes might be more adaptive or, failing to find such a peak, becoming extinct. Demes were thus constantly being challenged to undergo a shift in the balance of their gene frequencies owing to the constant interaction of the small subpopulations with their microniches (hence the name “shifting balance theory”).
Associated with the shifting balance theory was what Wright referred to as genetic drift, the process by which, in small populations, gene frequencies could become fixed, either at 100 percent or 0 percent, simply by the vagaries of random processes such as chance matings, differential survival, and reproductive success. Genetic drift became one of Wright’s seminal contributions to evolutionary thinking. However, genetic drift was misunderstood by many of Wright’s contemporaries, who thought that he was suggesting natural selection had little or no role in evolution and that most change was due to chance. Nevertheless, Wright always maintained that selection also played a major part in long-range evolutionary processes; his shifting balance theory emphasized only the role of random effects due to small population size.
A third major contribution to mathematical population genetics came from John Burdon Sanderson Haldane (1892–1964), the son of the well-known Oxford physiologist John Scott Haldane (1860–1936). Because of his strong biochemical as well as mathematical background, Haldane emphasized the ways in which genetic variations might act physiologically, for example, in slowing down or speeding up metabolic reactions, and thus provide an understanding of how selection might actually affect gene function. Such changes could also be computed mathematically. More than Fisher or Wright, Haldane also emphasized the importance of gene interactions as targets of selection, and brought embryological considerations into the evolutionary process by pointing out how different selection pressures must come into play at each stage in an organism’s life cycle.
The Second Phase: The Genetics of Natural Population
One of the earliest applications of Mendelian genetics to the study of natural populations appeared in the work of Sergei Chetverikov (1880–1959), a Russian butterfly taxonomist who was well acquainted with the problem of variation in natural populations. In 1922, at the Institute of Experimental Biology in Moscow, he was introduced by its director, Nikolai Koltsov (1872–1940), to the laboratory cultures of Drosophila melanogaster that H. J. Muller (1890–1967) had just brought from the US laboratory of Thomas Hunt Morgan (1866–1945). Because Drosophila was the premier experimental organism for studying genetics, the cultures brought by Muller were highly useful, since they contained point mutations at known positions on the chromosomes. Chetverikov was able to crossbreed these known stocks with flies from natural populations, to uncover variations that were masked in the wild by their dominant alleles. Chetverikov’s experiments suggested for the first time that there was considerably more variation in natural populations than had previously been thought. This was important because the rate of evolution, as predicted by Fisher and others on theoretical grounds, depended on the presence of a large amount of variation. Thus, determining how much variation was actually present became an important empirical investigation during the period of the evolutionary synthesis. The Russian school, under Chetverikov and others, contributed significantly to this effort. Unfortunately, owing to World War I (1914–1918), the upheaval of the Bolshevik Revolution (1917), and the fact that most Western biologists did not read Russian, the work of Chetverikov and his colleagues was not known outside the USSR until the 1940s, when it was brought to light and translated by Theodosius Dobzhansky (1900–1975).
A major figure in the second phase of the synthesis, Theodosius Dobzhansky brought the Russian perspective combining field and laboratory investigations to bear on determining the amount of variation in natural populations and the effect of natural selection on those variations. Dobzhansky’s 1937 book, Genetics and the Origin of Species, had already outlined the close relationship between genetics, cytogenetics (the microscopic study of chromosome structure and configuration of known genetic strains), and evolution, and became one of the major works outlining the new synthetic approach to evolutionary theory. In a 1947 paper, Dobzhansky reported on a series of innovative field studies on variation and selection, in which he introduced two quite novel approaches. First, he sought to inspect the genotype directly, by observing cytologically the frequency of chromosome variants (inversions) in natural populations, using the methods of cytogenetics he had learned from working in the Morgan laboratory at Columbia and Caltech. Second, he carried the work a step further by developing an experimentally based laboratory approach that allowed him to test under controlled conditions hypotheses about the fitness of different chromosomal variants found in natural populations.
Surveying populations of Drosophila pseudoobscura taken at regular geographic intervals across the Southwest from California to Texas, Dobzhansky found that different populations could be characterized by different frequencies of certain chromosomal inversion patterns. These patterns were detected microscopically but had no visible phenotypic effect by which they could be recognized or spotted. More interesting, Dobzhansky found that the frequency of four such arrangements he studied changed within any one population in a regular, cyclic pattern over the course of the year. Dobzhansky hypothesized that these changes were due to changes in selection pressures during different seasons. From an evolutionary point of view, it was clear that selection favored maintaining a range of variations (called balanced polymorphism) within a population subjected to regular changing conditions.
Dobzhansky then proceeded to study this phenomenon under controlled conditions in the laboratory. Until the 1940s it had been thought that evolution by selection was too slow a process to be observed by human beings in their lifetime—that is, it was a process whose effects could be seen only in the span of geologic time. Dobzhansky’s suggestion that seasonal fluctuation in certain genotypes was produced by changing selection pressures almost seemed preposterous. He pointed out, however, that different chromosomal arrangements very likely had differential adaptive capacities, one type being more adapted to, say, a particular temperature or humidity level. In the lab Dobzhansky exposed flies with different chromosomal inversions to varying conditions and observed that he could alter the frequency of one inversion type over another simply by changing the environment (temperature was the most effective selective agent). He was witnessing evolution right before his eyes.
Dobzhansky’s experiments emphasized two important points. The first was that, at last, even the theory of evolution by natural selection could be subject to experimental and quantitative tests. To Dobzhansky and many other biologists at the time it seemed that a problem that had been refractory to experimental study for over a century had at last yielded to rigorous analysis. The second point that Dobzhansky emphasized was that what appeared to be adaptive were not individual genes or their phenotypic traits but, instead, the whole complex of genetic traits carried on a chromosome, a part of a chromosome, or within the entire complex of chromosomes in the population. Selection operates at multiple levels.
The Third Phase: Taxonomy, Paleontology, and Embryology in Relation to Evolution
At the end of The Origin Darwin had prophesied that accepting his paradigm of evolution by natural selection would “revolutionize” a variety of fields of natural history, including taxonomy. As far as the latter was concerned, it did not turn out that way at the outset, but by the 1930s and 1940s taxonomists like the German émigré to the United States Ernst Mayr had begun to apply aspects of Darwin’s population approach to taxonomic questions, particularly the nature of species and species definitions. Mayr argued in his 1942 book, Systematics and the Origin of Species, that the focus of taxonomy should be on the range of variation in populations, not on characterizing the most “typical” member of the group. This was a shift, Mayr noted, from the older “typological” or “essentialist” view of species to a modern, dynamic, or population view. A species was now defined by the variability it displayed, as this was the creative reservoir on which natural selection acted. Taxonomists, he argued, should define their species as active populations in the process of evolving, not simply as the static products of evolution. Mayr called this the “biological species concept” and defined it primarily in terms of reproductive compatibility (see chapter VI.1). This became a highly debated point over the ensuing years, since many biologists, botanists, and microbiologists, for example, felt Mayr’s definition was too narrow to encompass the taxa with which they worked (see chapter VI.1).
In the nineteenth century paleontology provided the only hard evidence that evolution had actually occurred, but it was always problematic (see chapter II.9). First, it did not provide the intermediate forms that Darwin’s gradualist paradigm required. Also, since fossils are nonliving and are often fragmentary, their life histories are difficult to reconstruct and their taxonomy difficult (many fossils cannot be keyed to the species level with any certainty). Moreover, paleontology was tied professionally to geology, as fossils are often the key to stratigraphy, that is, to correlating strata from one region to another. By the 1930s and 1940s, however, several paleontologists, principally George Gaylord Simpson (1902–1984) and Norman D. Newell (1909–2005), both at the American Museum of Natural History in New York, argued for a closer alliance of paleontology with biology. Simpson had been strongly influenced by Dobzhansky’s Genetics and the Origin of Species and sought to bring the perspective of genetics, population thinking, and statistics to bear on paleontology. Both Simpson and Newell claimed that paleontologists should think of fossils as “once-living organisms” that existed in populations. In Tempo and Mode in Evolution (1944) Simpson argued that paleontology could complement population genetics by revealing large-scale trends and developments over time (tempo, or rates of change, for example) that study of living populations could not. Paleontology could thus contribute to understanding macroevolution, and population genetics could focus on microevolution. Simpson and others were responsible for transforming paleontology into paleobiology and bringing the field into the evolutionary synthesis.
Simpson’s challenge was taken up by a younger generation of paleontologists such as Stephen Jay Gould (1941–2002), Thomas J. Schopf (1939–1984), and Niles Eldredge (b. 1943), among others. Gould and Eldredge, for example, published a highly influential paper in 1972, “Punctuated Equilibria: An Alternative to Phyletic Gradualism,” that challenged the orthodoxy of slow, gradual change over evolutionary time. Their view of “punctuated equilibrium” characterized macroevolution as a series of rapid transformations followed by long periods of very slow change or stasis (such as the relatively small fossil sample from the pre-Cambrian compared with the Cambrian “explosion,” where most of the major body plans of the animal kingdom were laid down). For Gould and Eldredge this sharp change from one series of strata to another was not the result of an imperfect geological record but an actual picture of how evolution works. Punctuated equilibrium was a challenge to the orthodoxy of Darwinian gradualism (figure 4).

Figure 4. Comparison of evolutionary change according to the theory of phyletic gradualism (top) and the punctuated equilibrium of Gould and Eldredge (bottom). Punctuated equilibrium emphasizes sharp breaks when evolutionary change is rapid, followed by periods of slow change (vertical lines).
Gould, in particular, was also influential in bringing the perspectives of developmental biology back into evolutionary theory after its long hiatus. Gould emphasized that evolutionary lineages were always under developmental constraints that limited the paths they could follow. That is, genetic variations that seriously altered developmental sequences would be selected against in favor of those that worked within the limits of existing embryonic processes. Development, and the constraints it imposed, represented an intermediate set of phenotypes between the genotype of the fertilized egg and the adult phenotype. Gould’s emphasis on the importance of development led to the current interest in “evo-devo” (evolutionary-developmental biology; see chapters V.11 and V.12). In recent years, evo-devo has been highly informed by new knowledge in molecular genetics, methods of gene signaling, and genetic control mechanisms (see chapters V.1–V.3 and V.6–V.8).
9. EVOLUTIONARY THEORY IN THE ERA OF MOLECULAR BIOLOGY
With the rapid development of molecular biology after the discovery of the molecular structure of deoxyribonucleic acid (DNA) in 1953, evolutionary biology made great strides in several areas: more precise determination of evolutionary rates, the molecular basis of evolution, molecular taxonomy, comparative genomics of related lineages, and mechanisms of variation in both structural genes (those that code for functional proteins) and control elements (genes and gene products that control the expression of other genes).
Once the structure of DNA was clear, the molecular basis for variation (mutation) could be understood as base-pair substitutions and their effects on the subsequent proteins for which the genes code. With techniques such as chromatography and later electrophoresis (both means of separating molecules with only slightly different structures), it became possible by the 1950s and 1960s to compare molecules (proteins such as the hemoglobins and later DNA itself) from different organisms and work out “molecular phylogenies,” that is, the historical lineages of the molecules themselves (see chapter V.1). These often, but not always, confirmed existing lineages based on more conventional morphology. Further, since the rate of base-pair substitution could be determined experimentally, differences in similar proteins or the DNA that produced them yielded a more precise measure of divergence rates and times (what was called the molecular clock; see chapter V.1). Having a calibrated clock aided, for example, in determining more accurately the time of divergence between humans and our pongid ancestry, or between current human geographic populations.
A consequence of accumulating data from molecular genetics was the recognition that many mutations at the DNA level are the result of random drift rather than selective pressures. Such observations led the Japanese geneticist Motoo Kimura (1924–1994) in a 1968 article to propose the neutral theory of molecular evolution, which argued that most mutations in DNA are selectively neutral and remain in the genome because they are not eliminated by natural selection (see chapter V.1). Kimura’s idea was given broader coverage by Jack L. King and Thomas H. Jukes in another article, “Non-Darwinian Evolution,” in 1969. Kimura emphasized that the neutral theory did not dispense with natural selection as having a key role in the evolution of adaptations at the phenotypic level; but he did point out that the theory aimed to counter the prevailing idea that most mutations were of positive or negative selective value, and thus their frequency in a population was exclusively the result of selection. In many ways Kimura’s theory was an extension of Wright’s concept of genetic drift. However, it became highly controversial because it seemed to contradict the central role given to selection throughout the evolutionary synthesis. Although the controversy had died down to some degree by the 1990s, it is still often invoked as a null hypothesis in testing the role of variations in evolution. It remains an issue of debate, however, especially in discussions of the relative importance of the role of drift with respect to population size. Especially in taxonomy, comparisons of selected DNA segments have helped delineate monophyletic taxa with a precision not possible with traditional comparative methods. Particularly important has been the elucidation of the many mechanisms by which gene expression is controlled, from transcription of messenger RNA (mRNA), to alternative splicing (varying ways in which the same mRNA transcript can be cut and modified), to posttranslational modification of proteins (including how proteins are folded into their tertiary structure). With the advent of rapid, automated sequencing equipment, the complete genomes of a number of organisms have now been elucidated (mouse, fruit fly, nematodes, yeast, humans, to name a few). Comparative analysis using computers has revealed a remarkable number of conserved sequences of DNA, such as the “homeobox genes” that serve similar or many times quite different functions in widely divergent organisms (the “homeobox,” a 180-nucleotide sequence, is found in both Drosophila and humans, where it functions in laying down basic patterns of segmentation in the anterior-posterior axis). Even more striking, similarly related sequences have been found in regulatory genes, indicating that evolution can act on existing genetic systems by slight modifications of control elements that can have large effects at the genotypic level (see chapters V.2, V.3, and V.6–V.8). All these discoveries have suggested that evolution may proceed by modular modification of a number of existing genetic elements, just as a contractor can produce a wide variety of houses by arranging and rearranging a few basic room designs. Far fewer genes appear to be needed than previously believed to produce infinite variation for evolutionary change. New mechanisms of variation, their effects on development, and their evolution through time now form the forefront of twenty-first-century evolutionary theory (see chapters V.13 and V.14).
Bowler, P. J. 2009. Evolution: The History of an Idea. Berkeley, CA: University of California Press. The best single treatment of the evolution idea from the seventeenth century through the present day. Well written and clear, this is an often-used textbook in courses on the Darwinian Revolution.
Browne, J. 1995. Charles Darwin. Vol. 1, Voyaging. London: Jonathan Cape, 2002. Vol. 2, The Power of Place: The Origin and After—the Years of Fame. New York: Alfred A. Knopf. A fascinating and scrupulously detailed biographical study of Darwin, his influences, and the development of his work.
Browne, J. 2006. Darwin’s Origin of Species: A Biography. A much-shortened excerpt from the two-volume biography that focuses on the factors and events leading up to the publication of The Origin, its reception, and legacy. Beautifully written and an easy, quick introduction to Darwin’s major work.
Darwin, C. 1859. On the Origin of Species by Means of Natural Selection, or the Preservation of Favoured Races in the Struggle for Existence. Facsimile of the first edition, with an introduction by Ernst Mayr. Cambridge, MA: Harvard University Press. Harvard Press has also issued, more recently, The Annotated Origin, with an “Introduction” and explanatory notes by James T. Costa (2009). Both books are facsimile reproductions of the original edition, but the latter has useful explanatory notes on a variety of issues, including updated discussions of Darwin’s observations (for example, on ecology), and historical notes identifying people, places, practices.
Gould, S. J. 2002. The Structure of Evolutionary Theory. Cambridge, MA: Harvard University Press. This massive publication is both historical and current, in that it covers most of the history of evolutionary theory in the pre-1950 period but also discusses late twentieth-century developments in both micro- and macroevolution.
Kohn, D., ed. 1985. The Darwinian Heritage. Princeton, NJ: Princeton University Press. A collection of essays by major historians and philosophers of science, discussing everything from the structure of Darwin’s argument to his influence by early nineteenth-century romanticism.
Mayr, E. 1982. The Growth of Biological Thought: Diversity, Evolution and Inheritance. Cambridge, MA: Harvard University Press. Not really the growth of biological thought but primarily of evolutionary biological thought, Mayr’s tome covers 3000 years of natural history and evolutionary thinking. A great reference and compendium, though, like all the author’s work, written from his distinct perspective as a proponent of neo-Darwinism and an architect of the “evolutionary synthesis” of the 1930s–1960s.
Mayr, E. 1985. Darwin’s five theories of evolution. Chap. 25 in D. Kohn, ed., The Darwinian Heritage. Princeton, NJ: Princeton University Press. Lays out the five components of Darwin’s theory or paradigm: Evolution as Such, Common Descent, Gradualism, Multiplication of Species, and Natural Selection.
Provine, W. 1971. The Origins of Theoretical Population Genetics. Chicago, IL: University of Chicago Press. Despite its age, this remains one of the most concise and readable accounts of the first phase (mathematical population genetics) of the evolutionary synthesis.
Ruse, M. 1997. From Monad to Man: The Concept of Progress in Evolutionary Biology. Cambridge, MA: Harvard University Press. An interesting sweep of the history of evolutionary thinking, both pre- and post-Darwin, and the central role that notions of progress have played to the present day (includes modern evolutionary biologists such as E. O. Wilson, Stephen J. Gould, Richard Lewontin, Richard Dawkins, and John Maynard-Smith).